Introduction
It has been studied that instances of instinctive
behavior and emotion are controlled by smelling in many animals
(1,2). The olfactory bulb is the first relay
of the olfactory system, and its synaptic, phylogenetic and
cortical structures are well conserved compared with other cortical
structures of the brain (3,4). The
olfactory bulb is one of the special substructure as seen in the
brain, because it connects to the limbic system such as the
hippocampus and amygdala (5,6).
Mitral cells in the olfactory bulb play a key role in the olfactory
system, because they receive olfactory information from olfactory
sensory neurons, and axons of the mitral cells transfer the
information to the olfactory cortex (7–9).
The case of aging brings physiological changes in
the central nervous system (CNS) (10,11).
Studies show that alterations have been also discovered in aged
olfactory bub (12,13). In particular, it has been reported
that growth factors are changed in the brain during normal aging
(14,15). For example, insulin-like growth
factor-I (IGF-I) is well known to be a multifunctional protein in
the CNS, because in that case it acts as a controller of brain
development and neural plasticity (16). In addition, IGF-I promotes neuronal
survival and protection against neuronal damage induced by brain
damage, including ischemia-reperfusion injury (17). Furthermore, we have noted that some
studies have shown that exogenous IGF-I promotes neurogenesis in
the case of an aged brain (18,19).
To the best of our knowledge, however, in this case
we have noted that there are few reports on age-induced alterations
of IGF-I and its receptor (IGF-IR) in the olfactory bulb.
Therefore, in the present study, we investigated expression
patterns of IGF-I and IGF-IR at postnatal month (PM) 3 as a young
group, PM 6 as an adult group, and PM 24 as an aged group, in the
olfactory bulb of the gerbil, which shows physiologically inherent
attributes in auditory processes, reproductive and nervous systems
(12,20,21).
Materials and methods
Experimental animals
Gerbils were obtained from the Experimental Animal
Center, Kangwon National University, Chuncheon, Republic of Korea.
The gerbils were divided into 3 groups (n=14, in each group): i)
Young group at PM 3 group; ii) adult group at PM 12; and iii) aged
group at PM 24. The gerbils were housed in standard conditions
under suitable temperature (23°C) and humidity (60%), control with
a 12 h light and dark cycle per day and provided with freely
accessible feed and water. Experimental protocol was approved
(approval no. KW-160802-2) by Institutional Animal Care and Use
Committee (IACUC) at Kangwon National University and adhered to
guidelines that are in compliance with the current international
laws and policies (Guide for the Care and Use of Laboratory
Animals, The National Academies Press, 8th edition, 2011).
Western blotting
In order to examine alterations of IGF-I and IGF-IR
levels in the olfactory bulb, western blot analyses were carried
out as we previously described (12). In short, 21 gerbils (7 gerbils in
each group) were sacrificed, and their brains were removed. Their
olfactory bulbs were dissected with a surgical blade and
homogenized in 50 mM PBS (pH 7.4) containing EGTA (pH 8.0), 0.2%
NP-40, 10 mM EDTA (pH 8.0), 15 mM sodium pyrophosphate, 100 mM
β-glycerophosphate, 50 mM NaF, 150 mM NaCl, 2 mM sodium
orthovanadate, 1 mM PMSF and 1 mM DTT. After centrifugation, each
protein level in the supernatant was determined using a Micro BCA
protein assay kit with bovine serum albumin as a standard (Pierce;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Aliquots
containing 20 µg of total protein were boiled in loading buffer
containing 150 mM Tris (pH 6.8), 3 mM DTT, 6% SDS, 0.3% bromophenol
blue and 30% glycerol. The aliquots were then loaded onto a 10%
polyacrylamide gel. After electrophoresis, the gel was transferred
to nitrocellulose transfer membrane (Pall Crop, East Hills, NY,
USA). Rabbit anti-IGF-I (1:1,000, cat. no. sc-9013; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), rabbit anti-IGF-IR (1:1,000,
cat. no. sc-712; Santa Cruz Biotechnology, Inc.) or mouse
anti-β-actin (1:5,000, cat. no. A5441; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany,) were used as primary antibodies. The primary
antibodies were incubated overnight at 4°C. Peroxidase-conjugated
donkey anti-rabbit IgG (1:1,000, cat. no. sc-2305; Santa Cruz
Biotechnology, Inc.) or goat anti-mouse IgG (1:1,000, cat. no.
sc-2031; Santa Cruz Biotechnology, Inc.) were used as secondary
antibodies (2 h at room temperature). Antibody binding was detected
with an enhanced luminol-based chemiluminescent (ECL) kit (cat. no.
32106; Pierce; Thermo Fisher Scientific, Inc.).
