Introduction
Dysfunction caused by hypoxia is an important
component of numerous diseases (1). Tissue hypoxia is induced by decreased
blood flow, arterial blood oxygen partial pressure and interrupted
blood flow (2). When oxygen supply
is insufficient or the balance between O2 demand and
supply is disrupted, distant organs can be damaged, and neuronal
injury (2) or microcirculation
disturbance in perfusion organs, such as the liver, kidneys and
gastrointestinal tract (3), may
occur. The heart is at the center of blood and oxygen supply to all
organs. However, hypoxia is also a major contributor to
cardiomyocyte pathophysiology, with a role in diseases including
congenital heart disease, myocardial infarction (4–6) and
ischemic heart disease (7,8).
The metabolic center of oxygen consumption is the
mitochondria, which can produce up to 90% of the adenosine
triphosphate (ATP) required for cell function (9–12).
Previous studies have demonstrated that the mitochondria act as a
power source for the cell and as an effector of other critical
events, including apoptosis and cell death (13,14).
Therefore, it is important to understand their function,
particularly with regard to metabolism. In contrast to other
subcellular structures that are regulated by karyogenes,
mitochondria contain their own transcription system. Mitochondrial
DNA (mtDNA) is a double-stranded circular DNA containing 37 genes
and encoding 13 polypeptides that are involved in the synthesis of
ATP (15,16). Although ~1,500 proteins participate
in the regulation of mitochondrial structure and function, because
of the limited coding capacity of mtDNA, <1% of these proteins
are encoded by mitochondria, and the remaining proteins are encoded
by nuclear genes (17–19). Therefore, the biological processes
of mitochondria are regulated by both mitochondrial and nuclear
genes.
Nuclear respiratory factor-1 (NRF-1), which was
initially discovered during the investigation of cytochrome
c transcriptional activation (20), is essential for early embryogenesis
in mammals, and loss of NRF-1 results in a peri-implantation lethal
phenotype. Furthermore, NRF-1−/− blastocysts exhibited
decreased mtDNA amounts (21).
NRF-1 also serves an important role in the integration of nuclear
and mitochondrial interactions (20,22–24).
For example, NRF-1 mediates the transcription of mtDNA by affecting
the promoter region of mitochondrial transcription factor A (mtTFA;
also termed Tfam) (25), thus
altering mitochondrial biogenesis (26–28).
Nuclear factor (NF)-κB can regulate the NRF-1 gene directly
via the lipopolysaccharide-receptor pathway, leading to increased
mitochondrial mRNA transcription and enrichment of mtDNA copy
number (29). Furthermore, in
aerobic cardiac cells, NRF-1 is associated with the transcriptional
control of complex II and prevention of pseudo-hypoxic gene
expression (30).
Cobalt chloride (CoCl2) is often used as
a hypoxia mimic agent in vivo and in vitro (31,32)
and it have been demonstrated to activate hypoxia-associated
signals, such as stabilizing hypoxia inducible factor-1α (HIF-1α)
(33,34). HIF-1α can be hydroxylated and then
ubiquitinated for degradation by the proteasome in normoxic
conditions (35–37); however, under hypoxic conditions or
in the presence of low oxygen concentrations, the α subunit is not
hydroxylated, allowing HIF-1α to enter the nucleus inducing the
transcription of certain hypoxia response elements (38–40).
Therefore, in the present study, it was aimed to
further elucidate the role of NRF-1 in hypoxia. To this end, the
effects of NRF-1 overexpression in H9C2 cardiomyoblasts on
CoCl2-stimulated hypoxia were investigated.
Materials and methods
Materials
The lentiviral expression vector
pLenti6.3-NRF1-IRES2-EGFP and lentiviral packaging plasmids (pLP1,
pLP2 and pLP/VSVG) were purchased from Invitrogen (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). H9C2 cells were purchased from
cell bank of the Chinese Academy of Sciences (Shanghai, China).
Plasmid extraction and purification kits purchased from Axygen
(Corning Incorporated, Corning, NY, USA). TRIzol reagent, 0.25%
Trypsin, Dulbecco's modified Eagle's medium (DMEM), fetal bovine
serum (FBS) and 293T cells were purchased from Invitrogen (Thermo
Fisher Scientific, Inc.). The Cell Counting Kit-8 (CCK-8) was
purchased from TransGen Biotech (Beijing, China). Hoechst 33342 was
purchased from Beyotime Institute of Technology (Haimen, China).
TransScript Reverse Transcriptase and qPCR SuperMix were purchased
from TransGen Biotech.
