Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignancies worldwide (1).
Although great progress has been achieved in the diagnosis and
treatment of HCC, the overall survival of HCC patients remains poor
(2). Alteration of metabolism is a
hallmark of cancer cells (3).
Thus, targeting metabolic pathways may be a promising strategy for
the treatment of HCC.
The mevalonate (MVA) pathway is responsible for
cholesterol synthesis in mammals (4). 3-Hydroxy-3-methylglutaryl-CoA
reductase (HMGCR) is a rate-limiting enzyme for the MVA pathway
(5). Numerous studies have
demonstrated the oncogenic roles of the MVA pathway in
tumorigenesis (6,7). It has been reported that enzymes
involved in the MVA pathway were upregulated in breast cancer and
HMGCR promoted the transformation of benign breast epithelium
(2). In gastric cancer cells,
HMGCR has been reported to activate the hedgehog signaling pathway
and promotes the growth and migration of cancer cells (8). In HCC, HMGCR has been demonstrated to
regulate the activation of yes-associated protein and the
chemoresistance of cancer cells (2). The protein expression level of HMGCR
is tightly controlled at multiple levels. Sterol regulatory
element-binding protein 2 is a key modulator of HMGCR via
regulation of its transcription (9). On the other hand, ubiquitin fusion
degradation (UFD) and autocrine motility factor receptor (GP78)
promoted the degradation of the HMGCR protein (10). In addition, several microRNAs have
been reported to target HMGCR (11). Statins, which are HMGCR inhibitors,
have attracted attention for their role in lowing cholesterol and
anti-cancer activity (2).
Therefore, increased understanding of the regulation of HMGCR may
improve the treatment of HCC.
Heat-shock protein 90 (HSP90) is a molecular
chaperon and serves an important role in the protein folding,
stability and degradation of proteins referred to as ‘client
proteins’ (12). Numerous
oncogenes (including epidermal growth factor receptor, human
epidermal growth factor receptor and protein kinase B) are client
proteins (13–15). Due to environmental stress
(hypoxia, low pH and lack of nutrients), cancer cells are more
dependent on the function of HSP90, which may explain the
significant upregulation of HSP90 in cancer tissues (16,17).
Several studies have reported the function of HSP90 in HCC by
regulating cell proliferation and apoptosis, and the inhibitor for
HSP90, 17-DMAG, has shown anti-cancer activity (18,19).
However, the detail underlying mechanism of the function of HSP90
in HCC remain largely unknown. This study examined the expression
pattern of HSP90 in HCC and investigated the underlying molecular
mechanism.
Materials and methods
Cell culture
L02, 7404, huh-7, Hep3B, QGY and MHCC97 cell lines
were obtained from American Type Culture Collection (Manassas, VA,
USA). Cells were cultured in Dulbecco's Modified Eagle's medium
(DMEM; Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) containing antibiotics (100 U/ml penicillin and 0.1 mg/ml
streptomycin; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and
10% fetal bovine serum (FBS; Sigma-Aldrich, Merck KGaA), and
incubated at 37°C in an atmosphere containing 5%
CO2.
Clinical HCC samples and tissue
array
Human clinical HCC samples were collected from
patients that received surgery at Inner Mongolia People's Hospital
(Hohhot, China). This study was approved by the ethics committee of
Inner Mongolia People's Hospital, and written informed consent was
obtained from patients. Tissues were stored at −80°C. The tissue
microarray was constructed by the Shanghai Outdo Biotech Co., Ltd.
(Shanghai, China), which included 297 patients (266 males and 31
females; aged 58–68 years) who received surgery at Inner Mongolia
People's Hospital between 01/07/2009 and 06/04/2012. Patients did
not receive chemotherapy or radiotherapy prior to operation.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA and miRNA was extracted from the cell
lines using TRIzol (Thermo Fisher Scientific, Inc.). RT was
performed using RNA to cDNA EcoDry Premix (Takara Biomedical
Technology (Beijing) Co., Ltd., Beijing, China), according to the
manufacturer's protocol. Subsequently, qPCR was performed in
triplicate with a QuantiTect SYBR Green PCR kit (Qiagen GmbH,
Hilden, Germany). The PCR protocol was as follows: 95°C for 5 min,
followed by 40 cycles of 95°C for 30 sec, 60°C for 30 sec and 68°C
for 3 min. Following a final hold step at 68°C for 10 min, the
samples were stored at 10°C. The relative gene expression levels
were calculated using the 2-ΔΔCq method (20). Primers for HSP90 were as follows:
Forward, 5′-GTTAACTGGTACCAAGAAAA-3′ and reverse,
5′-CGGATTTTGTCCAATGCATC-3′. Primers for GAPDH were as follows:
Forward, 5′-GGAGCGAGATCCCTCCAAAAT-3′ and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′.
Plasmid construction
The coding sequence of HMGCR was inserted into
pcDNA3.1 (Addgene, Inc., Cambridge, MA, USA) containing myc tag by
HindIII and EcoRV, the coding sequence of HSP90 was
inserted into P23 (Addgene, Inc.) containing Flag tag and green
fluorescent protein by XbaI and EcoRI, and the coding
sequence of UFD was inserted into pCMV-HA (Addgene, Inc.) using
PstI and BamHI. All restriction enzymes were obtained
from Takara Biomedical Technology (Beijing) Co., Ltd. An empty
vector was used as the control. The expression vectors were
transfected into 7404 and Huh-7 cells using
Lipofectamine® 2000 reagent (Thermo Fisher Scientific,
Inc.) for 8 h at 37°C, following the manufacturer's protocol. A
total of 3×105 cells were seeded in 6-well plates and
cells were transfected with 1 µg/µl plasmids using
Lipofectamine® 2000. Cells were selected using
fluorescence-activated cell sorting by detecting expression of
green fluorescent protein (independently expressed).
Knocking down the expression of HMGCR
in HCC cells
The lentivirus (multiplicity of infection=1) used to
knock down the expression of HMGCR was purchased from Shanghai
GeneChem Co., Ltd. (Shanghai, China). The sequences for HMGCR-short
hairpin (sh)RNA were as follows: Forward,
5′-AAGTCATAGTGGGGACAGTGA-3′; reverse, 5′-AAACTCTGATGGAAACTCATG-3′.
