Introduction
Hypoxic ischemic encephalopathy (HIE) is the most
common brain injury caused by hypoxia and/or ischemia during the
perinatal period, which is a leading cause of perinatal mortality
in obstetrics and gynecology departments (1). HIE frequently leads to neurological
sequelae, including seizures, learning impairment, mental
retardation, epilepsy, visual impairment and cerebral palsy,
unconsciousness and muscle weakness (2,3).
Research has indicated that neuroprotection has a crucial role in
the progression of perinatal HIE in pediatric patients in
low-income countries (4).
Additionally, reports have suggested that apoptosis rate,
inflammatory cytokines and oxidative stress levels may regulate
neuroprotection in neurons during the progression of HIE. Evidence
has suggested that Tanshinone IIA (Tan-IIA) is a potential agent
for the treatment of cardiovascular and cerebrovascular diseases
(5). Therefore, the therapeutic
effects of Tan-IIA were investigated in a HIE mouse model.
Tan-IIA is a traditional Chinese medicine, extracted
from danshen that has been clinically used for treatment of various
human diseases (5,6). Research has indicated that Tan-IIA
administration may improve biochemical changes associated with
cardiac functions and increase fetal systolic pressure (7). In addition, Tan-IIA exhibits
therapeutic potential with protective effects on cardiovascular
functions (8). Furthermore,
Tan-IIA is reported to regulate mitogen-activated protein kinase
signaling pathway to enhance neuron regeneration (9). In this study, the beneficial effects
of Tan-IIA on apoptosis, inflammation and oxidative stress were
investigated in a mouse model of HIE. The potential neuroprotective
effects of Tan-IIA were primarily examined.
Currently, apoptosis of neurons has an essential
role in progression of HIE, with potential markers of
apoptosis-associated proteins are upregulated during neonatal HIE
injury (10). The role of
inflammatory cytokines in neuronal apoptosis of neonatal rats with
HIE has been reported previously and the results indicated that
inhibition of inflammatory cytokines, including tumor necrosis
factor-α (TNF-α) and interleukin-6 (IL-6), suppresses HIE-induced
neuronal apoptosis (11). In
addition, endoplasmic reticulum stress also was associated with the
activation of activating transcription factor-6 (ATF6) and
caspase-12 induced by apoptosis in neonatal HIE during the
perinatal period, and results indicated that apoptosis promotes HIE
brain injury, mediated by upregulation of endoplasmic reticulum
stress (12). Furthermore, Johnson
et al (13) demonstrated
that perinatal inflammation and infection caused by ischemia is
associated with correction of metabolic acidosis in HIE.
Furthermore, Chapados and Cheung (14) suggested that oxidative stress is
increased in rat pups following HIE and decreasing oxidative stress
is beneficial for improving HIE of the newborn rats. These reports
suggest that inflammation, oxidative stress and apoptosis are
associated with the progression of HIE during the perinatal
period.
In the current study, the regulatory effects of
Tan-IIA on the progression of HIE during perinatal period were
investigated. The inhibitory effects of Tan-IIA on inflammation,
oxidative stress and apoptosis was analyzed in a mouse model of
HIE. Body weight, brain water content and blood-brain barrier
permeability were also determined in newborn mice with HIE
following Tan-IIA treatment.
Materials and methods
Ethics statement
This preclinical investigation was conducted by
accordance with the recommendations in the Guide for the Care and
Use of Laboratory Animals of Renmin Hospital of Wuhan University
(Wuhan, China). This study was approved by the ethics committee of
Renmin Hospital of Wuhan University. All surgery and experiments
were performed under anesthetic.
Cell culture
Neurons were isolated from experimental mice and
cultured in Dulbecco's modified Eagle's medium with 10% fetal
bovine serum and incubated overnight at 37°C humidified atmosphere
of 5% CO2. Neurons were treated with 10 mg/ml Toll-like
receptor-4 (TLR-4) and/or Tan-IIA with PBS as control for 24 h for
further analysis in vitro.
Animal study
A HIE mouse model was conducted as previously
described (15). Postnatal day 10
C57BL/6J mice were purchased from Slac Shang Experimental Co., Ltd.
(Shanghai, China) and anesthetized to make a unilateral right
common carotid artery ligation by performing a 5-0 surgical silk
suture. Mice were then divided into two groups: One treated with
Tan-IIA (5 mg/kg) and the other treated with PBS daily for 15 days
by oral administration.
Counting of lymphocytes and
monocytes
The percentage of lymphocytes and monocytes was
calculated according to electrical impedance method (16) and laser scattering method by blood
cytometer (Vi-cell XR, Beckman Coulter, Inc., Brea, CA, USA). The
normal reference values of lymphocytes and monocytes in rats were
lymphocytes (62.76 to 90.19%): Mononuclear cells (0–3%).
ELISA
In the protein detection assay, mouse IL-1 (cat. no.
MLB00C; R&D Systems, Inc., Minneapolis, MN, USA), mouse IFN-γ
(cat. no. MIF00; R&D Systems, Inc., Minneapolis, MN, USA),
TNF-α (cat. no. P06804; R&D Systems, Inc.), C-X-C motif
chemokine 10 (CXCL10; cat. no. 15945; R&D Systems, Inc.) and
chemokine (C-C motif) ligand 12 (CCL12; cat. no. ABIN924766;
antibodies-online GmbH, Aachen, Germany) ELISA kits were used to
analyze serum levels of inflammatory factors in mice treated with
Tan-IIA or PBS. The procedures were conducted according to the
manufacturer's instructions. The final results were recorded at 450
nm using an ELISA plate reader.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from hippocampus using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Total RNA was used to produce cDNA by one step RT-PCR kit
(RR037A, Takara Biotechnology, Co., Inc., Tokyo, Japan) and the
protocol is 37°C for 15 min, 85°C for 5 sec The prepared cDNA was
used to analyze reactive oxygen species (ROS), superoxide dismutase
(SOD), catalase (CAT), glutathione (GSH), SRY-box 2 (Sox2) and
nerve growth factor (NGF) with β-actin as an endogenous control.
The amplified PCR products were quantified by measuring the
calculated quantitation cycle (Cq) of sample mRNA using
SYBR® qPCR: Premix Ex Taq™ II (TIi RNase H
Plus) kit and the thermocycling conditions were: Preheating 92°C (1
min 30 sec), followed by 28 cycles of 92°C (30 sec) and 65°C (1
min) (RR420A, Takara Biotechnology, Co., Inc.). All primers were
synthesized by Invitrogen (Thermo Fisher Scientific, Inc.; Table I). Relative changes in mRNA
expression were calculated by 2−ΔΔCq (17). The results are expressed as the
n-fold difference relative to β-actin.
 | Table I.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Target gene | Forward primer | Reverse primer |
|---|
| SOD |
5′-AATGTGTCCATTGAAGATCGTGTGA-3′ |
5′-GCTTCCAGCATTTCCAGTCTTTGTA-3′ |
| GSH |
5′-AATCCTGCTTGGGTATCAGG-3′ |
5′-GAGACCCAGTCTCAGGGAAA-3′ |
| CAT |
5′-GAGCCTCCTAGAAAGATCTAC-3′ |
5′-GCCAGCCTAGGGCTGAGCTG-3′ |
| Sox-2 |
5′-GGTCGAGGTAGTAGACCTTACA-3′ |
5′-GTTCGTCTCTGTGGTCAGATTC-3′ |
| NGF |
5′-TTTGTCTAACCCTAACTGAGAAGG-3′ |
5′-CTCTAGAATGAACGGTGGAAGG-3′ |
| β-actin |
5′-GTGGGCGCCCAGGCACCA-3′ |
5′-CTCCTTAATGTCACGCACGATTT-3′ |
Western blot analysis
Neurons were isolated from experimental mice and
homogenized in lysate buffer (cat. no. 3096; Bio-Rad Laboratories,
Inc., Hercules, CA, USA). The supernatant was acquired by
centrifugation at 13,400 × g for 25 min at 4°C; total protein was
quantified using a Bicinchoninic Acid Protein Assay kit (cat. no.