NRF-1 transfection
293T packaging cells (1×107) were plated
in 10-cm plates before transfection. PLenti6.3-NRF1-IRES2-EGFP
plasmids (3 µg) and 9 µg packaging plasmids (3 µg pLP1, 3 µg pLP2
and 3 µg pLP/VSVG) were co-transfected into the 293T cells using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and inoculated in a 10 cm culture dish before
transfection. Virus-containing supernatant was isolated under
50,000 × g at 4°C and collected after 2 h. Virus was added to the
H9C2 cells (1×105/ml) in the presence of 8 µg/ml
polybrene (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany).
Following transfection for 48 h, the target cells were subjected to
1 µg/ml puromycin for selection. The transfected cells were
designated as NRF1-transfected H9C2 (NRF1-H9C2) cells and empty
virus-transfected as pLenti-H9C2 cells.
Cell culture and treatment
NRF1-H9C2 or pLenti-H9C2 cells (5×106)
were cultured in 10-cm culture plates in DMEM supplemented with 10%
FBS and 2 mM glutamine and incubated in a humidified incubator with
an atmosphere containing 5% CO2 and 21% O2 at
37°C. Chemical hypoxia was induced by adding the hypoxia-mimetic
agent CoCl2 (Sigma-Aldrich; Merck KGaA) at 200 or 400
µM, and cells were then incubated for 6 or 24 h (41,42).
Determination of cell viability
5×104 NRF1-H9C2 and pLenti-H9C2 cells
(5×106) were seeded in 96-well plates and treated with
200 or 400 µM CoCl2 for 6 or 24 h. Subsequently, 10 µl
CCK-8 reagent was added to each well, and the plates were incubated
at 37°C for 3 h. Absorbance was measured at 450 nm using a
microplate reader. The cell viability (%) relative to the control
was calculated as follows: Relative cell viability (%) = optical
density (OD) sample/OD control ×100. Each group was analyzed using
five wells, and the experiment was repeated at least three
times.
Analysis of mitochondrial membrane
potential (MMP)
Cells (5×105) were seeded in 6-well
plates and the cells were stained with 2.5 nM tetramethylrhodamine
ethyl ester (Sigma-Aldrich; Merck KGaA) for 30 min at 37°C
(43), then washed twice with PBS
and analyzed using an Accuri C6 flow cytometer (BD Biosciences,
Franklin Lakes, NJ, USA) and BD Accuri C6 Software (version 1.0; BD
Biosciences).
Analysis of apoptosis by Hoechst 33342
staining
Cells (2×105) were seeded in 24-well
plates and propagated to ~80% confluence, then washed with PBS
twice, stained with Hoechst 33342 (50 µg/ml) at 37°C, and incubated
in the dark. After 20 min, the nuclear morphology of the cells was
observed to determine the occurrence of apoptosis using a
fluorescence microscope.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol reagent in
accordance with the manufacturer's protocol. cDNA was synthesized
from total RNA using Transcript Reverse Transcriptase (Transgene
Biotech; the reaction system is presented as Table I) then incubated for 30 min at 42°C
and heated at 85°C for 5 sec. Fluorescent qPCR was performed using
Transcript Green Two-Step qRT-PCR Super Mix (Transgene Biotech).
Specific primers were used to amplify NRF-1, NRF-2, mtTFA,
HIF-1α, mitochondrial single stranded DNA binding protein
(mtSSB), Bcl-associated X protein (Bax), B-cell
lymphoma-2 (Bcl-2), and GAPDH, as presented in
Table II. All fluorescent qPCR
assays were performed in duplicate 20-µl reactions, and detection
was carried out using a Light Cycler 2 real-time PCR machine
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). The thermocycling
conditions were: 95°C for 2 min followed by 35 cycles of
amplification (95°C for 15 sec, 55°C for 30 sec and 72°C for 30
sec). The data was quantified by 2−ΔΔCq method (44).
 | Table I.cDNA reaction system. |
Table I.
cDNA reaction system.