The sequence for control shRNA was as follows:
5′-AGGGGAGGGGAAAAATTTT-3′. Cells were selected with 1 µg/ml
puromycin for 1 week. Following recovery for 7 days, subsequent
assays were performed.
Western blotting
Proteins were extracted from HCC cells using
radioimmunoprecipitation assay lysis buffer (Cell Signaling
Technology, Inc., Danvers, MA, USA) containing protease and
phosphatase inhibitors. Cell lysates were centrifuged (12,000 × g
for 20 min at 4°C) and the concentration was quantified using a
Bradford assay. A total of 20 µg proteins were separated by 10%
SDS-PAGE and transferred to polyvinylidene difluoride membranes
(EMD Millipore, Billerica, MA, USA). Membranes were blocked with 5%
milk at room temperature for 1 h, and probed with primary and
secondary antibodies. The immunoreactive protein bands were
visualized using an Enhanced Chemiluminescence kit (Pierce; Thermo
Fisher Scientific, Inc.). Antibodies to HMGCR (1:1,000; cat no.
ab174830) and HSP90 (1:2,000; cat. no. ab13492) were purchased from
Abcam (Cambridge, UK), and antibodies to myc tag (1:1,000; cat. no.
sc-4084), GAPDH (1:5,000; cat. no. sc-293335), ubiquitin (1:750;
cat. no. sc-8017) and Flag tag (1:1,000; cat. no. sc-51590) were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Horseradish peroxidase-conjugated secondary antibodies, mouse IgG
(1:1,000; cat. no. 7076) and rabbit IgG (1:1,000; cat. no. 7074)
were obtained from Cell Signaling Technology, Inc.
Immunofluorescent staining
Cells were cultured on glass coverslips. Following
24 h, cells were fixed with pre-chilled methanol at −20°C for 5
min. The following procedure was performed according to standard
protocols. Briefly, after washing in PBS, the cells were blocked
with PBS containing 3% bovine serum albumin (BSA; Shanghai
Shenggong Biology Engineering Technology Service, Ltd., Shanghai,
China) for 30 min at room temperature and then incubated with HMGCR
(1:100; cat. no. ab174830) and HSP90 (1:200; cat. no. ab13492; both
Abcam) antibodies in blocking buffer overnight at 4°C, followed by
incubation with a 1:1,000 dilution of the corresponding Alexa Fluor
488-conjugated secondary antibody (cat. no. 150077; Abcam) in
blocking buffer at room temperature for 120 min. Hoechst staining
(1:2,000 dilution in PBS) was performed to label the nuclei.
Fluorescence was monitored using an inverted confocal laser
microscopy (Carl Zeiss AG, Oberkochen, Germany).
GST pull-down assay
The fusion protein GST-HSP90 was purified using
Sepharose® 4B beads (GE Healthcare Life Sciences, Little
Chalfont, UK) according to the manufacturer's protocol. Cell
lysates were prepared using cell lysis buffer (cat. no. 9803; Cell
Signaling Technology, Inc.). Following centrifugation (4°C, 10,000
× g, 20 min) the supernatant was incubated with 5 µg GST fusion
protein overnight at 4°C. The Sepharose® 4B beads were
added and incubated with cell lysate for an additional 4 h.
Subsequently, the beads were washed and the protein pulled down and
detected by western blot analysis.
Statin treatment
Lovastatin was purchased from Sigma-Aldrich (Merck
KGaA). Prior to treatment, Lovastatin was activated in 70% ethanol
containing 0.1 N NaOH at 50°C for 60 min and then neutralized with
70% ethanol containing 0.1 N HCl at room temperature for 1 h.
Activated lovastatin (10 µM) was added to the medium the day after
cells were plated, and the medium was changed every day. The cells
were treated with lovastatin for a total of 7 days.
MTT assay
MTT assay was use to examine the growth of HCC
cells. A total of 1,000 cells were seeded in 96-well plates
containing 200 µl DMEM and cultured for various durations. Every 2
days, 20 µl MTT was added into the medium and incubated with cells
for 4 h. Then, the supernatant was cleared and cells were resolved
with 200 µl DMSO. The absorabcne was measured at a wavelength of
540 nm.
Anchorage-independent growth
assay
For the soft agar assay, 6-well plates were coated
with a bottom layer (0.5% agarose and 10% FBS in DMEM).
Subsequently, the upper layer (0.35% agarose and 10% FBS in DMEM)
containing a total of 5,000 cells/well was plated. Following 14
days of incubation, the colonies were counted and measured using an
inverted microscope (Carl Zeiss AG). All experiments were performed
at least three times.
Immunohistochemistry
Tissue microarray was fixed with 4% formalin at 4°C
overnight, embedded in paraffin, cut into 5 µm-thick consecutive
sections and then deparaffinized at room temperature using xylene
and ethanol. Antigen recovery was performed in sodium citrate
solution (pH 6.0) for 20 min at 95°C. Subsequently, the sections
were washed three times for 5 min in 0.01 mol/l PBS containing 8
mmol/l Na2HPO4, 2 mmol/l
NaH2PO4 and 150 mmol/l NaCl. The sections
were then blocked for 1 h in 0.01 mol/l PBS containing 0.3% Triton
X-100 and 5% BSA (Shanghai Shenggong Biology Engineering Technology
Service, Ltd.) at room temperature, followed by incubation with
anti-HSP90 antibody (1:100; cat. no. ab13492; Abcam) at 4°C for at
least 8 h. Following brief washing with 0.01 mol/l PBS, sections
were incubated with 0.01 mol/l PBS containing horseradish
peroxidase-conjugated anti-mouse IgG (1:500; cat. no. 7076; Cell
Signaling Technology, Inc.) for 2 h at room temperature, followed
by development with 0.003% H2O2 and 0.03%
3,3′-diaminobenzidine (DAB) in 0.05 mol/l Tris-HCl (pH 7.6).
Immunohistochemistry for each sample was repeated three times.
Slides were then developed with DAB and counterstained with
hematoxylin at room temperature for 2 min. The slides were
evaluated using a light microscope.
Staining was assessed using the H-score method
(21), according to the following
equation: H-score=Σ(i × Pi). The staining intensity i) of the
tumour cells was graded from 0–3, and Pi was the percentage of
stained cells at each staining intensity. Tissues with a final
HSP90 H-score <45 were classified as ‘HSP90 low expression’ and
those with a final score ≥45 were classified as ‘HSP90 high
expression’.
