Introduction
Recently, infertility has become one of the more
common diseases endangering human health, as a result of the
long-term impact of industrialization, and contaminated
environments, water and food on human health. In addition to
reproductive system diseases, unknown harmful substances are the
leading cause of infertility, habitual abortion and embryonic
defects (1,2). In China, the number of patients with
infertility has increased rapidly in the past 10 years, and
infertility currently affects ~15% of females of reproductive age
(3). With the development of
detection techniques, such as polymerase chain reaction (PCR),
fluorescence in situ hybridization (FISH), and microarray
analysis, researchers have demonstrated that the incidence of
chromosomal abnormalities in human embryos is very high for women
that use assisted reproductive techniques (4). It is considered that the incidence of
chromosomal abnormalities in embryos increases with advancing age
in females (5,6); however, a significant upward trend in
such abnormalities has been observed among young patients (4). Aneuploid embryos are primarily
derived from oocytes, and their formation is prevalent among young
women and may be associated with various environmental changes.
Extensive studies have confirmed that over 50% of
reproductive and developmental defects are caused by chemical
substances and environmental factors, including heavy metals, such
as lead, cadmium, nickel and chromium, and organic compounds,
including dioxins, aldehydes and phenols (7–9).
However, the causes of chromosome abnormalities, as well as the
signaling pathways and molecular mechanisms involved in oocytes,
remain unclear. It has been reported that heavy metals can trigger
oxidative stress, which may reduce the level of antioxidants in
cells and also cause abnormal chromosomal division. This finding
suggests that the formation of aneuploidy may be associated with
the levels of active oxygen and antioxidants in oocytes and that
heavy metal coupling agents and antioxidants may effectively reduce
chromosomal abnormalities. In the present study, we explored the
effects of heavy metal coupling agents and antioxidants on the
ageing of mouse oocytes.
Materials and methods
Grouping and treatment of mice
Female C57BL/6 mice (body weight, 15±2 g) at 3 weeks
of age were obtained from Guangdong Medical Laboratory Animal
Center. All experimental animals were housed individually under a
12/12 h light/dark cycle in a specific pathogen-free animal
facility at the Laboratory Animal Center of Southern Medical
University (Guangdong, China), with controlled temperature (23±2°C)
and relative humidity (40–60%). Food and water were available ad
libitum during the experimental period. The experimental
procedures and animal conditions were approved by the Animal Ethics
Committee of The Third Affiliated Hospital of Guangzhou Medical
University.
The mice were divided into four groups as described
below (n=12 per group). Mice in the heavy metal coupling agent
group (group A) were given pure water containing 0.3 mmol/l EDTA
and 0.15 mmol/l sodium citrate (Sigma-Aldrich; Merck KGaA,
Darmstadt Germany). Mice in the antioxidant group (group B) were
given pure water supplemented with 40 mg/l lipoic acid and 100 mg/l
acetyl carnitine (GNC Company, Propinsi Lampung, Indonesia). All
concentrations were selected on basis of preliminary experiments
(data not shown). Mice in the mixed group (group C) were given pure
water supplemented with heavy metal coupling agents and
antioxidants. Mice in the normal control group (group D) were given
pure water. The pure water was processed using aseptic filtration.
Mice were randomly selected from each group for subsequent
experiments following 3, 6, 9 and 12 months of treatment.
Oocyte collection
Female mice were superovulated by intraperitoneal
injection of 10 IU of pregnant mare serum gonadotropin (PMSG),
followed 46–48 h later by 10 IU of human chorionic gonadotropin
(HCG; both Ningbo Second Hormone Factory, Zhejiang, China).
Drinking water was provided to mice from each group during the
promotion of ovulation. The mice were sacrificed by cervical
dislocation after 12–13 h to collect the oocytes. Briefly, the
abdominal cavity was opened exposing the uterus, fallopian tubes
and ovaries, followed by separation of blood vessels and adipose
tissue. Then, the fallopian tubes were removed and quickly placed
in human fallopian tube fluid (HTF) which was artificially
prepared, and the fallopian tubes of mice in the same group were
grouped together. Following rinsing with HTF three times, the
magnum of the fallopian tubes was removed to release cumulus-oocyte
complexes (COCs). The COCs were washed twice with HTF and incubated
with hyaluronidase (1:9; Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) to remove granular lymphocytes and obtain
MII-stage nude oocytes. The oocytes were counted following three
washes with HTF and incubated at 37°C in a humidified 5% CO2
incubator in 40 µl drops of HTF medium covered with mineral oil
(Sigma-Aldrich; Merck KGaA). The mice reared for 12 months did not
have a sufficient number of oocytes, thus, only the number of
oocytes in this age group was counted.
