Introduction
Diabetic mellitus (DM) is a progressive disease and
is usually associated with numerous complications (1–4). The
incidence of DM is predicted to reach 7.7% by 2030 on a global
scale (5,6). Diabetic nephropathy (DN) is one of
the most common microvascular complications associated with DM and
has become the second leading cause of end-stage renal disease in
China (7). Numerous studies have
demonstrated that well-managed blood glucose, blood pressure and
blood lipid levels, and medicinal application of
angiotensin-converting enzyme inhibitors or angiotensin receptor
blockers, may postpone disease progression (8,9). The
exact mechanisms underlying the pathogenesis of DN remain unclear.
A previous study revealed that the onset of DN may be pertinent to
oxidative stress in living cells (10). Podocytes are considered to be the
most critical component of glomerular permselectivity (11). Changes in podocyte morphology and
dysfunction are closely associated with renal diseases (12,13).
The apoptosis of podocytes has been regarded as an important factor
in the progression of DN (14).
Therefore, investigating the molecular mechanism associated with
the inhibition of podocyte injury is important for the development
of novel therapeutic strategies for the treatment of patients with
DN.
Krüppel-like factors (KLFs) are a type of
transcriptional regulatory factor. KLFs are associated with cell
proliferation, migration, apoptosis and tissue remodeling (15,16).
Furthermore, KLFs are associated with the development of numerous
diseases, including cardiovascular disease and cancer (17–19).
KLF5, a member of the KLF family, can be regulated via
phosphorylation, acetylation and ubiquitination following
translation (20,21). KLF5 is associated with hypertensive
nephropathy and diabetic retinopathy (22). However, the effects of KLF5 on DN
are not fully understood.
Puromycin aminonucleoside (PAN) may disrupt the
morphology of podocytes, trigger the overproduction of reactive
oxygen species and induce nephrosis. Therefore, PAN is frequently
used to establish a nephropathy model (23,24).
The aim of the present study was to investigate the effect of KLF5
on PAN-induced injury of podocytes and to determine the underlying
molecular mechanism. Therefore, the results of the present study
may further the understanding of molecular mechanisms associated
with DN and provide a potential therapeutic target for DN.
Materials and methods
Cell culture and cell treatment
Podocytes (MPC-5 cells) were acquired from the Bena
Culture Collection (Beijing, China). Podocytes were cultured in Ham
Nutrient Mix F12-Dulbecco modified Eagle medium (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 1% penicillin-streptomycin
G (Biochrom, Ltd., Cambridge, UK) and 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.). Podocytes were seeded in
6-well plates at a concentration of 3×105 cells/ml and
subsequently treated with PBS (control), or with PAN (0, 5, 10, 20,
40, 60, 80 and 100 µg/ml) for 0, 6, 12, 24 and 48 h time intervals
at 37°C.
Plasmid construction
Mouse KLF5 complementary (c)DNA clone was purchased
from GeneCopoeia Inc. (Rockville, MD, USA). KLF5 was amplified with
PrimSTAR DNA polymerase (Takara Bio, Inc., Otsu, Japan) using
primers with methylation-sensitive EcoRI and XhoI
restriction sites. The PCR thermocycling was set as: 95°C, 3 min;
30 cycles of 95°C, 30 sec; 58°C, 30 sec; 72°C, 90 sec; 72°C, 10
min. The primer sequences of KLF5 were as follows: Forward,
5′ATGCCCACGCGGGTGCTGACC3′ and reverse, 5′ATGAAGCGCCACCAGAACTGA3′.
PCR products were then digested with EcoRI and XhoI
restriction endonucleases and subsequently inserted into the pcDNA
3.1 vector (Invitrogen; Thermo Fisher Scientific, Inc.).
Cell viability assay
Treated podocytes (2×103 cells/well) were
seeded into a 96-well plate. The cells were treated with PAN at
different concentrations (0, 5, 10, 20, 40, 60, 80 and 100 µg/ml)
and maintained at 37°C for 0, 6, 12, 24 and 48 h time intervals.
Following this, 20 µl of MTT solution was added into each well and
incubated for 30 min. A total of 150 µl dimethylsulfoxide was
subsequently was added into each well. Finally, a microplate reader
was used to determine the absorbance at 490 nm. Treatment with 60
µg/ml PAN for 6 h was selected for subsequent analysis as this
resulted in decreased cell viability (described below).
Cell transfection
Podocytes were plated in 6-well plates at a density
of 1×106 cells per well. Cells were treated with PBS or
60 µg/ml PAN for 6 h at 37°C. KLF5 overexpression (1 ug) or
negative control plasmids (pcDNA 3.1; 1 µg) were transfected into
podocytes using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. After 12 h following transfection, the cells were
used for subsequent experimentation.
RT-qPCR assay
Total RNA was obtained using TRIzol (Invitrogen;
Thermo Fisher Scientific, Inc.). RNA was reversely transcribed to
cDNA using a miScript II RT Kit (Qiagen GmbH, Hilden, Germany). The
temperature protocol or RT was 25°C for 5 min, 37°C for 60 min,
85°C for 5 min and then held at 4°C. The mRNA expression levels
were subsequently determined using the SYBR-Green PCR Master Mix
kit (Takara Bio, Inc.) and the ABI 7500 system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The thermocycling conditions were
set as: 5 min pretreatment at 95°C, followed by 28 cycles of 95°C
for 15 sec and 60°C for 30 sec, a final extension at 72°C for 10
min. The specific primers used were as follows: KLF5 forward,
5′-TTTCTGTCCCTACCCAGCAG-3′ and reverse, 5′-AGTAAGTGGCCTGTTGTGGA-3′;
Bax forward, 5′-GAGCTGCAGAGGATGATTGC-3′ and reverse,
5′-CCAATGTCCAGCCCATGATG-3′; Bcl-2 forward,
5′-GCCTTCTTTGAGTTCGGTGG-3′ and reverse, 5′-GAAATCAAACAGAGGCCGCA-3′;
Caspase-3 forward, 5′-TTGCCACCTGTCCAGTTTTG-3′ and reverse,
5′-AGGAGTGAGTGGTCTTGCTC-3′; caspase-8 forward,
5′-TTTCTGTCCCTACCCAGCAG-3′ and reverse, 5′-AGTAAGTGGCCTGTTGTGGA-3′;
Caspase-9 forward, 5′-GCCCCATATGATCGAGGACA-3′ and reverse,
5′-CAGAAACGAAGCCAGCATGT-3′; cyclin D1 forward,
5′-GCTGCTCCTGGTGAACAAGC-3′ and reverse, 5′-TTGCGTCTCAGCTCAGGGAC-3′;
and c-myc forward, 5′-CCACAGCAAACCTCCTCACA-3′ and reverse,
5′-TCCAACTTGACCCTCTTGGC-3′. GAPDH forward,
5′-GGGTCCCAGCTTAGGTTCAT-3′; GAPDH reverse,
5′-CATTCTCGGCCTTGACTGTG-3′. Data were quantified using the
2−ΔΔCq method (25).