Immunoprecipitation assay
The immunoprecipitation assay was conducted as
previously described (4). Cells
were washed with ice-cold PBS and lysed in Tris-buffered saline (pH
7.4) containing 50 mM Tris, 150 mM NaCl, 1% NP-40, 1 mM EDTA, 1 mM
Na3VO4, 10 mM NaF, 2.5 mg/ml aprotinin and
leupeptin, 1 mM β-glycerophosphate and 4-(2-aminoethyl)
benzenesulfonyl fluoride hydrochloride and 10 mM iodoacetate.
Lysates were incubated on ice for 15 min prior to removal of
cellular debris and nuclei via centrifugation at 10,000 × g for 20
min at 4°C. Cell lysates were incubated overnight with the HMGCR
antibody (1:1,000; cat. no. ab174830; Abcam) at 4°C. IgG (1:1,000;
cat. no. 7074; Cell Signaling Technology, Inc.) was used as
control. Protein A Sepharose beads (GE Healthcare Life Sciences) in
a 1:1 mixture with 50 mM Tris buffer (pH 7.0) were added to the
cell lysates and incubated for an additional 4 h at 4°C. The
immunoprecipitates were washed four times in Tris-buffered saline
and boiled for 5 min in 40 µl Laemmli buffer containing 0.02% blue
bromophenol and 2% β-mercaptoethanol.
Cell migration assay
Cell migration was evaluated using a Boyden chamber.
The lower chamber was loaded with 0.152 ml medium containing 10%
FBS. After 8 h, cells that migrated to the lower surface of filters
were detected with traditional hematoxylin and eosin staining for 5
min at room temperature. The migrated cells were counted under an
inverted microscope and four fields of view were taken for each
group. The experiments were repeated three times.
Overexpression of RasV12 in normal
hepatic cells
The RasV12 plasmid (cat. no. 46746) was purchased
from Addgene, Inc. and its coding sequence was inserted into a P23
lentiviral vector (cat. no. 100633; Addgene, Inc.) containing Flag
tag and green fluorescent protein using BamHI and
EcoRI. The lentivirus was produced using 293T cells as
previously described (6), and used
to infect L02 at a multiplicity of infection of 1. Green
fluorescent protein-positive cells were sorted using
fluorescence-activated cell sorting and collected. Following a
two-week recovery period, subsequent experiments were
performed.
In vivo metastasis assay
Control cells (7404/P23) and HSP90 overexpressed
cells (7404/Flag-HSP90) were labeled with the luciferase gene. A
total of 1×106 cells were injected into nude mice
through the tail vein. A total of 12 mice (male, five weeks old,
~20 g) were obtained from the Shanghai Laboratory Animal Center
(Shanghai, China) and each group contained six mice. The mice were
housed under standard conditions of 25°C with 27% humidity, free
access to food and water, and a 12 h light/dark cycle. The mice
