Introduction
Liposarcoma is the most frequently occurring
soft-tissue tumor and accounts for >20% of all soft tissue
sarcomas (1,2). Liposarcoma has been classified into
four major histological sub-types: Well-differentiated liposarcoma,
dedifferentiated liposarcoma, myxoid liposarcoma and pleomorphic
liposarcoma (3). The 5-year
survival rate is affected by different sub-types, with 75–100% in
differentiated tumors. However, pleomorphic liposarcoma has the
worst 5-year survival rate of 0–20% (4). In clinical practice, surgical
approaches are first-line therapy in the management of liposarcoma,
accompanied with radiotherapy and chemotherapy (5). Currently, there is no therapeutic
option for aggressive and metastatic liposarcoma (5). Therefore, exploring novel and
specific therapeutic targets for liposarcoma is important.
Perilipin-1 (PLIN1) is a member of the
perilipin/APRP/TIP47 (PAT) protein family and exists in at least 3
protein isoforms (molecular weight ~56 kDa) (6). PLIN1 is a biomarker of adipocyte
differentiation and involved in regulating lipid droplet biogenesis
and hydrolysis (7). In db/db mice,
PLIN1 loss-of-function accelerates lipolysis and decreases the size
of lipid droplets in adipocytes, causing resistance to diet-induced
obesity (8). In addition,
downregulation of PLIN1 expression is also associated with the
acceleration of lipolysis in 3T3-L1 adipocytes (9). In humans, the expression of PLIN1 in
adipose tissues is positively correlated with insulin sensitivity
(10). Previous studies have
revealed that PLIN1 can regulate spermatogenesis (11), atherosclerotic lesions (12) and osteogenic differentiation
(13). Notably, PLIN1 is known to
be an independent poor prognostic factor in metastatic breast
cancer and can inhibit cell proliferation, migration, invasion and
tumorigenesis in mice (7,14). Only one study, to the best of the
authors' knowledge, has examined the expression of PLIN1 in
liposarcoma (6) but the
association between PLIN1 and cell proliferation, migration and
apoptosis has not been investigated in vitro.
The present study examined the expression of PLIN1
in liposarcoma tissues and identified that PLIN1 was significantly
overexpressed. In addition, PLIN1 levels in liposarcoma cell lines
were downregulated by short hairpin RNA (shRNA) and cell
proliferation, migration and apoptosis experiments were performed
to elucidate its function in vitro.
Materials and methods
Patients and specimens
A total of 12 pairs of liposarcoma tumor tissues and
matched adjacent non-tumorous adipose tissues were collected from
patients (8 males and 4 females; age range: 15–67 years; mean age:
48.8±14.0 years) who had undergone surgery at the Department of
Oncology, People's Hospital of Rizhao (Rizhao, China) between
January 2014 and June 2016. None of the patients had been subjected
to preoperative radiotherapy or chemotherapy and all had been
diagnosed with liposarcoma based on histopathological evaluation.
All collected tissue samples were immediately stored in liquid
nitrogen. Human samples were obtained with written informed consent
from all patients. The study was approved by the Ethics Committee
of the People's Hospital of Rizhao (Approval Number:
KYLL-20140903).
Cell culture
T778 (well differentiated) and LPS141
(dedifferentiated) liposarcoma cell lines were purchased from the
Cell Bank of China Academy of Sciences (Shanghai, China). The cell
lines were cultured in RPMI medium 1640 (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin at 37°C in 5% CO2 and were
plated in a 6-well plate at a density of 2×105 cells per
well. Following incubation for 2 days, cells were collected for
proliferation, migration and apoptosis assays.
Cell counting kit (CCK)-8
proliferation assay
The CCK-8 proliferation assay was conducted as
previously described (15). The
proliferation of cells (1×104) was measured using a
CCK-8 Cell Proliferation/Viability Assay kit (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan).
Construction of shRNA-PLIN1 eukaryotic
expression plasmid
shRNA was constructed to specifically target PLIN1
using shRNA design tools (http://rnaidesigner.thermofisher.com/rnaiexpress/).
Using BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi), it was
verified that the designed shRNA targeted only the PLIN1. The shRNA
was synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China).
The target site in human gene encoding PLIN1 was: shRNA-PLIN1 sense
strand, 5′-GGGGACACAGTGTGCATTA-3′, shRNA-PLIN1 antisense strand,
5′-TGGCACATACCCTGCAGAAGA-3′; shRNA-Con sense strand,
5′-GCCCTCGGTGTCCTACTTCA-3′, shRNA-Con antisense strand,
5′-ATTTGAAGTAGGACACCGAGG-3′. Each DNA was used to transform the
E. coli strain DH5α and purified with a plasmid purification
kit (Qiagen, Inc., Valencia, CA, USA). A total of 1 µg plasmid was
transfected into T778 and LPS141 cells using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientifc, Inc.) for 48 h at 37°C, as previously described
(16). The infected cells were
selected with puromycin (2 mg/ml) 48 h following transfection. Then
the clones were selected and cultured for further
experimentation.