Tissue preparation for histology
For histochemical and immunohistochemical analyses,
21 gerbils (7 gerbils in each group) were anesthetized with
pentobarbital sodium (30 mg/kg, i.p.; JW Pharmaceutical Co., Ltd.,
Seoul, Korea) and perfused transcardially with 0.1 M
phosphate-buffered saline (PBS, pH 7.4) followed by 4%
paraformaldehyde in 0.1 M phosphate-buffer (PB, pH 7.4). Their
olfactory bulbs were removed and post-fixed in the same fixative
for 6 h, infiltrated by 30% sucrose solution (in 0.1 M PB, pH 7.4)
for cryoprotection, and serially sectioned into 30 µm thickness in
a cryostat (Leica Microsystems GmbH, Wetzlar, Germany).
Cresyl violet (CV) histochemistry
To examine change of cellular morphology in the
olfactory bulb with age, CV histochemistry was carried out
according to our published protocol (22). In short, the sections were mounted
on gelatin-coated microscopy slides. CV acetate (Sigma-Aldrich;
Merck KGaA) was dissolved at 1.0% (w/v) in distilled water, and
glacial acetic acid (Sigma-Aldrich; Merck KGaA) was added to this
solution. The sections were stained by CV solution and dehydrated.
Finally, the stained sections were mounted Canada balsam (Kanto,
Tokyo, Japan).
Immunohistochemistry for IGF-I and
IGF-IR
To compare neuronal distribution and change in the
olfactory bulb between the 3 groups, IGF-I and IGF-IR
immunohistochemistry was performed according to our published
method (23). Briefly, the
sections were incubated with diluted rabbit anti-IGF-I (1:200, cat.
no. sc-9013; Santa Cruz Biotechnology, Inc.) and rabbit anti-IGF-IR