were treated with lovastatin (50 mg/kg) every three days. Control
mice were treated with ethanol. After 6 weeks, the mice were
injected intraperitoneally with 150 mg/kg luciferin (Sigma-Aldrich;
Merck KGaA) and after 5 min the metastasis foci were examined using
Living Image 2.0 software (PerkinElmer, Inc., Waltham, MA, USA).
The mice were sacrificed on the sixth week following imaging.
Statistics analysis
Each experiment was repeated at least three times.
The data are presented as the means ± standard deviation.
Statistical analysis was performed using SPSS 15.0 (SPSS, Inc.,
Chicago, IL, USA). Statistical comparisons between two groups were
performed using Student's t-test. For comparison among multiple
groups, a two-way analysis of variance (ANOVA) was performed.
Scheffe analysis was performed following ANOVA. For survival
analysis, Kaplan-Meier curves were constructed and the P-value was
determined using logrank test. For the association between the
HSP90 expression and clinical features, the χ2 test was
performed. P<0.05 was considered to indicate a statistically
significant difference. The correlation between HSP90 and HMGCR
expression was calculated using the Pearson's correlation
coefficient.
Results
P90 is upregulated in HCC and is
associated with clinical features
To examine the expression pattern of HSP90, this
study used a HCC tissue microarray that included 297 patients with
HCC. Immunohistochemistry staining was performed to evaluate the
expression of HSP90 in HCC tissues. High HSP90 expression (score
>50) was found in ~80% (236/297) clinical samples, and the
expression of HSP90 was associated with tumor size and tumor nodes
metastasis (TNM) stage (Table I).
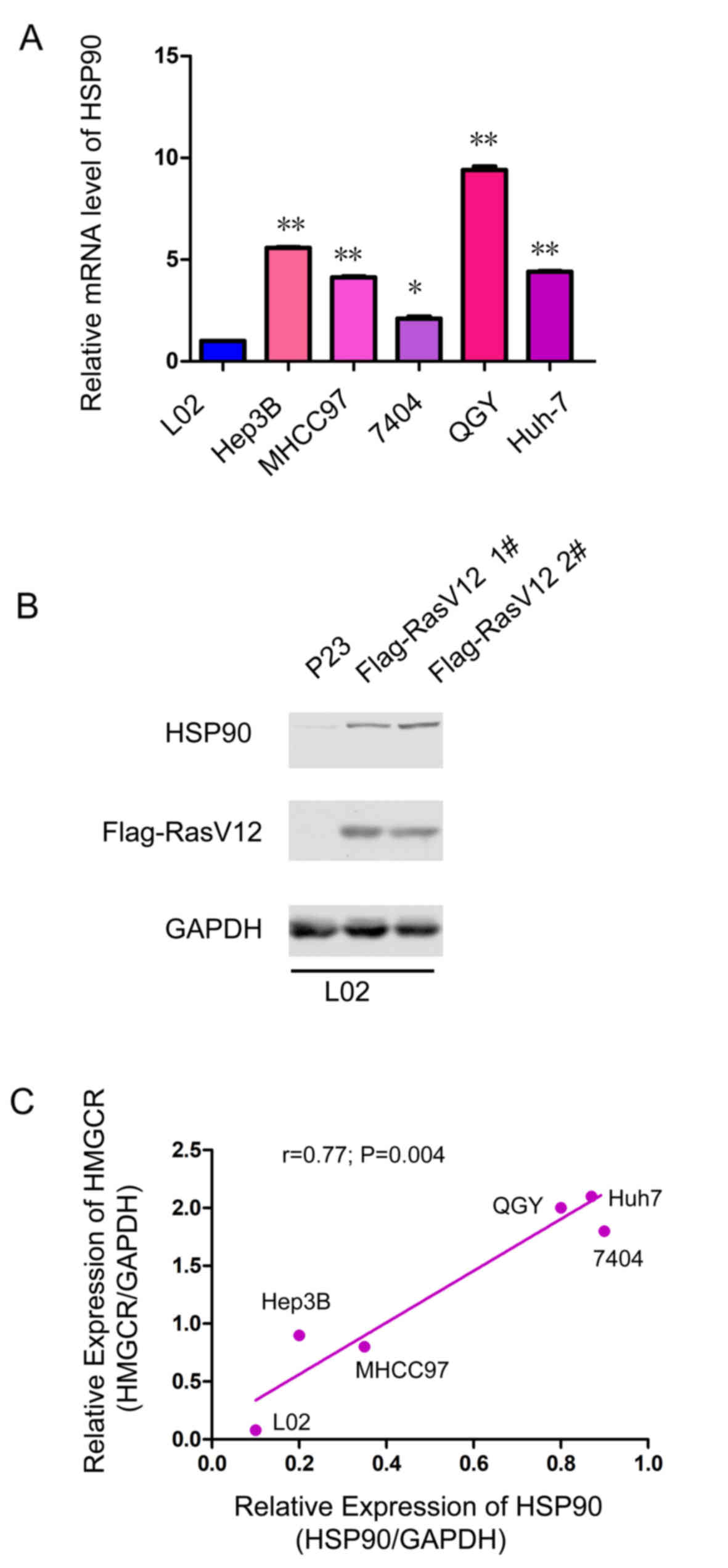
In addition, the expression of HSP90 in a normal liver cell line
(L02) and HCC cell lines (Hep3B, MHCC97, 7404, QGY and Huh-7) was
investigated. Lower expression levels of HSP90 mRNA was observed in
normal liver cells (L02), and higher HSP90 mRNA expression levels
were observed in HCC cell lines (Fig.
1A). In addition, it was found that oncogenic RasV12 expression
was associated with enhanced expression of HSP90, which may explain
the upregulation of HSP90 in HCC tissues (Fig. 1B). Furthermore, the expression of
HSP90 was positively associated with the protein expression level
of HMGCR (Fig. 1C), suggesting
that HSP90 may modulate the expression of HMGCR.
 | Table I.Association between expression of
HSP90 and clinical features. |
Table I.
Association between expression of
HSP90 and clinical features.
|
|
| HSP90 |
|
|
|---|
|
|
|
|
|
|
|---|
| Characteristic | Total (n=297) | Low (n=61) | High (n=236) | χ2 | P-value |
|---|
| Sex |
|
|
| 0.412 | 0.521 |
