Introduction
With continuous promotion of castration and
anti-androgen therapy, clinical treatment of androgen independent
protate cancer or castration-resistant prostate cancer (CRPC) has
become difficult. It is not uncommon that CRPC develops metastases
that chemotherapy and radiotherapy have limited effects on, which
seriously affects patients' quality of life. Therefore, research on
mechanisms of CRPC progression seems particularly important
(1–3). The tumor microenvironment is
essential for tumor genesis and tumor development (4,5),
with hypoxia a strong research topic in recent years. Hypoxia can
induce vascular formation in tumors, and is also widely involved in
tumor formation, development, metastasis and recurrence (6–8).
Hypoxia accelerates epithelial-mesenchymal transition, invasion,
and metastasis in prostate cancer. Also, hypoxia may lead to a
decreased sensitivity to radiotherapy and chemotherapy in prostate
cancer treatment (9–12). However, there have been scant
studies on hypoxia-induced immune evasion in prostate cancer.
Therefore, we carried out this study to discover the role of
hypoxia in tumor immune regulation.
Hypoxia is involved in immune evasion of a variety
of tumors (13) involving many
types of immune cells, including T cells, natural killer (NK)
cells, macrophages and dendritic cells, that can inhibit or kill
tumors (13). Hypoxia may lead to
upregulation of the expression of stem cell marker Nanog and
transforming growth factor beta 1, resulting in low immune killing
capacity of T lymphocytes and macrophages against tumor cells
(14). It was discovered in a lung
cancer and melanoma study that hypoxia could induce miR-210
expression, which decreased tumor cell susceptibility to
antigen-specific cytotoxic T lymphocytes and led to tumor formation
and development (15). The NK cell
mediated immune response can kill tumor cells directly with no
dependence on antibodies or complements, which is a unique
advantage in tumor immunity. By improving immune killing ability of
NK cells against tumor cells, tumor formation and development can
be effectively controlled. Suppression of expression of NK cell
activating receptors MICA and MICB on the tumor cell surface by
hypoxia can cause immune evasion from NK cells in pancreatic
cancer, osteosarcoma, multiple myeloma and other malignant tumors
(16–19). The role of hypoxia regarding NK
cell immune evasion in prostate cancer is rarely reported. In a
study of DU145 and PC3 in prostate cancer cells, hypoxia inhibited
the expression of NKG2D ligands on the surface of the tumor cells,
thereby inhibiting the killing of tumor cells by activated NK cells
(20,21).
The mechanism of hypoxia-mediated immune evasion is
unknown. Many studies have indicated that programmed cell death
ligand 1 (PD-L1) plays an important role in tumor immune evasion
(22,23). A study of non-small cell lung
cancer showed that tumor cells overexpress PD-L1, thereby binding
PD-1 receptors on the surface of T cells and inhibiting T cell
immune attack, resulting in immune evasion (24,25).
Studies of ovarian cancer, melanoma, bladder cancer, laryngeal
squamous cell carcinoma and other malignant tumors have indicated
that downregulation of PD-1/PD-L1 improves tumor susceptibility to
immune cells (26–30). PD-1 is expressed in multiple immune
cells, including T cells, B cells, NK cells and dendritic cells
(31,32). The effect of PD-L1 in immune
evasion of CRPC from NK cells, which is rarely studied, became the
research direction of these experiments.
NKG2D is an important receptor for activation of NK
cells. The upregulation of expression of NKG2D ligands, MHC class I
chain-related proteins A and B (MICA and MICB) and UL16-binding
proteins ULBP-1, ULBP-2, ULBP-3, can promote immune cytolytic
activity of NK cells to tumors (33). This receptor has become the subject
of enhancing the anti-tumor immunity of NK cells in this study.
Previous work has shown that the combination of PD-1 and PD-L1
constitutes the PD-1-PD-L1 signaling pathway; in addition to this
pathway's immunosuppressive effects through T cells (34), it can also inhibit the anti-tumor
activity of NK cells by inhibiting the function of NKG2D ligands on
the surface of tumor cells (35).
Interestingly, based on a review of the literature, we found that
the JAK1,2/Stat3 signaling pathway is the upstream
regulator of PD-L1, which also includes other widely studied
regulators such as Akt, MAPK, MEK, NFκB, and mTOR (36–40).
We selected CRPC cell lines C4-2 and CWR22Rv1 as
subjects of study. After induction of the cells by hypoxia, we
assayed NK cell mediated cytotoxicity and colony formation to
determine whether there were any changes in CRPC cell immune
susceptibility to NK cells. We investigated the underlying
molecular mechanisms of the changes to seek an effective way to
enhance the killing ability of NK cells against CRPC cells, with
the aim of inhibiting or killing tumors and offering a new approach
to CRPC immunotherapy.
Materials and methods
Cell culture
The C4-2 cells used in the assays were generously
donated by the China Center for Type Culture Collection (CCTCC).
The CWR22Rv1 and NK92 cells were purchased from the American Type
Culture Collection (ATCC, Manassas, VA, USA) and the cell culture
medium was RPMI 1,640 medium (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% charcoal stripped fetal bovine
serum (Thermo Fisher Scientific, Inc.). We used an anoxic incubator
(1%O2, 5%CO2, 94%N2) (Sanyo,
Osaka, Japan) and a normoxic incubator (21%O2,
5%CO2, 74%N2) (Sanyo). C4-2 and CWR22Rv1
lines of CRPC cells grown inananoxic incubator for 24 h served as
anoxic cells. NK92 cells were cultured in Minimum Essential Medium
containing sodium bicarbonate (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany), IL-2 (Bio-Techne, Minneapolis, MN, USA),
inositol, folic acid, 12.5% horse serum (Sigma-Aldrich; Merck
KGaA), 2-mercaptoethanol (Bio-Rad Laboratories, Inc., Hercules, CA,
USA), and 12.5% FBS (HyClone; GE Healthcare Life Sciences, Logan,
UT, USA).
NK cell mediated cytotoxicity
assay
Tumor cells were seeded at 2,000 cells/well in a
96-well plate and cultured overnight. After aspirating all the
medium, NK cells were added at a ratio of tumor cells to NK cells
of 1:1, 1:5, or 1:15; the inhibitors PD-L1 Ab (329710; BioLegend,
Inc., San Diego, CA, USA), JAK inhibitor 1 (CAS457081-03-7; EMD
Millipore, Billerica, MA, USA), and Stattic were added at a ratio
of 1:1,000 at the same time. Cells were cultured for 4 h, then a 50
µl aliquot of medium was used in a LDH cytotoxic assay using the
LDH cytotoxic assay kit (88954; Thermo Fisher Scientific, Inc.).
The experimental release was corrected by subtracting the amount
released spontaneously in cells at corresponding dilutions. The
percentage cytotoxicity was expressed as
Experimental value-Effector cells
spontaneous control-Target cells spontaneous controlTarget cell
maximum control-Target cells spontaneous control×arg
and used to calculate the immune killing ability of
NK cells against tumor cells.
Colony formation assay
Using the gradient dilution cell-count method, we
seeded 200 cells in 6 cm dishes (REF353002; Corning Incorporated,
Corning, NY, USA). After overnight culture, tumor cells and NK
cells were incubated for 4 h at 1:1, 1:5 and 1:15 ratios. After
aspirating all the medium and removing NK cells, tumor cells were
continuously cultured for 14 days after replacing the conventional
culture medium. Then the target cells were fixed with formaldehyde
for 15 min, stained with crystal violet for 30 min, and observed
and photographed under the microscope. The protocol was repeated
three times in each group and the results were averaged.