Immunohistochemical staining and
western blotting
The tissues were fixed in 4% formaldehyde at room
temperature for 24 h and embedded in paraffin. The
paraffin-embedded tumor tissues and adipose tissues were cut into
~3 µm sections, which were dehydrated at 65°C for 24 h,
deparaffinized in xylene and rehydrated in a descending ethanol
series. Subsequently, the sections were evaluated
immunohistochemically using anti-human PLIN1 (cat. no. ab61682,
dilution, 1:100; Abcam, Cambridge, UK), as previously described
(17). Image visualization was
performed under a microscope (Leica DM 2500; Leica Microsystems
GmbH, Wetzlar, Germany) and the integrated optical density (IOD)
was measured by Image Pro-Plus 6 software (Media Cybernetics, Inc.,
Rockville, MD, USA).
Protein was extracted using radioimmunoprecipitation
assay lysis buffer (Beyotime Institute of Biotechnology, Haimen,
China). The concentration was determined using the bicinchoninic
acid kit for protein determination (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Samples containing 50 µg of protein were
separated by 10% SDS-PAGE gel and transferred onto nitrocellulose
membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
membranes were blocked with 10% (w/v) non-fat dry milk in
Tris-buffered saline containing 0.1% (w/v) Tween 20 at room
temperature for 2 h. Primary antibody PLIN1 (cat. no. ab61682,
dilution, 1:1,000; Abcam, Cambridge, UK) was incubated for 2 h at
room temperature and membranes were washed three times (10
min/wash) in PBS. Next, the membranes were incubated with the
appropriate horseradish peroxidase-conjugated secondary antibody
(cat. no. sc-516102; dilution: 1:10,000; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA) at room temperature for 2 h, and then
visualized using SuperSignal™ West Pico PLUS Chemiluminescent
Substrate (cat. no. 34580; Thermo Fisher Scientific, Inc.).
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH; cat. no. 2118;
dilution: 1:2,000; Cell Signaling Technology, Inc., Danvers, MA,
USA) was used as the control antibody. Signals were
densitometrically assessed using Quantity One software version 4.5
(Bio Rad Laboratories, Inc.).
Wound healing assay
T778 and LPS141 cells (2×105) were
trypsinized and reseeded in each well of a new 6-well plate.
Following 24 h incubation at 37°C in 5% CO2, the
confluent cell monolayers were scratched with a 10 µl sterile
pipette tip. Then the non-adherent cells were washed off with
sterilized PBS and serum-free medium was added to the wells. The
gap area caused by the scratch was observed using an inverted
microscope (Olympus Corporation, Tokyo, Japan). Images were
captured from 3 random non-overlapping areas in each well at 12 h
post-scratch. The scratch width between the two linear regions was
measured to assess the capacity of cells to migrate.
Flow cytometry for the detection of
cell cycle progression and apoptosis
T778 and LPS141 cells were collected following
digestion and were washed twice with PBS and centrifuged at 1,200 ×
g for 5 min at 4°C. The supernatant was discarded and the cells
resuspended and fixed in ice-cold 75% ethanol then stored at 4°C.
Following two washes in PBS, the cells were stained with propidium
iodide (PI; cat. no. F10797; Invitrogen; Thermo Fisher Scientific,
Inc.) at 4°C for 30 min in the dark and acquired on a flow
cytometer. Flow cytometric analysis of the percentage of cells in
G1, S and G2 phases (BD Biosciences, Franklin
Lakes, NJ, USA) was performed. The data were processed by Cell
Quest Software (version 5.1, BD Biosciences). The cell apoptosis
assay was determined as previously described (15). The Annexin V-FITC apoptosis
detection kit was purchased from Invitrogen (Thermo Fisher
Scientific, Inc.).
Statistical analysis
The experiments were repeated three times and the
data are presented as the means ± standard deviation for each
group. All statistical analyses were performed using PRISM version
7.0 (GraphPad Software, Inc., La Jolla, CA, USA). Student's t-test
was used to analyze two-group differences. Inter-group differences
were analyzed by one-way analysis of variance, followed by a post
hoc Tukey's test for multiple comparisons. P<0.05 was considered
to indicate a statistically significant difference.
