Introduction
Intracranial aneurysm (IA), also termed cerebral
aneurysm, is a localized abnormal widening of a cerebral artery
lumen or arterial wall due to increased intracranial pressure
(1). The estimated prevalence of
IA is 2.3% worldwide (2). Usually,
IAs can be divided in two subtypes: Unruptured IAs (UIA) and
ruptured IAs (RIA). An estimated 50–80% of IAs will not rupture
during a person's lifetime (3). As
aneurysmal rupture is the main event leading to the occurrence of
subarachnoid hemorrhage (4) and it
often results in high disability or death rates (5), the management of UIAs for preventing
RIAs is crucial (6). Genetic
factors have a crucial role in regulating RIA pathogenesis
(7); therefore, it is important to
identify the genetic risk factors and elucidate the molecular
mechanisms involved in IA.
Many genes, microRNAs (miRNAs) and transcription
factors (TFs) have been associated with the progression of IA.
Previous studies have revealed that the downregulation of elastin
and lim domain kinase 1 may increase the risk of IA by weakening
the altered arterial wall (8). In
addition, the expression levels of colony stimulating factor 3 and
AXL receptor tyrosine kinase are increased in patients with RIA
(9). The upregulated ATP binding
cassette subfamily A member 1, which is associated with
intracellular lipid accumulation, may lead to IA by increasing the
occurrence of inflammation in the IA wall (10). Additionally, the dysregulation of
miRNA-9 is involved in IA by suppressing the proliferation and
reducing the contractility of smooth muscle cells (11) and the upregulation of miRNA-92a,
which negatively regulates the expression of kruppel-like factor 2
in a rabbit model, is also responsible for IA progression (12). The ETS proto-oncogene 1 TF promotes
the development of IA via mediating the overexpression of monocyte
chemoattractant protein-1 in IA walls (13). However, current research only
partially explains the pathogenetic mechanisms of IA and further
studies aimed at identifying IA-associated genes, miRNAs and TFs
and elucidating their involvement in molecular mechanisms are
required.
Nakaoka et al (14) have generated a gene expression
profile of patients with IA and identified the differentially
expressed genes (DEGs), revealing that the majority of DEGs are
associated with inflammatory and immune responses. However, they
did not use bioinformatics tools for an in-depth analysis of the
miRNAs and TFs associated with IA. Therefore, the present study
re-analyzed their expression data using a series of bioinformatics
methods to identify crucial IA-associated genes, miRNAs and TFs
that will allow the identification of the underlying mechanisms
associated with IA.
Materials and methods
Affymetrix microarray data
The microarray dataset GSE54083, downloaded from the
Gene Expression Omnibus (www.ncbi.nlm.nih.gov/geo), was analyzed using the
GPL4133: Agilent-014850 Whole Human Genome Microarray 4×44K G4112F
(Agilent Technologies, Inc., Santa Clara, CA, USA) platform. A
total of 18 samples were present in this dataset, including 8
ruptured IA (RIA) and 10 superficial temporal artery (STA) samples.
All 8 RIA samples were from females (age, 28–88 years) and the 10
STA samples included 8 females and 2 males (age, 34–61 years). All
specimens were obtained through surgery at the Tokyo Metropolitan
Fuchu Hospital (Tokyo, Japan), and all patients enrolled in the
experiment provided written informed consent approved by the Ethics
Committee of Tokyo Metropolitan Fuchu Hospital, Tokyo Women's
University (Tokyo, Japan) (14).
Data preprocessing
Raw probe-level data was downloaded, and expression
profile data preprocessing was performed using a robust multiarray
average method based on the affy package (version 1.52.0;
www.bioconductor.org/packages/release/bioc/html/affy.html)
in R version 3.3.2 (15). Data
preprocessing included background correction, filling in of missing
data and quartile normalization.