Western blot analysis
Total proteins were prepared using a
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Shanghai, China) containing a protease inhibitor
cocktail (P8340; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
Protein concentrations were determined using a Bradford Protein
Assay kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Proteins
(30 µg) were separated via 10% SDS-PAGE gels and then transferred
onto polyvinyl difluoride (PVDF) membranes (PerkinElmer, Inc.,
Waltham, MA, USA). Following this, the membranes were blocked with
5% skimmed milk (BD Biosciences, Franklin Lakes, NJ, USA) at room
temperature for 2 h. The membranes were subsequently incubated with
the following primary antibodies overnight at 4°C: Anti-GAPDH
(1:2,000; cat. no. ab8245; Abcam, Cambridge, UK), anti-KLF5
(1:1,000; cat. no. ab24331; Abcam), anti-B cell lymphoma 2 (Bcl-2)
associated X (Bax; 1:1,000; cat. no. ab32503; Abcam), Bcl-2
(1:1,000; cat. no. ab32124; Abcam), anti-caspase-3 (1:1,500; cat.
no. ab13586; Abcam), anti-caspase-8 (1:1,500; cat. no. ab25901;
Abcam), anti-caspase-9 (1:1,500; cat. no. ab25758; Abcam),
anti-cyclin D1 (1:1,000; cat. no. ab134175; Abcam), anti-c-myc
(1:1,000; cat. no. ab39688; Abcam), anti-phosphorylated
(p)-extracellular signal-regulated protein kinase (ERK)1/2
(1:1,200; cat. no. 4370; Cell Signaling Technology, Inc., Danvers,
MA, USA), anti-ERK1/2 (1:1,000; cat. no. ab184699; Abcam),
anti-p-p38 (1:1,000; cat. no. ab47363; Abcam), anti-p38 (1:1,000;
cat. no. ab170099; Abcam). PVDF membranes were subsequently
incubated with a horseradish peroxidase-conjugated secondary
antibody (1:5,000; cat. no. sc-2004, Santa Cruz Biotechnology,
Inc., Dallas, TX, USA) at room temperature for 1 h. Finally, the
proteins were visualized using ECL Western Blotting Substrate
(Pierce; Thermo Fisher Scientific, Inc.) in an enhanced
chemiluminescence detection system (GE Healthcare, Chicago, IL,
USA). The gray value was determined by Quantity One 4.6.2 software
(Bio-Rad Laboratories, Inc.).
Immunofluorescence (IF) staining
Podocytes were fixed with 4% paraformaldehyde at 4°C
for 20 min, and then permeabilized with 0.2% Triton X-100 for 3
min. Following this, cells were washed with PBS and then blocked
using 10% goat serum (Thermo Fisher Scientific, Inc.) for 30 min at
room temperature. Cells were subsequently incubated with 4 µg/ml
anti-synaptopodin antibodies (cat. no. ab220345; Abcam) overnight
at 4°C. Following this, cells were washed using PBS and then
incubated with Alexa-Fluor® 633-conjugated secondary
antibodies (1:500; A-21070, Thermo Fisher Scientific, Inc.) at 37°C
for 1 h. Cells were subsequently stained using
4′-6-diamidino-2-phenylindole (Thermo Fisher Scientific, Inc.) for
5 min at room temperature. A fluorescence microscope
(magnification, ×200; Olympus Corporation, Tokyo, Japan) was used
to visualize the results.
Flow cytometry
Podocytes were treated as described above. Cells
(2×105 cells/well) were seeded into 24-well plates and
incubated overnight at 37°C. To investigate the cell cycle, cells
were centrifuged at 1,000 × g for 5 min at 4°C and subsequently
fixed using 70% (v/v) ethanol for 1 h at 4°C. Following this, cells
were washed with PBS and then incubated with 500 µl propidium
iodide (PI)/RNase buffer (BD Biosciences, Franklin Lakes, NJ, USA)
at room temperature for 30 min. Finally, a FACS-Calibur flow
cytometer (BD Biosciences) was used to investigate cell cycle
distribution, and ModFit LT 2.0 software (Verity Software House,
Inc., Topsham, ME, USA) was used to analyze the results. To
investigate apoptosis, cells were washed with PBS and then
re-suspended with 0.5 ml binding buffer containing 5 µl Annexin
V-fluorescein isothiocyanate and PI double stain (BD Biosciences)
for 20 min at room temperature in the dark. Following this, the
apoptotic rate was determined using a FACS-Calibur flow cytometer
(BD Biosciences) and ModFit LT 2.0 software.
Statistical analysis
All data were analyzed by SPSS 13.0 (SPSS, Inc.,
Chicago, IL, USA) using one-way analysis of variance with Tukey's
test. Data are presented as the mean ± standard deviation. All
experiments were performed in triplicate. P<0.05 was considered
to indicate a statistically significant difference.
Results
PAN inhibits the proliferation of
podocytes
Initially, synaptopodin expression in podocytes was
investigated using immunofluorescence. The results revealed that
the expression of synaptopodin was positive in podocytes (Fig. 1A). To further investigate the
effect of PAN on the proliferation ability of podocytes, podocytes
were treated with PBS (control) or PAN (5, 10, 20, 40, 60, 80, and
100 µg/ml) for 0, 6, 12, 24 and 48 h time intervals. The results
revealed that the viability of podocytes were markedly inhibited
following treatment with PAN in a dose- and time-dependent manner
(Fig. 1B; P<0.05). It was
observed that the cell viability significantly decreased when the
cells were treated with 60 µg/ml PAN for 6 h. Therefore, the
present study selected 60 µg/ml PAN to treat the cells for 6 h.