(1:200, cat. no. sc-712; Santa Cruz Biotechnology, Inc.) and
exposed to biotinylated goat anti-rabbit IgG and avidin-biotin
complex subsequently (1:200; Vector Laboratories, Burlingame, CA,
USA). Finally, they were visualized by staining with
3,3′-diaminobenzidine (Sigma-Aldrich, Merck KGaA).
Data analysis
Western blot analysis was done according to our
published method (12). In short,
the bands were scanned, and the quantification of the western
blotting was done using Scion Image software (Scion Corp.,
Frederick, MD, USA), which was used to count relative optical
density (ROD). The ROD was presented in graphs as percentage of the
young group.
To analyze immunoreactivity of IGF-I and IGF-IR,
respectively, 7 sections per animal in each group were selected and
analyzed according to our published method (24). In brief, digital images were taken
through a light microscope (BX53, Olympus, Germany) equipped with
digital camera (DP72, Olympus) connected to a PC monitor. The
images were calibrated into an array of 512 × 512 pixels
corresponding to a tissue area of 140 × 140 µm (×40 primary
magnification). Each immunoreactivity was measured by a 0–255 gray
scale system, and the background density was subtracted. A ratio of
the relative immunoreactivity (RI) for each antibody was calibrated
as % using Adobe Photoshop version 8.0 and then analyzed using NIH
Image 1.59 software. A ratio of the RI was calibrated as percentage
of the young group.
Statistical analysis
Data were expressed as the mean ± standard error of
mean (SEM). The data were elevated by one-way analysis of variance