Total RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
When the adherent tumor cells had grown over about
70% of the surface, PD-L1 Ab, JAK inhibitor 1, Stattic, LY294002,
SB203580, and U0126 were added separately at a ratio of 1:1,000,
then cells were cultured for 6 h, and total RNA was extracted. We
used Superscript III transcriptase (Invitrogen; Thermo Fisher
Scientific, Inc.) to reverse-transcribe total RNA (1 µg). Following
primer setup, (the primer solutions included RT buffer, dNTPs, RT
random primers, MultiScribe reverse transcriptase and RNase-free
water), we conducted qPCR with a Bio-Rad CFX96 reaction system
(reactions included cDNA, RNAase-free water, SYBR Green and
primers; Table I). Using GAPDH as
a reference, the expression of target gene mRNA was measured by the
intensity of green fluorescence.
 | Table I.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Direction | Sequence |
|---|
| GAPDH | Forward |
5′-AACGGATTTGGTCGTATTGGG-3′ |
|
| Reverse |
5′-CCTGGAAGATGGTGATGGGAT-3′ |
| PD-L1 | Forward |
5′-GCTATGGTGGTGCCGACTAC-3′ |
|
| Reverse |
5′-TTGGTGGTGGTGGTCTTACC-3′ |
| MICA | Forward |
5′-ACTGCTTGAGCCGCTGAGA-3′ |
|
| Reverse |
5′-GAGGTGCAAAAGGGAAGATGC-3′ |
| MICB | Forward |
5′-GGGGCGCAGGTGACTAAAT-3′ |
|
| Reverse |
5′-CCTACGTCGCCACCTTCTCA-3′ |
| ULBP-1 | Forward |
5′-CAGCAGACGATGAGGACATT-3′ |
|
| Reverse |
5′-GACAGAAAGTGGCAGAAGGTG-3′ |
| ULBP-2 | Forward |
5′-CATTACTTCTCAATGGGAGACTGT-3′ |
|
| Reverse |
5′-TGTGCCTGAGGACATGGCGA-3′ |
| ULBP-3 | Forward | 5′-
ATTCTTCCGTACCTGCTATT-3′ |
|
| Reverse |
5′-GCTATCCTTCTCCCACTTCT-3′ |
Western blot analysis
Tumor cells were collected after centrifugation and
the supernatant removed. Then cell lysate was added, the protein
concentration measured, sodium dodecyl sulphate-polyacrylamide gel
electrophoresis performed, and the protein transferred to
polyvinylidene difluoride membranes (EMD Millipore). After adding
blocking solution and incubating with primary antibodies (1:1,000),
the membranes were incubated with horseradish peroxidase labeled
secondary antibody (1:5,000), followed by capture of images with an
ECL imaging system (Thermo Fisher Scientific, Inc.). Primary
antibodies used in the assay included p-HIF-1α (EP1215Y; ab210073;
Abcam, Cambridge, UK), p-PD-L1 (MAB1086; R&D Systems, Inc.,
Minneapolis, MN, USA), JAK1 (pY1022, A7125; Assay
Biotech, Fremont, CA, USA), p-JAK2 (pY1007 + 1008,
601–670; Abbomax, Inc., San Jose, CA, USA), p-MAPK (9101S; Cell
Signaling Technology, Inc., Danvers, MA, USA), p-MEK (Ser 217/221,
9121; Cell Signaling Technology, Inc.), p-Akt (S473, 9271; Cell
Signaling Technology, Inc.), p-Stat3 (Y705, ab76315; Abcam), p-NFκB
(S536, ab86299; Abcam), and GAPDH (2118S; Cell Signaling
Technology, Inc.).
Signaling pathway inhibitors
We added appropriately diluted inhibitors of various