| Male | 266 | 56 | 210 |
|
|
|
Female | 31 | 5 | 26 |
|
|
| Age (years) |
|
|
| 0.597 | 0.44 |
|
≥45 | 182 | 40 | 142 |
|
|
|
<45 | 115 | 21 | 94 |
|
|
| Tumor size |
|
|
| 7.204 | 0.007b |
| ≥6 | 248 | 44 | 204 |
|
|
|
<6 | 49 | 17 | 32 |
|
|
| Histological
differentiation |
|
|
| 2.879 | 0.09 |
|
Well | 258 | 49 | 209 |
|
|
|
Poor | 39 | 12 | 27 |
|
|
| Organ
metastasis |
|
|
| 0.004 | 0.947 |
|
Yes | 74 | 15 | 59 |
|
|
| No | 223 | 46 | 177 |
|
|
| TNM |
|
|
| 7.217 | 0.027a |
| I | 40 | 13 | 27 |
|
|
| II | 243 | 48 | 195 |
|
|
| III,
IV | 14 | 0 | 14 |
|
|
| Tumor number |
|
|
| 0.006 | 0.939 |
| ≥2 | 30 | 6 | 24 |
|
|
| ≤1 | 267 | 55 | 212 |
|
|
| Lymph node
metastasis |
|
|
| 0.281 | 0.596 |
|
Yes | 19 | 3 | 16 |
|
|
| No | 278 | 58 | 220 |
|
|
HSP90 elevates the protein expression
level of HMGCR
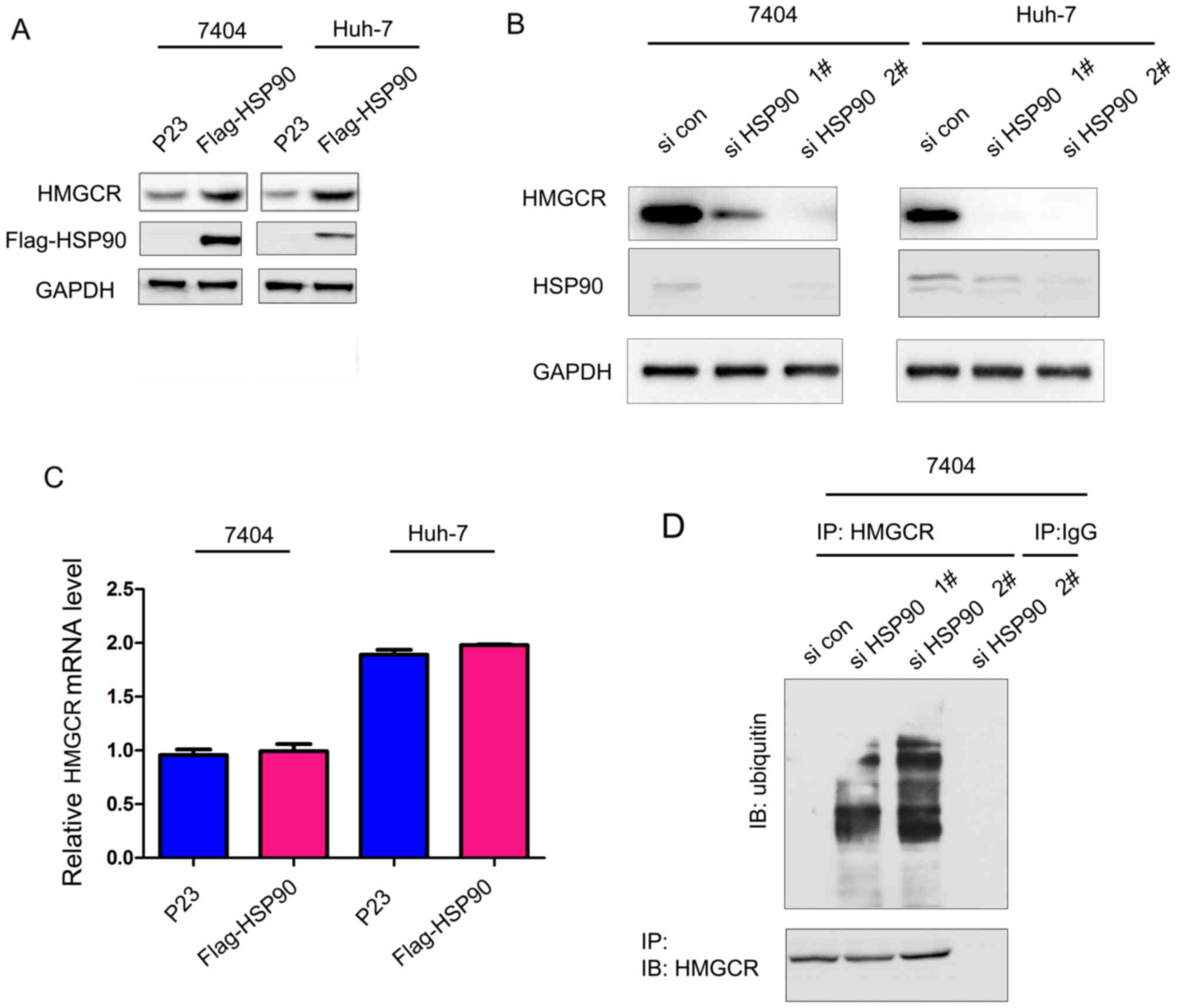
Subsequently, the present study examined the
modulation of HMGCR by HSP90. Overexpression of HSP90 in 7404 (P53
mutant) and Huh-7 (P53 mutant) cells significantly upregulated the
expression of HMGCR (Fig. 2A).
However, knock-down of HSP90 expression by two independent small
interfering RNA sequences significantly decreased the protein level
of HMGCR (Fig. 2B). In addition,
the expression of HSP90 did not affect the mRNA expression levels
of HMGCR (Fig. 2C), suggesting
that HSP90 regulates the protein levels of HMGCR at the
post-translation level. Consistent with this hypothesis, it was
found that knock-down HSP90 promoted the ubiquitination of HMGCR
(Fig. 2D), suggesting that HSP90
upregulates the protein levels of HMGCR by inhibiting its
degradation.
HSP90 interacts with HMGCR
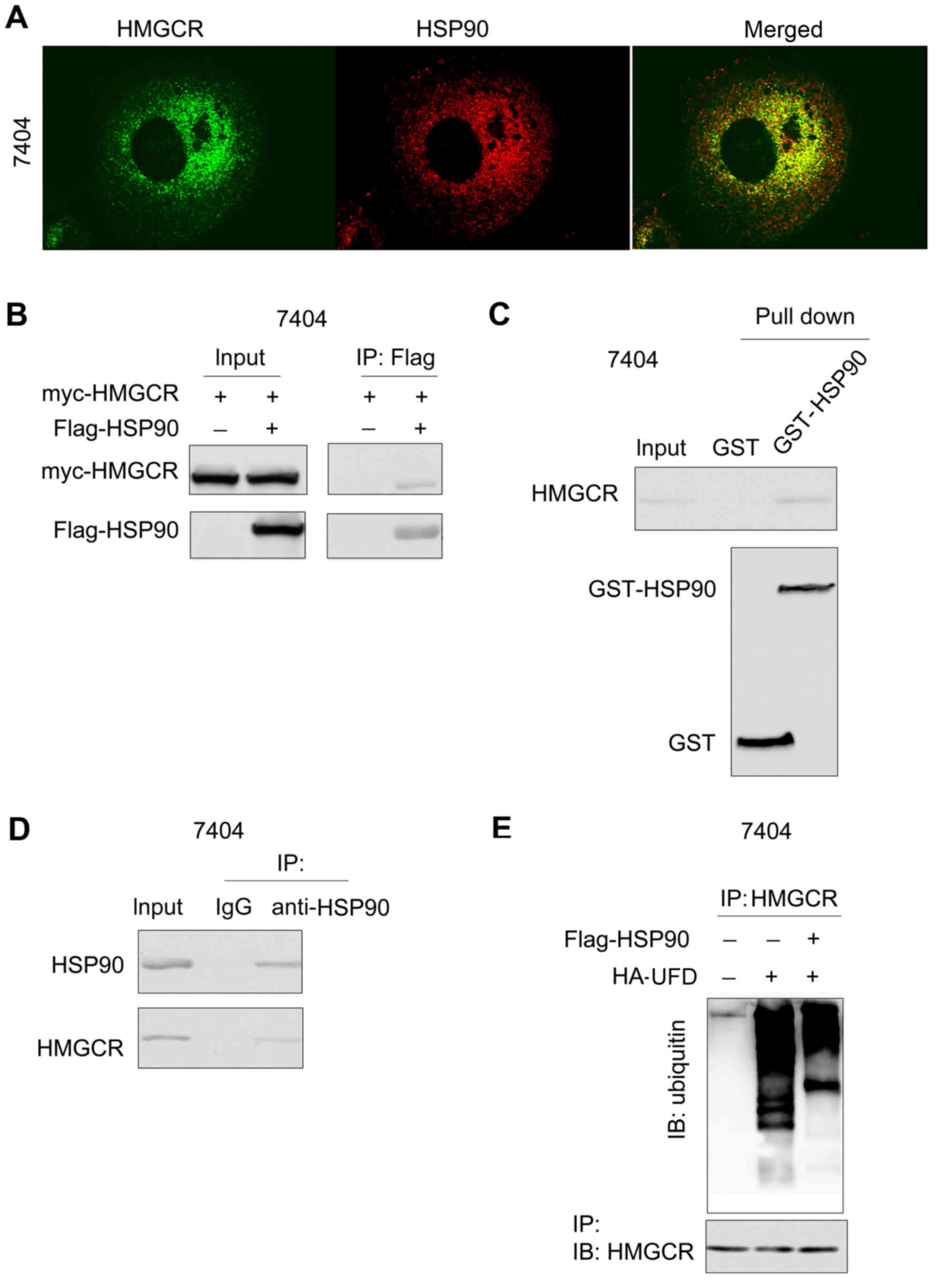
To explore the underlying mechanism by which HSP90
may inhibit the degradation of HMGCR, the interaction between HSP90
and HMGCR was examined. Immunofluorescent staining demonstrated the
co-localization of HSP90 and HMGCR (Fig. 3A). Consistent with these
observations, exogenously expressed HSP90 (Flag-HSP90) and HMGCR
(myc-HMGCR) formed a complex in 7404 cells (Fig. 3B). In addition, the fusion protein
GST-HSP90 formed a complex with endogenously expressed HMGCR
(Fig. 3C). Furthermore,
endogenously expressed HMGCR interacted with HSP90 in the
immunoprecipitation assay using anti-HSP90 antibody (Fig. 3D). These data demonstrated an
interaction between HMGCR and HSP90. The study subsequently
investigated the biological significance of the interaction between
HSP90 and HMGCR. Overexpression of HSP90 attenuated the degradation
of HMGCR induced by the expression of UFD (Fig. 3E). Taken together, these results
suggested that HSP90 interacted with HMGCR and inhibited its
degradation.