Results
PLIN1 is overexpressed in liposarcoma
tissues compared with normal adipose tissue
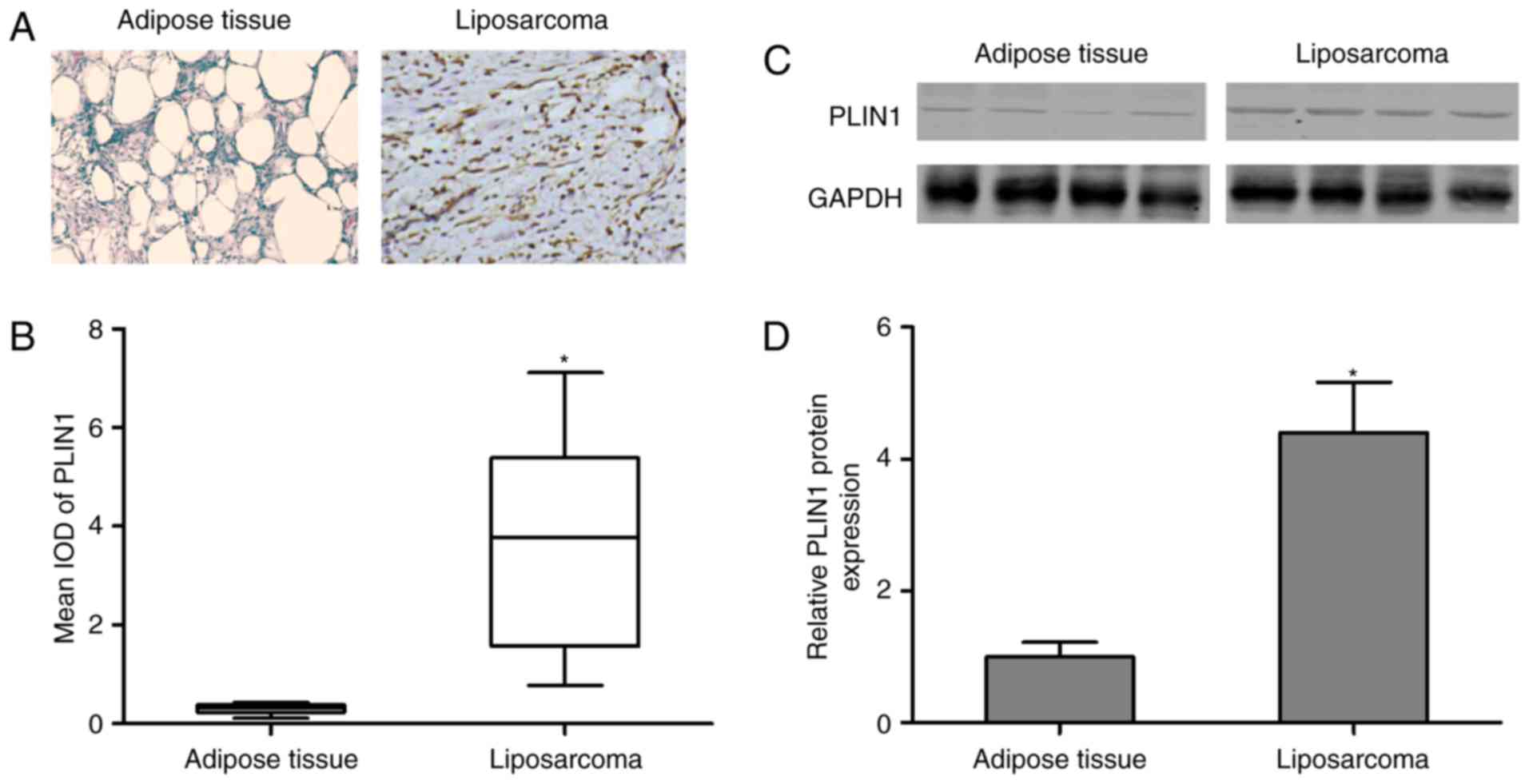
Immunohistochemical staining was performed in normal
control and liposarcoma tumor tissues, and the results demonstrated
that PLIN1 expression increased in liposarcoma tumor tissues,
revealing a statistically significant difference compared with
normal control (Fig. 1A and B). To
further confirm PLIN1 expression, western blotting analysis was
performed and identified that PLIN1 protein expression was
significantly upregulated in liposarcoma tumor tissues compared
with the control group (Fig. 1C and
D). These findings validated the hypothesis that PLIN1 is
closely associated with liposarcoma.
Knockdown of PLIN1 inhibits
proliferation of liposarcoma cells
To investigate its function in vitro, shRNA
to inhibit the levels of PLIN1 was designed. As illustrated in
Fig. 2A, the levels of PLIN1 were
significantly inhibited by shRNA-PLIN1 in T778 and LPS141 cells
compared with the negative control (Negative-Con) or shRNA-Con
group. No significant difference was observed in the levels of
PLIN1 between the Negative-Con and shRNA-Con group. Next, the
proliferation of T778 and LPS141 cells following transfection with
shRNA-PLIN1 was analyzed using the CCK-8 assay. The results
revealed that the proliferation of T778 and LPS141 cells was
markedly suppressed by PLIN1 depletion compared with the
Negative-Con or shRNA-Con group at days 3 and 5 (Fig. 2B). No significant difference was
observed in the proliferation curves of Negative-Con and shRNA-Con
transfected into T778 or LPS141 cells. These data indicated that
PLIN1 loss-of-function inhibited the proliferation of liposarcoma
cells.