cell signaling pathways, JAK inhibitor 1 (5 µM) (CAS457081-03-7;
EMD Millipore), Stattic (10 µM) (CAS19983-44-9; EMD Millipore),
PD-L1 Ab (329710; BioLegend, Inc.), LY294002, SB203580 (both
Sigma-Aldrich; Merck KGaA) and U0126 (Cell Signaling Technology,
Inc.), which respectively inhibit JAK1, JAK2, Stat3, Akt, MAPK and
MEK, to hypoxia-treated cells before co-incubation with tumor
cells.
Statistical analysis
All data were reported as the mean ± standard
deviation of 3 experimental repeats., and were analyzed using SPSS
19.0 software (IBM Corp., Armonk, NY, USA). The differences between
two groups were analyzed by a two-tailed Student's t-test. One-way
analysis of variance followed by Fisher's least significant
difference post hoc test were used for comparisons among multiple
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
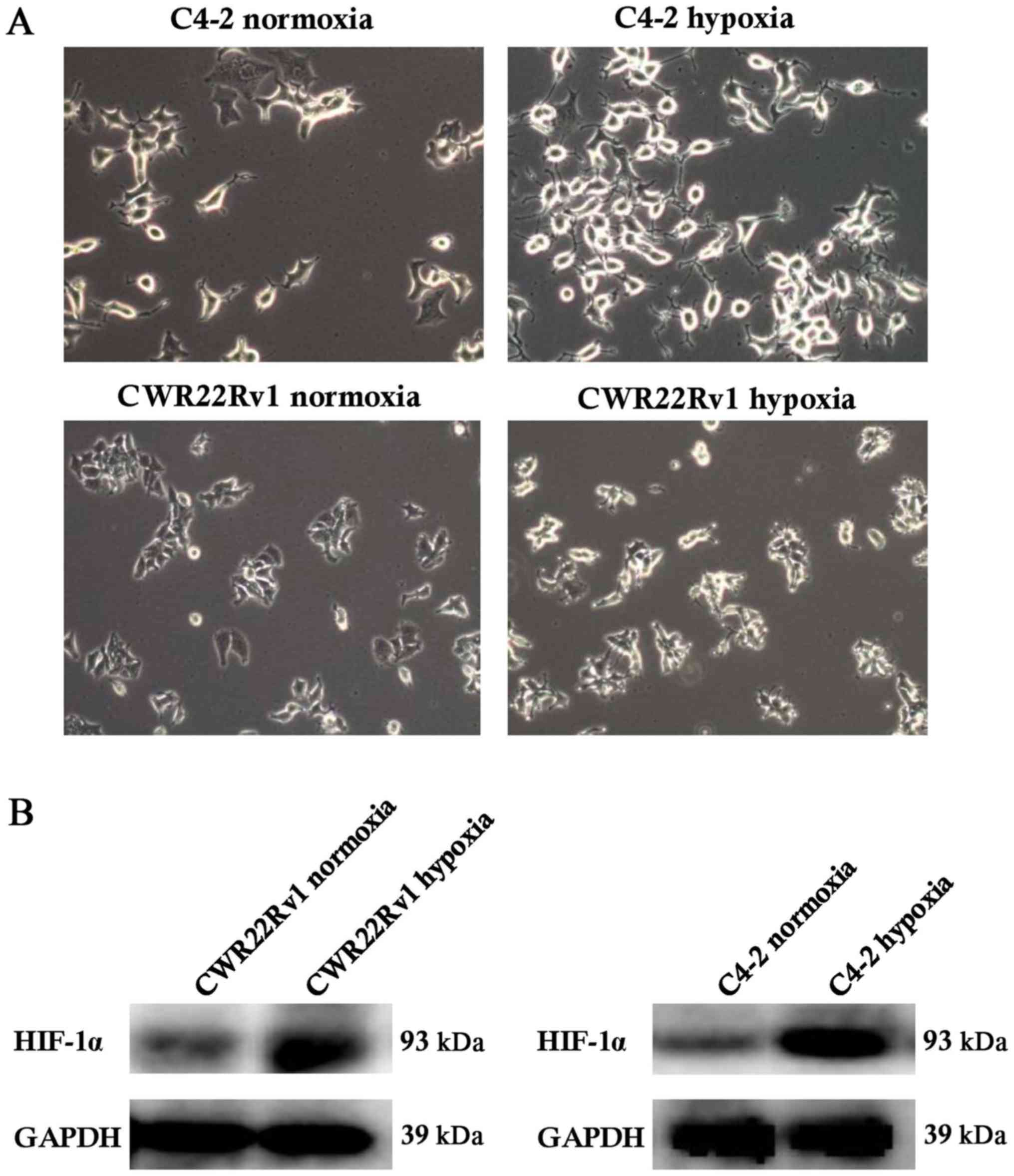
Hypoxia-induced C4-2 and CWR22Rv1
cells showed changes in shape and increased expression of
hypoxia-inducible factor-1α (HIF-1α)
After culture of C4-2 and CWR22Rv1 lines of CRPC
cells in a hypoxic incubator for 24 h, the expected changes in cell
morphology, such as to spindle-shaped and polygonal cells, and an
increase in cell refractive index were observed (Fig. 1A). Normoxic cultured cells were the
control group. We observed significantly upregulated expression of
HIF-1α in tumor cells exposed to hypoxia. While HIF-1α can be
easily degraded by the intracellular oxygen-dependent
ubiquitin-proteasome pathway under normoxic conditions, under
hypoxic conditions, its expression is stable (33), suggesting that the assayed C4-2 and
CWR22Rv1 cells were effectively induced by hypoxia (Fig. 1B).
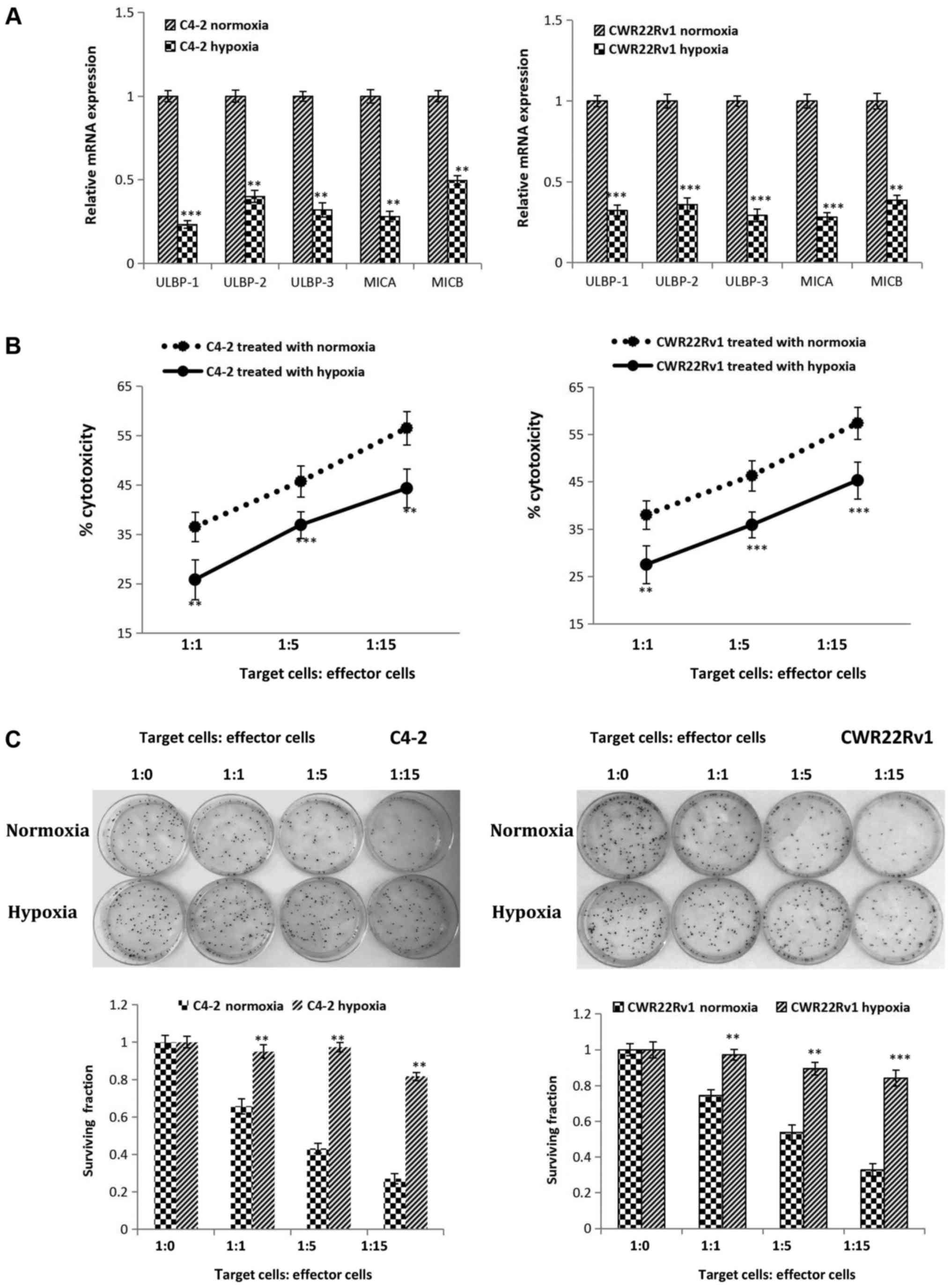
Hypoxia reduced expression of NKG2D
activating ligandsin C4-2 and CWR22Rv1 cell lines and led to
increased tolerance to immune killing of NK cells
We also detected changes in expression of genes
encoding NKG2D ligands in hypoxia-induced C4-2 and CWR22Rv1 cells.
Gene expression of these ligands (ULBP-1, ULBP-2, ULBP-3, MICA and
MICB, all found on the surface of target cells) significantly
decreased (Fig. 2A). In
hypoxia-induced CRPC cells co-cultured for 4 h with various ratios
of NK cells, lactate dehydrogenase release showed that the killing
ability of NK cells significantly decreased against hypoxia induced
target cells (Fig. 2B). The colony
formation assay suggested that as the ratio of co-cultured NK cells
to tumor cells increased, the clone-forming ability of the hypoxia
group showed no obvious changes, while a significant decrease was
observed in the normoxia group. These results indicated that
hypoxia increased survival of target cells co-cultured with NK
cells, which was consistent with the findings in the NK cell
mediated cytotoxicity assay (Fig.
2C). In conclusion, hypoxia can lead to CRPC cell immune
tolerance to NK cells.