Identification of DEGs and
hierarchical clustering analysis
Following data preprocessing, DEGs between RIA and
STA samples were analyzed using Bayes methods based using the limma
package version 3.30.3 (www.bioconductor.org/packages/release/bioc/html/limma.html)
in R (16), and raw P-values were
revised using the Benjamini and Hochberg method (17). The cut-off criteria for defining
DEGs were adjusted P<0.05 and |log2 fold change (FC)|
>2. Clustering analysis was then conducted using the pheatmap
package in R (cran.r-project.org/web/packages/pheatmap/index.html).
Functional and pathway enrichment
analyses of DEGs
The Database for Annotation, Visualization and
Integrated Discovery version 6.8 (DAVID; david-d.ncifcrf.gov) provides functional
classification and annotation analyses of associated genes
(18). In the present study, the
DAVID database was used to perform Gene Ontology (GO) and Kyoto
Encyclopedia of Genes and Genomes (KEGG) enrichment analyses of
upregulated and downregulated DEGs. The significant GO terms and
KEGG pathways enriched with upregulated and downregulated DEGs were
selected with a criterion of P<0.05.
Protein-protein interaction (PPI)
network construction
The Search Tool for the Retrieval of Interacting
Genes version 9.1 (STRING; www.string-db.org) database can annotate all
functional interactions among proteins by computational prediction
methods (19), and the present
study used STRING to predict the interactions among proteins
encoded by DEGs. The PPI pairs with a combined score >0.4 were
identified, and the interaction pairs were visualized by
constructing a PPI network using Cytoscape version 3.0.
Subsequently CytoNCA version 2.1.6 (apps.cytoscape.org/apps/cytonca), a Cytoscape plugin,
was used for centrality analysis by examining the network topology
(20), and the hub nodes were
identified by a high score based on the scale-free property of the
network.
Integrated network construction of
TF-target-miRNA
Prior to integrated network construction, the TF and
miRNA targets were identified. The miR2Disease database (www.mir2disease.org) was used to search for
IA-associated miRNAs (21) and the
miRNA target genes were predicted from MiRwalk2, MiRDB, RNA22,
miRanda, RNAhybrid and TargetScan databases. A gene was categorized
as a miRNA target when identified in at least 4 out of the
aforementioned 6 databases. Genes shared between predicted miRNA
targets and DEGs were selected for analyses to predict whether TFs
were among DEGs and miRNA targets, which were performed based on
ITFP (itfp.biosino.org/itfp) and TRANSFAC
(www.gene-regulation.com/pub/databases.html)
databases. Subsequently, TF targets that overlapped between the
predicted TF target and TF DEGs were screened. Finally, the
regulation relationships between the TF-target and miRNA-target
were integrated, and the TF-target-miRNA network was constructed
using Cytoscape (www.cytoscape.org).
Results
DEG screening and hierarchy cluster
analysis
Based on the aforementioned criteria, a total of 777
DEGs were identified when the RIA samples were compared with the
STA samples, including 402 that were upregulated and 375 that were
downregulated. In addition, the hierarchy cluster analysis
indicated that DEGs could correctly distinguish between RIA and STA
samples based on significantly different expression patterns,
confirming that the results were reliable and could be applied to
subsequent analyses (Fig. 1).