Evaluation of the mitochondrial
membrane potential in mature oocytes using JC-1 dye
A JC-1 mitochondrial membrane potential detection
kit (Sigma-Aldrich; Merck KGaA) was used to evaluate the
mitochondrial membrane potential. First, 1X working solution was
prepared with 200X JC-1, dyeing buffer and pure water according to
the manufacturer's protocol, then preheated at 37°C in a water
bath. Next, 50 µl of 1X working solution was divided into several
droplets, which were covered with mineral oil and preheated at
37°C. Subsequently, 15–20 MII-stage oocytes were incubated for
20–25 min with the working solution at 37°C in 5% CO2, followed by
2–3 washes. JC-1 mitochondrial fluorescence was visualized with a
laser-scanning confocal microscope (Nikon-C2Si; Nikon Corp., Tokyo,
Japan), and ImageJ software was used for quantitative analysis.
Chromosome examination in mature
oocytes
MII-stage oocytes were digested in 3% trypsin for
2–5 min to clear the zona pellucida and then quickly transferred to
the fresh HTF droplets. Appropriately sized circles were drawn on a
clean glass slide using a crayon, and each circle was filled with
an appropriate amount of 1% paraformaldehyde (Sigma-Aldrich; Merck
KGaA) with 5–10 oocytes. After the nuclear membrane was ruptured,
the oocytes were fixed, dried and marked, and stored in a −20°C
freezer. For imaging, the glass slides were removed, stained for 10
min with 10 µg/ml propidium iodide (PI) in the dark, covered with a
cover glass, mounted with nail polish, and then stored at −20°C.
The number of chromosomes was calculated in each oocyte.
Examination of spindle morphology in
mature oocytes
First, oocytes were fixed. Briefly, 25 MII-stage
oocytes from each group with normal morphology were fixed with 4%