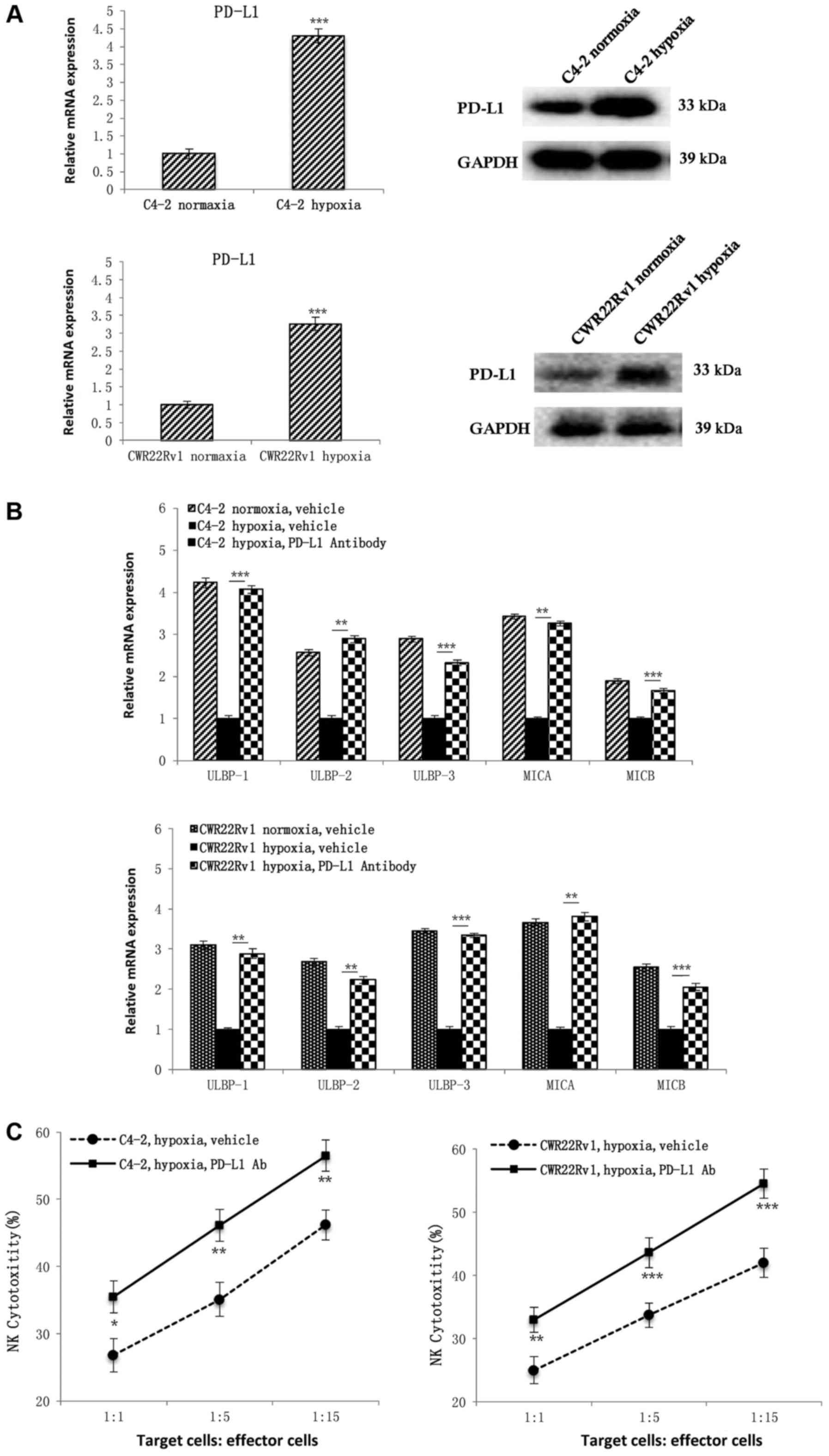
Hypoxia-induced C4-2, and CWR22Rv1
CRPC cells had increased PD-L1 expression, resulting in immune
escape from NK cells
In further study on the mechanisms affecting
expression of PD-L1, both PD-L1 gene and protein expression in C4-2
and CWR22Rv1 cells were higher after hypoxia induction than under
normoxic conditions (Fig. 3A). To
determine whether hypoxia induces CRPC cells to evade NK cell
immunity, we added PD-L1 antibodies to hypoxia-induced CRPC cells
and detected upregulation of NKG2D ligand expression (Fig. 3B). We assayed NK cell mediated
cytotoxicity, and discovered that addition of PD-L1 antibodies
significantly increased the susceptibility of hypoxia-induced CRPC
cells to NK cell immunity (Fig.
3C), which confirmed that PD-L1 protein is involved in
hypoxia-induced CRPC cell immune evasion.
PD-L1 expression is inhibited by
blocking the JAK1,2/Stat3 signaling pathway
To explore the mechanism behind PD-L1 regulation of
CRPC cells against NK cell immunity, we selected well-studied
signaling pathways of PD-L1 regulation, including JAK1,
JAK2, Stat3, PI3K/Akt, MAPK, MEK, and NFκB (34–39).
We first detected differences in expression of these molecular
markers in both normoxia and hypoxia groups by western blotting,
and found that the expression of JAK1, JAK2,
Stat3, Akt, MAPK, MEK in hypoxia group was increased compared with
the normoxia group (Fig. 4A);
thus, we think that these proteins may be involved in the
regulation of PD-L1 under hypoxic conditions in the tumor. To
observe their regulation of PD-L1, we added inhibitors of these
proteins' corresponding signaling pathways JAK inhibitor 1 (which
can simultaneously inhibit the expression of JAK 1 and JAK 2),
Stattic, LY294002, SB203580 and U0126 at a ratio of 1:1,000 to
hypoxic tumor cells. After 6 h, the level of PD-L1 was detected by
RT-qPCR. The untreated group served as a control. The results
showed that JAK inhibitor 1 and Stattic significantly down-regulate
PD-L1 expression. PI3k inhibitor LY294002 could also significantly
reduce the expression of PD-L1, but JAK inhibitor 1 and Stattic
inhibited PD-L1 more. So we chose JAK inhibitor 1 and Stattic as
the research subjects. Of course, given this result, PI3k inhibitor
LY294002 will also be our future research direction (Fig. 4B).