23228; Beyotime Institute of Biotechnology, Nanjing, China).
Samples (20 µg) were electrophoresed via 15% SDS-PAGE and
transferred to a polyvinylidene fluoride membrane (EMD Millipore,
Billerica, MA, USA).
Proteins of caspase-3 (cat. no. ab2172; Abcam,
Cambridge, UK), caspase-9 (cat. no. ab32539; Abcam), Bcl-2
apoptosis regulator (Bcl-2; cat. no. ab692; Abcam), P53 (cat. no.
ab61241; Abcam), TLR-4 (cat. no. ab22048; Abcam), nuclear factor-κB
(NF-κB; cat. no. ab28849; Abcam), ATF-6 (cat. no. ab11909; Abcam),
the transcription factor C/EBP homologous protein (CHOP; cat. no.
ab171894; Abcam) and apoptotic peptidase activating factor-1
(Apaf-1; cat. no. ab32372; Abcam) were detected. Transmembrane
proteins were extracted using Transmembrane Protein Extraction kit
(Qiagen Sciences, Inc., Gaithersburg, MD, USA) according to the
manufacturer's instructions. For western blotting, primary
antibodies were added after blocking (5% skimmed milk) and were
incubated for 1 h at 37°C and washed with PBS three times.
Subsequently, the nitrocellulose membrane was incubated with
secondary antibodies, for 24 h at 4°C and washed with PBS three
times. The results were visualized by using browser-based Digital
Darkroom software (Darkroom Software LLC, Plano, TX, USA) of
FluorChem R chemi-luminescence detection system (ProteinSimple, San
Jose, CA, USA).
Flow cytometry analysis
Apoptosis rates of neurons were evaluated using
Annexin V-fluorescein isothiocyanate (Annexin V-FITC) and propidium
iodide (PI) apoptosis detection kit (BD Biosciences, San Jose, CA,
USA). Neurons were collected and suspended with Annexin V-FITC and
PI according to the manufacturer's instructions. Fluorescence was
measured with a FACS scan flow cytometer (BD Biosciences).
Immunohistochemistry
Brains tissues were fixed in formalin fluid for 24 h
at room temperature and embedded in paraffin; 8 µm sections were
used for further analysis. The paraffin sections were incubated in
hydrogen peroxide (4.0%) for 10–15 min at 37°C and subsequently
rinsed three times with PBS, 5 min per wash. The sections were
blocked with blocking solution (pH 7.2–7.4, NaCl 137 mmol/l, KCl
2.7 mmol/l, Na2HPO4 10 mmol/l, KH2PO4 2 mmol/l) at room temperature
for 30 min and stained with 0.1% cresyl violet at 4°C for 12 h. All
sections were washed three times with PBS. Cerebral infarction area
was measured under a microscope, with Stereo Investigator software
(MBF Bioscience, Williston, VT, USA) as previously described
(18).
Detection of NF-κB activity
The activity of NF-κB in experimental mice rats was
detected according to the manufacturer' protocols using an NF-κB
Activation-Nuclear Translocation Assay kit (cat. no. SN368,
Beyotime Institute of Biotechnology).
Brain water content
The experimental mice were sacrificed under
isoflurane anesthesia at 15 days post-treatment with Tan-IIA. Brain
hemispheres were weighed prior to and following drying. Brain water
content (%) was calculated as (wet weight-dry weight)/wet weight
×100.
Evan's blue dye extravasation
assay
Evan's Blue dye extravasation was performed
according to a previously described report (19). Evan's Blue dye was administered by
intraperitoneal injection. Experimental mice were sacrificed to
measure blood-brain barrier permeability 24 h after administration.
The brain hemispheres were homogenized (50% tri-chloroacetic acid;
3:1) and analyzed at 620 nm by spectrophotometry.
ROS productions assay
Reactive Oxygen Species Assay Kit (University of
Rochester Medical Center, Rochester, NY, United States) was used to
evaluate the changes of ROS productions in neurons isolated from
experimental mice. Briefly, neurons were cultured in for 24 h at
37°C. Supernatants were removed and incubated with
dichlorofluorescin (10 µmol/l) for 20 min at 37°C. Cells were
washed with serum-free medium and were measured at 480 nm using an
ELISA plate reader.
Transfection of small interfering RNA
(Si-RNA)
All siRNAs were synthesized by Invitrogen (Thermo
Fisher Scientific, Inc.) including Si-RNA-TLR-4 (Si-TLR-4) or
Si-RNA-vector (Si-vector). A total of 120 pmol Si-TLR-4 (sense,
5′-UCCCCAAGUCAAUCUCUCUTT-3′ and
anti-sense-5′-AGAGAGAUUGACUUGGGGATT-3′) or Si-vector (sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and
anti-sense-5′-ACGUGACACGUUCGGAGAATT-3′) was delivered into the
neurons cells by electroporation. Standard electroporation was
performed using an Amaxa Cell line Nucleofector Kit L (Lonza Group,
Ltd., Basel, Switzerland) according to the manufacturer's
protocols. Briefly, 5×106 cells were suspended in 100 ìl
of nucleofector solution, Si-RNA-TLR-4 or negative control
(Si-RNA-vector) was added to the suspension, and then
electroporation was conducted at 140 V (250 ms/pulse, eight pulses;
10 µsec).
Fluoro-Jade C staining assay
Fluoro-Jade C staining was used to label
degenerating neurons using Fluoro-Jade C Ready-to-Dilute Staining
kit (cat. no. TR-100-FJ; Biosensis Pty Ltd., Thebarton, Australia)
to identify degenerating neurons. Neurons were isolated from
experimental mice and cultured in 6-well plates for 12 h at 37°C.
Cell culture medium was removed and labeled with 1 mg/ml DAPI for
15 min or 0.001% Fluoro-Jade C for 20 min according to
manufacturer's instructions. Fluoro-Jade C-labeled degenerating
neurons were visualized with blue light excitation while DAPI
counter stained cell nuclei were visualized with ultra-violet
illumination.
Short-term neurobehavioral
analysis
The geotactic reflex test was performed as
previously described (20).
Neurobehavioral analysis of mice was performed at baseline and the
72-h after treatment with Tan-IIA or PBS.
Statistical analysis
SPSS 12.0 statistical analysis software (SPSS Inc.,
Chicago, IL, USA) was used for statistical analysis. All data are
presented as the mean ± standard error from three or more
experimental repeats. Unpaired data were analyzed by Student's
t-test. Comparisons between multiple groups were analyzed by
one-way ANOVA. P<0.05 was considered to indicate a statistically
significant difference.