Lovastatin, the inhibitor of HMGCR,
inhibits the growth and migration of HCC cells
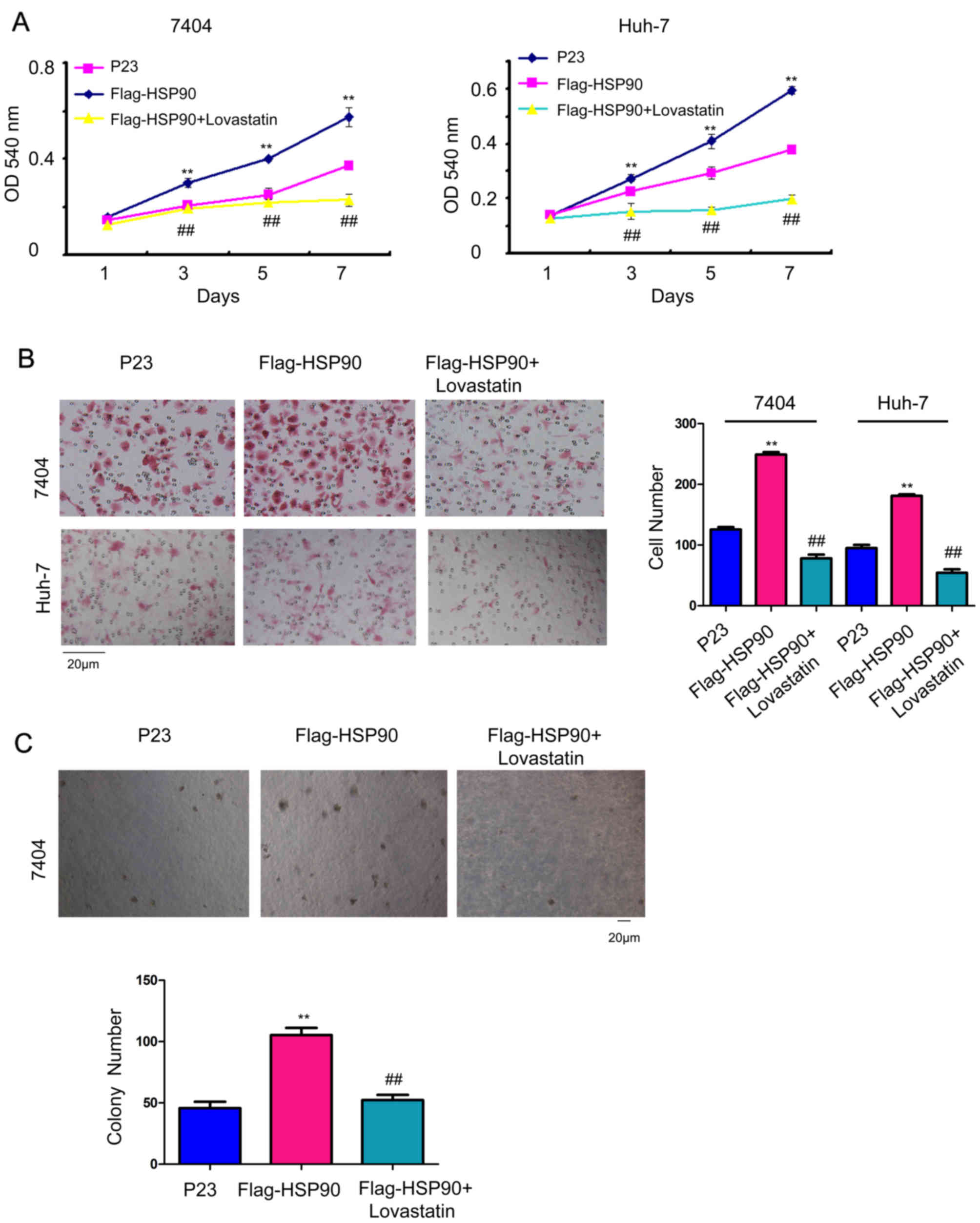
To study whether HMGCR mediated the biological
functions of HSP90 in HCC cells, 10 µM lovastatin, an inhibitor of
HMGCR, was used to rescue the functions of HSP90. HSP90 promoted
the growth (Fig. 4A), migration
(Fig. 4B) and
anchorage-independent growth of HCC cells (Fig. 4C), which was attenuated by the
treatment of lovastatin. These results demonstrated that HSP90
regulated cell growth and migration by elevating the activity of
HMGCR.