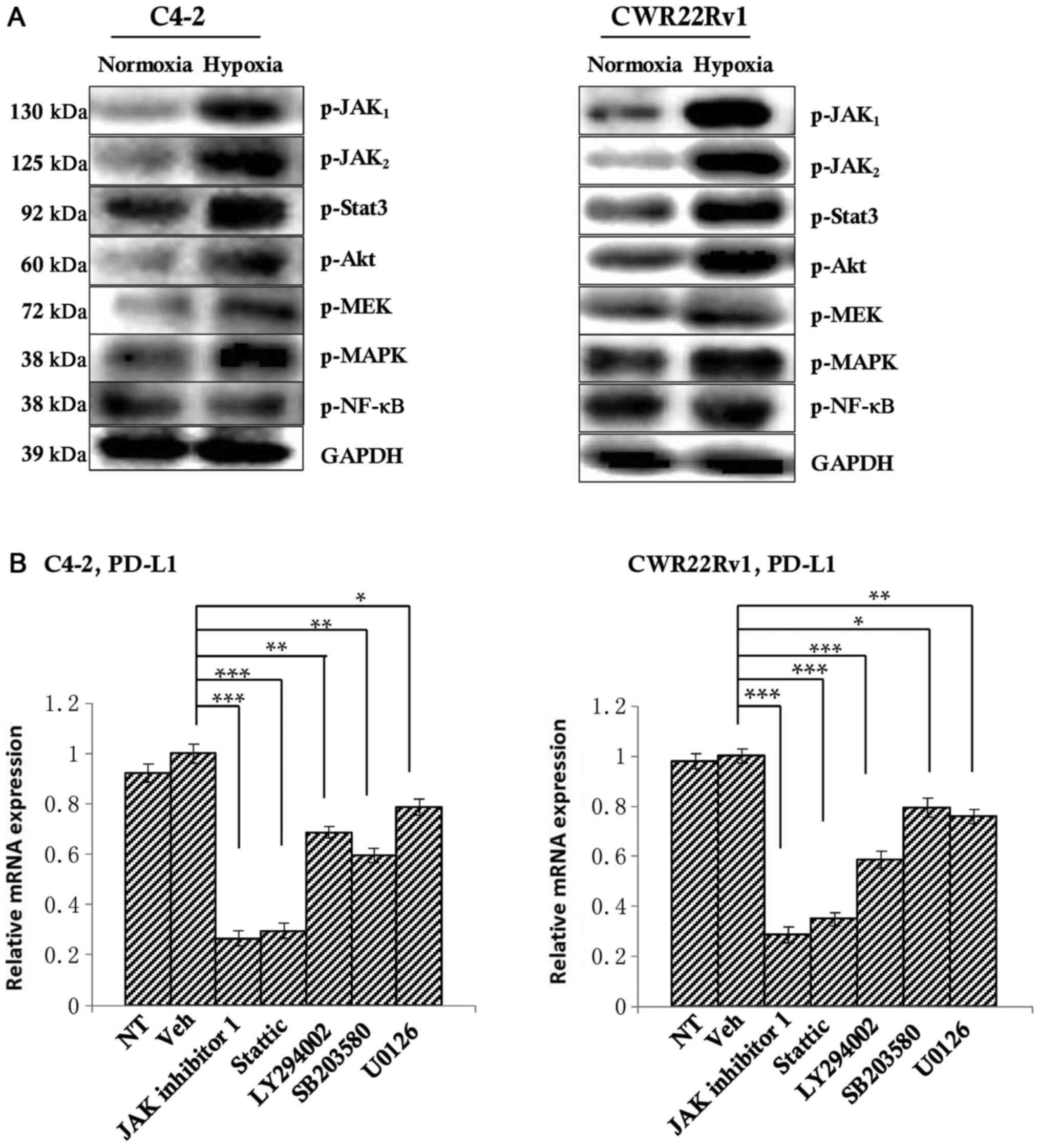 | Figure 4.Detection of the upstream regulatory
factors of PD-L1. (A) Western blotting indicated an upward tendency
in the expression of p-JAK1, p-JAK2, p-Akt,
p-Stat3, p-MAPK and p-MEK in the hypoxia group. (B) Once the
corresponding inhibitors were added, PD-L1 expression in all of the
above signaling pathways decreased to varying degrees, with
significant decreases observed following the addition of JAK
inhibitor 1 and Stattic, suggesting that the inhibition of the
JAK1,2/Stat3 signaling pathway may inhibit PD-L1
expression. *P<0.05, **P<0.01 and ***P<0.001, as
indicated. PD-L1, programmed death-ligand 1; JAK, Janus kinase;
Akt, protein kinase B; Stat, signal transducer and activator of
transcription; NF, nuclear factor; MAPK, mitogen-activated protein
kinase; MEK, MAPK kinase; p-, phosphorylated; NT, not treated. |
JAK1,2/Stat3 signaling pathway inhibition
increases the susceptibility of CRPC cells to NK cell immunity,
with a combination of PD-L1 antibodies and JAK inhibitor 1/Stattic
more effective either alone. CRPC cells in the hypoxia group were
used as the study subject, and the JAK1,2 inhibitor JAK
inhibitor 1 or the Stat3 inhibitor Stattic were added. The
untreated group was used as the control. RT-qPCR showed increased
expression of NKG2D ligands (Fig.
5A), suggesting that inhibition of the JAK1,2/Stat3
signaling pathway may improve the immune cytolytic activity of NK
cells toward CRPC cells under hypoxic conditions.
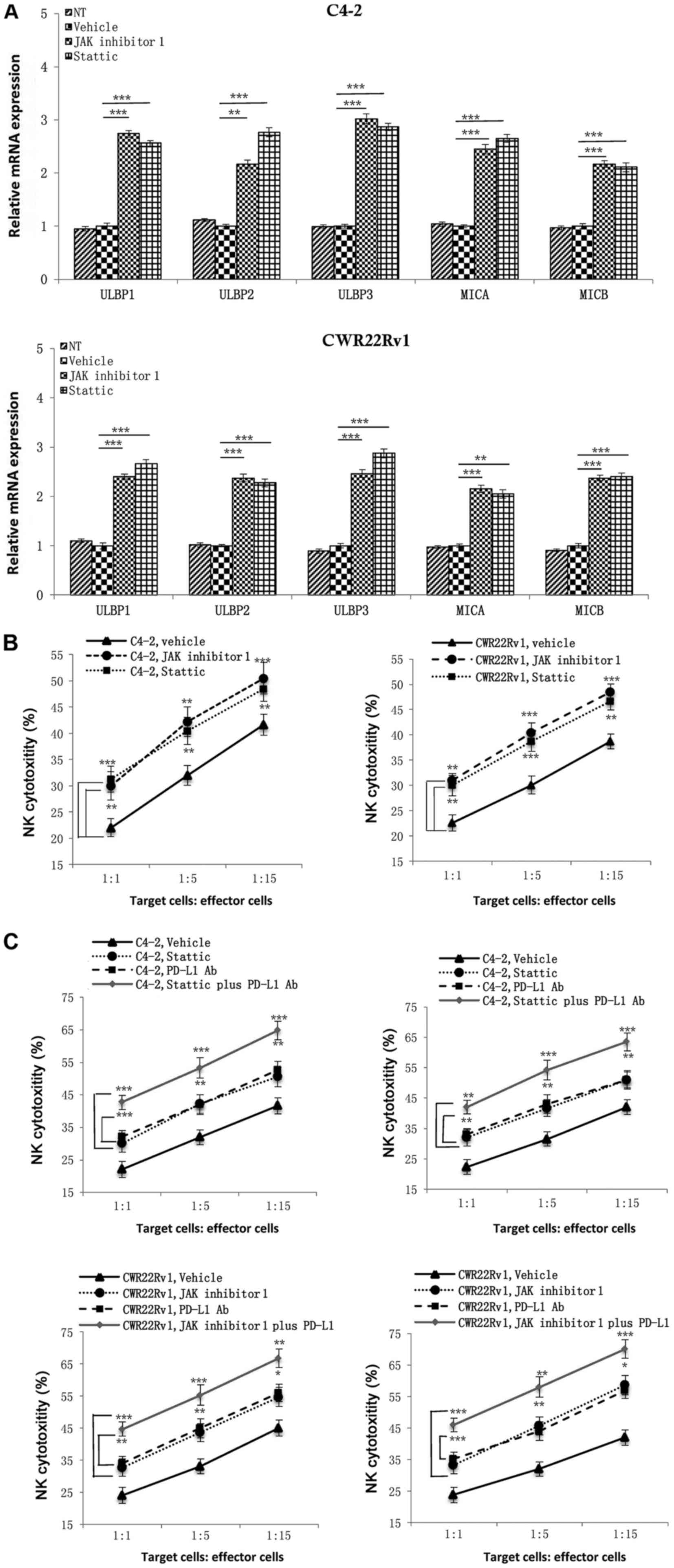 | Figure 5.Effects of JAK1,2 and
Stat3 single inhibition and JAK1,2/PD-L1 and Stat3/PD-L1
combined inhibition on NK cell mediated cytotoxicity. (A)
Inhibition of the expression of JAK1,2 or Stat3
upregulated NKG2D ligand expression in castration-resistant
prostate cancer cells under hypoxia. (B) NK cell cytotoxic effects
on tumor cells were significantly increased following the addition
of JAK inhibitor 1 or Stattic in the hypoxia group. (C)
Combinations of JAK inhibitor 1/PD-L1 antibody or Stattic/PD-L1
antibody were superior to single applications, indicating that
combined inhibition of the PD-L1 and JAK1,2/Stat3
signaling pathways enhanced NK cell immune killing ability against
hypoxia-induced CRPC cells. *P<0.05, **P<0.01 and
***P<0.001, as indicated. JAK, Janus kinase; Stat, signal
transducer and activator of transcription; NKG2D, natural killer
group 2D; NK cells, natural killer cells; PD-L1, programmed
death-ligand 1; MICA, MHC class I chain-related protein A; MICB,
MHC class I chain-related protein B; ULBP, UL16 binding protein;
NT, not treated. |
Since blocking the JAK1,2/Stat3 signaling
pathway can inhibit PD-L1 expression in hypoxic CRPC cells and may
upregulate expression of NKG2D ligands of CRPC cells, to test
whether this also enhances the immune killing function of NK cells
against tumors by down-regulating PD-L1, we used hypoxic CRPC cells
as targets and added the same JAK1,2/Stat3 signaling
pathway inhibitors to an NK cell mediated cytotoxicity assay. Both
JAK inhibitor 1 and Stattic significantly increased susceptibility
of CRPC cells to NK cell immunity compared to an untreated group
(Fig. 5B). Finally, we added
combinations of PD-L1 antibody/JAK inhibitor 1 and PD-L1
antibody/Stattic to the NK cell mediated cytotoxicity assay and
discovered that combined treatments were superior to each single
application. Combined treatments achieved better results in
enhancing NK cell immune killing function against CRPC cells, which
could provide new approaches to CRPC targeted therapy (Fig. 5C).
Discussion
In clinical practice, the incidence of both local
progression and distant metastasis in CRPC patients has
significantly increased, seriously affecting patients' quality of
life (40,41). Although the mechanisms behind this
increase are not yet fully understood, hypoxia, as an important
feature of the tumor microenvironment, plays an essential role
(42,43). In this study, CRPC cell lines C4-2
and CWR22Rv1 were grown in an incubator under conditions of
hypoxia. We observed changes in the morphology of these cells to