Results
Tan-IIA inhibits inflammatory
cytokines in peripheral blood of mice with perinatal HIE
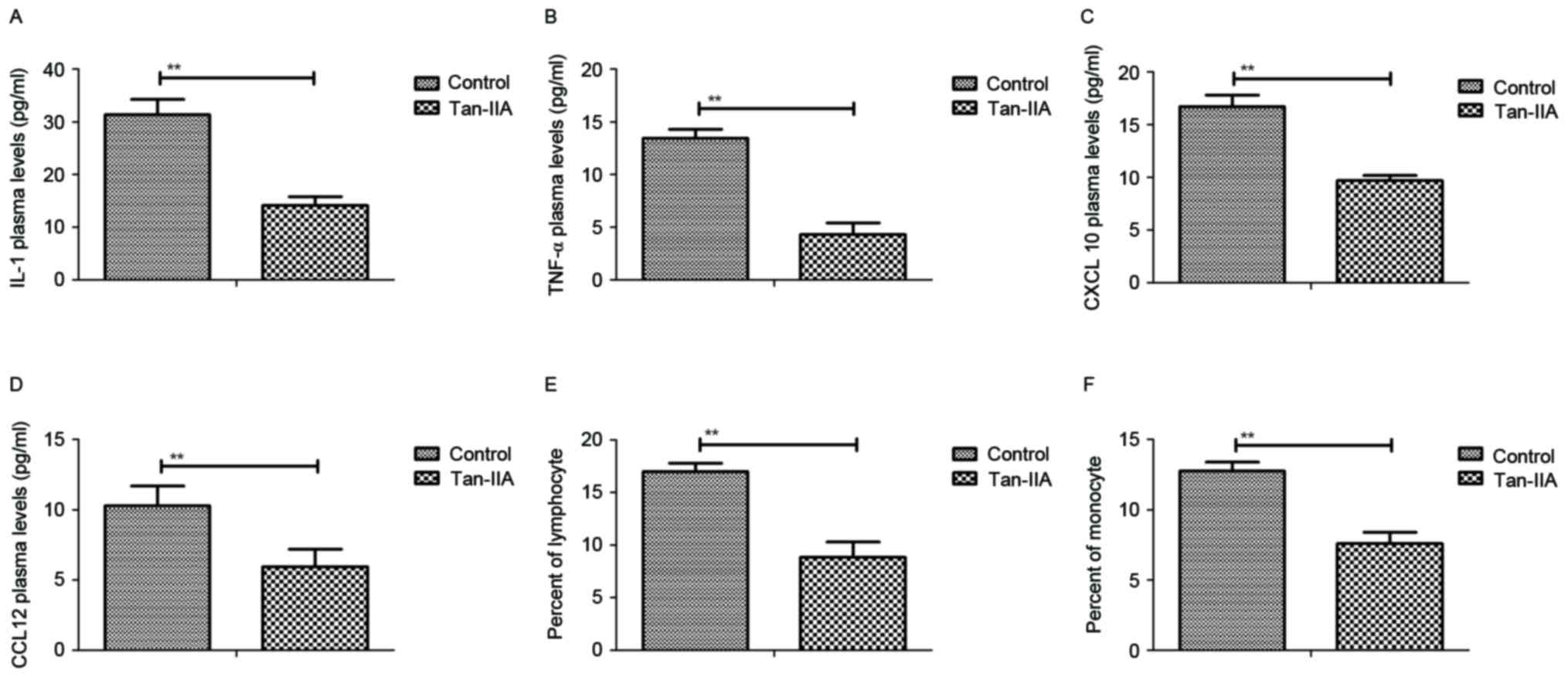
As presented in Fig.
1A, Tan-IIA treatment decreased production of inflammatory
cytokine IL-1 in the peripheral blood of mice with perinatal HIE.
Additionally, TNF-α plasma concentration levels were decreased by
Tan-IIA (Fig. 1B). Results also
demonstrated that serum levels of CXCL10 and CCL12 were
downregulated in mice with perinatal HIE following treatment with
Tan-IIA (Fig. 1C and D). Further,
counting with a blood cytometer revealed that the percentage of
lymphocytes and monocytes in peripheral blood were decreased by
Tan-IIA in mice with perinatal HIE compared with control treatment
(Fig. 1E and F). These data
indicate that Tan-IIA significantly reduces the level of
inflammatory cytokines in peripheral blood of mice with perinatal
HIE.
Tan-IIA decreases endoplasmic
reticulum stress and oxidative stress in mice with perinatal
HIE
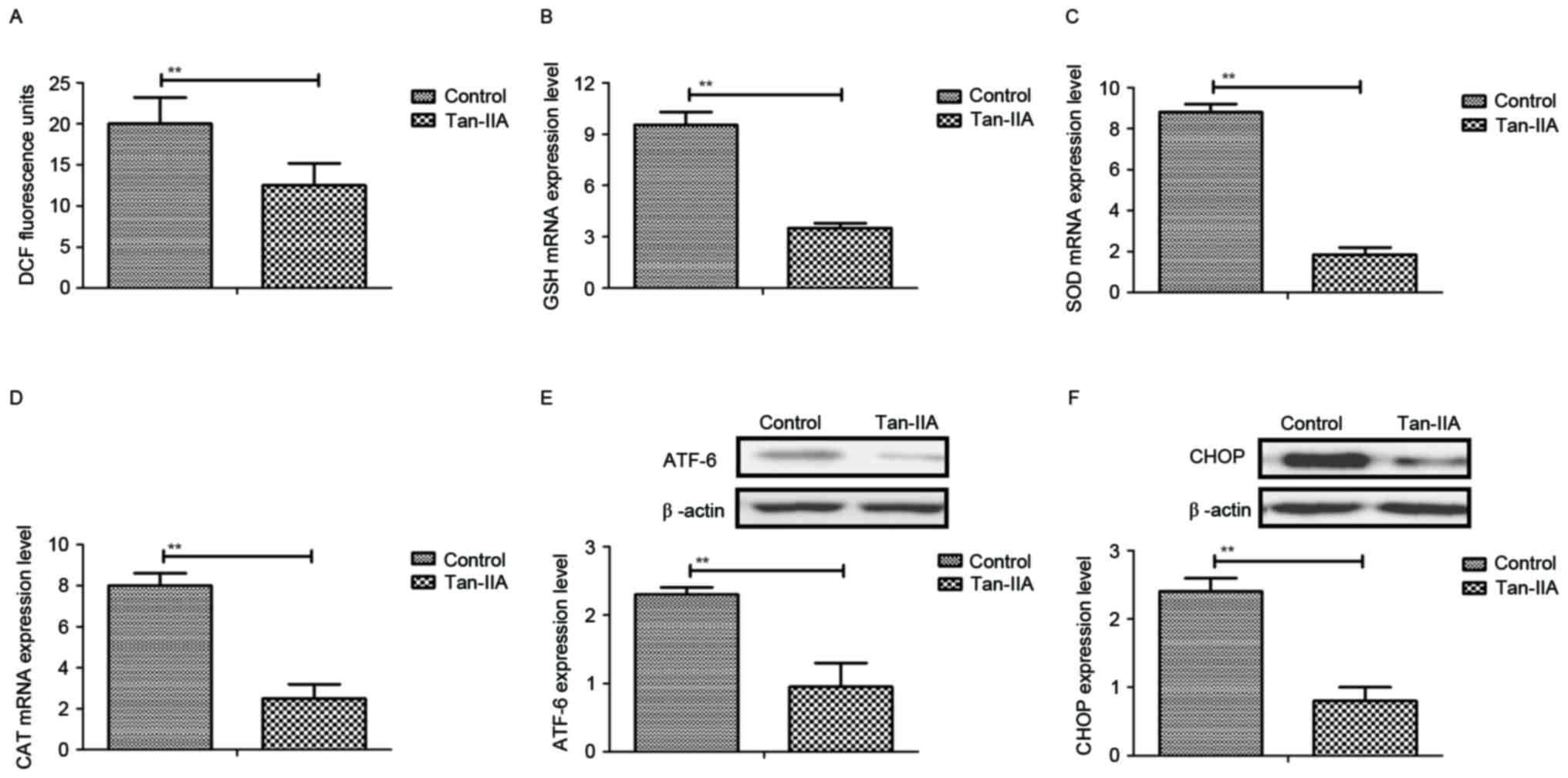
Following analysis of Tan-IIA-mediated inflammatory
cytokines, endoplasmic reticulum and oxidative stress were examined
in neurons from Tan-IIA-treated HIE mice. The results demonstrated
that ROS productions and GSH mRNA expression levels were
downregulated by Tan-IIA in serum in experimental mice (Fig. 2A and B). In addition, the mRNA
expression levels of SOD and CAT were also decreased by Tan-IIA in
the serum of experimental mice compared with the control group
(Fig. 2C and D). Furthermore,
protein expression levels of ATF-6 and CHOP were decreased in
neurons following treatment with Tan-IIA (Fig. 2E and F). These data indicate that
Tan-IIA treatment decreases endoplasmic reticulum stress and
oxidative stress in mice with perinatal HIE, which may contribute
to reduced apoptosis of neurons in the central nervous system
following HIE.