Functional and pathway enrichment
analyses of DEGs
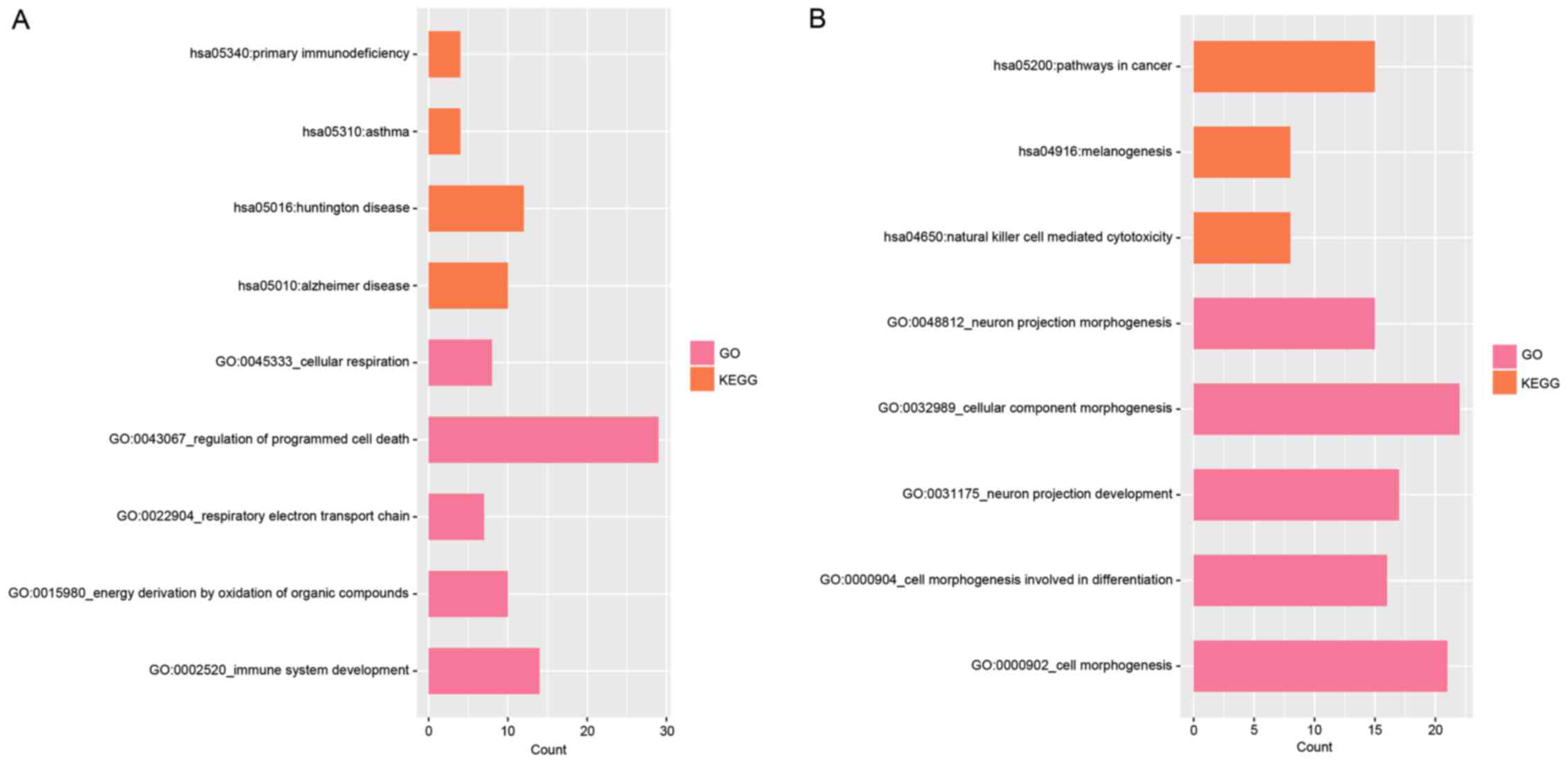
With a threshold of P<0.05, the upregulated DEGs
were enriched in 37 GO terms and 4 KEGG pathways. The top 5 GO
terms and 4 KEGG pathways are presented in Fig. 2A and mainly involved immune system
development (GO terms, P=0.006), regulation of programmed cell
death (GO terms, P=0.007) and primary immunodeficiency (KEGG,
P=0.047) categories. The downregulated DEGs were closely associated
with 135 GO terms and 3 KEGG pathways, which included natural
killer cell-mediated cytotoxicity (KEGG, P=0.029) and neuron
projection development (GO terms, P=1.828×10−5)
(Fig. 2B).
PPI network analysis
PPIs encoded by DEGs were identified using the
STRING database and a subset of 239 upregulated DEGs with 440
interactions was included in the upregulated PPI network (Fig. 3). For the downregulated PPI
network, there were 202 DEGs and 374 interactions (Fig. 4). In addition, the top 10
upregulated nodes, including CD40 and CD40LG and
downregulated nodes, such as DRD2 and TGFB1 with a
high degree score are presented in Table I.
 | Table I.Top 10 upregulated and downregulated
nodes of the protein-protein interaction network centrality
analysis. |
Table I.