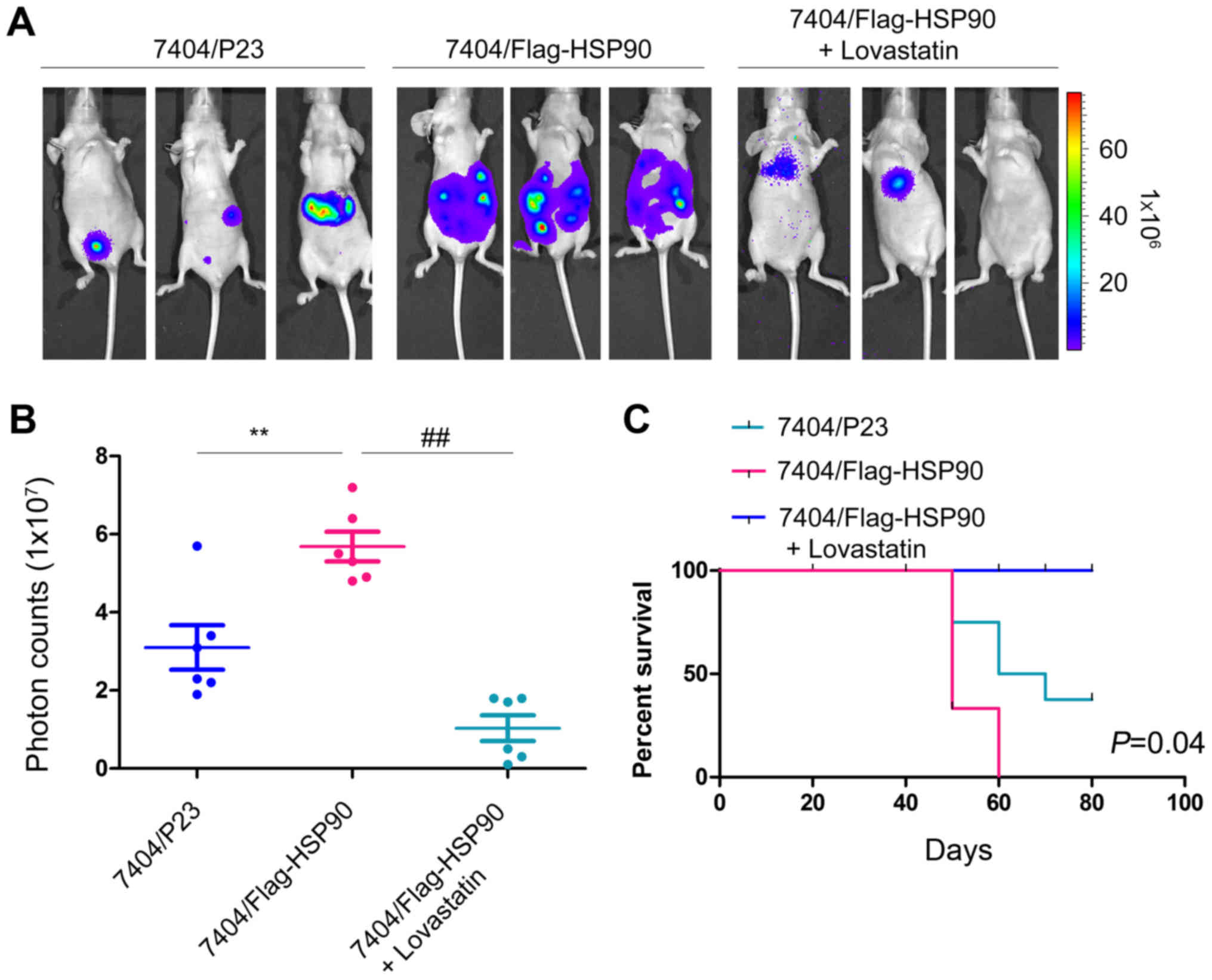
Lovastatin inhibits the metastasis of
HCC cells in vivo
As presented in Table
I, the expression of HSP90 was positively associated with the
tumor TNM stage. Subsequently, this study examined whether HSP90
promoted the metastasis of HCC cells and whether its promotion on
metastasis of HCC cells was dependent on the activity of HMGCR. As
demonstrated in Fig. 5A, forced
expression of HSP90 promoted the metastasis of 7404 cells, which
were was injected into nude mice through the tail vein.
Furthermore, the administration of lovastatin effectively blocked
the metastasis of 7404 cells in vivo, which was demonstrated
by the fluorescence intensity, photon counts and prolonged the
survival of the mice (Fig. 5A-C).
Taken together, these data suggested that HSP90 promoted the
metastasis of HCC cells, which was dependent on the activity of
HMGCR.
Discussion
Although several studies have demonstrated the
oncogenic roles of HSP90 in the progression of HCC by promoting the
proliferation and inhibiting apoptosis of cancer cells, the
detailed underlying mechanism of the oncogenic roles of HSP90 in
HCC remains poorly understood (22,23).
The findings of the current study demonstrated that the expression
of HSP90 was significantly associated with clinical features. In
addition, the expression of HSP90 was induced upon the activation
of Ras signaling. Furthermore, HSP90 inhibited the degradation of
HMGCR, the rate-limiting enzyme of the mevalonate pathway, which
suggested a role for HSP90 in cholesterol metabolism. Notably,
lovastatin, an inhibitor of HMGCR, reduced the malignant behavior
of cancer cells via overexpression of HSP90 both in vitro
and in vivo, suggesting that the clinical use of statins may
be beneficial for HCC treatment. These data suggested that HSP90
promoted the progression of HCC by activating the mevalonate
pathway.
An important finding of this study was the
regulation of HMGCR degradation by HSP90. A previous study
demonstrated that UFD and the E3 ligase GP78 cooperated to promote
the ubiquitination and degradation of HMGCR (10). The present study demonstrated an
interaction between HSP90 and HMGCR, and HSP90 inhibited the
ubiquitination of HMGCR induced by UFD. Based on these
observations, it is speculated that HSP90 may compete with UFD for
binding with HMGCR.
Another notable finding was the induction of HSP90
by RasV12. Aberrant activation of RasV12 signaling is observed in
~100% of HCC clinical samples (24), which may provide an explanation for
the upregulation of HSP90 in HCC samples. In addition, HSP90 has
been reported to positively regulate β-catenin/T-cell factor
signaling (25), suggesting that
HSP90 may be an important protein for various signaling
pathways.
In the current study, lovastatin, an inhibitor of
HMGCR, exhibited anti-cancer activity in HCC cells. Lovastatin
impaired the growth, migration and metastasis of HCC cells driven
by HSP90. Combination of
17-dimethylaminoethylamino-17-demethoxygeldanamycin (an inhibitor
of HSP90) and lovastatin (the inhibitor for HMGCR) may be useful
for HCC therapy.
In conclusion, the present study may have elucidated
a mechanism through which HSP90 promotes the progression of HCC,
and HSP90 may be a promising therapeutic target. Further
investigation of the therapeutic effects of HSP90 and HMGCR
inhibitors for HCC will provide novel insights.