polygonal and spindle shapes, and high expression of HIF-1α that
was dependent on the oxygen concentration, consistent with
hypoxia-induced changes in the cells, which may provide fundamental
data for further exploration of the effects of hypoxia on tumor
immunity. In the hypoxic microenvironment of prostate cancer,
HIF-1α and related proteins present a major research direction. Our
focus on the JAK1,2/Stat3-PD-L1 signaling pathway has
not been studies before, and our results suggest this pathway was
upregulated in a hypoxic microenvironment, resulting in immune
escape of CRPC from NK cells.
The occurrence and development of tumors are closely
related to the immunity of the human body, in which the direct
killing function of NK cells against tumors has a unique advantage;
however, there are scant studies on the mechanisms of NK cell
immunity against CRPC. Our study took this as a starting point to
explore the mechanisms of NK cell immunity against CRPC in a
hypoxic microenvironment. Firstly we examined the expression of
ligands of NKG2D, a major activating receptor of NK cells, which
include the ULBP family (ULBP-1, ULBP-2 and ULBP-3) and MICA/MICB
(44). Expression of all these
markers showed a downward trend, consistent with NKG2D playing an
important role in tumor immunity of NK cells. CRPC cells also had
stronger immune tolerance to NK cells in a hypoxic
microenvironment, confirming a stronger immune evasion function
under these conditions.
To explore the mechanisms of immune evasion, we
selected PD-L1, a relatively well-studied protein in tumor targeted
therapy (45,46), as a research subject. PD-L1
expression of hypoxia-induced CRPC cells significantly increased,
and by adding PD-L1 antibodies, the expression of NKG2D ligands was
reversed, while the immune killing ability of NK cells against
tumor cells was significantly enhanced. The combination of PD-1 and
PD-L1 constitutes the PD-1-PD-L1 signaling pathway, thus affecting
immune cytolytic activity of immune cells to tumor cells, which is
mainly reflected in the following aspects: (1) Inhibition of function of TIL cells by
(a) inhibiting the activation of TIL cells, (b) influencing Th cell
differentiation, (c) inhibiting the production of effector
cytokines, (d) promoting the secretion of suppressive cytokines,
and (e) increasing TIL apoptosis, thus resulting in tumor immune
escape (34); (2) Inhibiting the function of NK cells,
thereby reducing their anti-tumor effect (35); (3)
Expression of PD-L1 on TAM, which can cause synergistic
stimulation, inhibiting the immune susceptibility of tumor cells
(47).
During examination of PD-L1 upstream regulatory
factors, we found that the JAK1,2/Stat3 signaling
pathways played an important role, and by adding appropriate
antibodies (JAK inhibitor 1 or PD-L1 antibody), PD-L1 expression of
CRPC cells was inhibited under hypoxic conditions. The
JAK1,2/Stat3 signaling pathway is widely involved in
proliferation, differentiation, migration and other processes of
cells (48). It plays an important
role in occurrence and development of many tumors including
prostate cancer (49–53). The results of our study also
indicated that in a hypoxic microenvironment, the
JAK1,2/Stat3 signaling pathway promotes immune evasion
of NK cells by CRPC cells. The current targeted immunotherapy of
PD-1/PD-L1 against tumors has achieved good results, effectively
reducing tumor size and prolonging the survival of patients.
However, clinical observation shows that there is still some degree
of immunosuppression, which may be related to the complex immune
escape mechanism of the tumor itself. Combined administration of
drugs through multiple ways may be an effective choice for tumor
targeted immunotherapy (54).
Thus, we studied the combined inhibition of JAK1,2/PD-L1
and Stat3/PD-L1, and observed that a combined inhibition was more
effective in enhancing NK cell cytotoxicity toward hypoxia-induced
CRPC cells. Given these results, we may develop a combination
inhibitor of JAK1,2/Stat3 and PD-L1, and
conduct related animal studies and clinical trials, thereby
providing a more effective means of clinical immunotherapy of
CRPC.