Top 10 upregulated and downregulated
nodes of the protein-protein interaction network centrality
analysis.
| A, Upregulated |
|---|
|
|---|
| Gene | Degree | Betweenness | Closeness |
|---|
| KIT | 20.0 | 4938.772 | 0.07957205 |
| MAPK14 | 17.0 | 9604.479 | 0.08109029 |
| CD40 | 17.0 | 6098.7065 | 0.08153477 |
| CD79A | 14.0 | 4231.3047 | 0.0800269 |
| CD36 | 13.0 | 4260.1753 | 0.07986577 |
| CHD4 | 12.0 | 5844.3154 | 0.07989258 |
| POLR2K | 11.0 | 3625.8403 | 0.0771725 |
| SF3A2 | 11.0 | 1057.66 | 0.07345679 |
| SF3B3 | 11.0 | 1219.1407 | 0.073615834 |
| CD40LG | 10.0 | 4938.772 | 0.07957205 |
|
| B,
Downregulated |
|
| Gene | Degree | Betweenness | Closeness |
|
| TGFB1 | 16.0 | 3138.2646 | 0.03716029 |
| EDN1 | 14.0 | 3436.2148 | 0.037284363 |
| ADCY2 | 14.0 | 3696.108 | 0.037353653 |
| SAA1 | 14.0 | 2775.5098 | 0.036921382 |
| UBL4A | 14.0 | 4874.1514 | 0.03745807 |
| CCT5 | 13.0 | 2006.9825 | 0.03694853 |
| PRKCA | 13.0 | 5347.6064 | 0.037486013 |
| DRD2 | 13.0 | 1943.1166 | 0.036914602 |
| HSPD1 | 13.0 | 2716.8848 | 0.037249815 |
| RPS20 | 12.0 | 1213.8475 | 0.03707119 |
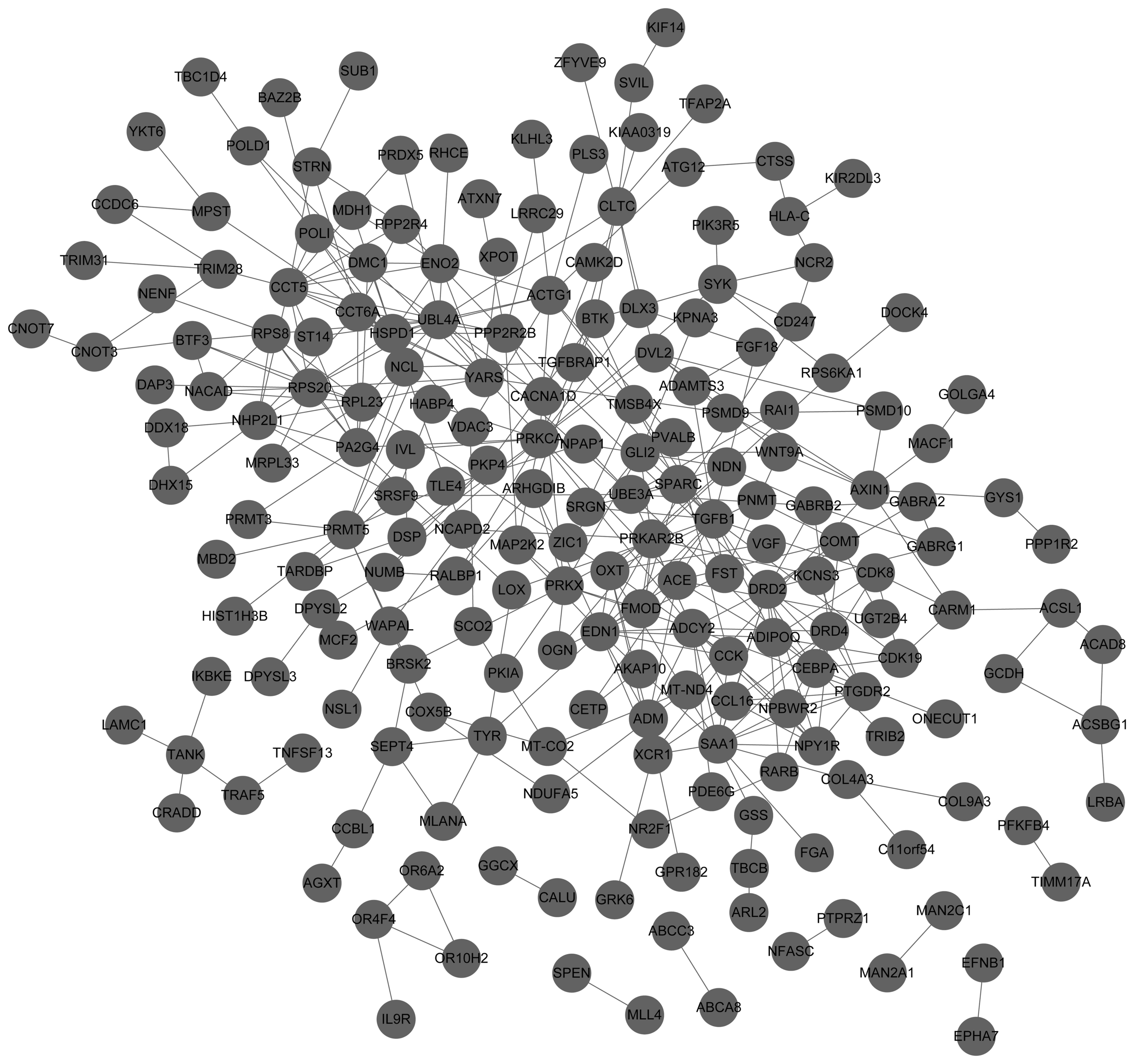
Integrated network analysis
Based on the miR2Disease database, 12 IA-associated
miRNAs were identified, and among these miRNAs, 9, including
hsa-miR-125a, hsa-miR-125b, hsa-miR-145, hsa-miR-146a, hsa-miR-21
and hsa-miR-214 were confirmed to be regulators of the
aforementioned DEGs. Additionally, a total of 18 upregulated DEGs,
such as MCM4, MRPS12 and SF3B2 and 15
downregulated DEGs, including TRIM28 that had a regulatory