(ANOVA) with a post hoc Bonferroni's multiple comparison test to
express differences among the 3 groups. Data was analysed using
SPSS software (version 12.0; SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
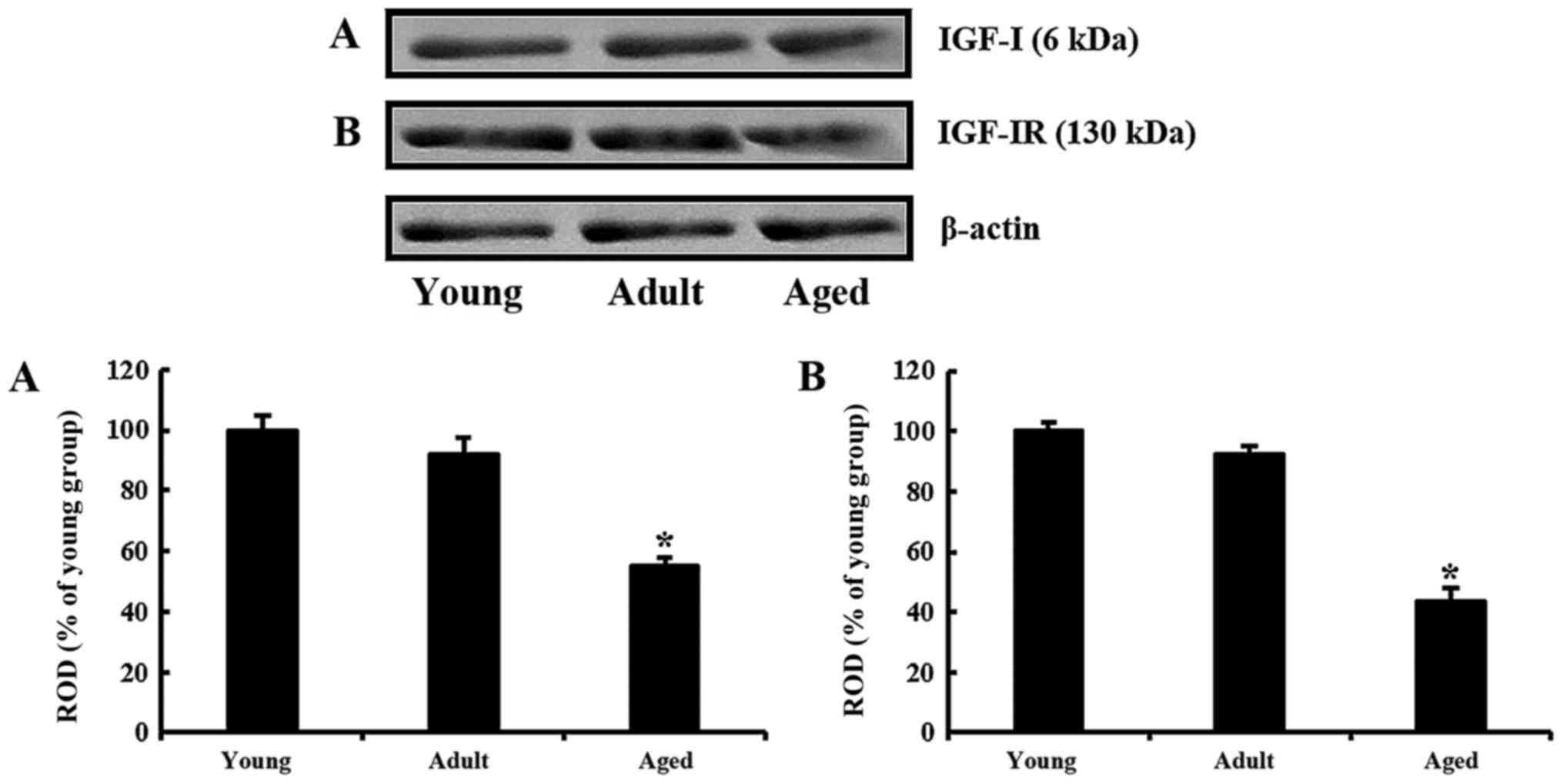
IGF-I and IGF-IR levels
Age-dependent changes in IGF-I and IGF-IR levels in
the olfactory bulb were identified by western blot analyses
(Fig. 1). IGF-I level in the adult
group was decreased to approximately 91% of the young group and
very low (approximately 54% of the young group) in the aged group
(Fig. 1A). Similarly, as the
animals were getting older, IGF-IR levels were continuously
decreased. The level in the adult was approximately 92% of the
young group and approximately 43% of the young group in the aged
(Fig. 1B).
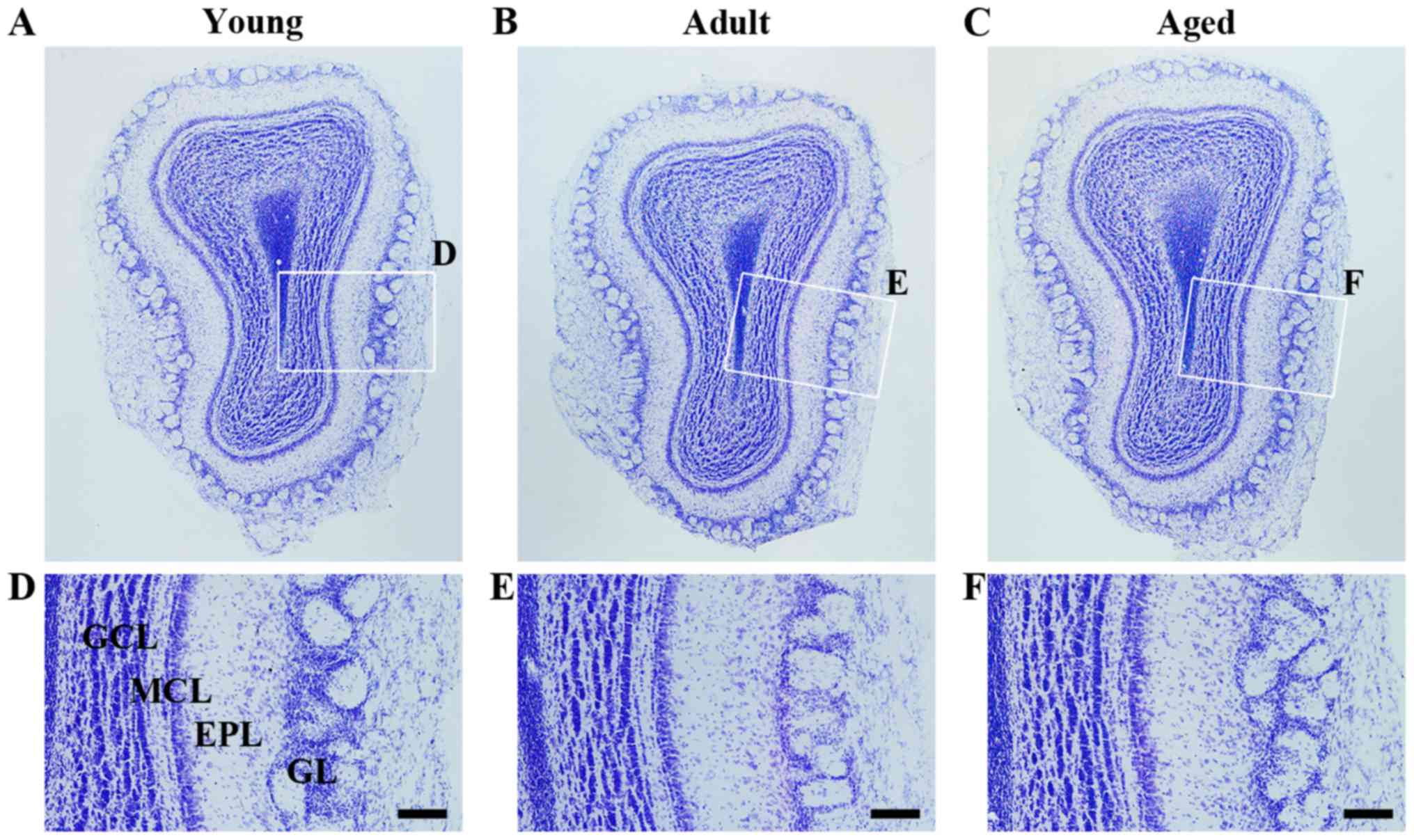
CV positive cells
To investigate alteration of cellular morphology in
the olfactory bulb, CV staining was performed in each group.
Regardless of age, CV positive cells in the olfactory bulb were
clearly observed. As the animals were getting older, significant
change in cellular morphology was not found in the olfactory bulb
(Fig. 2A-C). In addition, among
the three groups, significant differences in cellular distribution
were not found (Fig. 2D-F).
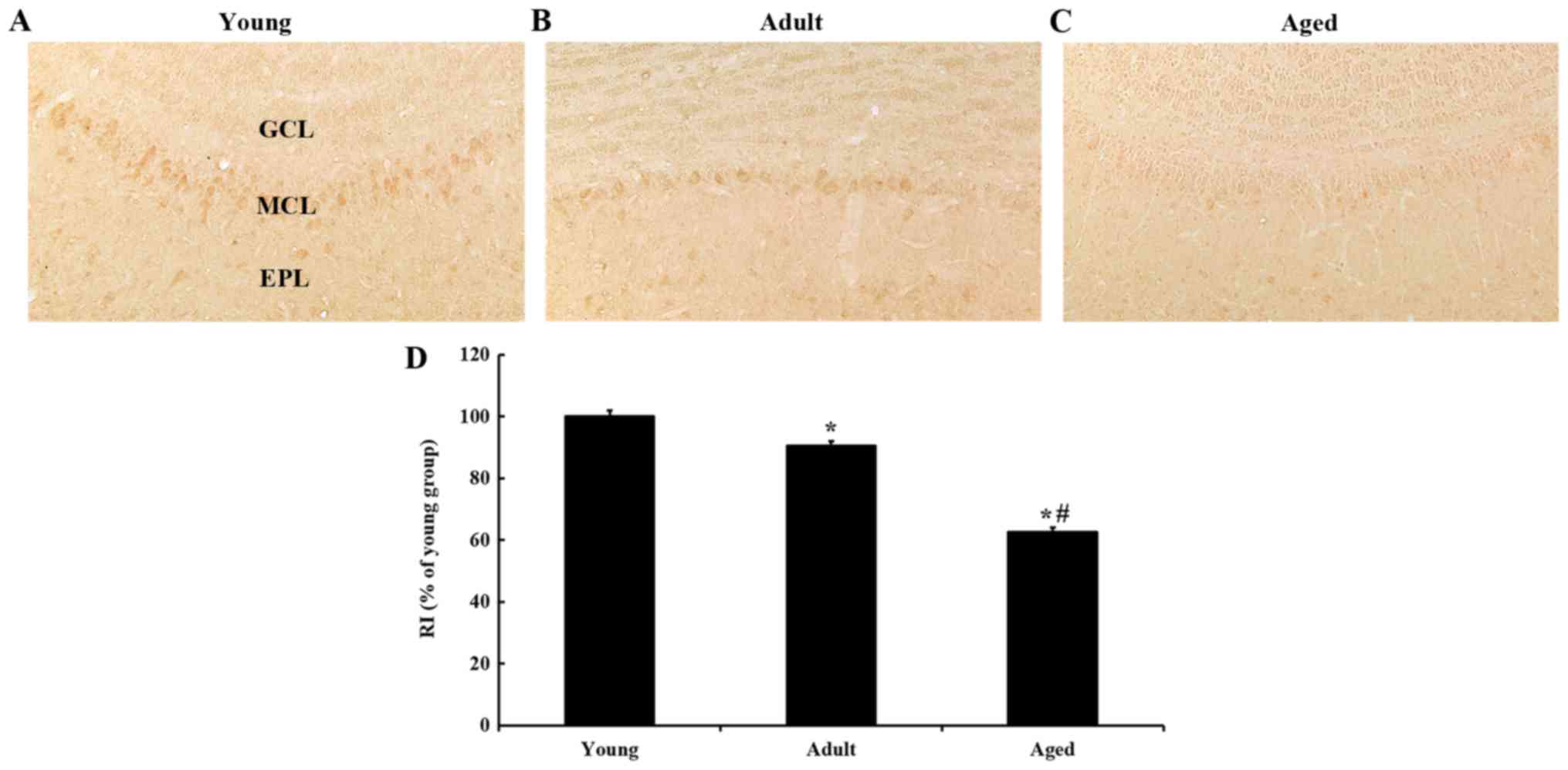
IGF-I immunoreactivity
To examine changes in IGF-I immunoreactivity in the
olfactory bulb, immunohistochemistry for IGF-I was carried out in
each age group (Fig. 3). In the
young group, IGF-I immunoreactivity was the strongest in cells of
the mitral cell layer (MCL) (Fig.