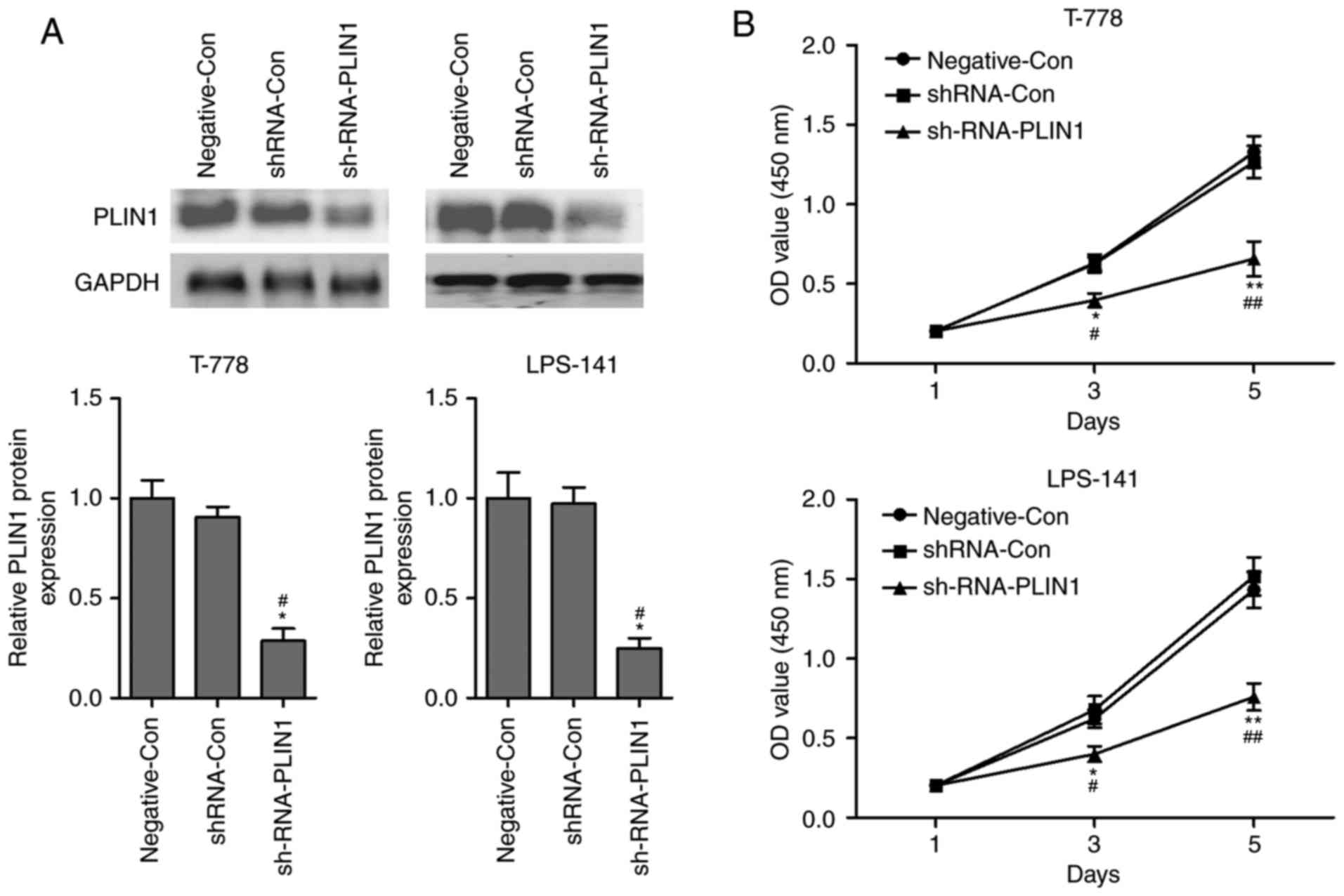 | Figure 2.Silencing of PLIN1 suppresses
proliferation of liposarcoma cells. (A) Following transfection with
Negative-Con, siRNA-Con or siRNA-PLIN1, the protein levels of PLIN1
were detected using western blotting in T778 and LPS141 cells. (B)
Following transfection with Negative-Con, shRNA-Con or shRNA-PLIN1,
cells proliferation of T778 and LPS141 cells was monitored by CCK-8
assay at days 1, 3 and 5. n=3 in each group. *P<0.05,
**P<0.01 vs. shRNA-Con group; #P<0.05,
##P<0.01 vs. Negative-Con. PLIN1, perilipin-1; Con,
control; sh, short hairpin; OD, optical density. |
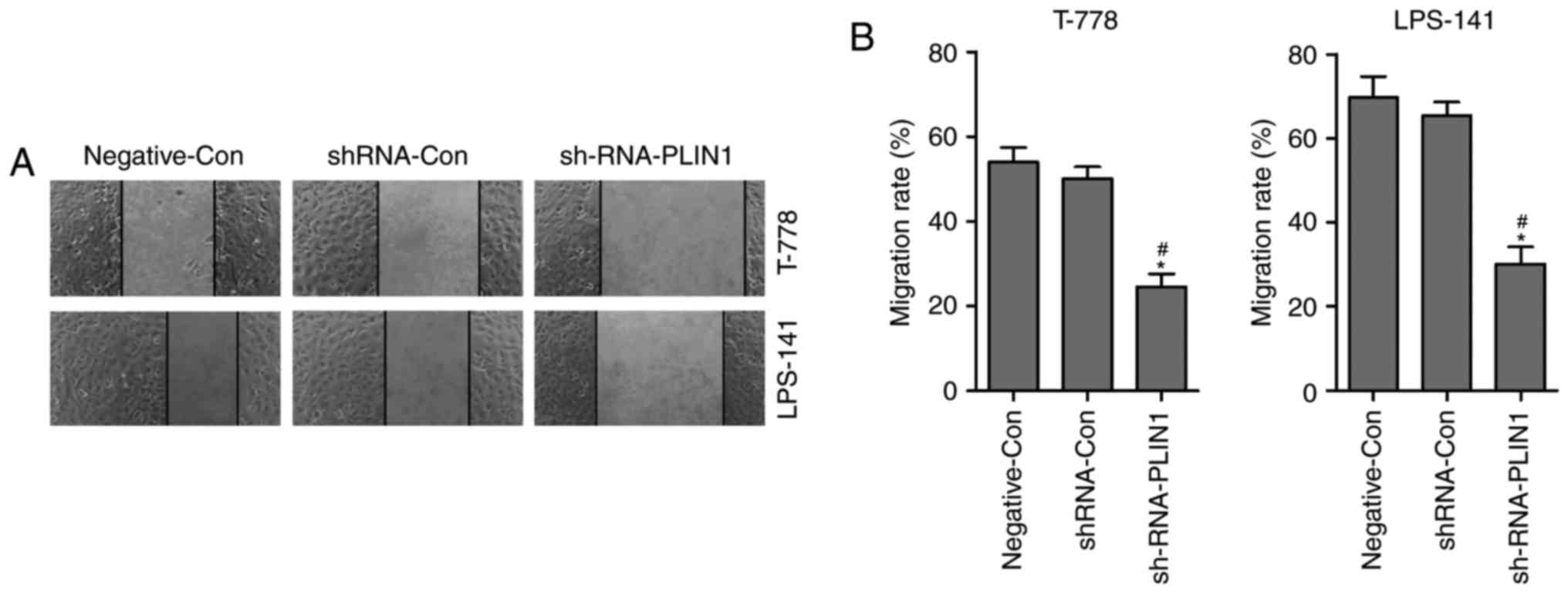
Knockdown of PLIN1 inhibits migration
of liposarcoma cells
The effect of PLIN1 knockdown on cell migration was