function were categorized as TFs (Table II). Additionally, several DEGs
with a high degree score, such as STRN and NOS1 are
presented in Fig. 5.
| Table II.Upregulated genes and the
downregulated genes that could also be taken as transcription
factors. |
Table II.
Upregulated genes and the
downregulated genes that could also be taken as transcription
factors.
| Upregulated | Downregulated |
|---|
| ARHGAP25, CCNE1,
CIAO1, CIRBP, STF, CSTF2, GRSF1, GTF3C3, HOMER3, KLC1, MCM4,
MRPS12, POLL, SF3A2, SF3B2, SF3B3, SSBP1, TCF19 | ATXN7, BAZ2B,
DDX18, EML4I, GF2BP3, KLHL3, NCAPD2, NCL, PA2G4, POLD1, PRH1,
PRMT3, PSMD10, TANK, TRIM28 |
Discussion
In the present study, 402 upregulated and 375
downregulated DEGs, were identified in the RIA samples. The
majority of the DEGs were predicted to be associated with immune
system development (e.g., CD40LG and CD40) and neuron
projection development (e.g., STRN) functions and in terms
of pathways, such as primary immunodeficiency (e.g., CD40LG)
and natural killer cell-mediated cytotoxicity (e.g., MCM4).
In addition, CD40 (adjusted P=0.00001) and NOS1 were
hub genes in the constructed PPI networks. Additionally, DEGs, such
as CD40LG (adjusted P=0.0023), NOS1 (adjusted
P=0.00021), STRN (adjusted P=0.00011) and MYH11
(adjusted P=0.0011), the miRNA hsa-miR-125b and the TF MCM4
were highlighted in the TF-target-miRNA networks.
Inflammation has an important role in IA formation
and rupture (22,23). Prior to IA rupture, inflammatory
changes in the IA walls, such as T cells and macrophage
infiltration and complement activation, are major events (24,25).