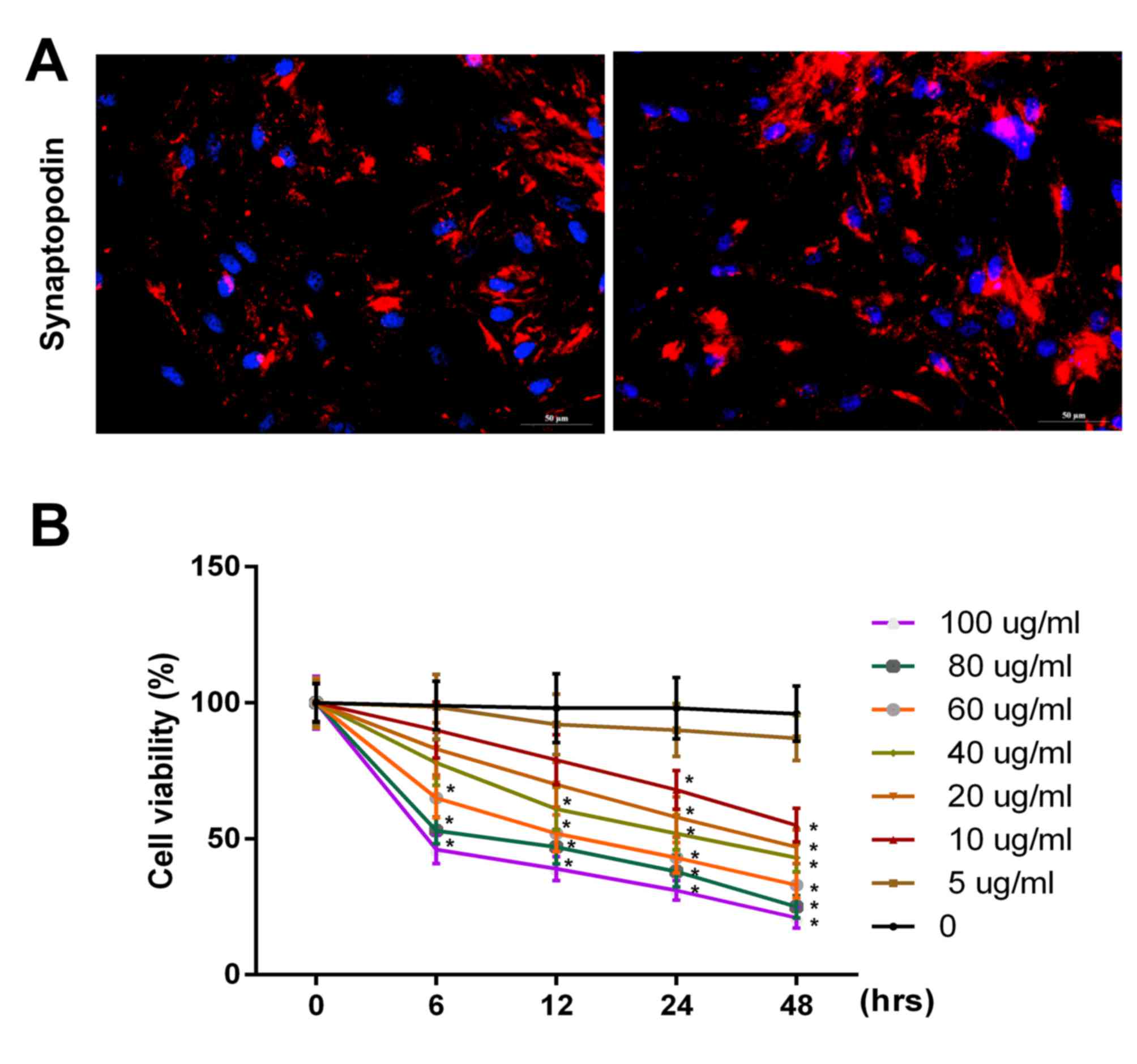 | Figure 1.PAN inhibited the proliferative
ability of podocytes in a dose- and time-dependent manner. (A)
Synaptopodin expression in podocytes was detected via
immunofluorescence assay using a fluorescence microscope
(magnification, ×200). (B) Cell viability in podocytes was analyzed
using an MTT assay following g treatment with PBS (control), or PAN
(5, 10, 20, 40, 60, 80 and 100 µg/ml) for 0, 6, 12, 24 and 48 h
time intervals. PAN, puromycin aminonucleoside. *P<0.05 vs. 0
h. |
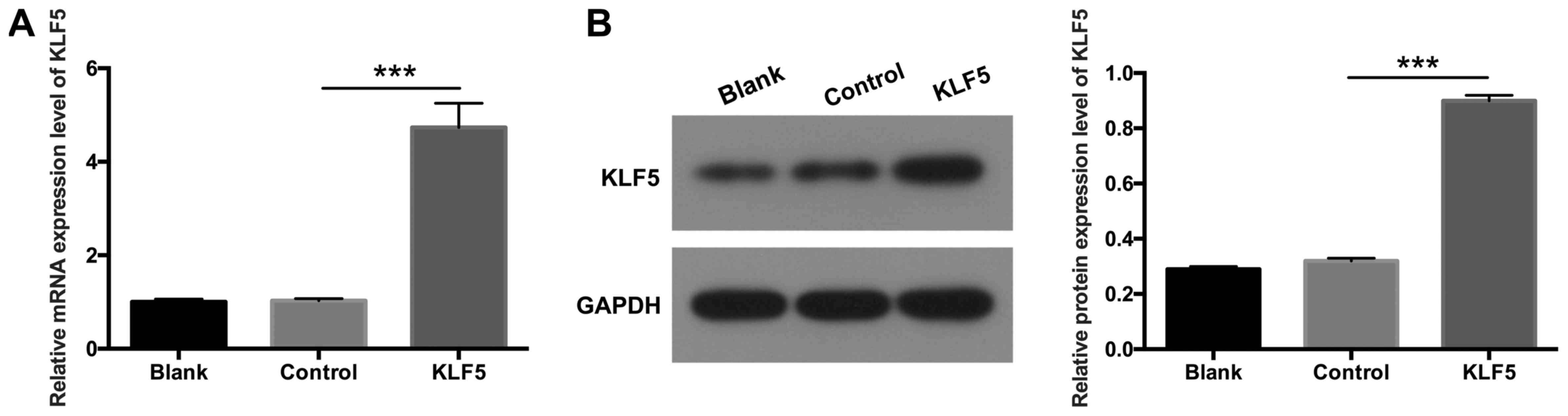
KLF5 is overexpressed in transfected
podocytes
To further investigate the potential role of KLF5 in
podocytes, KLF5 was overexpressed in podocytes via transfection
with pCDNA3.1-KLF5 plasmids. RT-qPCR and western blot assays were
performed to determine the expression levels of KLF5. The results
demonstrated that KLF5 expression was significantly increased in
podocytes transfected with pCDNA3.1-KLF5 compared with the NC group
(Fig. 2; P<0.001), which
suggested that transfection to establish the podocyte injury model
was successful.