3A). As the animals were getting older, IGF-I immunoreactivity
in the MCL was continuously decreased. RI in the MCL in the adult
was approximately 90% of the young group (Fig. 3B and D), and approximately 62% of
the young group in the aged group (Fig. 3C and D).
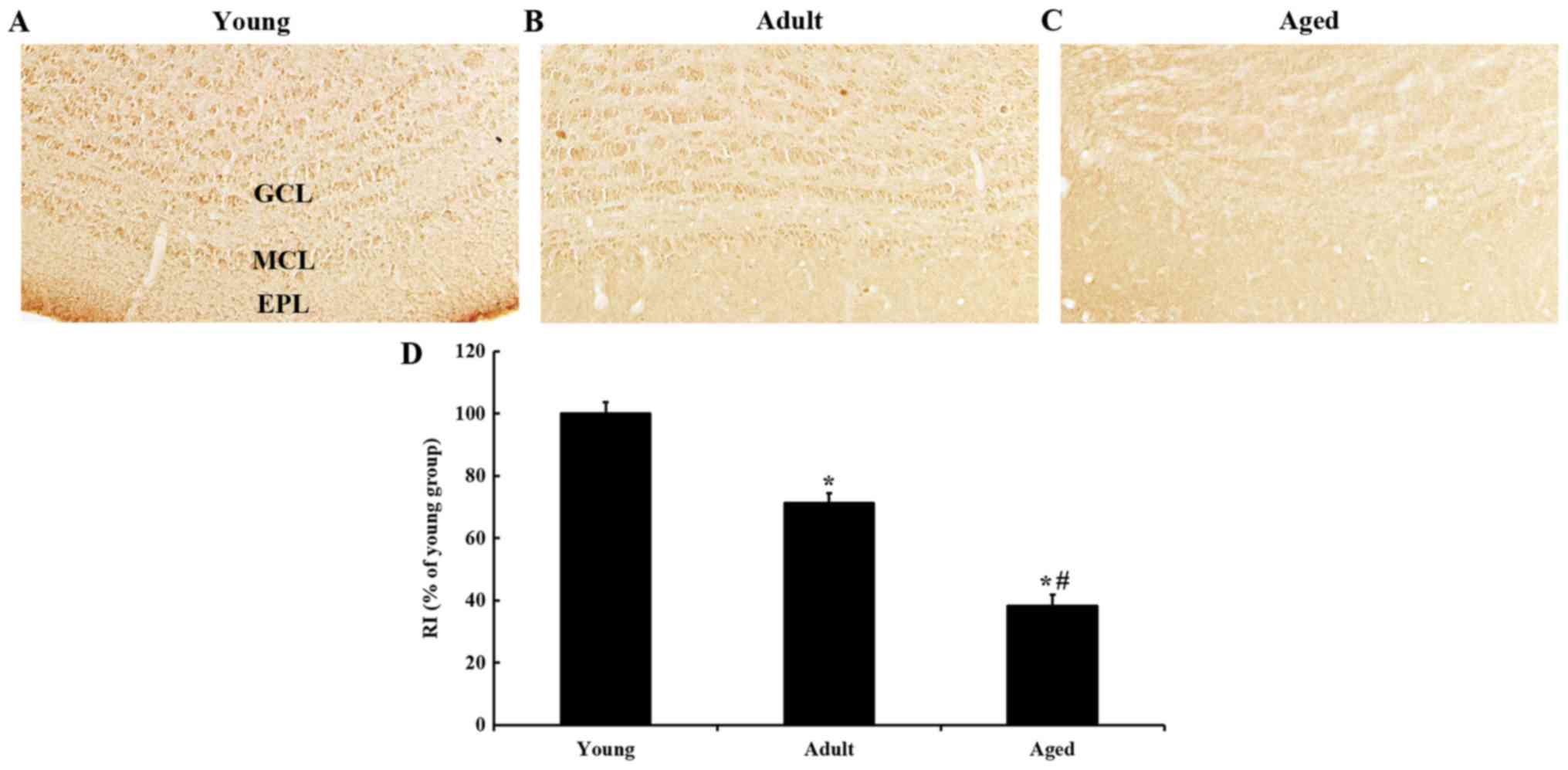
IGF-IR immunoreactivity
IGF-IR immunoreactivity in the young group was found
in all layers of the olfactory bulb, and the immunoreactivity was
strong (Fig. 4A). IGF-IR
immunoreactivity was decreased with age; the immunoreactivity in
the adult group was approximately 71% of the young group (Fig. 4B), and approximately 38% of the
young group in the aged group (Fig. 4C
and D).