Pyysalo et al (26) have
reported that the etiology of the inflammatory alteration in
patients with IA may be associated with odontogenic bacteria.
Bacterial DNA was detected in 58% IA samples from the ruptured IA
sac tissue, and the overexpression of toll-like receptor 2
(TLR2) has also been reported (26), suggesting that the inflammation was
bacteria-driven. Similarly, the findings of the present study
revealed that the expression of TLR2 (adjusted P=0.00002)
was upregulated in IA samples. In addition, the present study
revealed that the majority of the upregulated DEGs were enriched in
the function of ‘immune system development’ (e.g., CD40LG
and CD40) and the pathway of ‘primary immunodeficiency’
(e.g., CD40LG). The CD40 molecule (CD40) is expressed
on macrophages, and matrix metalloproteinase-2 activation in the
CD40LG pathway is crucial for abdominal aortic aneurysm
(27). In addition, Ochs et
al (28) have indicated that
mutation of CD40LG may lead to primary immunodeficiency, and
T-cell immunodeficiency may function as a key player in the
formation of IA. Therefore, this suggested that CD40 and
CD40LG contribute to the progression of IA by participating
in immune system development and primary immunodeficiency.
Additionally, striatin (STRN), a neuronal calmodulin-binding
protein, was downregulated in neuronal dendrites and spines
(29), and the present study found
17 downregulated DEGs (e.g., STRN) that were predicted to be
enriched in neuron projection development. Therefore, the present
study suggested that STRN may be associated with the
development of IA via influencing the neuron projections
development of IA cells.
Liu et al (30) have demonstrated that miR-125b,
which is involved in modulating the cell proliferation of vascular
smooth muscle cells, is closely associated with IA. The integrated
network analysis revealed that nitric oxide synthase 1
(NOS1), a target gene of miR-125b, was predicted to be
associated with IA. The overexpression of NOS1 (also termed
neuronal NOS) may promote vascular nitric oxide (NO) release, and
NO is responsible for macrophage-mediated vascular smooth muscle
cell apoptosis (31,32). Therefore, the present study
proposed that regulation of NOS1 by miR-125b may contribute
to the progression of IA via modulating cell proliferation and
apoptosis of vascular smooth muscle cells. Additionally, the
mutation of minichromosome maintenance complex component 4
(MCM4) is involved in natural killer cell deficiency in
humans (33), and the natural
killer cell cytotoxicity pathway is closely associated with
abdominal aortic aneurysms (34).
In the present study, the MCM4 TF was shown to be at a high
degree in the integrated network and was predicted to regulate the
expression of MYH11. Additionally, mutation of myosin heavy
chain 11 (MYH11) was inversely correlated with RIA (35), which suggested that the MCM4
TF regulates MYH11 for participating in the development of
IA.
In conclusion, immune system development-associated
DEGs, such as CD40LG and CD40, and regulation of cell
proliferation and apoptosis of vascular smooth muscle cells by DEGs
and miRNAs, such as NOS1 and miR-125b, as well as neuron
projection development-associated DEGs, such as STRN, may
contribute to the progression of IA. Additionally, these may
provide potential therapeutic targets and act as diagnostic
biomarkers for IA. However, further studies are required to verify
these findings.
Acknowledgements
The present study was supported by Shanghai
Municipal Commission of Health and Family Planning (grant no.
201440319) and Pudong New Area Municipal Commission of Health and
Family Planning (grant no. PW2014A-15).
References
|
1
|
Chalouhi N, Hoh BL and Hasan D: Review of
cerebral aneurysm formation, growth, and rupture. Stroke.
44:3613–3622. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blignaut Gerrit, Loggenberg Eugene, Vries
D and Coert: The radiological appearance of intracranial aneurysms
in adults infected with the human immunodeficiency virus (HIV).