determined by a wound healing assay. T778 and LPS141 cells were
seeded, and then scratches made 24 h later. Transfection of
shRNA-PLIN1 led to retarded wound closing compared with the
Negative-Con or shRNA-Con groups in T778 and LPS141 cells for 12 h
(Fig. 3A and B). However, no
significant difference was observed in the wound closing results
between the Negative-Con and shRNA-Con groups in T778 and LPS141
cells (Fig. 3A and B). These
findings indicated that PLIN1 loss-of-function inhibited the
migration of liposarcoma cells.
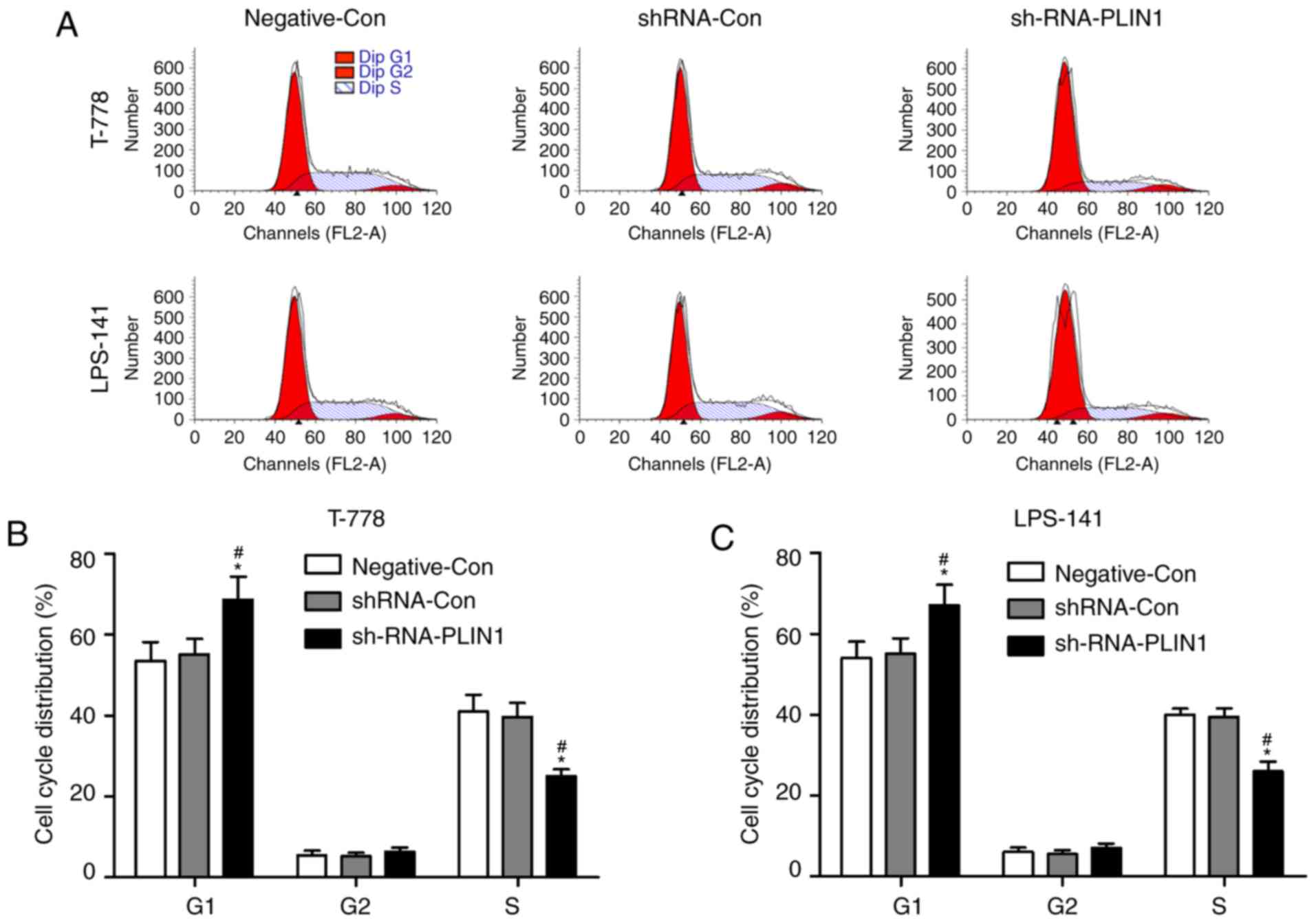
Knockdown of PLIN1 induces cell cycle
arrest and apoptosis of liposarcoma cells
Flow cytometry analysis was performed to further
evaluate whether silencing of PLIN1 regulates liposarcoma cell
proliferation by altering the progression of cell cycle and