Acknowledgements
Not applicable.
Funding
This study was supported by the Nature Science
Foundation of Inner Mongolia (grant no. 2015MS08144).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LD and RG designed the present study. LX, CZ, HL and
ZC performed the experiments.
Ethics approval and consent to
participate
This study was approved by the ethics committee of
Inner Mongolia People's Hospital, and written informed consent was
obtained from patients.
Patient consent for publication
Written informed consent was obtained from
patients.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dai XY, Zhuang LH, Wang DD, Zhou TY, Chang
LL, Gai RH, Zhu DF, Yang B, Zhu H and He QJ: Nuclear translocation
and activation of YAP by hypoxia contributes to the chemoresistance
of SN38 in hepatocellular carcinoma cells. Oncotarget. 7:6933–6947.
2016.PubMed/NCBI
|
|
3
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hashemi M, Hoshyar R, Ande SR, Chen QM,
Solomon C, Zuse A and Naderi M: Mevalonate cascade and its
regulation in cholesterol metabolism in different tissue in health
and disease. Curr Mol Pharmacol. 10:13–26. 2016. View Article : Google Scholar
|
|
5
|
Gabor KA and Fessler MB: Roles of the
mevalonate pathway and cholesterol trafficking in pulmonary host
defense. Curr Mol Pharmacol. 10:27–45. 2016. View Article : Google Scholar
|
|
6
|
Hashimoto A, Oikawa T, Hashimoto S, Sugino
H, Yoshikawa A, Otsuka Y, Handa H, Onodera Y, Nam JM, Oneyama C, et
al: P53- and mevalonate pathway-driven malignancies require Arf6
for metastasis and drug resistance. J Cell Biol. 213:81–95. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zahra Bathaie S, Ashrafi M, Azizian M and
Tamanoi F: Mevalonate Pathway and Human Cancers. Curr Mol
Pharmacol. 10:77–85. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chushi L, Wei W, Kangkang X, Yongzeng F,
Ning X and Xiaolei C: HMGCR is up-regulated in gastric cancer and
promotes the growth and migration of the cancer cells. Gene.
587:42–47. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mukherjee M, Basu Ball W and Das PK:
Leishmania donovani activates SREBP2 to modulate macrophage
membrane cholesterol and mitochondrial oxidants for establishment
of infection. Int J Biochem Cell Biol. 55:196–208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cao J, Wang J, Qi W, Miao HH, Wang J, Ge
L, DeBose-Boyd RA, Tang JJ, Li BL and Song BL: Ufd1 is a cofactor
of gp78 and plays a key role in cholesterol metabolism by
regulating the stability of HMG-CoA reductase. Cell Metab.
6:115–128. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Singh R, Yadav V, Kumar S and Saini N:
MicroRNA-195 inhibits proliferation, invasion and metastasis in
breast cancer cells by targeting FASN HMGCR, ACACA and CYP27B1. Sci
Rep. 5:174542015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Richter K and Buchner J: Hsp90: Twist and
fold. Cell. 127:251–253. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sawai A, Chandarlapaty S, Greulich H,
Gonen M, Ye Q, Arteaga CL, Sellers W, Rosen N and Solit DB:
Inhibition of Hsp90 down-regulates mutant epidermal growth factor
receptor (EGFR) expression and sensitizes EGFR mutant tumors to
paclitaxel. Cancer Res. 68:589–596. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Citri A, Gan J, Mosesson Y, Vereb G,
Szollosi J and Yarden Y: Hsp90 restrains ErbB-2/HER2 signalling by
limiting heterodimer formation. EMBO Rep. 5:1165–1170. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sato S, Fujita N and Tsuruo T: Modulation
of Akt kinase activity by binding to Hsp90. Proc Natl Acad Sci USA.
97:10832–10837. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Minet E, Mottet D, Michel G, Roland I,
Raes M, Remacle J and Michiels C: Hypoxia-induced activation of
HIF-1: Role of HIF-1alpha-Hsp90 interaction. FEBS Lett.
460:251–256. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee YJ, Curetty L, Hou ZZ, Kim SH, Kim JH
and Corry PM: Effect of pH on quercetin-induced suppression of heat
shock gene expression and thermotolerance development in HT-29
cells. Biochem Biophys Res Commun. 186:1121–1128. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou CC, Yang F, Yuan SX, Ma JZ, Liu F,
Yuan JH, Bi FR, Lin KY, Yin JH, Cao GW, et al: Systemic genome
screening identifies the outcome associated focal loss of long
noncoding RNA PRAL in hepatocellular carcinoma. Hepatology.
63:850–863. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao S, Li H, Jiang C, Ma T, Wu C, Huo Q
and Liu H: 17-Demethoxy-reblastatin, an Hsp90 inhibitor, induces
mitochondria-mediated apoptosis through downregulation of Mcl-1 in
human hepatocellular carcinoma cells. J Bioenerg Biomembr.
47:373–381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)). Methods. 25402–408. (25)2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cai Z, Qian ZY, Jiang H, Ma N, Li Z, Liu
LY, Ren XX, Shang YR, Wang JJ, Li JJ, et al: HPCL3s promotes
metastasis of hepatocellular carcinoma by activating β-catenin/TCF
signaling. Cancer Res. 78:2536–2549. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang C, Zhang Y, Guo K, Wang N, Jin H, Liu
Y and Qin W: Heat shock proteins in hepatocellular carcinoma:
Molecular mechanism and therapeutic potential. Int J Cancer.
138:1824–1834. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu X, Chen S, Tu J, Cai W and Xu Q: HSP90
inhibits apoptosis and promotes growth by regulating HIF-1alpha
abundance in hepatocellular carcinoma. Int J Mol Med. 37:825–835.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Neuzillet C, Tijeras-Raballand A, de
Mestier L, Cros J, Faivre S and Raymond E: MEK in cancer and cancer
therapy. Pharmacol Ther. 141:160–171. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cooper LC, Prinsloo E, Edkins AL and
Blatch GL: Hsp90alpha/beta associates with the
GSK3beta/axin1/phospho-beta-catenin complex in the human MCF-7
epithelial breast cancer model. Biochem Biophys Res Commun.
413:550–554. 2011. View Article : Google Scholar : PubMed/NCBI
|