paraformaldehyde at room temperature for 30 min. Following
permeabilization with 0.5% Triton X-100 in phosphate-buffered
saline (PBS) for 20 min, the oocytes were blocked in a blocking
solution (PBS containing 1% bovine serum albumin) for 1 h and then
incubated with an α-tubulin antibody (Sigma-Aldrich; Merck KGaA)
diluted in blocking solution for 1.5 h at room temperature or
overnight at 4°C. After the oocytes were washed with a washing
solution (PBS with 0.1% Tween-20 and 0.01% Triton X-100) three
times (5 min/wash), the chromosomes were stained with PI
(Sigma-Aldrich; Merck KGaA) for 15 min, and the oocytes were then
mounted on glass slides and observed using a laser-scanning
confocal microscope.
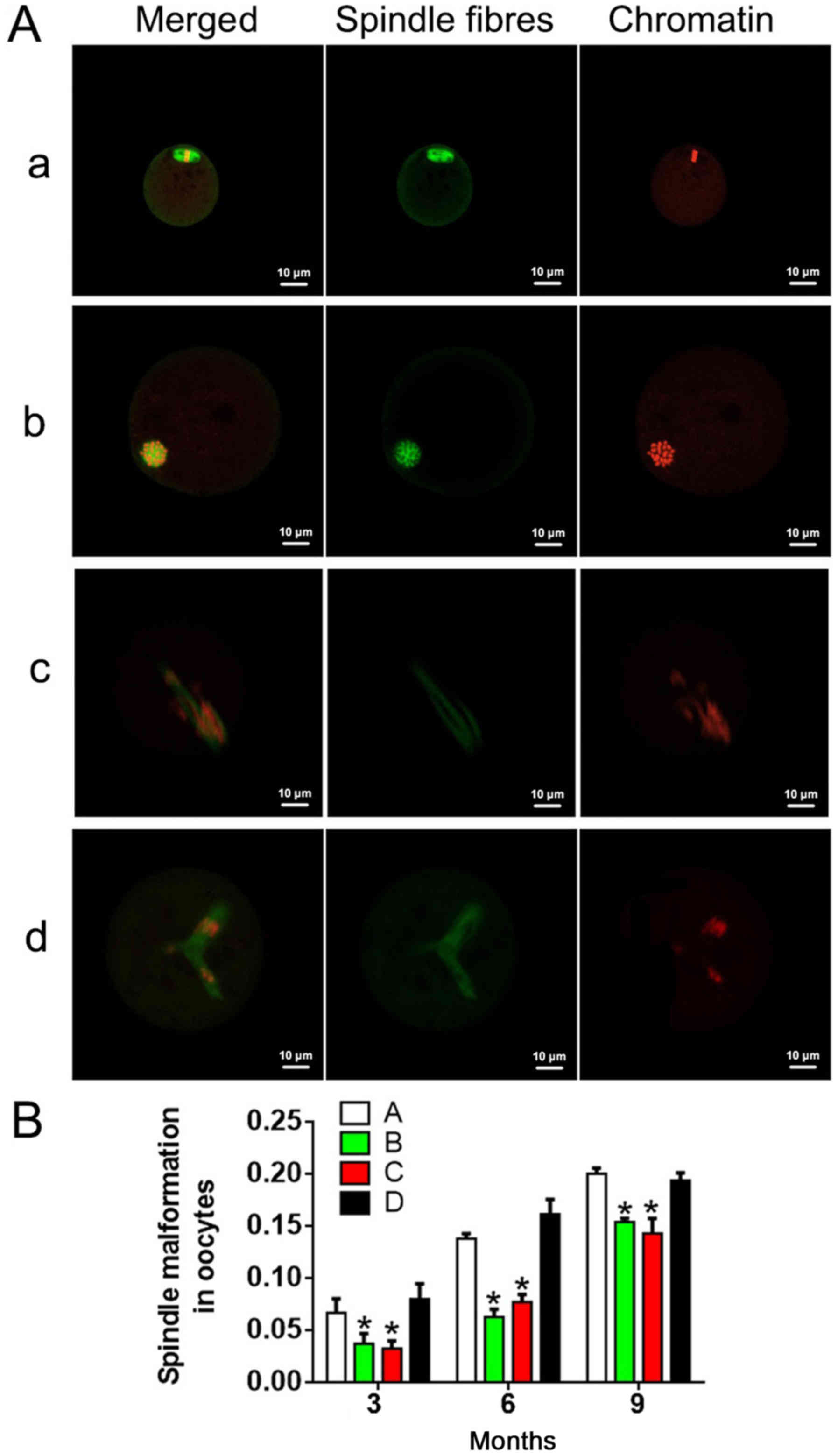
Next, the spindles were observed. In normal oocytes,
the spindles exhibit a fusiform shape and have a granular structure
at the two poles, with regularly arranged spindle microtubules
(green) connecting the spindle poles to the chromosomes (red) that
are arranged in order on the equatorial plate. However, in an
abnormal state, the spindle length is contracted, the microtubules
are irregular or completely absent, and the chromosomes are
arranged outside of the equatorial plate, with a dispersed
distribution, or only lightly stained.
Statistical analysis
Statistical analyses were performed using SPSS
version 13.0 (SPSS Inc., Chicago, IL, USA). All experimental data
are presented as percentages and were analyzed using the
χ2 test. A repeated-measures analysis of variance was
also performed and post hoc multiple factor analysis of variance
with Scheffé's test were applied to compare the quantitative data,
with α=0.05 as a test standard. P<0.05 was considered to
indicate a statistically significant difference.