Acknowledgements
The authors would like to thank Dr. Soo Ok Lee
(Department of Urology, The Second Affiliated Hospital of Soochow
University, Suzhou, Jiangsu, China) for assisting with the
preparation of the manuscript.
Funding
The present study was supported by the Pre Research
Fund Project of The Second Affiliated Hospital of Soochow
University (grant no. SDFEYBS1707), The National Natural Science
Foundation of China (grant nos. 81472776 and 81773221), and by
Preponderant Discipline Construction Funding of the Second
Affiliated Hospital of Soochow University (grant no.
XKQ2015008).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
LJX, QM and JZ performed the experiments and
statistical analyses, and created the figures. JL and BXX
contributed to the generation of the knockdown cell lines. JG and
CYS provided and performed the staining of human tissues. YCZ and
YBZ assisted with the interpretation of data and reviewed the
manuscript. DRY and YXS conceived the idea and wrote the
manuscript. All authors reviewed and agreed to the information in
this manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cornford P, Bellmunt J, Bolla M, Briers E,
De Santis M, Gross T, Henry AM, Joniau S, Lam TB, Mason MD, et al:
EAU-ESTRO-SIOG Guidelines on prostate cancer. Part II: Treatment of
relapsing, metastatic, and castration-resistant prostate cancer.
Eur Urol. 71:630–642. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lowrance WT, Roth BJ, Kirkby E, Murad MH
and Cookson MS: Castration-resistant prostate cancer: AUA Guideline
Amendment 2015. J Urol. 195:1444–1452. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Crawford ED, Higano CS, Shore ND, Hussain
M and Petrylak DP: Treating patients with metastatic castration
resistant prostate cancer: A comprehensive review of available
therapies. J Urol. 194:1537–1547. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qiu J, Jiang W, Yang Y, Feng C, Chen Z,
Guan G, Zhuo S and Chen J: Monitoring changes of tumor
microenvironment in colorectal submucosa using multiphoton
microscopy. Scanning. 37:17–22. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Casey SC, Amedei A, Aquilano K, Azmi AS,
Benencia F, Bhakta D, Bilsland AE, Boosani CS, Chen S, Ciriolo MR,
et al: Cancer prevention and therapy through the modulation of the
tumor microenvironment. Semin Cancer Biol. 35 Suppl:S199–S223.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wolff M, Kosyna FK, Dunst J, Jelkmann W
and Depping R: Impact of hypoxia inducible factors on estrogen
receptor expression in breast cancer cells. Arch Biochem Biophys.
613:23–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu X, Qiao B, Liu Q and Zhang W:
Upregulation of extracellular matrix metalloproteinase inducer
promotes hypoxia-induced epithelial-mesenchymal transition in
esophageal cancer. Mol Med Rep. 12:7419–7424. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Clavo B, Robaina F, Fiuza D, Ruiz A,
Lloret M, Rey-Baltar D, Llontop P, Riveros A, Rivero J, Castañeda
F, et al: Predictive value of hypoxia in advanced head and neck
cancer after treatment with hyperfractionated radio-chemotherapy
and hypoxia modification. Clin Transl Oncol. 19:419–424. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li W, Dong Y, Zhang B, Kang Y, Yang X and
Wang H: PEBP4 silencing inhibits hypoxia-induced
epithelial-to-mesenchymal transition in prostate cancer cells.
Biomed Pharmacother. 81:1–6. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li M, Wang YX, Luo Y, Zhao J, Li Q, Zhang
J and Jiang Y: Hypoxia inducible factor-1α-dependent epithelial to
mesenchymal transition under hypoxic conditions in prostate cancer
cells. Oncol Rep. 36:521–527. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang W, Liu M, Guan Y and Wu Q:
Hypoxia-responsive Mir-301a and Mir-301b promote radioresistance of
prostate cancer cells via downregulating NDRG2. Med Sci Monit.
22:2126–2132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nomura T, Yamasaki M, Hirai K, Inoue T,
Sato R, Matsuura K, Moriyama M, Sato F and Mimata H: Targeting the
Vav3 oncogene enhances docetaxel-induced apoptosis through the
inhibition of androgen receptor phosphorylation in LNCaP prostate
cancer cells under chronic hypoxia. Mol Cancer. 12:272013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barsoum IB, Koti M, Siemens DR and Graham
CH: Mechanisms of hypoxia-mediated immune escape in cancer. Cancer
Res. 74:7185–7190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hasmim M, Noman MZ, Messai Y, Bordereaux
D, Gros G, Baud V and Chouaib S: Cutting edge: Hypoxia-induced
Nanog favors the intratumoral infiltration of regulatory T cells
and macrophages via direct regulation of TGF-β1. J Immunol.
191:5802–5806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Noman MZ, Buart S, Romero P, Ketari S,
Janji B, Mari B, Mami-Chouaib F and Chouaib S: Hypoxia-inducible
miR-210 regulates the susceptibility of tumor cells to lysis by
cytotoxic T cells. Cancer Res. 72:4629–4641. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu Y, Hu J, Sun W, Duan X and Chen X:
Hypoxia-mediated immune evasion of pancreatic carcinoma cells. Mol
Med Rep. 11:3666–3672. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yamada N, Yamanegi K, Ohyama H, Hata M,
Nakasho K, Futani H, Okamura H and Terada N: Hypoxia downregulates
the expression of cell surface MICA without increasing soluble MICA
in osteosarcoma cells in a HIF-1α-dependent manner. Int J Oncol.
41:2005–2012. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sarkar S, Germeraad WT, Rouschop KM,
Steeghs EM, van Gelder M, Bos GM and Wieten L: Hypoxia induced
impairment of NK cell cytotoxicity against multiple myeloma can be
overcome by IL-2 activation of the NK cells. PLoS One.
8:e648352013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Labiano S, Palazon A and Melero I: Immune
response regulation in the tumor microenvironment by hypoxia. Semin
Oncol. 42:378–386. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hamilton TK, Hu N, Kolomitro K, Bell EN,
Maurice DH, Graham CH and Siemens DR: Potential therapeutic
applications of phosphodiesterase inhibition in prostate cancer.
World J Urol. 31:325–330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Siemens DR, Hu N, Sheikhi AK, Chung E,
Frederiksen LJ, Pross H and Graham CH: Hypoxia increases tumor cell
shedding of MHC class I chain-related molecule: Role of nitric
oxide. Cancer Res. 68:4746–4753. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barsoum IB, Smallwood CA, Siemens DR and
Graham CH: A mechanism of hypoxia-mediated escape from adaptive
immunity in cancer cells. Cancer Res. 74:665–674. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen J, Jiang CC, Jin L and Zhang XD:
Regulation of PD-L1: A novel role of pro-survival signalling in
cancer. Ann Oncol. 27:409–416. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shi MH, Xing YF, Zhang ZL, Huang JA and
Chen YJ: Effect of soluble PD-L1 released by lung cancer cells in
regulating the function of T lymphocytes. Zhonghua Zhong Liu Za
Zhi. 35:85–88. 2013.(In Chinese). PubMed/NCBI
|
|
25
|
Akbay EA, Koyama S, Carretero J, Altabef
A, Tchaicha JH, Christensen CL, Mikse OR, Cherniack AD, Beauchamp
EM, Pugh TJ, et al: Activation of the PD-1 pathway contributes to
immune escape in EGFR-driven lung tumors. Cancer Discov.