Tan-IIA suppresses apoptosis of
neurons in mice with perinatal HIE
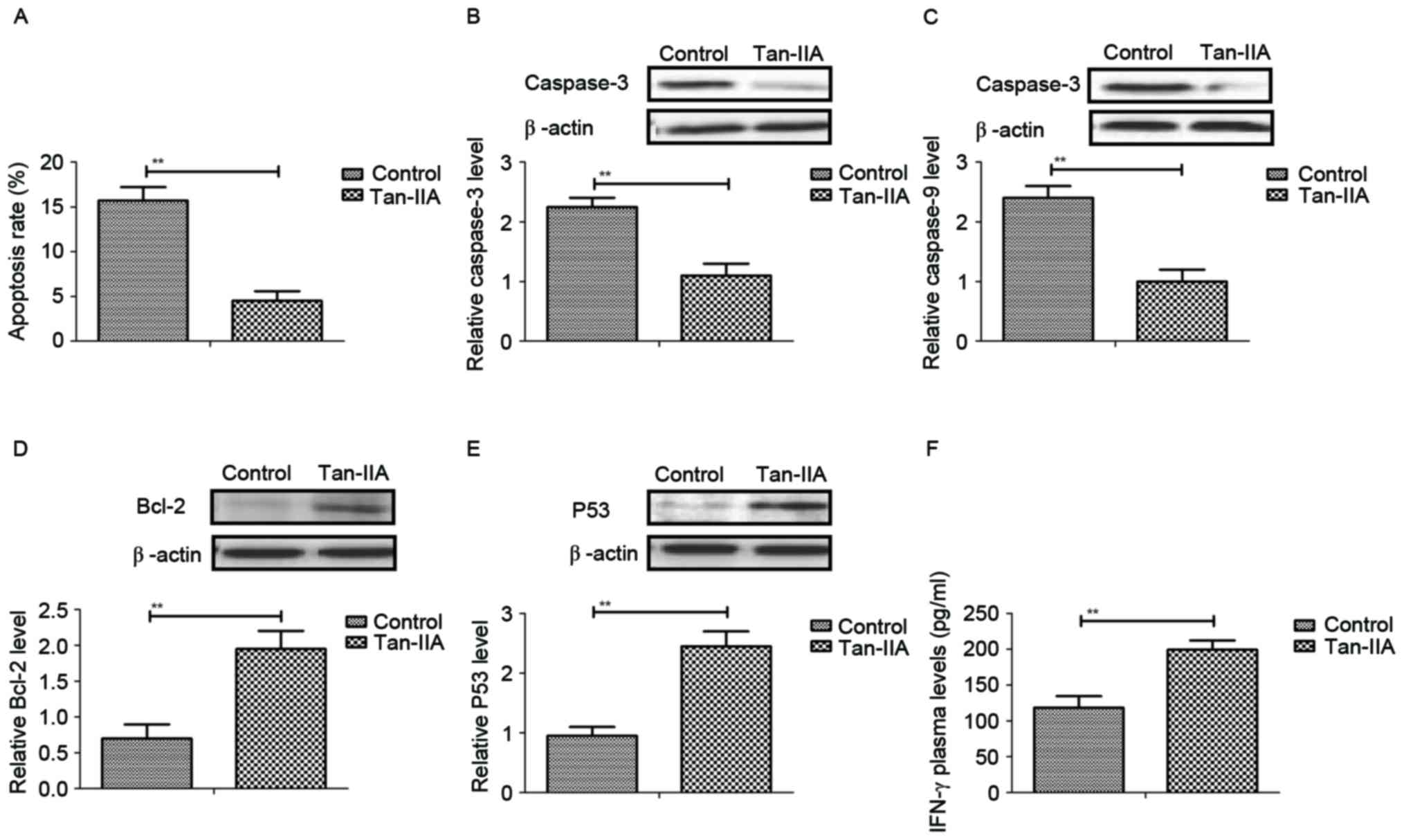
Subsequently, the anti-apoptosis effects of Tan-IIA
on neurons in mice model of HIE. As presented in Fig. 3A, the apoptosis rate of neurons was
decreased by 15 days of Tan-IIA treatment. The expression levels of
pro-apoptosis protein, caspase-3 and caspase-9, were suppressed by
Tan-IIA compared with control in neurons from HIE model mice
(Fig. 3B and C). However, the
expression levels of anti-apoptosis proteins, Bcl-2 were
upregulated by Tan-IIA treatment in neurons from HIE mice compared
with the control (Fig. 3D and E).
Notably, serum levels of IFN-γ anti-cytokine were upregulated by
Tan-IIA compared with the control group (Fig. 3F). The data demonstrate that
Tan-IIA can markedly inhibit apoptosis of neurons in mice with
perinatal HIE.