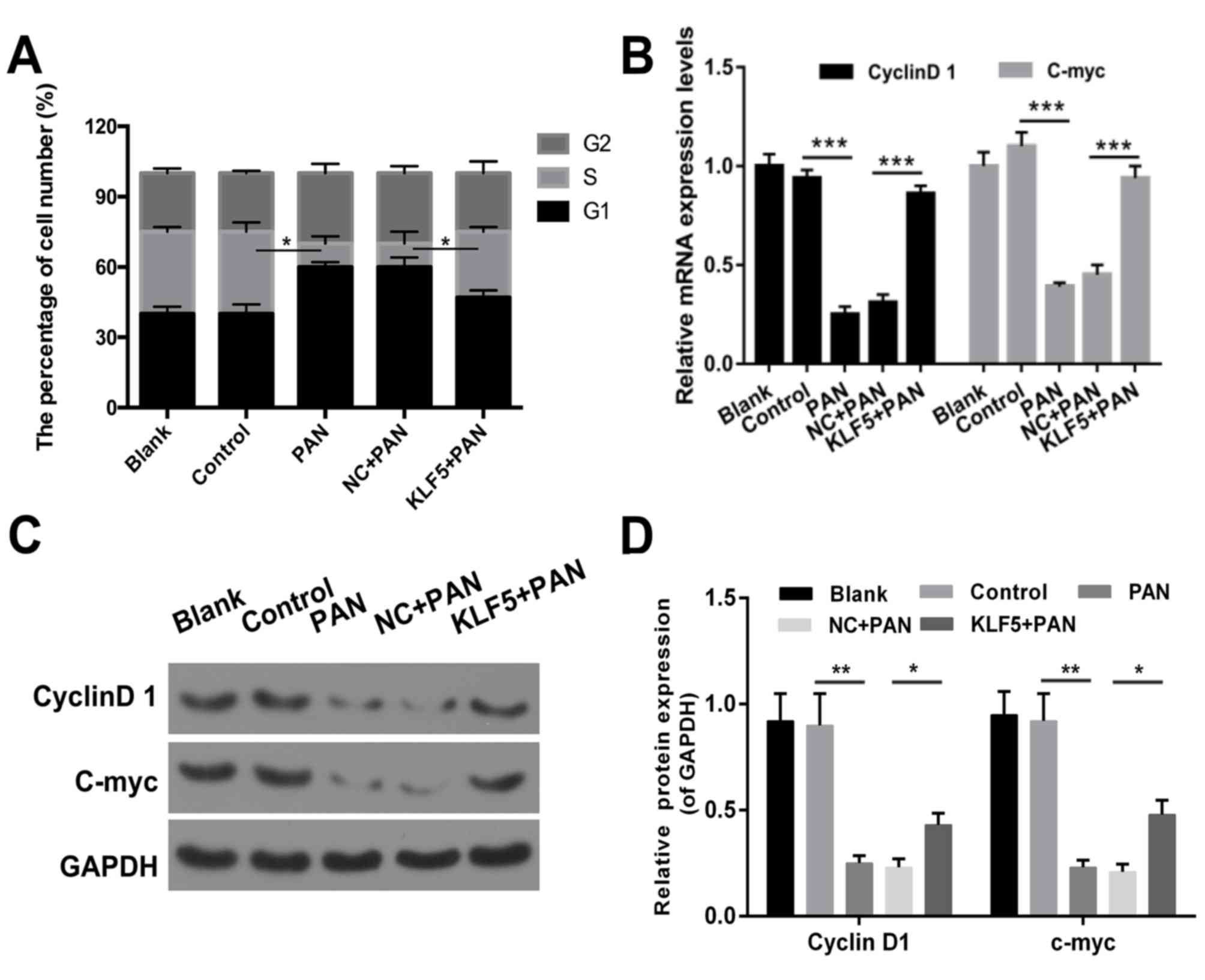
Overexpression of KLF5 inhibits cell
cycle arrest of PAN-treated podocytes
The results revealed that the proportion of cells in
the G1 phase was increased in the PAN group compared with the
control group (Fig. 3A). The
number of cells in the G1 phase was decreased in the KLF5+PAN group
compared with the NC+PAN group (Fig.
3A, P<0.05). In addition, the results demonstrated that
treatment with PAN promoted cell cycle arrest in podocytes
(P<0.05); however, this effect was markedly attenuated following
overexpression of KLF5 (Fig. 3A,
P<0.05). In addition, the expression levels of cyclin D1 and
c-myc were investigated via RT-qPCR and western blot assays, and
the results revealed that cells treated with PAN exhibited
significantly decreased expression levels of cyclin D1 and c-myc
compared with the NC group (P<0.01), whereas cells
overexpressing KLF5 exhibited significantly increased expression
levels of cyclin D1 and c-myc compared with the NC + PAN group
(Fig. 3B-D, P<0.05).
Overexpression of KLF5 suppresses the
apoptosis of PAN-treated podocytes
Flow cytometry was performed to determine whether
KLF5 has an important role in the apoptosis of podocytes. The
results demonstrated that the apoptosis of podocytes was
significantly increased in the PAN group compared with the NC group
(P<0.001; Fig. 4A); however,
this effect was significantly attenuated following overexpression
of KLF5 (P<0.05; Fig. 4A).
Furthermore, the expression levels of genes associated with
apoptosis (Bax, Bcl-2, caspase-3, caspase-8 and caspase-9) were
investigated using RT-qPCR and western blot assays. The results
revealed that treatment with PAN significantly enhanced the
expression levels of Bax, caspase-3, caspase-8 and caspase-9
exhibited by podocytes (P<0.01 and P<0.001; Fig. 4B and C); however, overexpression of
KLF5 significantly attenuated this effect (P<0.05 and P<0.01;
Fig. 4B and C). In addition,
podocytes treated with PAN exhibited significantly decreased Bcl-2
expression, and overexpression of KLF5 significantly attenuated
this effect (P<0.05, P<0.01 and P<0.001; Fig. 4B and C).