3:1355–1363. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mahoney KM, Freeman GJ and McDermott DF:
The next immune-checkpoint inhibitors: PD-1/PD-L1 blockade in
melanoma. Clin Ther. 37:764–782. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mandai M, Hamanishi J, Abiko K, Matsumura
N, Baba T and Konishi I: Anti-PD-L1/PD-1 immune therapies in
ovarian cancer: Basic mechanism and future clinical application.
Int J Clin Oncol. 21:456–461. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bardoli AD, Afshar M, Viney R, Foster M,
Porfiri E, Zarkar A, Stevenson R, James ND, Bryan RT and Patel P:
The PD-1/PD-L1 axis in the pathogenesis of urothelial bladder
cancer and evaluating its potential as a therapeutic target. Future
Oncol. 12:595–600. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Brower V: Anti-PD-L1 antibody active in
metastatic bladder cancer. Lancet Oncol. 16:e112015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vassilakopoulou M, Avgeris M, Velcheti V,
Kotoula V, Rampias T, Chatzopoulos K, Perisanidis C, Kontos CK,
Giotakis AI, Scorilas A, et al: Evaluation of PD-L1 expression and
associated tumor-infiltrating lymphocytes in laryngeal squamous
cell carcinoma. Clin Cancer Res. 22:704–713. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yao S and Chen L: PD-1 as an immune
modulatory receptor. Cancer J. 20:262–264. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang BY, Zhan YP, Zong WJ, Yu CJ, Li JF,
Qu YM and Han S: The PD-1/B7-H1 pathway modulates the natural
killer cells versus mouse glioma stem cells. PLoS One.
10:e01347152015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Joo HY, Yun M, Jeong J, Park ER, Shin HJ,
Woo SR, Jung JK, Kim YM, Park JJ, Kim J and Lee KH: SIRT1
deacetylates and stabilizes hypoxia-inducible factor-1α (HIF-1α)
via direct interactions during hypoxia. Biochem Biophys Res Commun.
462:294–300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Doi T, Ishikawa T, Okayama T, Oka K,
Mizushima K, Yasuda T, Sakamoto N, Katada K, Kamada K, Uchiyama K,
et al: The JAK/STAT pathway is involved in the upregulation of
PD-L1 expression in pancreatic cancer cell lines. Oncol Rep.
37:1545–1554. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bellucci R, Martin A, Bommarito D, Wang K,
Hansen SH, Freeman GJ and Ritz J: Interferon-γ-induced activation
of JAK1 and JAK2 suppresses tumor cell susceptibility to NK cells
through upregulation of PD-L1 expression. Oncoimmunology.
4:e10088242015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Atefi M, Avramis E, Lassen A, Wong DJ,
Robert L, Foulad D, Cerniglia M, Titz B, Chodon T, Graeber TG, et
al: Effects of MAPK and PI3K pathways on PD-L1 expression in
melanoma. Clin Cancer Res. 20:3446–3457. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang L, Huang F, Mei J, Wang X, Zhang Q,
Wang H, Xi M and You Z: Posttranscriptional control of PD-L1
expression by 17β-estradiol via PI3K/Akt signaling pathway in
ERα-positive cancer cell lines. Int J Gynecol Cancer. 27:196–205.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang X, Zhou J, Giobbie-Hurder A, Wargo J
and Hodi FS: The activation of MAPK in melanoma cells resistant to
BRAF inhibition promotes PD-L1 expression that is reversible by MEK
and PI3K inhibition. Clin Cancer Res. 19:598–609. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gowrishankar K, Gunatilake D, Gallagher
SJ, Tiffen J, Rizos H and Hersey P: Inducible but not constitutive
expression of PD-L1 in human melanoma cells is dependent on
activation of NF-κB. PLoS One. 10:e01234102015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hernandez RK, Cetin K, Pirolli M, Quigley
J, Quach D, Smith P, Stryker S and Liede A: Estimating high-risk
castration resistant prostate cancer (CRPC) using electronic health
records. Can J Urol. 22:7858–7864. 2015.PubMed/NCBI
|
|
41
|
Kamoto T: Evaluation and diagnosis for
castration resistant prostate cancer: CRPC. Nihon Rinsho.
72:2103–2107. 2014.(In Japanese). PubMed/NCBI
|
|
42
|
Walsh JC, Lebedev A, Aten E, Madsen K,
Marciano L and Kolb HC: The clinical importance of assessing tumor
hypoxia: Relationship of tumor hypoxia to prognosis and therapeutic
opportunities. Antioxid Redox Signal. 21:1516–1554. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jensen LD: The circadian clock and hypoxia
in tumor cell de-differentiation and metastasis. Biochim Biophys
Acta. 1850:1633–1641. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lanier LL: NKG2D receptor and its ligands
in host defense. Cancer Immunol Res. 3:575–582. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Swaika A, Hammond WA and Joseph RW:
Current state of anti-PD-L1 and anti-PD-1 agents in cancer therapy.
Mol Immunol. 67:4–17. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ohaegbulam KC, Assal A, Lazar-Molnar E,
Yao Y and Zang X: Human cancer immunotherapy with antibodies to the
PD-1 and PD-L1 pathway. Trends Mol Med. 21:24–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cai B, Cai JP, Luo YL, Chen C and Zhang S:
The specific roles of JAK/STAT signaling pathway in sepsis.
Inflammation. 38:1599–1608. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Teng Y, Ross JL and Cowell JK: The
involvement of JAK-STAT3 in cell motility, invasion, and
metastasis. JAKSTAT. 3:e280862014.PubMed/NCBI
|
|
49
|
O'Shea JJ, Holland SM and Staudt LM: JAKs
and STATs in immunity, immunodeficiency, and cancer. N Engl J Med.
368:161–170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang SW and Sun YM: The IL-6/JAK/STAT3
pathway: Potential therapeutic strategies in treating colorectal
cancer (Review). Int J Oncol. 44:1032–1040. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu RY, Zeng Y, Lei Z, Wang L, Yang H, Liu
Z, Zhao J and Zhang HT: JAK/STAT3 signaling is required for
TGF-β-induced epithelial-mesenchymal transition in lung cancer
cells. Int J Oncol. 44:1643–1651. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lapeire L, Hendrix A, Lambein K, Van
Bockstal M, Braems G, Van Den Broecke R, Limame R, Mestdagh P,
Vandesompele J, Vanhove C, et al: Cancer-associated adipose tissue
promotes breast cancer progression by paracrine oncostatin M and
Jak/STAT3 signaling. Cancer Res. 74:6806–6819. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Duzagac F, Inan S, Simsek Ela F, Acikgoz
E, Guven U, Khan SA, Rouhrazi H, Oltulu F, Aktug H, Erol A and
Oktem G: JAK/STAT pathway interacts with intercellular cell
adhesion molecule (ICAM) and vascular cell adhesion molecule (VCAM)
while prostate cancer stem cells form tumor spheroids. J BUON.
20:1250–1257. 2015.PubMed/NCBI
|
|
54
|
Kim JM and Chen DS: Immune escape to
PD-L1/PD-1 blockade: Seven steps to success (or failure). Ann
Oncol. 27:1492–1504. 2016. View Article : Google Scholar : PubMed/NCBI
|