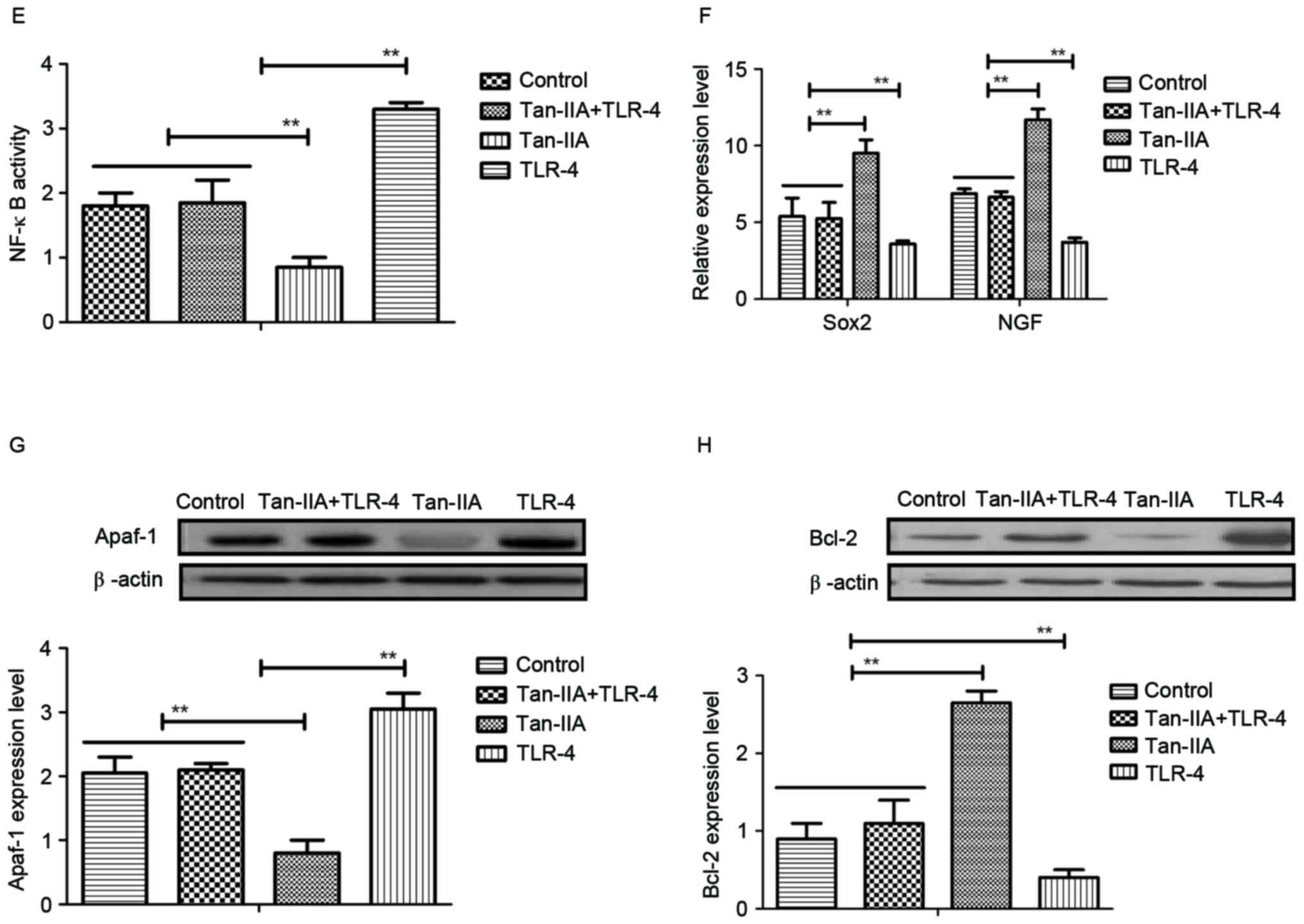
Tan-IIA regulates neuroactive factors
through inhibition of TLR-4-mediated NF-κB pathway
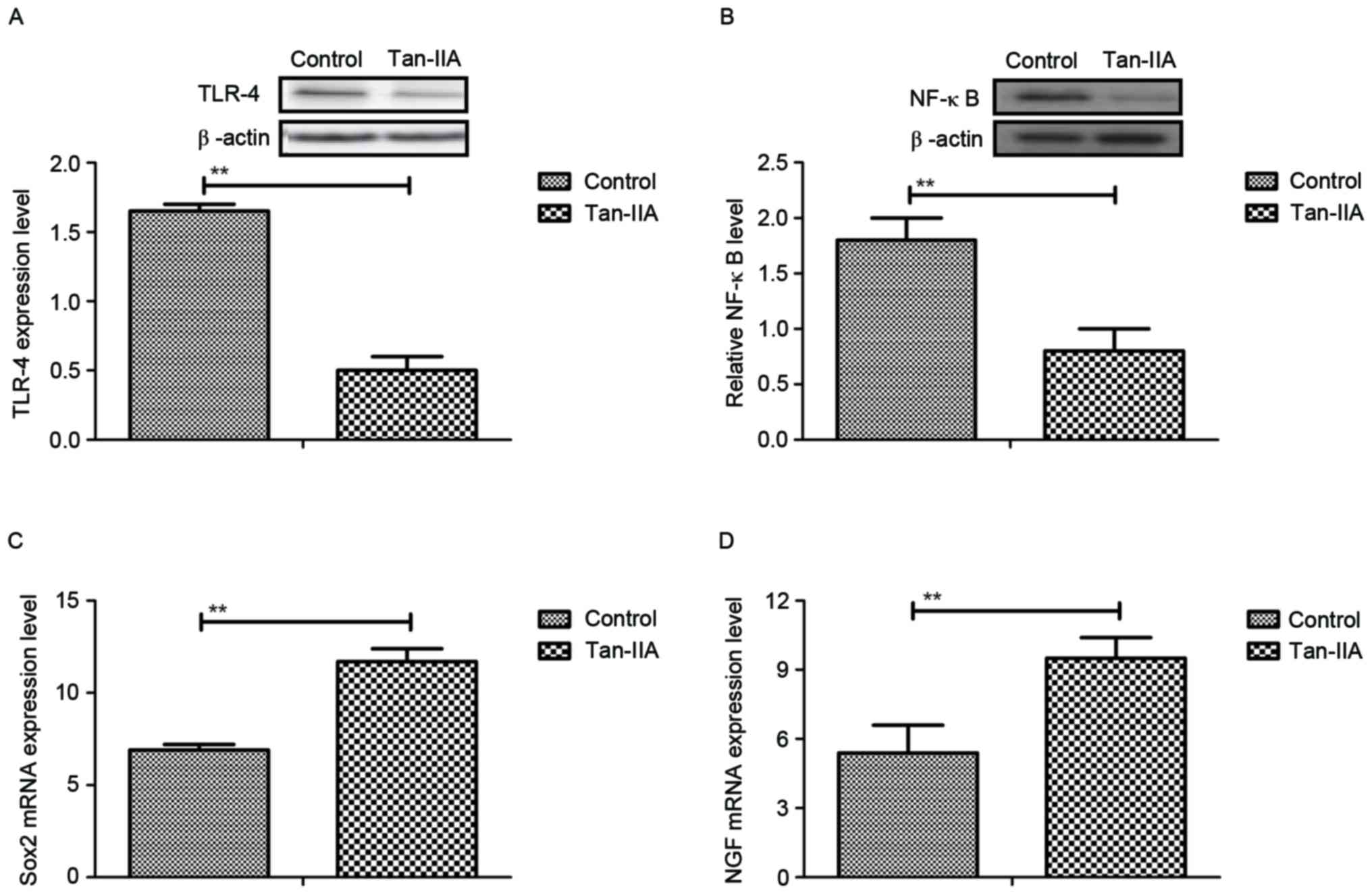
The molecular mechanisms of Tan-IIA-mediated
neuroprotective effects in neurons isolated from experimental mice
were investigated. TLR-4 expression levels were downregulated by
Tan-IIA treatment (Fig. 4A). NF-κB
expression was also decreased by Tan-IIA treatment in neurons
isolated from experimental mice compared with control (Fig. 4B). Tan-IIA treatment promoted the
mRNA expression of neuroactive factors Sox2 and NGF in neurons
(Fig. 4C and D). In vitro
experiments using cells used extracted from the HIE model control
demonstrated that TLR-4 addition abolished the inhibitory effects
of Tan-IIA on NF-κB activity (Fig.
4E). In addition, TLR-4 addition inhibited production of Sox2
and NGF promoted by Tan-IIA (Fig.
4F). Furthermore, TLR-4 addition upregulated Apaf-1 expression,
and downregulated Bcl-2 expression in neurons (Fig. 4G and H). These results suggest that
Tan-IIA regulates neuroactive factors and apoptosis of neurons
through inhibition of TLR-4-mediated pathways.
Knockdown of TLR-4 blocks
Tan-IIA-improved inflammation, stress and apoptosis in neurons in
HIE
The effects of TLR-4 knockdown were analyzed using
Si-TLR-4 to determine the effects on inflammation, stress and
apoptosis in Tan-IIA treated neurons following HIE. As presented in
Fig. 5A, IL-1 and TNF-α expression
levels were increased by TLR-4 knockdown in Tan-IIA-treated neurons
isolated from HIE mice. TLR-4 knockdown also abrogated expression
levels of CAT, SOD and GSH in neurons isolated from Tan-IIA-treated
HIE mice (Fig. 5B). The results
demonstrated that TLR-4 knockdown abolished Tan-IIA-inhibited
apoptosis of neurons isolated from Tan-IIA-treated HIE mice
(Fig. 5C). NF-κB, ATF-6 and CHOP
expression levels were upregulated by Si-TLR-4 neurons isolated
from Tan-IIA-treated HIE mice (Fig.