Discussion
As a matter of course, aging gives rise to
morphological and physiological changes in the various regions of
the brain. In this study, we had focused on the characterization of
the mitral cells, which display important roles in the synaptic
circuit of the olfactory system (7). Axons of mitral cells transfer
olfactory information to diverse subregions in the brain including
the olfactory cortex (8,9). In this study, we found that cells in
the olfactory bulb were not significantly changed in microscopic
morphology during the incidence of normal aging. Up to date,
researchers have studied age-dependent alterations in aged
olfactory bulbs. For example, it is noted that a hindrance to
delivering olfactory information is induced by a selective
disruption of synaptic circuits (13).
It has been studying that neuroactive substances in
various regions of the brain are continuously altered during normal
aging. For example, dynamin 1, which is known as a regulator of
presynaptic endocytosis in the hippocampus, is known to be
decreased with age (25). In
addition, calcium binding proteins are differently changed
according to the proteins in the somatosensory cortex with age
(26). In this case, some studies
have reported that IGF-I and IGF-IR play key roles in the olfactory
system. For example, IGF-I and IGF-IR are involved in the
transmission of olfactory information and development of olfactory
axons (27,28). In this respect, in this study, we
had meticulously examined IGF-I and IGF-IR expressions in the
gerbil olfactory bulb during normal aging, and we found that IGF-I
and IGF-IR expressions were in deed strong in young olfactory
bulbs, and noted as significantly reduced in aged ones.
To the best of our knowledge, our literature review
and studies regarding alterations of IGF-I and IGF-IR in the
olfactory bulb with aging have been poorly produced. In this
regard, we report that IGF-I and IGF-IR were strongly expressed in
mitral cells of young gerbils, and they were significantly reduced
in the mitral cells of aged gerbils. It has been reported that
there are instances of IGF-I modulating synaptic formation and
secretions of neurotransmitters in the brain (29,30).
In the hippocampus, IGF-I plays an important role in synaptogenesis
in the dentate gyrus (31).
It is noted that some researchers have reported that
an age-induced decrease of IGF-I expression in the brain is related
to an identified cognitive impairment (32–35).
On the other hand, there is a split in age-dependent alteration of
IGF-IR expression in this case. Some papers have shown that the
expression of IGF-IR is decreased or increased or sustained in aged
hippocampi (35–38). We have recently reported that
expressions of IGF-I and IGF-IR are decreased in aged mouse
hippocampus and somatosensory cortex (33).
Although, in this study, we found that the
morphology and distribution pattern of olfactory cells was not
decidedly different regardless of age, IGF-I and IGF-IR were
strongly expressed in mitral cells of young gerbils, and the
expressions were significantly decreased in the mitral cells with
time. Therefore, based on our present and precedent researches, we
suggest that IGF-I and IGF-IR must be a key factor in olfactory
transmission through mitral cells. In this regard, noted subsequent
decreases of IGF-I and IGF-IR expressions with age may be related
with olfactory decline. Further studies must be needed to study
characteristic roles of IFG-I and IGF-IR in disorders of the
olfactory system.
Acknowledgements
Not applicable.
Funding
This study was supported by the Priority Research
Centers Program (grant no. NRF-2009-0093812) through the National
Research Foundation of Korea funded by the Ministry of Science, ICT
and Future Planning. It was also supported by the Bio-Synergy
Research Project (grant no. NRF-2015M3A9C4076322) of the Ministry
of Science, ICT and Future Planning through the National Research
Foundation.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SYC and MHW were responsible for the experimental
design, data, collection, data analysis and manuscript writing.
TKL, JCL, JHA, JHP and MCS performed the experiments. BHC, JHC,
HAL, JHC, IKH and IJK performed data analysis and critical comments
on the whole process of this study. All authors have read and
approved the final version of manuscript.