Results
Antioxidants effectively maintain
oocyte numbers
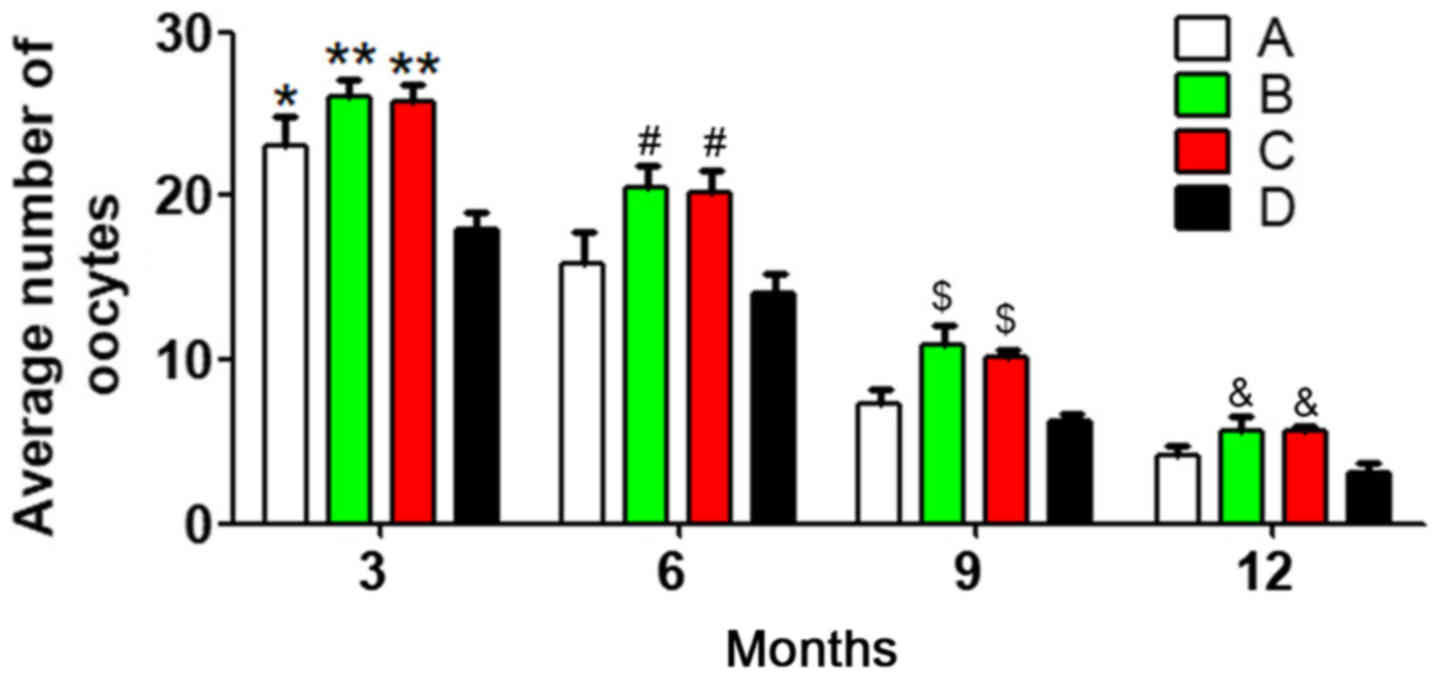
To reduce individual differences, 10 mice were
randomly selected from each group at each age stage, and the
average number of oocytes was calculated. Analysis of variance and
multiple-factor repetitive analysis revealed that the primary
effect of age in each group was significantly different
(P<0.05), the differences in the oocyte numbers at different
ages were statistically significant (P<0.05), and different
associations were observed between different groups and different
age stages (P<0.05). There were significant differences in the
number of oocytes between group D and the other three groups at 3
months (P<0.05). In addition, at 6, 9 and 12 months, the oocyte
number was significantly different between group B and group D, and
between group C and group D (both P<0.05) (Fig. 1). These results demonstrated that
with prolonged treatment times, the oocyte number was
better-maintained in the antioxidant group compared with that in
the heavy metal coupling agent and normal control groups.
Meanwhile, the oocyte number in the heavy metal coupling agent
group was significantly different from that in the normal control
group at 3 months, indicating that heavy metal coupling agents
serve a role in maintaining oocyte numbers, but the effect was not
as evident as that of antioxidants. One possible reason might be
that the mice were not exposed to many heavy metals during
treatment, which made the heavy metal coupling agents less
useful.
Antioxidants effectively alleviate the
decrease in mitochondrial activity
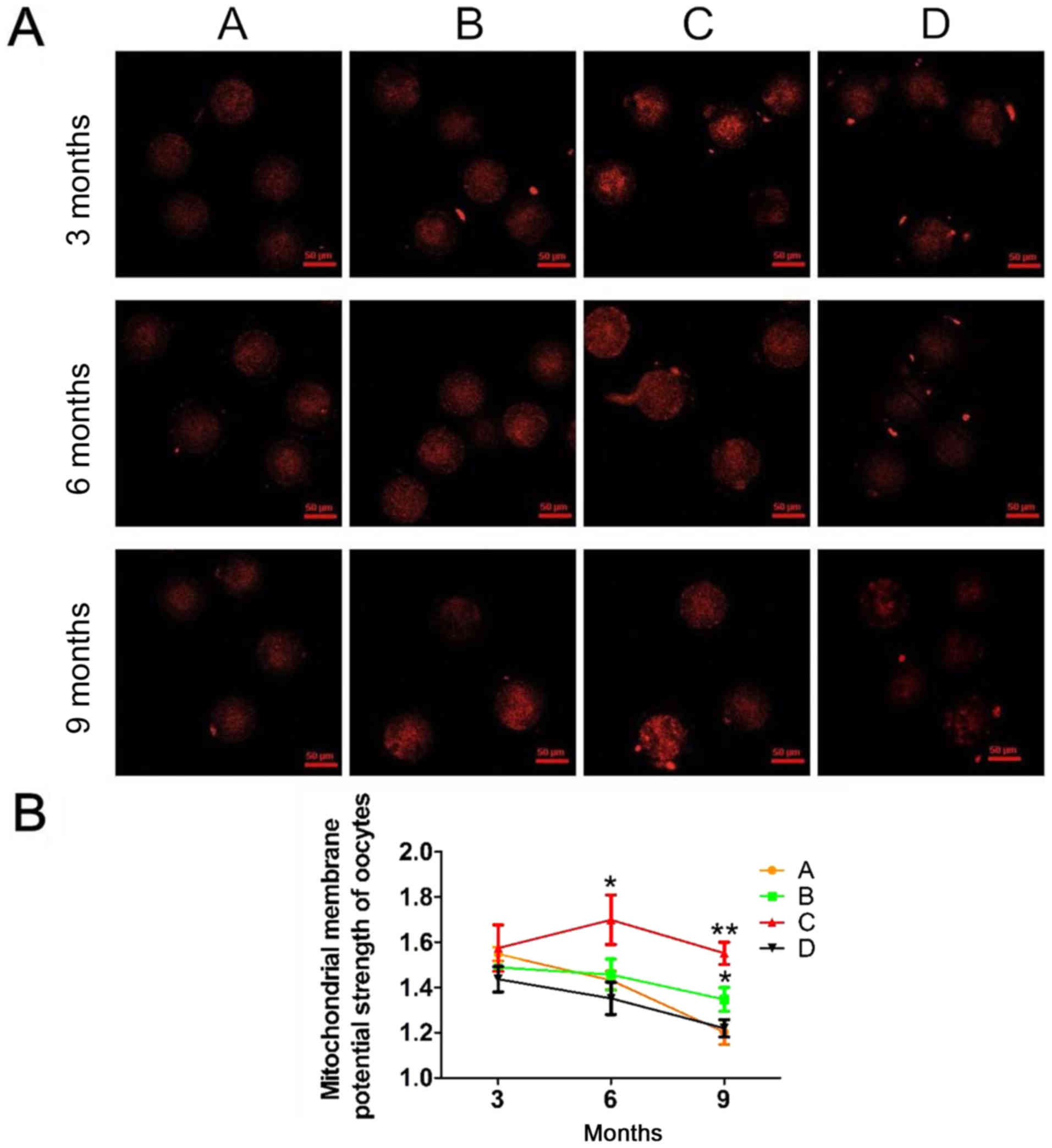
In our experiment, mitochondrial activity in mouse
oocytes decreased with age (Fig.