5D). TLR-4 knockdown increased ROS production in neurons
isolated from Tan-IIA-treated HIE mice (Fig. 5E). These results suggest that TLR-4
knockdown can block Tan-IIA-induced effects on inflammation, stress
and apoptosis in neurons from HIE model mice.
 | Figure 5.TLR-4 knockdown blocks
Tan-IIA-improved inflammation, stress and apoptosis in neurons
following HIE. (A) TLR-4 knockdown increased IL-1 and TNF-α
expression levels in neurons isolated from Tan-IIA-treated HIE
mice. (B) TLR-4 knockdown increases expression levels of CAT, SOD
and GSH in neurons isolated from HIE mice. (C) TLR-4 knockdown
upregulates apoptosis of neurons isolated from HIE mice. (D) TLR-4
knockdown upregulates NF-κB expression in neurons in isolated from
HIE mice. (E) TLR-4 knockdown increases ROS production in neurons
isolated from HIE mice. **P<0.05. HIE, hypoxic ischemic
encephalopathy; IL-1, interleukin-1; ROS, reactive oxygen species;
TNF-α, tumor necrosis factor-α; Si, small interfering RNA; TLR-4,
Toll-like receptor-4; CAT, catalase; SOD, superoxide dismutase;
GSH, glutathione; ATF-6, activating transcription factor-6; CHOP,
C/EBP homologous protein. |
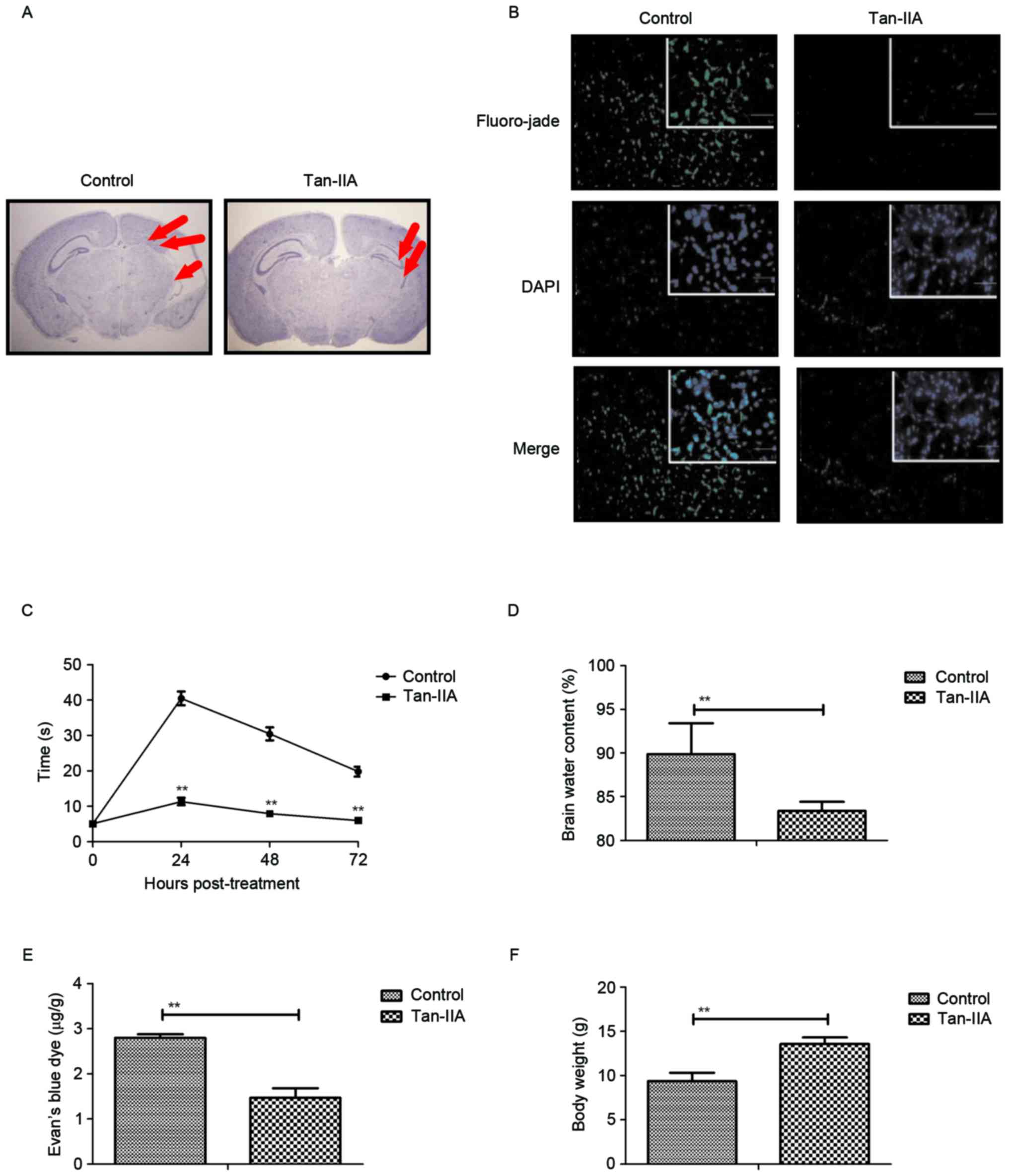
Tan-IIA improves infarct volume and
neuronal degeneration in mice with perinatal HIE
Finally, the effect of Tan-IIA on infarct volume and
neuronal degeneration in mice with perinatal HIE was investigated.
As presented in Fig. 6A, Tan-IIA
treatment decreased infarct volume caused by perinatal HIE. In
order to analyze the efficacy of Tan-IIA on apoptosis of
hippocampal cells, Fluoro-Jade C staining was performed.
Immunohistochemistry demonstrated that Tan-IIA treatment decreased
the number of damaged neurons in the hippocampal region of the
brain of HIE mice (Fig. 6B). In
addition, neurobehavioral tests demonstrated that Tan-IIA treatment
markedly improved physical dysfunction of mice with perinatal HIE
determined by short-term neurobehavioral analysis (Fig. 6C). Furthermore, Tan-IIA treatment
markedly reduced brain water content and blood barrier permeability
in brain in mice with perinatal HIE (Fig. 6D and E). Furthermore, body weight
of mice with perinatal HIE was also increased by Tan-IIA treatment
compared with the control (Fig.
6F). These results suggest that Tan-IIA is beneficial for
improving the infarct volume and neuronal degeneration in mice with
perinatal HIE.
Discussion
HIE is severe brain disease caused by brain hypoxia
and/or ischemic damage, which leads to a series of brain
dysfunction, such as perinatal asphyxia and various neurological
sequelae for newborns (21).
Clinical investigation has indicated that HIE often results in
mental retardation, epilepsy and cerebral palsy (22). Currently, although more and more
therapeutic methods have been proposed, the therapeutic efficacy is
limited for patients with HIE following neuroprotection
pharmacotherapy, regarding improvement of inflammation, apoptosis
and oxidative stress in the central nervous system (23). Tan-IIA is a traditional Chinese
medicine that is used as drug for the treatment of cardiovascular
and cerebrovascular diseases (5).
The current study investigated the therapeutic effects of Tan-IIA
and analyzed the molecular mechanism of Tan-IIA-mediated benefits
for HIE. The data indicated that Tan-IIA improved inflammation,
oxidative stress and apoptosis in a mouse model of HIE.
Inflammation is an important feature of patients
with HIE, and HIE may also lead to other conditions, including
cranial nerve palsy (24). A
previous study reported that inflammation is associated with
metabolic acidosis in HIE, which may aggravate neurological
outcomes (13). Girard et
al (25) reported that
administration of IL-1 receptor antagonist exerts neuroprotective
effects during perinatal inflammation and/or hypoxic-ischemic
injuries. In addition, the role of TNF-α in neuronal apoptosis in
neonatal rats with HIE has been investigated and may presents an
important role in the progression of HIE (11,26).