apoptosis. T778 and LPS141 cells were transfected with
Negative-Con, shRNA-Con or shRNA-PLIN1 for 24 h, and cell cycle
distributions were determined by PI staining. As presented in
Fig. 4A and B, transfection with
shRNA-PLIN1 led to increase of cells in the G1 phase and
decrease of cells in the S phase, while transfection with
shRNA-PLIN1 had no evident effect on the G2 phase of the
cell cycle in T778 and LPS141 cells. In addition, no significant
difference was observed in cell cycle distribution between
Negative-Con and shRNA-Con transfected into T778 or LPS141 cells.
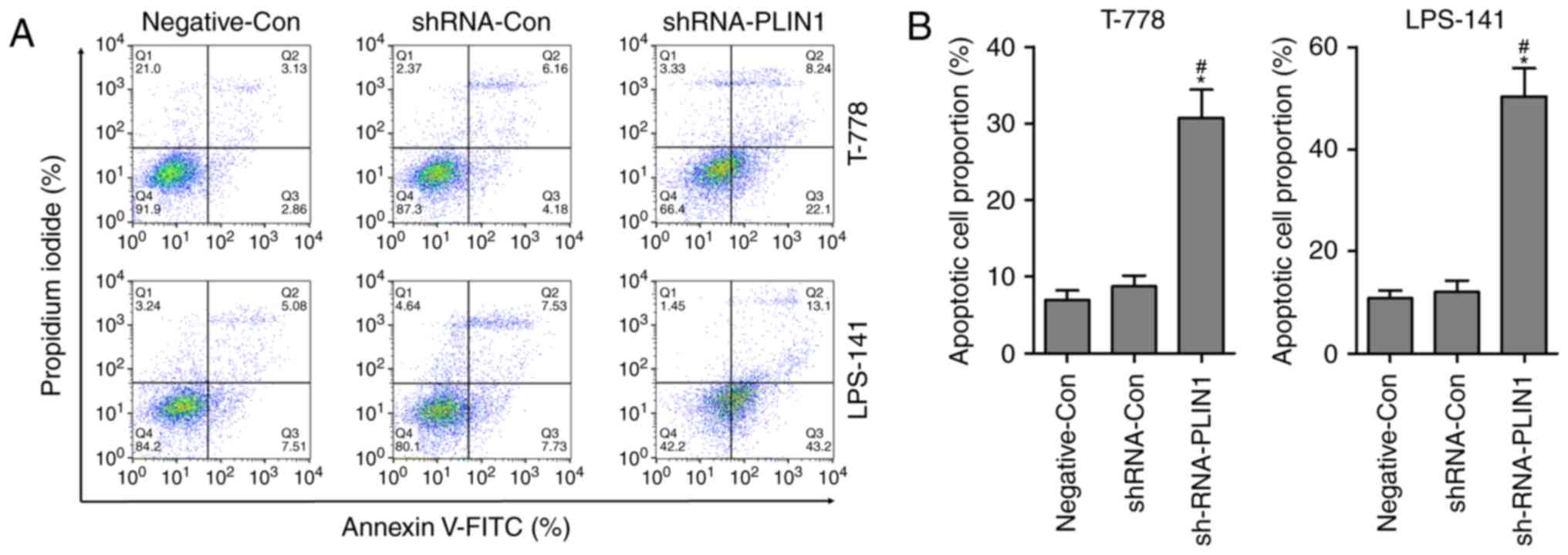
The rate of apoptosis was significantly increased when T778 and
LPS141 cells were transfected with shRNA-PLIN1 compared with the
Negative-Con or shRNA-Con group (Fig.