Ethics approval and consent to
participate
The experimental protocol was approved by
Institutional Animal Care and Use Committee at Kangwon National
University (approval no. KW-160802-2).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Saraiva LR, Kondoh K, Ye X, Yoon KH,
Hernandez M and Buck LB: Combinatorial effects of odorants on mouse
behavior. Proc Natl Acad Sci USA. 113:E3300–E3306. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stowers L, Cameron P and Keller JA:
Ominous odors: Olfactory control of instinctive fear and aggression
in mice. Curr Opin Neurobiol. 23:339–345. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Halász N and Shepherd GM: Neurochemistry
of the vertebrate olfactory bulb. Neuroscience. 10:579–619. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shepherd GM: Neurobiology. Modules for
molecules. Nature. 358:457–458. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kikusui T, Kajita M, Otsuka N, Hattori T,
Kumazawa K, Watarai A, Nagasawa M, Inutsuka A, Yamanaka A, Matsuo
N, et al: Sex differences in olfactory-induced neural activation of
the amygdala. Behav Brain Res. 346:96–104. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pena RR, Medeiros DC, Guarnieri LO, Guerra
JB, Carvalho VR, Mendes EMAM, Pereira GS and Moraes MFD: Home-cage
odors spatial cues elicit theta phase/gamma amplitude coupling
between olfactory bulb and dorsal hippocampus. Neuroscience.
363:97–106. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan Q, Mutoh H, Debarbieux F and Knöpfel
T: Calcium signaling in mitral cell dendrites of olfactory bulbs of
neonatal rats and mice during olfactory nerve Stimulation and
beta-adrenoceptor activation. Learn Mem. 11:406–411. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Igarashi KM, Ieki N, An M, Yamaguchi Y,
Nagayama S, Kobayakawa K, Kobayakawa R, Tanifuji M, Sakano H, Chen
WR and Mori K: Parallel mitral and tufted cell pathways route
distinct odor information to different targets in the olfactory
cortex. J Neurosci. 32:7970–7985. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nagayama S, Enerva A, Fletcher ML,
Masurkar AV, Igarashi KM, Mori K and Chen WR: Differential axonal
projection of mitral and tufted cells in the mouse main olfactory
system. Front Neural Circuits. 4:pii: 120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He WB, Zhang JL, Hu JF, Zhang Y, Machida T
and Chen NH: Effects of glucocorticoids on age-related impairments
of hippocampal structure and function in mice. Cell Mol Neurobiol.
28:277–291. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gallagher M, Bizon JL, Hoyt EC, Helm KA
and Lund PK: Effects of aging on the hippocampal formation in a
naturally occurring animal model of mild cognitive impairment. Exp
Gerontol. 38:71–77. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Choi JH, Yoo KY, Lee CH, Park OK, Yan BC,
Li H, Hwang IK, Park JH, Kim SK and Won MH: Comparison of newly
generated doublecortin-immunoreactive neuronal progenitors in the
main olfactory bulb among variously aged gerbils. Neurochem Res.
35:1599–1608. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Richard MB, Taylor SR and Greer CA:
Age-induced disruption of selective olfactory bulb synaptic
circuits. Proc Natl Acad Sci USA. 107:15613–15618. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fenn AM, Smith KM, Lovett-Racke AE,
Guerau-de-Arellano M, Whitacre CC and Godbout JP: Increased
micro-RNA 29b in the aged brain correlates with the reduction of
insulin-like growth factor-1 and fractalkine ligand. Neurobiol
Aging. 34:2748–2758. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Voss MW, Erickson KI, Prakash RS, Chaddock
L, Kim JS, Alves H, Szabo A, Phillips SM, Wójcicki TR, Mailey EL,
et al: Neurobiological markers of exercise-related brain plasticity
in older adults. Brain Behav Immun. 28:90–99. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dyer AH, Vahdatpour C, Sanfeliu A and
Tropea D: The role of insulin-like growth factor 1 (IGF-1) in brain
development, maturation and neuroplasticity. Neuroscience.
325:89–99. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Park JH, Park OK, Yan B, Ahn JH, Kim IH,
Lee JC, Kwon SH, Yoo KY, Lee CH, Hwang IK, et al: Neuroprotection
via maintenance or increase of antioxidants and neurotrophic
factors in ischemic gerbil hippocampus treated with tanshinone I.
Chin Med J (Engl). 127:3396–3405. 2014.PubMed/NCBI
|
|
18
|
Tang JJ, Podratz JL, Lange M, Scrable HJ,
Jang MH and Windebank AJ: Mechano growth factor, a splice variant
of IGF-1, promotes neurogenesis in the aging mouse brain. Mol
Brain. 10:232017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Morel GR, León ML, Uriarte M, Reggiani PC
and Goya RG: Therapeutic potential of IGF-I on hippocampal
neurogenesis and function during aging. Neurogenesis (Austin).
4:e12597092016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Campos SG, Zanetoni C, Scarano WR,
Vilamaior PS and Taboga SR: Age-related histopathological lesions
in the Mongolian gerbil ventral prostate as a good model for
studies of spontaneous hormone-related disorders. Int J Exp Pathol.