Furthermore, chemokines CXCL10 and CCL12 have been identified to be
involved in intracranial inflammation and contribute to nerve
injury (27,28). The results of the current study
demonstrated that Tan-IIA reduced the plasma concentration of IL-1,
TNF-α, CXCL10 and CCL12 in the mouse model of HIE. Inflammatory
responses were also downregulated by Tan-IIA treatment.
Oxidative and reductive stress are essential for
dynamic phases experienced by cells undergoing adaptation towards
endogenous or exogenous noxious stimuli (29). Mitochondrial malfunction is the
common denominator arising from the aberrant functioning of the
rheostat that maintains the homeostasis between oxidative and
reductive stress in neurons (30).
Maladaptation during oxidative stress may have a pivotal role in
the pathophysiology of HIE (14,31).
Demarest et al (32) have
analyzed the correlation of mitochondrial respiratory impairment
and oxidative stress in a rat model of neonatal HIE and the results
demonstrated that attenuation of oxidative stress can protect
neurons against the loss of mitochondrial glutathione peroxidase
activity. Evidence has also indicated that endoplasmic reticulum
stress is a crucial risk factor in the pathogenic progression of
HIE. Wang et al (33)
reported that notoginsenoside R1 protects neurons against neonatal
cerebral HIE through estrogen receptor-dependent activation of
endoplasmic reticulum stress pathways. In the current study,
Tan-IIA-mediated endoplasmic reticulum stress and oxidative stress
were investigated in HIE mice. The data demonstrated that Tan-IIA
may be an efficient agent for inhibition of endoplasmic reticulum
stress and oxidative stress in the central nervous system.
Apoptosis of neurons has a crucial role in the
initiation and development of HIE. In the current study, the effect
of Tan-IIA on apoptosis of neurons in mice with HIE was analyzed.
The results provided potential mechanisms of Tan-IIA-mediated
apoptosis resistance. A previous study reported that inhibition of
apoptosis can protect neurons against hypoxic-ischemic injury by
inhibiting parthanatos and necroptosis (34). In addition, Yan et al
(35) also reported that
upregulation of Bax and Bcl-2 expression inhibited neural apoptosis
in neonatal rats with hypoxic-ischemic brain damage. Furthermore,
research has indicated that NF-κB contributes to
6-hydroxydopamine-induced apoptosis of nigral dopaminergic neurons
through downregulation of P53 (36). Furthermore, blocking of the NF-κB
pathway inhibited apoptosis of neurons in transgenic X-linked
inhibitor of apoptotic protein mice and hippocampal rat (37). Notably, the present study
demonstrated that Tan-IIA decreases TLR-4 expression, which has
been reported to be positively associated with apoptosis of
neuroblastoma cells following ischemia-reperfusion injury in the
rat retina in vivo (38).
The findings of the current study illustrated that Tan-IIA
regulates apoptosis of neurons through inhibition of TLR-4-mediated
NF-κB signaling.
In conclusion, intravenous injection of Tan-IIA
treatment reduced neuronal apoptosis through TLR-4-mediated NF-κB
signaling in a neonatal HIE mouse model. Tan-IIA treatment reduced
the plasma levels inflammatory factors, IL-1, TNF-α, CXCL10 and
CCL12, and also reduced endoplasmic reticulum stress and oxidative
stress in the central nervous system. Notably, Tan-IIA treatment
markedly suppresses apoptosis of neurons and reduced the physical
dysfunction in mice following perinatal HIE. Tan-IIA treatment
improved the infarct volume and neuronal degeneration in mice with
perinatal HIE. These investigations suggest that Tan-IIA treatment
may be a potential agent for the treatment of HIE; however, a sham
group was not employed in the present study, which may pose a
limitation. Further preclinical and clinical investigations should
be performed to validate the efficacy and safety of Tan-IIA.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BZ made substantial contributions to the design of
the present study. CZF was responsible for cell culture, detection
of NF-κB activity and flow cytometry analysis. LX was responsible
for establishment of the rat model preparation and
immunohistochemistry. CL and HL analyzed the data and drafted the
manuscript. CHF performed the statistical analysis. WY conducted
the data collection. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the ethics
committee of Renmin Hospital of Wuhan University (Wuhan,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hochwald O, Jabr M, Osiovich H, Miller SP,
McNamara PJ and Lavoie PM: Preferential cephalic redistribution of
left ventricular cardiac output during therapeutic hypothermia for
perinatal hypoxic-ischemic encephalopathy. J Pediatr.
164:999–1004.e1. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Looney AM, Ahearne C, Boylan GB and Murray
DM: Glial Fibrillary acidic protein is not an early marker of
injury in perinatal asphyxia and Hypoxic-ischemic encephalopathy.
Front Neurol. 6:2642015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Khan RH, Islam MS, Haque SA, Hossain MA,
Islam MN, Khaleque MA, Chowdhury B and Chowdhury MA: Correlation
between grades of intraventricular hemorrhage and severity of
hypoxic ischemic encephalopathy in perinatal asphyxia. Mymensingh
Med J. 23:7–12. 2014.PubMed/NCBI
|
|
4
|
Tagin M, Abdel-Hady H, Rahman Ur S,
Azzopardi DV and Gunn AJ: Neuroprotection for perinatal hypoxic
ischemic encephalopathy in low-and Middle-income countries. J
Pediatr. 167:25–28. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li H, Han W, Wang H, Ding F, Xiao L, Shi
R, Ai L and Huang Z: Tanshinone IIA inhibits Glutamate-induced
oxidative toxicity through prevention of mitochondrial dysfunction
and suppression of MAPK activation in SH-SY5Y human neuroblastoma
cells. Oxid Med Cell Longev. 2017:45174862017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim EO, Kang SE, Im CR, Lee JH, Ahn KS,
Yang WM, Um JY, Lee SG and Yun M: Tanshinone IIA induces TRAIL
sensitization of human lung cancer cells through selective ER
stress induction. Int J Oncol. 48:2205–2212. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mao C, Zhang Y, Zhang Y, Cao L, Shao H,
Wang L, Zhu L and Xu Z: The effect of tanshinone IIA on the
cardiovascular system in ovine fetus in utero. Am J Chin Med.
37:1031–1044. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gao S, Liu Z, Li H, Little PJ, Liu P and
Xu S: Cardiovascular actions and therapeutic potential of
tanshinone IIA. Atherosclerosis. 220:3–10. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shen JL, Chen YS, Lin JY, Tien YC, Peng
WH, Kuo CH, Tzang BS, Wang HL, Tsai FJ, Chou MC, et al: Neuron
regeneration and proliferation effects of Danshen and Tanshinone
IIA. Evid Based Complement Alternat Med. 2011:3789072011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hernandez-Jimenez M, Sacristan S, Morales
C, García-Villanueva M, García-Fernández E, Alcázar A, González VM
and Martín ME: Apoptosis-related proteins are potential markers of
neonatal hypoxic-ischemic encephalopathy (HIE) injury. Neurosci
Lett. 558:143–148. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li SJ, Liu W, Wang JL, Zhang Y, Zhao DJ,
Wang TJ and Li YY: The role of TNF-α, IL-6, IL-10, and GDNF in
neuronal apoptosis in neonatal rat with hypoxic-ischemic
encephalopathy. Eur Rev Med Pharmacol Sci. 18:905–909.