5A and B). These findings indicated that PLIN1 loss-of-function
could induce G1 phase cell cycle arrest and apoptosis in
T778 and LPS141 cells. Based on aforementioned findings, it was
demonstrated that knockdown of PLIN1 inhibited the proliferation of
liposarcoma cells, at least partly, by inducing G1 phase
cell cycle arrest and apoptosis.
Discussion
The present study demonstrated robust protein
expression of PLIN1 in liposarcoma tumor tissues. Silencing of
PLIN1 inhibited the proliferation and migration of liposarcoma
cells. In addition, G1 phase cell cycle arrest and
apoptosis were induced by the knockdown of PLIN1 in liposarcoma
cells. These data suggested that upregulation of PLIN1 may serve an
oncogenic role in the pathogenesis and progression of liposarcoma.
Therefore, it is hypothesized that PLIN1 may be a potential
therapeutic target for the clinical management of liposarcoma.
PLIN1 is the major lipid droplet-coating protein of
mature adipocytes and has a high specificity for adipocytes of
white and brown fatty tissue (18,19).
PLIN1 is known to be highly expressed in Leydig cells and is
closely associated with steroidogenesis and spermatogenesis
(11). Straub et al
(20) demonstrated that PLIN1 is
present in hepatocellular carcinoma, sebaceous tumors and
mesenchymal tumors with adipogenic differentiation, but PLIN1 is
restricted to lipid droplets in tumors. The other members of the
PAT protein family, adipocyte-differentiation-related protein
(adipophilin, PLIN2) and tail-interacting protein of 47 kDa (TIP47,
PLIN3), are almost ubiquitously expressed in lipid droplets in
various tumor types and normal tissues (20,21).
Westhoff et al (6)
demonstrated that more than two-thirds of liposarcomas present
PLIN1 positivity and that all non-lipomatous sarcomas studied are
negative for this marker, suggesting that PLIN1 is a highly
specific marker for liposarcoma. The present study demonstrated
that PLIN1 protein expression was significantly upregulated in
liposarcoma tumor tissues, and this was verified by
immunohistochemical staining and western blotting. These findings
suggested that robust expression of PLIN1 may possess a potential
role in the pathogenesis and development of liposarcoma.
Increasing evidence suggests that PLIN1 is
positively associated with poor overall survival rates in
metastatic breast cancer and lung cancer (7,14).
Zhou et al (7) demonstrated
that PLIN1 mRNA expression is significantly downregulated in human
breast cancer, and that the overexpression of PLIN1 in MCF-7 and
MDA-MB-231 cells markedly suppresses cell proliferation, migration,
invasion and in vivo tumorigenesis in mice. Conversely, the
present study identified that PLIN1 was significantly increased in
liposarcoma tumor tissues and that downregulation of PLIN1 by shRNA
in T778 (well differentiated liposarcoma cell line) and LPS141
(dedifferentiated liposarcoma cell line) suppressed cell
proliferation and migration. These findings suggested that PLIN1
expression may be tissue-specific. A previous study indicated that
proliferation inhibition in liposarcoma cells may be a result of
cell cycle arrest (2), so the cell
cycle distributions were analyzed using flow cytometry to detect
the effects of shRNA-PLIN1 on cell cycle arrest in T778 and LPS141
cells. It was identified that shRNA-PLIN1 significantly affected
the cell cycle distributions, increasing the percentage of cells in
G1 phase and decreasing the percentage of cells in S
phase. Notably, apoptosis was also induced in T778 and LPS141 cells
transfected with shRNA-PLIN1.
In conclusion, the findings of the present study
indicated that the expression of PLIN1 was altered in liposarcoma
tumor tissues, and preliminarily determined that PLIN1 was a key
regulator of liposarcoma. In vitro experiments demonstrated
that silencing of PLIN1 inhibited the proliferation and migration
of liposarcoma cells by inducing G1 phase cell cycle
arrest and apoptosis. However, PLIN1-associated signaling pathways
were not involved in the present study, and PLIN1-induced apoptotic
mechanisms need to be further investigated in in vivo and
in vitro experiments.
Acknowledgements
The authors would like to thank the Department of
Pathology of the People's Hospital of Rizhao for their technical
support.
Funding
This work was supported by Shandong Medical and
Health Science and Technology Development Program (grant no:
2016WS0329).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
The study was designed by LXM, YXZ and MLH.
Literature research, data acquisition and data analysis were
performed by XMZ, SYS, ZJD, QM, BCL and YWS. Histological
examination was conducted by ZJD, QM, BCL and YWS. The manuscript
was prepared and edited by LXM and YXZ. The manuscript was reviewed
by LXM, YXZ and MLH. LXM, YXZ, MLH, XMZ, SYS, ZJD, QM, BCL and YWS
approved the final version of the manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the People's Hospital of Rizhao (Approval Number: KYLL-20140903).
Human samples were obtained with written informed consent from all
patients.
Patient consent for publication
All patients provided consent for publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Garg M, Kanojia D, Mayakonda A, Said JW,
Doan NB, Chien W, Ganesan TS, Chuang LS, Venkatachalam N, Baloglu
E, et al: Molecular mechanism and therapeutic implications of
selinexor (KPT-330) in liposarcoma. Oncotarget. 8:7521–7532. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu J, Zhong D, Wei Y, Wu X, Kang L and
Ding Z: Potassium channel ether a go-go1 is aberrantly expressed in
human liposarcoma and promotes tumorigenesis. Biomed Res Int.