89:13–24. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Laumen G, Tollin DJ, Beutelmann R and
Klump GM: Aging effects on the binaural interaction component of
the auditory brainstem response in the Mongolian gerbil: Effects of
interaural time and level differences. Hear Res. 337:46–58. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Park JH, Joo HS, Yoo KY, Shin BN, Kim IH,
Lee CH, Choi JH, Byun K, Lee B, Lim SS, et al: Extract from
Terminalia chebula seeds protect against experimental ischemic
neuronal damage via maintaining SODs and BDNF levels. Neurochem
Res. 36:2043–2050. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Park JH, Lee TK, Yan BC, Shin BN, Ahn JH,
Kim IH, Cho JH, Lee JC, Hwang IK, Kim JD, et al: Pretreated glehnia
littoralis extract prevents neuronal death following transient
global cerebral ischemia through increases of superoxide dismutase
1 and brain-derived neurotrophic factor expressions in the gerbil
hippocampal cornu ammonis 1 area. Chin Med J (Engl). 130:1796–1803.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee TK, Park JH, Ahn JH, Shin MC, Cho JH,
Bae EJ, Kim YM, Won MH and Lee CH: Pretreated duloxetine protects
hippocampal CA1 pyramidal neurons from ischemia-reperfusion injury
through decreases of glial activation and oxidative stress. J
Neurol Sci. 370:229–236. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yoo DY, Jung HY, Kim JW, Yim HS, Kim DW,
Nam H, Suh JG, Choi JH, Won MH, Yoon YS and Hwang IK: Reduction of
dynamin 1 in the hippocampus of aged mice is associated with the
decline in hippocampaldependent memory. Mol Med Rep. 14:4755–4760.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bae EJ, Chen BH, Shin BN, Cho JH, Kim IH,
Park JH, Lee JC, Tae HJ, Choi SY, Kim JD, et al: Comparison of
immunoreactivities of calbindin-D28k, calretinin and parvalbumin in
the striatum between young, adult and aged mice, rats and gerbils.
Neurochem Res. 40:864–872. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Thorne RG, Pronk GJ, Padmanabhan V and
Frey WH II: Delivery of insulin-like growth factor-I to the rat
brain and spinal cord along olfactory and trigeminal pathways
following intranasal administration. Neuroscience. 127:481–496.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Matsui H, Noguchi T, Takakusaki K and
Kashiwayanagi M: Co-localization of TRPV2 and insulin-like growth
factor-I receptor in olfactory neurons in adult and fetal mouse.
Biol Pharm Bull. 37:1907–1912. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nilsson L, Sara VR and Nordberg A:
Insulin-like growth factor 1 stimulates the release of
acetylcholine from rat cortical slices. Neurosci Lett. 88:221–226.
1988. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ramsey MM, Adams MM, Ariwodola OJ, Sonntag
WE and Weiner JL: Functional characterization of des-IGF-1 action
at excitatory synapses in the CA1 region of rat hippocampus. J
Neurophysiol. 94:247–254. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
O'Kusky JR, Ye P and D'Ercole AJ:
Insulin-like growth factor-I promotes neurogenesis and
synaptogenesis in the hippocampal dentate gyrus during postnatal
development. J Neurosci. 20:8435–8442. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Arvat E, Broglio F and Ghigo E:
Insulin-Like growth factor I: Implications in aging. Drugs Aging.
16:29–40. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee CH, Ahn JH, Park JH, Yan BC, Kim IH,
Lee DH, Cho JH, Chen BH, Lee JC, Cho JH, et al: Decreased
insulin-like growth factor-I and its receptor expression in the
hippocampus and somatosensory cortex of the aged mouse. Neurochem
Res. 39:770–776. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Molina DP, Ariwodola OJ, Weiner JL,
Brunso-Bechtold JK and Adams MM: Growth hormone and insulin-like
growth factor-I alter hippocampal excitatory synaptic transmission
in young and old rats. Age (Dordr). 35:1575–1587. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sonntag WE, Lynch CD, Bennett SA, Khan AS,
Thornton PL, Cooney PT, Ingram RL, McShane T and Brunso-Bechtold
JK: Alterations in insulin-like growth factor-1 gene and protein
expression and type 1 insulin-like growth factor receptors in the
brains of ageing rats. Neuroscience. 88:269–279. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dore S, Kar S, Rowe W and Quirion R:
Distribution and levels of [125I]IGF-I, [125I]IGF-II and
[125I]insulin receptor binding sites in the hippocampus of aged
memory-unimpaired and -impaired rats. Neuroscience. 80:1033–1040.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Le Grevès M, Le Grevès P and Nyberg F:
Age-related effects of IGF-1 on the NMDA-, GH- and IGF-1-receptor
mRNA transcripts in the rat hippocampus. Brain Res Bull.
65:369–374. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Stenvers KL, Lund PK and Gallagher M:
Increased expression of type 1 insulin-like growth factor receptor
messenger RNA in rat hippocampal formation is associated with aging
and behavioral impairment. Neuroscience. 72:505–518. 1996.
View Article : Google Scholar : PubMed/NCBI
|