2014.PubMed/NCBI
|
|
12
|
Liu L, Liu C, Lu Y and Jiang Y: ER stress
related factor ATF6 and caspase-12 trigger apoptosis in neonatal
hypoxic-ischemic encephalopathy. Int J Clin Exp Pathol.
8:6960–6966. 2015.PubMed/NCBI
|
|
13
|
Johnson CT, Burd I, Raghunathan R,
Northington FJ and Graham EM: Perinatal inflammation/infection and
its association with correction of metabolic acidosis in
hypoxic-ischemic encephalopathy. J Perinatol. 36:448–452. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chapados I and Cheung PY: Not all models
are created equal: Animal models to study hypoxic-ischemic
encephalopathy of the newborn. Commentary on Gelfand SL et
al: A new model of oxidative stress in rat pups (Neonatology
2008;94:293-299). Neonatology. 94:300–303. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cikla U, Chanana V, Kintner DB, Udho E,
Eickhoff J, Sun W, Marquez S, Covert L, Otles A, Shapiro RA, et al:
ERα signaling is required for TrkB-mediated hippocampal
neuroprotection in female neonatal mice after hypoxic ischemic
encephalopathy(1,2,3). eNeuro. 3:pii2016. View Article : Google Scholar
|
|
16
|
Can MM, Tanboğa IH, Türkyilmaz E, Karabay
CY, Akgun T, Koca F, Tokgoz HC, Keles N, Ozkan A, Bezgin T, et al:
The risk of false results in the assessment of platelet function in
the absence of antiplatelet medication: Comparison of the PFA-100,
multiplate electrical impedance aggregometry and verify now assays.
Thromb Res. 125:e132–e137. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time. quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hashiguchi A, Yano S, Nitta K, Ide W,
Hashimoto I, Kamada H and Kuratsu J: Hemisplenial-accompanied by
internal border-zone infarction: Clinical relevance of the splenium
of the corpus callosum as a border-zone area between anterior and
posterior cerebral arteries. J Neurol Neurosurg Psychiatry.
81:704–706. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bechet S, Hill F, Gilheaney Ó and Walshe
M: Diagnostic accuracy of the modified Evan's blue dye test in
detecting aspiration in patients with tracheostomy: A systematic
review of the evidence. Dysphagia. 31:721–729. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Farahat FM, Rohlman DS, Storzbach D,
Ammerman T and Anger WK: Measures of short-term test-retest
reliability of computerized neurobehavioral tests. Neurotoxicology.
24:513–521. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sant'Anna G, Laptook AR, Shankaran S, Bara
R, McDonald SA, Higgins RD, Tyson JE, Ehrenkranz RA, Das A,
Goldberg RN, et al: Phenobarbital and temperature profile during
hypothermia for hypoxic-ischemic encephalopathy. J Child Neurol.
27:451–457. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shankaran S: Hypoxic-ischemic
encephalopathy and novel strategies for neuroprotection. Clin
Perinatol. 39:919–929. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peliowski-Davidovich A: Hypothermia for
newborns with hypoxic ischemic encephalopathy. Paediat Child
Health. 17:41–43. 2012. View Article : Google Scholar
|
|
24
|
Barks JD, Liu YQ, Shangguan Y, Li J, Pfau
J and Silverstein FS: Impact of indolent inflammation on neonatal
hypoxic-ischemic brain injury in mice. Int J Dev Neurosci.
26:57–65. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Girard S, Sebire H, Brochu ME, Briota S,
Sarret P and Sébire G: Postnatal administration of IL-1Ra exerts
neuroprotective effects following perinatal inflammation and/or
hypoxic-ischemic injuries. Brain Behav Immun. 26:1331–1339. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aly H, Khashaba MT, El-Ayouty M, El-Sayed
O and Hasanein BM: IL-1beta, IL-6 and TNF-alpha and outcomes of
neonatal hypoxic ischemic encephalopathy. Brain Dev. 28:178–182.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gotsch F, Romero R, Friel L, Kusanovic JP,
Espinoza J, Erez O, Than NG, Mittal P, Edwin S, Yoon BH, et al:
CXCL10/IP-10: A missing link between inflammation and
anti-angiogenesis in preeclampsia? J Matern Fetal Neonatal Med.
20:777–792. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tomita K, Freeman BL, Bronk SF, LeBrasseur
NK, White TA, Hirsova P and Ibrahim SH: CXCL10-mediates macrophage,
but not other innate immune cells-associated inflammation in murine
nonalcoholic steatohepatitis. Sci Rep. 6:287862016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kurian GA, Rajagopal R, Vedantham S and
Rajesh M: The role of oxidative stress in myocardial ischemia and
reperfusion injury and remodeling: Revisited. Oxid Med Cell Longev.
2016:16564502016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Perrone S, Szabo M, Bellieni CV, Longini
M, Bangó M, Kelen D, Treszl A, Negro S, Tataranno ML and Buonocore
G: Whole body hypothermia and oxidative stress in babies with
hypoxic-ischemic brain injury. Pediatr Neurol. 43:236–240. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ten VS, Yao J, Ratner V, Sosunov S, Fraser
DA, Botto M, Sivasankar B, Morgan BP, Silverstein S, Stark R, et
al: Complement component c1q mediates mitochondria-driven oxidative
stress in neonatal hypoxic-ischemic brain injury. J Neurosci.
30:2077–2087. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Demarest TG, Schuh RA, Waddell J, McKenna
MC and Fiskum G: Sex-dependent mitochondrial respiratory impairment
and oxidative stress in a rat model of neonatal hypoxic-ischemic
encephalopathy. J Neurochem. 137:714–729. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Y, Tu L, Li Y, Chen D and Wang S:
Notoginsenoside R1 protects against neonatal cerebral
Hypoxic-ischemic injury through estrogen Receptor-dependent
activation of endoplasmic reticulum stress pathways. J Pharmacol
Exp Ther. 357:591–605. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kong D, Zhu J, Liu Q, Jiang Y, Xu L, Luo
N, Zhao Z, Zhai Q, Zhang H, Zhu M and Liu X: Mesenchymal stem cells
protect neurons against hypoxic-ischemic injury via inhibiting
parthanatos, necroptosis, and apoptosis, but not autophagy. Cell
Mol Neurobiol. 37:303–313. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yan SZ, Wang XL, Wang HY, Dong P and Zhao
YS: Effects of umbilical cord blood mononuclear cells
transplantation via lateral ventricle on the neural apoptosis and
the expression of Bax and Bcl-2 proteins in neonatal rats with
hypoxic-ischemic brain damage. Zhongguo Dang Dai Er Ke Za Zhi.
18:862–866. 2016.(In Chinese). PubMed/NCBI
|
|
36
|
Liang ZQ, Li YL, Zhao XL, Han R, Wang XX,
Wang Y, Chase TN, Bennett MC and Qin ZH: NF-kappaB contributes to
6-hydroxydopamine-induced apoptosis of nigral dopaminergic neurons
through p53. Brain Res. 1145:190–203. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kairisalo M, Korhonen L, Sepp M, Pruunsild
P, Kukkonen JP, Kivinen J, Timmusk T, Blomgren K and Lindholm D:
NF-kappaB-dependent regulation of brain-derived neurotrophic factor
in hippocampal neurons by X-linked inhibitor of apoptosis protein.
Eur J Neurosci. 30:958–966. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ulbrich F, Lerach T, Biermann J, Kaufmann
KB, Lagreze WA, Buerkle H, Loop T and Goebel U: Argon mediates
protection by interleukin-8 suppression via a TLR2/TLR4/STAT3/NF-κB
pathway in a model of apoptosis in neuroblastoma cells in vitro and
following ischemia-reperfusion injury in rat retina in vivo. J
Neurochem. 138:859–873. 2016. View Article : Google Scholar : PubMed/NCBI
|