2A). As shown in Fig. 2B, in
the four groups, mitochondrial activity, represented as red
fluorescence, was significantly lower at 9 months compared with at
3 or 6 months. In addition, groups B and C had significantly higher
mitochondrial activity levels compared with groups A and B at 9
months. Furthermore, no significant differences were observed in
mitochondrial activity between the four groups at 3 and 6 months.
These results suggested that antioxidants effectively alleviated
the reduction in mitochondrial activity.
Antioxidants reduce the incidence of
chromosomal abnormality
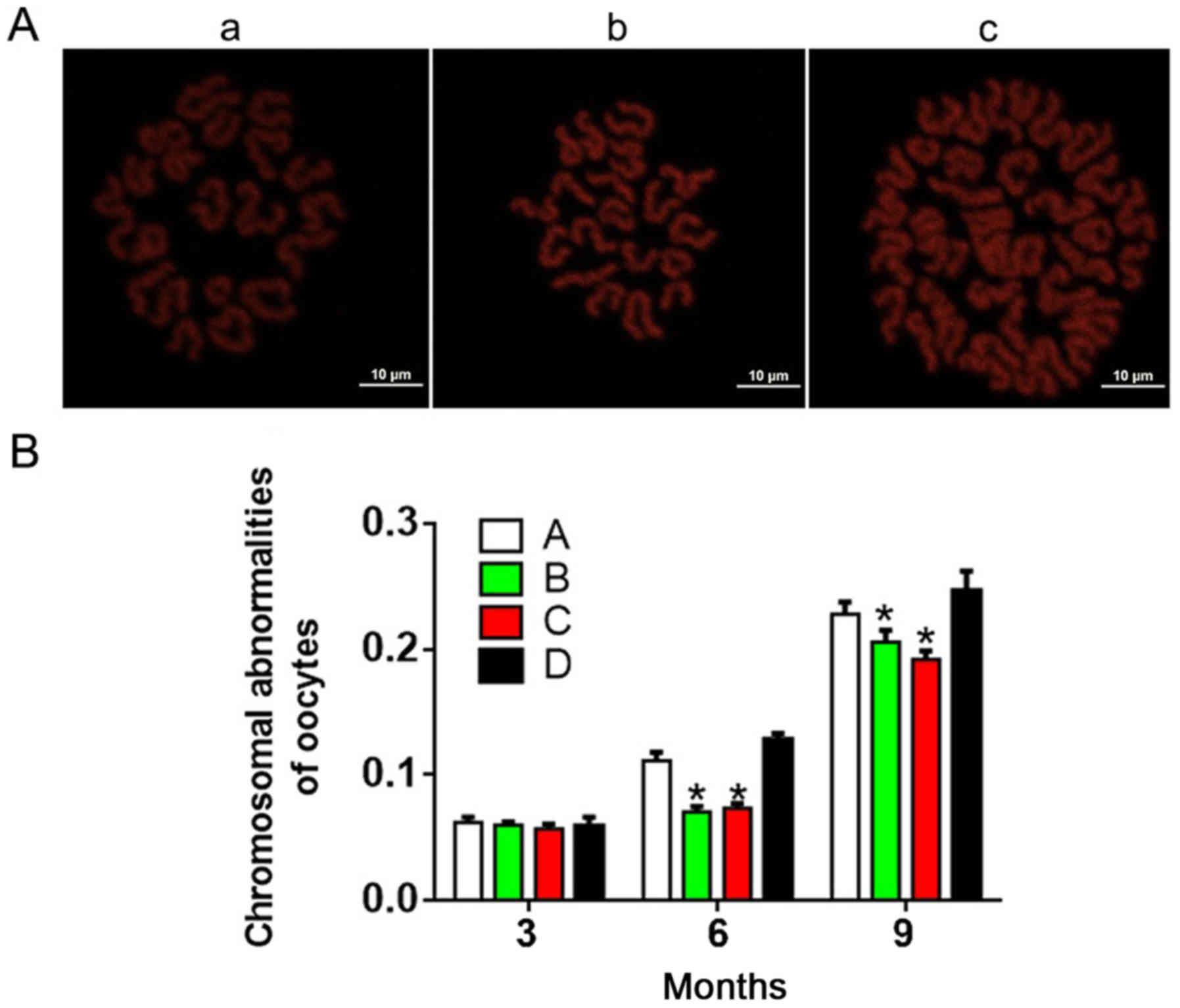
As shown in Fig. 3,
the incidence of chromosomal abnormality increased with age and/or
treatment time in each group. At 3 months, there were no
significant differences in chromosome abnormalities between the
four groups (P>0.05). Interestingly, groups A and D exhibited
significantly increased chromosomal abnormalities compared with
groups B and C at 6 and 9 months. These results indicated that
antioxidants reduced the incidence of chromosomal
abnormalities.