Orig Res Sa J Radiol. 18:1–4. 2014.
|
|
3
|
Brown RD Jr: Management of cerebral
aneurysms. Mayo Clinic Proceedings. 79:288–302. 2004. View Article : Google Scholar
|
|
4
|
Olafsson E, Hauser WA and Gudmundsson G: A
population-based study of prognosis of ruptured cerebral aneurysm:
Mortality and recurrence of subarachnoid hemorrhage. Neurology.
48:1191–1195. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Varble N and Meng H: Vortex imprints at
the wall, but not in the bulk, distinguish ruptured from unruptured
intracranial aneurysms. Presented at the DFD15 Meeting of The
American Physical Society. (abstract L23.00008). 2015, http://meetings.aps.org/link/BAPS.2015.DFD.L23.8
|
|
6
|
Lall RR, Eddleman CS, Bendok BR and Batjer
HH: Unruptured intracranial aneurysms and the assessment of rupture
risk based on anatomical and morphological factors: Sifting through
the sands of data. Neurosurg Focus. 26:E22009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mohan D, Munteanu V, Coman T and Ciurea
AV: Genetic factors involves in intracranial aneurysms-actualities.
J Med Life. 8:336–341. 2015.PubMed/NCBI
|
|
8
|
Akagawa H, Tajima A, Sakamoto Y, Krischek
B, Yoneyama T, Kasuya H, Onda H, Hori T, Kubota M, Machida T, et
al: A haplotype spanning two genes, ELN and LIMK1, decreases their
transcripts and confers susceptibility to intracranial aneurysms.
Hum Mol Genet. 15:1722–1734. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu J, Ma F, Yan W, Qiao S, Xu S, Li Y, Luo
J, Zhang J and Jin J: Identification of the soluble form of
tyrosine kinase receptor Axl as a potential biomarker for
intracranial aneurysm rupture. BMC Neurol. 15:232015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ollikainen E, Tulamo R, Lehti S,
Lee-Rueckert M, Hernesniemi J, Niemelä M, Ylä-Herttuala S, Kovanen
PT and Frösen J: Smooth muscle cell foam cell formation,
apolipoproteins and ABCA1 in intracranial aneurysms: Implications
for lipid accumulation as a promoter of aneurysm wall rupture. J
Neuropathol Exp Neurol. 75:689–699. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo J, Jin H, Jiang Y, Ge H, Wang J and Li
Y: Aberrant Expression of microRNA-9 contributes to development of
intracranial aneurysm by suppressing proliferation and reducing
contractility of smooth muscle cells. Med Sci Monit. 22:4247–4253.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu X, Zhang J, Huang Q, Yang P, Chen J and
Liu J: MicroRNA-92a regulates expression of kruppel-like factor2 in
rabbit model of intracranial aneurysm. Cell Mol Biol
(Noisy-le-grand). 61:44–48. 2015.PubMed/NCBI
|
|
13
|
Aoki T, Kataoka H, Nishimura M, Ishibashi
R, Morishita R and Miyamoto S: Ets-1 promotes the progression of
cerebral aneurysm by inducing the expression of MCP-1 in vascular
smooth muscle cells. Gene Ther. 17:1117–1123. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakaoka H, Tajima A, Yoneyama T, Hosomichi
K, Kasuya H, Mizutani T and Inoue I: Gene expression profiling
reveals distinct molecular signatures associated with the rupture
of intracranial aneurysm. Stroke. 45:2239–2245. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li C, Wei Z and Li H: Network-based
empirical bayes methods for linear models with applications to
genomic data. J Biopharm Stat. 20:209–222. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Statist Soc B. 57:289–300. 1995.
|
|
18
|
da Huang W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Franceschini A, Szklarczyk D, Frankild S,
Kuhn M, Simonovic M, Roth A, Lin J, Minguez P, Bork P, von Mering C
and Jensen LJ: STRING v9.1: Protein-protein interaction networks,
with increased coverage and integration. Nucleic Acids Res.