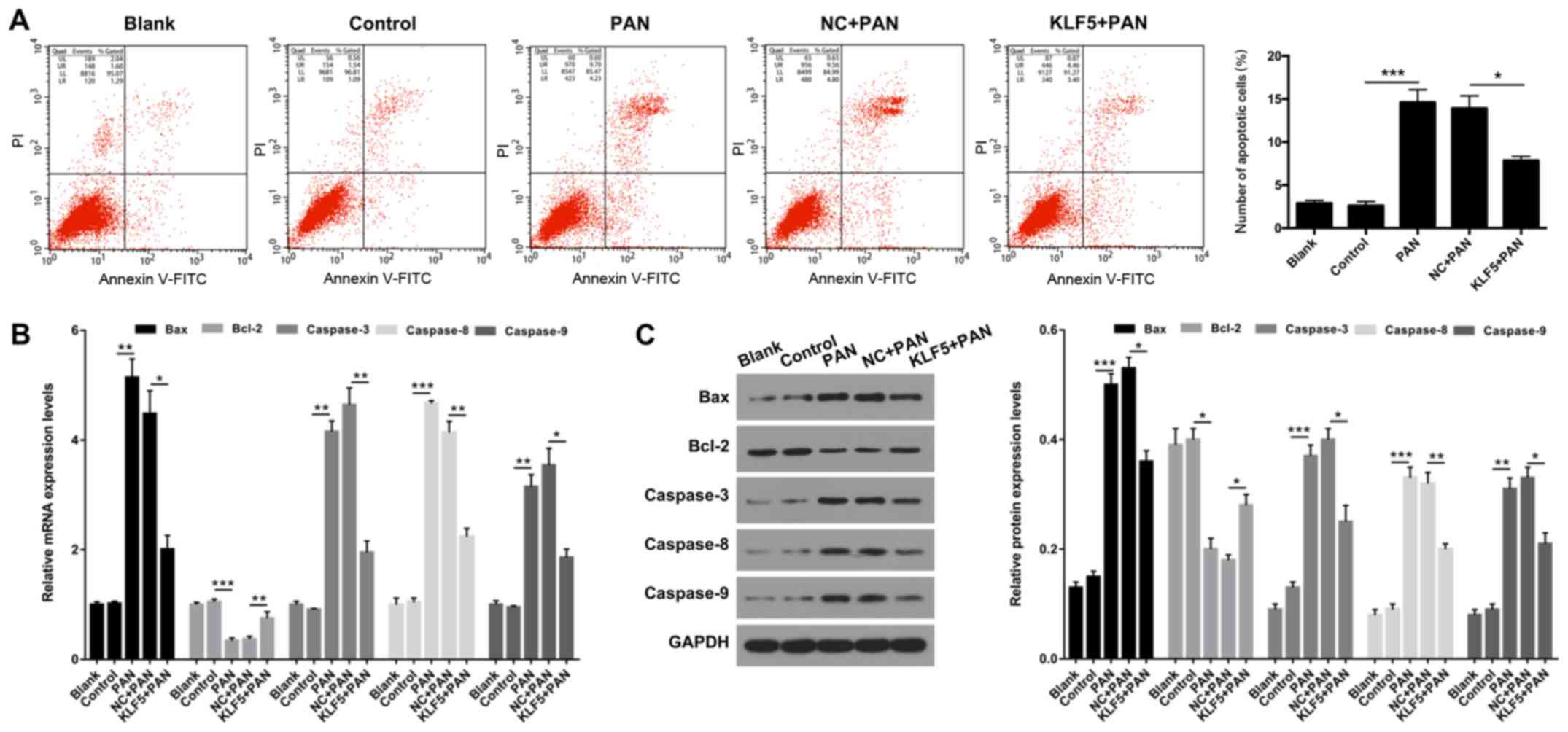 | Figure 4.Overexpression of KLF5 inhibited
PAN-induced apoptosis of podocytes. Podocytes were treated with PBS
(control), PAN (60 µg/ml), NC + PAN or KLF5 + PAN. (A) Cell
apoptosis was investigated using flow cytometry in treated
podocytes, and the number of apoptotic cells were quantitatively
analyzed. (B) mRNA expression levels of Bax, Bcl-2, caspase-3,
caspase-8 and caspase-9 were determined via by reverse
transcription-quantitative polymerase chain reaction assays. (C)
Protein expression levels of Bax, Bcl-2, caspase-3, caspase-8 and
caspase-9 were investigated using western blot analyses, and
quantitative analysis was performed using base gray values and
GAPDH. *P<0.05, **P<0.01 and ***P<0.001. PAN, puromycin
aminonucleoside; KLF5, Krüppel-like factor 5; NC, negative control;
Bcl-2, B cell lymphoma 2; Bax, Bcl-2 associated X; PI, propidium
iodide; FITC, fluorescein isothiocyanate. |
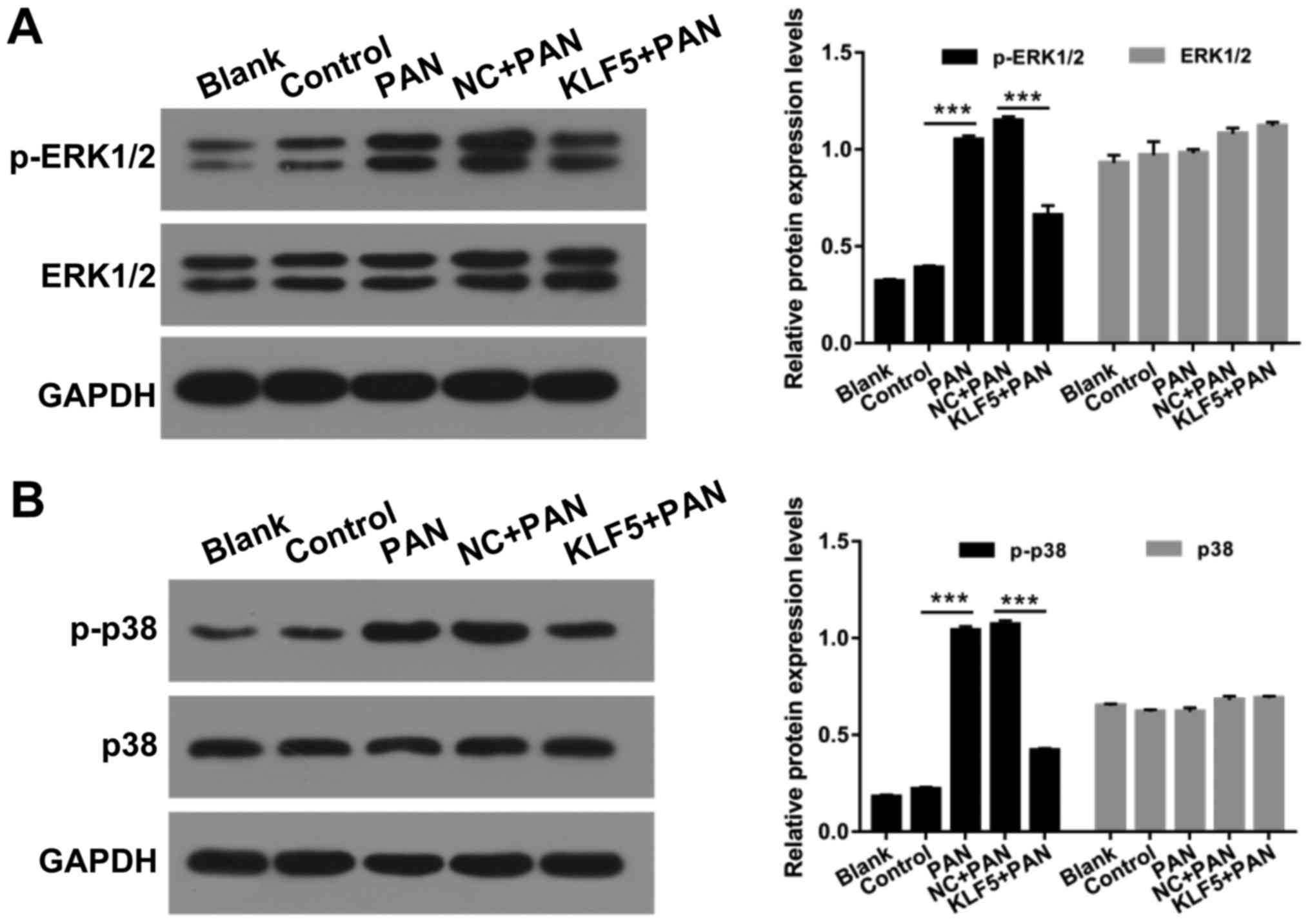
Overexpression of KLF5 suppresses the
activation of ERK/p38 mitogen-activated protein (MAP) kinase
pathway in PAN-treated podocytes
MAP kinases have important roles in numerous
cellular functions and are regulated via independent upstream
activation cascades (26). In the
present study, the effects of KLF5 and PAN on the ERK/p38 MAP
kinase pathway in podocytes were investigated. The protein levels
of p-ERK1/2, ERK1/2, p-p38 and p38 in podocytes were investigated
using a western blot assay. The results revealed that treatment
with PAN significantly enhanced ERK1/2 and p38 phosphorylation
compared with the control group; however, overexpression of KLF5
significantly attenuated these effects (P<0.001; Fig. 5).
Discussion
DN is one of the most common complications
associated with diabetes and has become the most frequent causative
factor resulting in the development of end-stage renal disease
(27). The pathogenesis of DN
arises from glucose and lipid metabolic disorders, abnormal
hemodynamics and oxidative stress (28,29).
The results of the present study demonstrated that podocyte injury
is an important factor resulting in the development of proteinuria
and glomerular sclerosis, and has significant effects on the
pathogenesis of DN (30).
Considering that synaptopodin is a podocyte marker (31), the expression levels of
synaptopodin in podocytes was investigated in the present study,
and the results revealed that synaptopodin was highly expressed in
podocytes.
Podocytes and the slit diaphragm within foot