Antioxidants reduce the incidence of
spindle abnormality
Immunofluorescence analysis was performed to detect
the condition of spindles in oocytes (Fig. 4A). As shown in Fig. 4B, the incidence of spindle
abnormality increased with age and/or treatment time in each group.
Groups B and C exhibited no significant differences in spindle
abnormality rate at any time point (P>0.05). There were
statistically significant differences in spindle abnormalities
between group A and group D at the same time points (P<0.05). At
6 and 9 months, the spindle abnormalities in groups B and C were
remarkably lower compared with that in groups A and D. These
results suggested that antioxidants reduced the incidence of
spindle abnormalities.
Discussion
Oocyte ageing, and germ cell dysfunction and
depletion, accompanied by follicular and ovarian atrophy and
endocrine abnormalities, have been widely suggested to be the major
causes of reproductive ageing, menopause and associated
pathologies, including infertility (10,11).
It has been reported that oxidative stress serves an important role
in oocyte ageing, and increased levels of reactive oxygen species
(ROS) have been identified in the follicular fluid of elderly women
(12,13). Oxidative stress is defined as an
imbalance between increased levels of ROS and low activity of
antioxidative mechanisms. A variety of destructive stimuli can
cause the overproduction of highly reactive molecules, such as ROS
and reactive nitrogen species (RNS), whereby the antioxidative
mechanisms are unable to eliminate them. A high concentration of
ROS can destroy DNA, protein and lipids, leading to impairments in
gene expression, damage to cell structures and functions, and even
cell death, which is considered to be a major mechanism involved in
organism ageing and diseases. Previous studies have demonstrated
that a high level of ROS is associated with reduced meiosis
(14,15), chromosome abnormalities, spindle
microtubule deformation (16–19)
and reduced embryonic development potential (20,21).
These findings suggest that oxidative stress may partially
contribute to the decline in fertility that is directly associated
with oocyte ageing. Thus, we speculated that oocyte ageing might be
effectively blocked by reducing ROS via antioxidant
supplementation.
There are two types of antioxidant defense systems
in the human body: The enzymatic antioxidant system, including
superoxide dismutase (SOD), catalase (CAT) and glutathione
peroxidase (GSH-Px); and the non-enzymatic antioxidant system,
including glutathione, α-lipoic acid, carotenoids, and vitamin C.
Our study revealed that the number and quality of mouse oocytes
increased following prolonged treatment with the antioxidants
lipoic acid and acetyl carnitine. Consistent with these results,
Tarín et al (22) reported
that oral administration of vitamin C and vitamin E could also
offset the negative effects of female ageing on the number and
quality of oocytes, and the response to oxidative stress has
clinical significance in preventing female age-associated
aneuploidy (23). Moreover, Liu
et al (24) demonstrated
that the quality and number of oocytes significantly increased
following N-acetyl-L-cysteine treatment. Collectively, these
studies suggest that antioxidants exert positive effects on
oocytes.
Increased aneuploidy is considered an important
indicator of oocyte senescence. In 1965, Harman first proposed the
Mitochondrial Free Radical Theory of Aging, postulating that
the destruction of DNA and cell membranes caused by peroxidation is
the primary reason for the decline in cell function with age
(25). Soon afterward, in 1972,
Harman put forward the concept of mitochondrial senescence
(26). Mitochondria are the most
abundant organelles in oocytes, providing energy for various
chemical reactions and other organelles. Thus, with fewer
mitochondria, the chemical reactions in oocytes cannot easily
proceed, reducing the activity of oocytes and embryos, and
subsequently leading to decreased fertility and increased
aneuploidy. Recently, an increasing number of studies have focused
on the association between mitochondrial metabolism and oocyte
maturation, fertilization and early embryonic development (15,27,28).