2014:3456782014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Franz H, Greschik H, Willmann D, Ozretić
L, Jilg CA, Wardelmann E, Jung M, Buettner R and Schüle R: The
histone code reader SPIN1 controls RET signaling in liposarcoma.
Oncotarget. 6:4773–4789. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Azar AR, Weynand B, Daumerie C and Coche
E: Metastatic liposarcoma of the thyroid gland. Br J Radiol.
76:750–752. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tseng WW, Somaiah N, Lazar AJ, Lev DC and
Pollock RE: Novel systemic therapies in advanced liposarcoma: A
review of recent clinical trial results. Cancers. 5:529–549. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Westhoff CC, Mrozinski J, Riedel I, Heid
HW and Moll R: Perilipin 1 is a highly specific marker for
adipocytic differentiation in sarcomas with intermediate
sensitivity. J Cancer Res Clin Oncol. 143:225–232. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou C, Wang M, Zhou L, Zhang Y, Liu W,
Qin W, He R, Lu Y, Wang Y, Chen XZ and Tang J: Prognostic
significance of PLIN1 expression in human breast cancer.
Oncotarget. 7:54488–54502. 2016.PubMed/NCBI
|
|
8
|
Martinez-Botas J, Anderson JB, Tessier D,
Lapillonne A, Chang BH, Quast MJ, Gorenstein D, Chen KH and Chan L:
Absence of perilipin results in leanness and reverses obesity in
Lepr (db/db) mice. Nat Genet. 26:474–479. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Souza SC, de Vargas LM, Yamamoto MT, Lien
P, Franciosa MD, Moss LG and Greenberg AS: Overexpression of
perilipin A and B blocks the ability of tumor necrosis factor alpha
to increase lipolysis in 3T3-L1 adipocytes. J Biol Chem.
273:24665–24669. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Puri V, Ranjit S, Konda S, Nicoloro SM,
Straubhaar J, Chawla A, Chouinard M, Lin C, Burkart A, Corvera S,
et al: Cidea is associated with lipid droplets and insulin
sensitivity in humans. Proc Natl Acad Sci USA. 105:7833–7838. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen M, Wang H, Li X, Li N, Xu G and Meng
Q: PLIN1 deficiency affects testicular gene expression at the
meiotic stage in the first wave of spermatogenesis. Gene.
543:212–219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao X, Gao M, He J, Zou L, Lyu Y, Zhang
L, Geng B, Liu G and Xu G: Perilipin1 deficiency in whole body or
bone marrow-derived cells attenuates lesions in
atherosclerosis-prone mice. PLoS One. 10:e01237382015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meyer MB, Benkusky NA, Sen B, Rubin J and
Pike JW: Epigenetic plasticity drives adipogenic and osteogenic
differentiation of marrow-derived mesenchymal stem cells. J Biol
Chem. 291:17829–17847. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jung YY, Kim HM and Koo JS: Expression of
lipid metabolism-related proteins in metastatic breast cancer. PLoS
One. 10:e01372042015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang HF, Zhang XZ, Liu BG, Jia GT and Li
WL: Circular RNA circ-ABCB10 promotes breast cancer proliferation
and progression through sponging miR-1271. Am J Cancer Res.
7:1566–1576. 2017.PubMed/NCBI
|
|
16
|
Seo JH, Jeong ES and Choi YK: Therapeutic
effects of lentivirus-mediated shRNA targeting of cyclin D1 in
human gastric cancer. BMC Cancer. 14:1752014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Peng YT, Shi XE, Li ZQ, He X and Sun YM:
Particularly interesting Cys-His-rich protein is highly expressed
in human intracranial aneurysms and resists aneurysmal rupture. Exp
Ther Med. 12:3905–3912. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brasaemle DL, Barber T, Wolins NE, Serrero
G, Blanchette-Mackie EJ and Londos C: Adipose
differentiation-related protein is an ubiquitously expressed lipid
storage droplet-associated protein. J Lipid Res. 38:2249–2263.
1997.PubMed/NCBI
|
|
19
|
Kimmel AR, Brasaemle DL, McAndrews-Hill M,
Sztalryd C and Londos C: Adoption of PERILIPIN as a unifying
nomenclature for the mammalian PAT-family of intracellular lipid
storage droplet proteins. J Lipid Res. 51:468–471. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Straub BK, Herpel E, Singer S, Zimbelmann
R, Breuhahn K, Macher-Goeppinger S, Warth A, Lehmann-Koch J,
Longerich T, Heid H and Schirmacher P: Lipid droplet-associated
PAT-proteins show frequent and differential expression in
neoplastic steatogenesis. Mod Pathol. 23:480–492. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Heid H, Rickelt S, Zimbelmann R, Winter S,
Schumacher H, Dörflinger Y, Kuhn C and Franke WW: On the formation
of lipid droplets in human adipocytes: The organization of the
perilipin-vimentin cortex. PLoS One. 9:e903862014. View Article : Google Scholar : PubMed/NCBI
|