processes are critical components of the glomerular filtration
barrier (32). Associated
molecules, including nephrin, podocin, CD2 associated protein and
α-actinin-4 have important roles in the development of proteinuria
(33). A human podocyte cell line
was used in the present study to investigate podocyte injury in
vitro. Furthermore, the present study established a podocyte
injury model in vitro via administration of PAN. This model
provided the possibility to further investigate molecular
mechanisms associated with DN (34–36).
The results revealed that treatment with PAN markedly inhibited the
proliferation of podocytes in a dose- and time-dependent
manner.
KLF-5 is an important member of the KLF protein
family, and is located at chromosome 13q21 and encodes a 55 kDa
protein that contains 457 amino acids (37). In normal tissue, KLF5 extensively
regulates numerous cellular processes, such as proliferation,
differentiation, movement, inflammation and pluripotency (38,39).
However, the role of KLF5 in DN remains unclear. In the present
study, the results revealed that PAN induced cell cycle arrest in
podocytes; however, overexpression of KLF5 significantly attenuated
this effect via upregulation of cyclin D1 and c-myc expression
levels. In addition, a previous study demonstrated that KLF5
inhibited the cell cycle progression of vascular smooth muscle
cells via activation of cyclin D1 (40). Furthermore, overexpression of KLF5
was revealed to significantly suppress PAN-mediated apoptosis of
podocytes. At a molecular level, the results of the present study
revealed that PAN enhanced the expression levels of Bax, caspase-3,
caspase-8 and caspase-9; however, treatment with PAN significantly
reduced Bcl-2 expression. Furthermore, overexpression of KLF5
significantly decreased Bax, caspase-3, caspase-8 and caspase-9
expression levels, whereas overexpression of KLF5 significantly
increased Bcl-2 expression in PAN-treated podocytes. A previous
study revealed that treatment with PAN induces apoptosis of
glomerular podocytes (41). A
further study demonstrated that the downregulation of KLF5 is
associated with G1 phase cell cycle arrest (42). The results of the present study
were therefore consistent with previous studies. Therefore, it was
concluded that overexpression of KLF5 alleviates PAN-mediated
podocyte injury.
MAP kinases are serine/threonine-specific protein
kinases (43–45). p38 MAP kinase and ERK are factors
of the MAP kinase protein family (46,47).