It has been reported that oocyte aging, mitochondrial apoptosis and
a decrease in mitochondrial membrane potential are associated with
increased age (29). In our study,
mitochondrial activity in oocytes was significantly lower at 9
months compared with at 3 and 6 months, which supports the findings
by Tarín et al (12),
whereby there is a significant correlation between age and
oxidative stress in mammals (15).
Additionally, mitochondrial activity was significantly decreased in
the antioxidant group compared with the heavy metal coupling agent
and normal control groups, indicating that antioxidants could
effectively maintain the activity of mitochondria, thus, deferring
the process of oocyte senescence. It is generally known that
spindles serve a regulatory role in chromosome arrangement and
separation (30,31). The majority of aneuploid embryos
originate from the two meiotic divisions, particularly from the
initial meiosis (32,33). In this study, we examined the
structures of chromosomes and spindles, and found that chromosomal
and spindle abnormalities increased in an age-dependent manner. We
also confirmed that antioxidants could effectively remove excess
ROS in cells and maintain mitochondrial activity, thereby
protecting the normal morphology of spindles and chromosomes in
oocytes.
It is traditionally considered that the number of
female oocytes is fixed at birth and decreases at various rates
during life until the ovarian reserve is exhausted, and menopause
is reached (34). The results of
this study confirmed that ovarian senescence indicated a decline in
the number and quality of oocytes. In addition, our experiments
demonstrated that lipoic acid and acetyl carnitine could not
prevent the negative effects caused by ovarian ageing but could
significantly delay this process. However, heavy metal coupling
agents, including EDTA and sodium citrate, showed no significant
effect on oocytes, possibly because the experimental mice were not
exposed to heavy metals. Understandably, research on the mechanism
of oocyte ageing is of great significance for women seeking to use
assisted reproductive strategies.
In conclusion, our data demonstrated that lipoic
acid and acetyl carnitine can delay the ageing process of germ
cells, which may be achieved by protecting oocytes against
senescence via the maintenance of mitochondrial activity, and
prevention of chromosomal and spindle abnormalities.
Acknowledgements
The authors would like to thank the Laboratory
Animal Center of Guangzhou Medical University (Guangdong, China)
for providing the animal breeding sites.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81270745), the
Science and Information Technology of Guangzhou Key Project (grant
no. 201508020258) and Guangdong Province Science and Technology
Project (grant no. 2016B030229008).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YXX, XFS and WHW conceived and designed the study.
YXX, LFL, STQ, BS, SMO and YHX performed the experiments. YXX and
YJX conducted the statistical analysis. YXX, XFS and WHW wrote the
paper. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental procedures and animal conditions
were approved by the Animal Ethics Committee of the Third
Affiliated Hospital of Guangzhou Medical University (Guangzhou,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Herrera LA, Prada D, Andonegui MA and
Dueñas-González A: The epigenetic origin of aneuploidy. Curr
Genomics. 9:43–50. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dulskiene V and Maroziene L: Impact on
environmental factors on the reproductive system and fetal
development. Medicina (Kaunas). 38:1072–1077. 2002.(In Lithuanian).
PubMed/NCBI
|
|
3
|
Montoya JM, Bernal A and Borrero C:
Diagnostics in assisted human reproduction. Reprod Biomed Online.
5:198–210. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baart EB, Martini E, van den Berg I,
Macklon NS, Galjaard RJ, Fauser BC and Van Opstal D:
Preimplantation genetic screening reveals a high incidence of
aneuploidy and mosaicism in embryos from young women undergoing
IVF. Hum Reprod. 21:223–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jones KT: Meiosis in oocytes:
Predisposition to aneuploidy and its increased incidence with age.
Hum Reprod Update. 14:143–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chiang T, Schultz RM and Lampson MA:
Meiotic origins of maternal age-related aneuploidy. Biol Reprod.
86:1–7. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zenzes MT: Smoking and reproduction: Gene
damage to human gametes and embryos. Hum Reprod Update. 6:122–131.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kumar S: Occupational exposure associated
with reproductive dysfunction. J Occup Health. 46:1–19. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gerhard I, Waibel S, Daniel V and
Runnebaum B: Impact of heavy metals on hormonal and immunological
factors in women with repeated miscarriages. Hum Reprod Update.
4:301–309. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tatone C, Amicarelli F, Carbone MC,
Monteleone P, Caserta D, Marci R, Artini PG, Piomboni P and
Focarelli R: Cellular and molecular aspects of ovarian follicle
ageing. Hum Reprod Update. 14:131–142. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Coccia ME and Rizzello F: Ovarian reserve.