41:(Database issue). D808–D815. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang Y, Li M, Wang J, Pan Y and Wu FX:
CytoNCA: A cytoscape plugin for centrality analysis and evaluation
of protein interaction networks. Bio Systems. 127:67–72. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang Q, Wang Y, Hao Y, Juan L, Teng M,
Zhang X, Li M, Wang G and Liu Y: miR2Disease: A manually curated
database for microRNA deregulation in human disease. Nucleic Acids
Res. 37:(Database issue). D98–D104. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chalouhi N, Ali MS, Jabbour PM,
Tjoumakaris SI, Gonzalez LF, Rosenwasser RH, Koch WJ and Dumont AS:
Biology of intracranial aneurysms: Role of inflammation. J Cereb
Blood Flow Metab. 32:1659–1676. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Aoki T, Fukuda M and Narumiya S: Chronic
inflammation in intracranial aneurysm formation. Inflamm Regen.
33:283–287. 2013. View Article : Google Scholar
|
|
24
|
Tulamo R, Frösen J, Junnikkala S, Paetau
A, Pitkäniemi J, Kangasniemi M, Niemelä M, Jääskeläinen J, Jokitalo
E, Karatas A, et al: Complement activation associates with saccular
cerebral artery aneurysm wall degeneration and rupture.
Neurosurgery. 59:1076–1077. 2006. View Article : Google Scholar
|
|
25
|
Kataoka K, Taneda M, Asai T, Kinoshita A,
Ito M and Kuroda R: Structural fragility and inflammatory response
of ruptured cerebral aneurysms. A comparative study between
ruptured and unruptured cerebral aneurysms. Stroke. 30:1396–1401.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pyysalo MJ, Pyysalo LM, Pessi T, Karhunen
PJ and Öhman JE: The connection between ruptured cerebral aneurysms
and odontogenic bacteria. Öhman JE. 84:1214–1218. 2013.
|
|
27
|
Nagashima H, Aoka Y, Sakomura Y, Uto K,
Sakuta A, Aomi S, Kurosawa H, Hagiwara N, Kawana M and Kasanuki H:
Matrix metalloproteinase 2 is suppressed by trapidil, a CD40-CD40
ligand pathway inhibitor, in human abdominal aortic aneurysm wall.
J Vasc Surg. 39:447–453. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ochs HD, Hollenbaugh D and Aruffo A: The
role of CD40L (gp39)/CD40 in T/B cell interaction and primary
immunodeficiency. Semin Immunol. 6:337–341. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bartoli M, Ternaux JP, Forni C, Portalier
P, Salin P, Amalric M and Monneron A: Down-regulation of striatin,
a neuronal calmodulin-binding protein, impairs rat locomotor
activity. J Neurobiol. 40:234–243. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu D, Han L, Wu X, Yang X, Zhang Q and
Jiang F: Genome-wide microRNA changes in human intracranial
aneurysms. BMC Neurol. 14:1882014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Boyle JJ, Weissberg PL and Bennett MR:
Human macrophage-induced vascular smooth muscle cell apoptosis
requires NO enhancement of Fas/Fas-L interactions. Arterioscler
Thromb Vasc Biol. 22:1624–1630. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li J, Feng W, Chen L and He J:
Downregulation of SMC1A inhibits growth and increases apoptosis and
chemosensitivity of colorectal cancer cells. J Int Med Res.
44:67–74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hughes CR, Guasti L, Meimaridou E, Chuang
CH, Schimenti JC, King PJ, Costigan C, Clark AJ and Metherell LA:
MCM4 mutation causes adrenal failure, short stature, and natural
killer cell deficiency in humans. J Clin Invest. 122:814–820. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hinterseher I, Schworer CM, Lillvis JH,
Stahl E, Erdman R, Gatalica Z, Tromp G and Kuivaniemi H:
Immunohistochemical analysis of the natural killer cell
cytotoxicity pathway in human abdominal aortic aneurysms. Int J Mol
Sci. 16:11196–11212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ravindra VM, Karsy M, Schmidt RH, Taussky
P, Park MS and Bollo RJ: Rapid de novo aneurysm formation after
clipping of a ruptured middle cerebral artery aneurysm in an infant
with an MYH11 mutation. J Neurosurg Pediatr. 18:463–470. 2016.
View Article : Google Scholar : PubMed/NCBI
|