The results of the present study suggested that treatment with PAN
induced the phosphorylation of ERK1/2 and p38. Overexpression of
KLF5 suppressed the phosphorylation of ERK/p38 MAP kinase in
PAN-treated podocytes. These results suggested that overexpression
of KLF5 protected podocytes from injury via inhibited activation of
the ERK1/2 and p38 pathways. However, a previous study demonstrated
that KLF5 may promote the activation of ERK1/2 in breast cancer
cells (48). This discrepancy may
be due to the use of different cells.
Limitations of the present study included that all
experiments were performed in vitro, and that the exact
mechanism underlying the regulation of KLF5 remains unclear. Thus,
future studies should perform further in vivo investigations
to determine the role of KLF5 in DN. In conclusion, the results of
the present study demonstrated that PAN inhibited the proliferation
of podocytes, and that overexpression of KLF5 attenuated
PAN-induced cell cycle arrest and apoptosis of podocytes.
Furthermore, the results demonstrated that overexpression of KLF5
suppressed the ERK/p38 MAP kinase pathway in PAN-treated podocytes.
The present study revealed that a potential therapeutic strategy
for the treatment of DN may comprise the upregulation of KLF5
expression.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Fund project in Shandong Province (grant no.
2017GSF21116).
Availability of data and materials
All data generated and/or analyzed during this study
are included in this published article.
Authors' contributions
YL wrote the main manuscript. YL, XH and XS
performed the experiments. YL and ZH designed the study. XS and XH
performed data analysis. YL, XS and ZH contributed to manuscript
revisions. All authors reviewed the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bikbova G, Oshitari T, Tawada A and
Yamamoto S: Corneal changes in diabetes mellitus. Curr Diabetes
Rev. 8:294–302. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bril V: Neuromuscular complications of
diabetes mellitus. Continuum (Minneap Minn) 20 (3 Neurology of
Systemic Disease). 1–544. 2014.
|
|
3
|
Nentwich MM and Ulbig MW: Diabetic
retinopathy-ocular complications of diabetes mellitus. World J
Diabetes. 6:489–499. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kaul K, Tarr JM, Ahmad SI, Kohner EM and
Chibber R: Introduction to diabetes mellitus. Adv Exp Med Biol.
771:1–11. 2012.PubMed/NCBI
|
|
5
|
Brinks R and Rathmann W: Response to
Monesi etal Prevalence, incidence and mortality of diagnosed
diabetes: Evidence from an Italian population-based study. Diabet
Med. 29:1085–1086; author reply 1085. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Farag YM and Gaballa MR: Diabesity: An
overview of a rising epidemic. Nephrol Dial Transplant. 26:28–35.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chao CT, Huang JW, Chiang CK, Chen YC,
Fang CC, Hu FC, Chang CC and Yen CJ: Diabetes mellitus, superoxide
dismutase and peroxisome proliferator activated receptor gamma
polymorphisms modify the outcome of end-stage renal disease
patients of Han Chinese origin. Nephrology (Carlton). 23:117–125.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tagawa A, Yasuda M, Kume S, Yamahara K,
Nakazawa J, Chin-Kanasaki M, Araki H, Araki S, Koya D, Asanuma K,
et al: Impaired podocyte autophagy exacerbates proteinuria in
diabetic nephropathy. Diabetes. 65:755–767. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yasuda-Yamahara M, Kume S, Tagawa A,
Maegawa H and Uzu T: Emerging role of podocyte autophagy in the
progression of diabetic nephropathy. Autophagy. 11:2385–2386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kamiyama M, Urushihara M, Morikawa T,
Konishi Y, Imanishi M, Nishiyama A and Kobori H: Oxidative
stress/angiotensinogen/renin-angiotensin system axis in patients
with diabetic nephropathy. Int J Mol Sci. 14:23045–23062. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Weening JJ and Rennke HG: Glomerular
permeability and polyanion in adriamycin nephrosis in the rat.
Kidney Int. 24:152–159. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Amann K, Nichols C, Tornig J, Schwarz U,
Zeier M, Mall G and Ritz E: Effect of ramipril, nifedipine, and
moxonidine on glomerular morphology and podocyte structure in
experimental renal failure. Nephrol Dial Transplant. 11:1003–1011.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mifsud SA, Allen TJ, Bertram JF, Hulthen
UL, Kelly DJ, Cooper ME, Wilkinson-Berka JL and Gilbert RE:
Podocyte foot process broadening in experimental diabetic
nephropathy: Amelioration with renin-angiotensin blockade.
Diabetologia. 44:878–882. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim D, Lim S, Park M, Choi J, Kim J, Han
H, Yoon K, Kim K, Lim J and Park S: Ubiquitination-dependent CARM1
degradation facilitates Notch1-mediated podocyte apoptosis in
diabetic nephropathy. Cell Signal. 26:1774–1782. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Courboulin A, Tremblay VL, Barrier M,
Meloche J, Jacob MH, Chapolard M, Bisserier M, Paulin R, Lambert C,
Provencher S and Bonnet S: Krüppel-like Factor 5 contributes to
pulmonary artery smooth muscle proliferation and resistance to
apoptosis in human pulmonary arterial hypertension. Respir Res.
12:1282011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tetreault MP, Yang Y and Katz JP:
Krüppel-like factors in cancer. Nat Rev Cancer. 13:701–713. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Courboulin A, Tremblay VL, Barrier M,
Meloche J, Jacob MH, Chapolard M, Bisserier M, Paulin R, Lambert C,
Provencher S and Bonnet S: Krüppel-like factor 5 contributes to
pulmonary artery smooth muscle proliferation and resistance to
apoptosis in human pulmonary arterial hypertension. Respir Res.
12:1282011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Limame R, Op de Beeck K, Lardon F, De
Wever O and Pauwels P: Krüppel-like factors in cancer progression:
Three fingers on the steering wheel. Oncotarget. 5:29–48. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yin KJ, Hamblin M, Fan Y, Zhang J and Chen
YE: Krüppel-like factors in the central nervous system: Novel
mediators in stroke. Metab Brain Dis. 30:401–410. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen C, Zhou Z, Guo P and Dong JT:
Proteasomal degradation of the KLF5 transcription factor through a
ubiquitin-independent pathway. FEBS Lett. 581:1124–1130. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong JT and Chen C: Essential role of KLF5
transcription factor in cell proliferation and differentiation and
its implications for human diseases. Cell Mol Life Sci.