Ann N Y Acad Sci. 1127:27–30. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tarín JJ: Potential effects of
age-associated oxidative stress on mammalian oocytes/embryos. Mol
Hum Reprod. 2:717–724. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tarín JJ: Aetiology of age-associated
aneuploidy: A mechanism based on the ‘free radical theory of
ageing’. Hum Reprod. 10:1563–1565. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tamura H, Takasaki A, Miwa I, Taniguchi K,
Maekawa R, Asada H, Taketani T, Matsuoka A, Yamagata Y, Shimamura
K, et al: Oxidative stress impairs oocyte quality and melatonin
protects oocytes from free radical damage and improves
fertilization rate. J Pineal Res. 44:280–287. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chaube SK, Prasad PV, Thakur SC and
Shrivastav TG: Hydrogen peroxide modulates meiotic cell cycle and
induces morphological features characteristic of apoptosis in rat
oocytes cultured in vitro. Apoptosis. 10:863–874. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang X, Wu XQ, Lu S, Guo YL and Ma X:
Deficit of mitochondria-derived ATP during oxidative stress impairs
mouse MII oocyte spindles. Cell Res. 16:841–850. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tarín JJ, Vendrell FJ, Ten J, Blanes R,
van Blerkom J and Cano A: The oxidizing agent tertiary butyl
hydroperoxide induces disturbances in spindle organization,
c-meiosis, and aneuploidy in mouse oocytes. Mol Hum Reprod.
2:895–901. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu L and Keefe DL: Ageing-associated
aberration in meiosis of oocytes from senescence-accelerated mice.
Hum Reprod. 17:2678–2685. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Choi WJ, Banerjee J, Falcone T, Bena J,
Agarwal A and Sharma RK: Oxidative stress and tumor necrosis
factor-alpha-induced alterations in metaphase II mouse oocyte
spindle structure. Fertil Steril. 88 4 Suppl:S1220–S1231. 2007.
View Article : Google Scholar
|
|
20
|
Tarín JJ, Gómez-Piquer V, Pertusa JF,
Hermenegildo C and Cano A: Association of female aging with
decreased parthenogenetic activation, raised MPF, and MAPKs
activities and reduced levels of glutathione S-transferases
activity and thiols in mouse oocytes. Mol Reprod Dev. 69:402–410.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bhattacharya S, Maheshwari A and Mollison
J: Factors associated with failed treatment: An analysis of 121,744
women embarking on their first IVF cycles. PLoS One. 8:e822492013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tarín JJ, Pérez-Albalá S and Cano A: Oral
antioxidants counteract the negative effects of female aging on
oocyte quantity and quality in the mouse. Mol Reprod Dev.
61:385–397. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tarín JJ, Vendrell FJ, Ten J and Cano A:
Antioxidant therapy counteracts the disturbing effects of diamide
and maternal ageing on meiotic division and chromosomal segregation
in mouse oocytes. Mol Hum Reprod. 4:281–288. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu J, Liu M, Ye X, Liu K, Huang J, Wang
L, Ji G, Liu N, Tang X, Baltz JM, et al: Delay in oocyte aging in
mice by the antioxidant N-acetyl-L-cysteine (NAC). Hum Reprod.
27:1411–1420. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harman D: The free radical theory of
aging: Effect of age on serum copper levels. J Gerontol.
20:151–153. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Harman D: The biologic clock: The
mitochondria? J Am Geriatr Soc. 20:145–147. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Takahashi T, Takahashi E, Igarashi H,
Tezuka N and Kurachi H: Impact of oxidative stress in aged mouse
oocytes on calcium oscillations at fertilization. Mol Reprod Dev.
66:143–152. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu L, Trimarchi JR and Keefe DL:
Involvement of mitochondria in oxidative stress-induced cell death
in mouse zygotes. Biol Reprod. 62:1745–1753. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang D, Keilty D, Zhang ZF and Chian RC:
Mitochondria in oocyte aging: Current understanding. Facts Views
Vis Obgyn. 9:29–38. 2017.PubMed/NCBI
|
|
30
|
Wang WH and Sun QY: Meiotic spindle,
spindle checkpoint and embryonic aneuploidy. Front Biosci.
11:620–636. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vogt E, Kirsch-Volders M, Parry J and
Eichenlaub-Ritter U: Spindle formation, chromosome segregation and
the spindle checkpoint in mammalian oocytes and susceptibility to
meiotic error. Mutat Res. 651:14–29. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hassold T and Hunt P: To err (meiotically)
is human: The genesis of human aneuploidy. Nat Rev Genet.
2:280–291. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hassold T, Hall H and Hunt P: The origin
of human aneuploidy: Where we have been, where we are going. Hum
Mol Genet 16 Spec No. 2:R203–R208. 2007. View Article : Google Scholar
|
|
34
|
Djahanbakhch O, Ezzati M and Zosmer A:
Reproductive ageing in women. J Pathol. 211:219–231. 2007.
View Article : Google Scholar : PubMed/NCBI
|