66:2691–2706. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Agardh E, Lundstig A, Perfilyev A, Volkov
P, Freiburghaus T, Lindholm E, Rönn T, Agardh CD and Ling C:
Genome-wide analysis of DNA methylation in subjects with type 1
diabetes identifies epigenetic modifications associated with
proliferative diabetic retinopathy. BMC Med. 13:1822015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kakimoto T, Okada K, Fujitaka K, Nishio M,
Kato T, Fukunari A and Utsumi H: Quantitative analysis of markers
of podocyte injury in the rat puromycin aminonucleoside nephropathy
model. Exp Toxicol Pathol. 67:171–177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Trachtman H, Del Pizzo R, Futterweit S,
Levine D, Rao PS, Valderrama E and Sturman JA: Taurine attenuates
renal disease in chronic puromycin aminonucleoside nephropathy. Am
J Physiol. 262:F117–F123. 1992.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Baĭramov RB and Abdullaeva RT: The impact
of early gastric cancer diagnosis on indices of survival in
patients after radical surgical intervention. Klin Khir. 1–21.
2013.
|
|
27
|
Krolewski AS, Skupien J, Rossing P and
Warram JH: Fast renal decline to end-stage renal disease: An
unrecognized feature of nephropathy in diabetes. Kidney Int.
91:1300–1311. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kowluru RA and Mishra M: Oxidative stress,
mitochondrial damage and diabetic retinopathy. Biochim Biophys
Acta. 1852:2474–2483. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Aoki Y, Yazaki K, Shirotori K, Yanagisawa
Y, Oguchi H, Kiyosawa K and Furuta S: Stiffening of connective
tissue in elderly diabetic patients: Relevance to diabetic
nephropathy and oxidative stress. Diabetologia. 36:79–83. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yasuno K, Kamiie J and Shirota K: Analysis
of ultrastructural glomerular basement membrane lesions and
podocytes associated with proteinuria and sclerosis in
Osborne-Mendel rats with progressive glomerulonephropathy. J Vet
Sci. 14:223–226. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yu H, Kistler A, Faridi MH, Meyer JO,
Tryniszewska B, Mehta D, Yue L, Dryer S and Reiser J: Synaptopodin
limits TRPC6 podocyte surface expression and attenuates
proteinuria. J Am Soc Nephrol. 27:3308–3319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kriz W and Lemley KV: Potential relevance
of shear stress for slit diaphragm and podocyte function. Kidney
Int. 91:1283–1286. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cara-Fuentes G, Clapp WL, Johnson RJ and
Garin EH: Pathogenesis of proteinuria in idiopathic minimal change
disease: Molecular mechanisms. Pediatr Nephrol. 31:2179–2189. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li X, Zhang X, Li X, Ding F and Ding J:
The role of survivin in podocyte injury induced by puromycin
aminonucleoside. Int J Mol Sci. 15:6657–6673. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu SY and Qi R: Role of bad in podocyte
apoptosis induced by puromycin aminonucleoside. Transplant Proc.
45:569–573. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zennaro C, Rastaldi MP, Pascolo L, Stebel
M, Trevisan E, Artero M, Tiribelli C, Di Maso V and Carraro M:
Podocyte expression of membrane transporters involved in puromycin
aminonucleoside-mediated injury. PLoS One. 8:e661592013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Marrero-Rodríguez D, Taniguchi-Ponciano K,
Jimenez-Vega F, Romero-Morelos P, Mendoza-Rodríguez M, Mantilla A,
Rodriguez-Esquivel M, Hernandez D, Hernandez A, Gomez-Gutierrez G,
et al: Krüppel-like factor 5 as potential molecular marker in
cervical cancer and the KLF family profile expression. Tumour Biol.
35:11399–11407. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sousa MI, Rodrigues AS, Pereira S,
Perestrelo T, Correia M and Ramalho-Santos J: Mitochondrial
mechanisms of metabolic reprogramming in proliferating cells. Curr
Med Chem. 22:2493–2504. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ci X, Xing C, Zhang B, Zhang Z, Ni JJ,
Zhou W and Dong JT: KLF5 inhibits angiogenesis in PTEN-deficient
prostate cancer by attenuating AKT activation and subsequent HIF1α
accumulation. Mol Cancer. 14:912015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Amirak E, Zakkar M, Evans PC and Kemp PR:
Perfusion of veins at arterial pressure increases the expression of
KLF5 and cell cycle genes in smooth muscle cells. Biochem Biophys
Res Commun. 391:818–823. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xiao H, Shi W, Liu S, Wang W, Zhang B,
Zhang Y, Xu L, Liang X and Liang Y: 1,25-Dihydroxyvitamin D(3)
prevents puromycin aminonucleoside-induced apoptosis of glomerular
podocytes by activating the phosphatidylinositol
3-kinase/Akt-signaling pathway. Am J Nephrol. 30:34–43. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Qin WW, Zhang R, Chen RA, Li GH, Ji YR,
Liu L and Wang T: MicroRNA-145 induces cell cycle arrest in G1
phase by directly targeting KLF5 in colon cancer. Int J Clin Exp
Pathol. 9:5197–5209. 2016.
|
|
43
|
Gaestel M: MAPK-activated protein kinases
(MKs): Novel insights and challenges. Front Cell Dev Biol.
3:882016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hettenhausen C, Schuman MC and Wu J: MAPK
signaling: A key element in plant defense response to insects.
Insect Sci. 22:157–164. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
O'Callaghan C, Fanning LJ and Barry OP:
p38δ MAPK: Emerging roles of a neglected isoform. Int J Cell Biol.
2014:2726892014.PubMed/NCBI
|
|
46
|
Zhang Q, Wang J, Duan MT, Han SP, Zeng XY
and Wang JY: NF-κB, ERK, p38 MAPK and JNK contribute to the
initiation and/or maintenance of mechanical allodynia induced by
tumor necrosis factor-alpha in the red nucleus. Brain Res Bull.
99:132–139. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cuadrado A and Nebreda AR: Mechanisms and
functions of p38 MAPK signalling. Biochem J. 429:403–417. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu R, Zheng HQ, Zhou Z, Dong JT and Chen
C: KLF5 promotes breast cell survival partially through fibroblast
growth factor-binding protein 1-pERK-mediated dual specificity
MKP-1 protein phosphorylation and stabilization. J Biol Chem.
284:16791–16798. 2009. View Article : Google Scholar : PubMed/NCBI
|