| Component | Volume |
|---|
| Total
RNA | 1 µl (50 ng-5
µg) |
| Oligo(dT)
Primer | 1 µl |
| 10 mM dNTPs | 1 µl |
|
5xBuffer | 4 µl |
| Ribonuclease
Inhibitor | 0.5 µl |
| TransScript
RT | 1 µl |
| RNase-free
Water | 11.5 µl |
| Total
volume | 20 µl |
| Table II.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table II.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Direction | Sequence
(5′-3′) |
|---|
| NRF-1 | Sense |
CCACGTTGGATGAGTACACG |
|
| Antisense |
CTGAGCCTGGGTCATTTTGT |
| NRF-2 | Sense |
CCGCTACACCGACTACGATT |
|
| Antisense |
ACCTTCATCACCAACCCAAG |
| mtTFA | Sense |
GCTTCCAGGAGGCTAAGGAT |
|
| Antisense |
CCCAATCCCAATGACAACTC |
| HIF-1α | Sense |
TGCTGATTTGTGAACCCATT |
|
| Antisense |
TCTGGCTCATAACCCATCAA |
| mtSSB | Sense |
AGCCAGCAGTTTGGTTCTTG |
|
| Antisense |
CCACTTTGCCTTCCACAAAT |
| Bax | Sense |
TTGCTACAGGGTTTCATCCA |
|
| Antisense |
CTGAGACACTCGCTCAGCTT |
| Bcl-2 | Sense |
GAGTACCTGAACCGGCATCT |
|
| Antisense |
GAAATCAAACAGAGGTCGCA |
| GAPDH | Sense |
ACAGCAACAGGGTGGTGGAC |
|
| Antisense |
TTTGAGGGTGCAGCGAACTTT |
Western blotting
Cells were lysed using cell lysis buffer (Beyotime
Institute of Biotechnology). Homogenate protein concentrations were
measured using a bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology). Cell lysates (50 µg) were separated by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis on 12%
gels and transferred onto polyvinylidene difluoride membranes (EMD
Millipore). The membranes were blocked with 5% nonfat milk in PBS
containing 0.1% Tween-20 for 2 h at room temperature and then
incubated overnight at 4°C with anti-NRF1 (1:1,000; ab175932),
anti-Bax (1:1,000; ab32503), anti-Bcl-2 (1:500; ab59348),
anti-poly(ADP-ribose) polymerase 1 (PARP; 1:500; ab32064) and
anti-β-actin (1:2,000; ab49900); all from Abcam (Cambridge, UK)].
Subsequently, the membranes were incubated with horseradish
peroxidase-linked secondary anti-rabbit (1:5,000; BA1054; Boster
Biological Technology, Pleasanton, CA, USA) at room temperature for
2 h. Antibody-bound protein was detected using a chemiluminescent
detection kit (Super Signal West Dura; Thermo Fisher Scientific,
Inc.), exposed to X-ray film, and densitometry was quantified by
Image Lab version 5.1 (Bio-Rad Laboratories, Inc.).
Annexin V-allophycocyanin
(APC)/7-aminoactinomycin D (7-AAD) staining
Cells were seeded in 24-well plates
(2×105 per well) and cultured with medium containing 400
µM CoCl2 for 6 h. Apoptotic cells were measured by flow
cytometry using Annexin V-APC and 7-AAD Apoptosis Detection reagent
(BD Biosciences). Briefly, cell were digested, collected, washed
and incubated at room temperature for 10 min with 1X binding buffer
containing Annexin V-conjugated APC and 7-AAD. Annexin V-APC
staining was used to detect exposure of phosphatidylserine. 7-AAD
is able to penetrate the cells when cell apoptosis and cell death
occurs. Annexin V-APC was used to detect early apoptotic cells, and
Annexin V-APC and 7-AAD were used to detect necrotic cells or/and
nonviable apoptotic cells. Cells were detected by BD Accuri C6 Flow
Cytometer and analyzed by BD Accuri™ C6 Software version
1.0 (BD Biosciences).
Detection of energy metabolism
An XFe24 Analyzer (Seahorse Bioscience;
Agilent Technologies, Inc., Santa Clara, CA, USA) was used to
detect cellular energy, and XF Cell Mito Stress Tests (Seahorse
Bioscience; Agilent Technologies, Inc.) were used to detect oxygen
consumption rate. Briefly, the day before the assay,
1×106 cells were seeded in Seahorse 24-well XF Cell
Culture Microplates, and the cells were then allowed to grow
overnight in a cell culture incubator containing 5% CO2
and 95% air at 37°C. The cells were then treated with 400 µM
CoCl2 at 37°C for 6 h. Additionally, a Hydrate Sensor
Cartridge was placed in a non-CO2 incubator (37°C)
overnight. The day of the assay, cells were washed with XF
Glycolysis Stress Test Assay Medium, stock compounds [1 µM
oligomycin, 2 µM carbonyl cyanide p-trifluoromethoxyphenylhydrazone
(FCCP) and 0.5 µM rotenone/antimycin A] were added, and the
cartridge ports were loaded. Following setting up the program on an
XF Controller, the plate was placed in the instrument for testing.
OCR was measured for about 3 min to detect basic OCR. Following
baseline measurements, OCR was detected after injection of
oligomycin, then FCCP was injected and the maximal OCR was
measured. Non-mitochondrial oxygen consumption level was detected
after the injection of rotenone/antimycin A.
Statistical analysis
Data were expressed as the mean ± standard error.
Statistical significance was assessed by two-tailed Student's
t-tests or one-way analysis of variance with post hoc comparisons
using the Student-Newman-Keuls test with GraphPad Prism 5.0
(GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
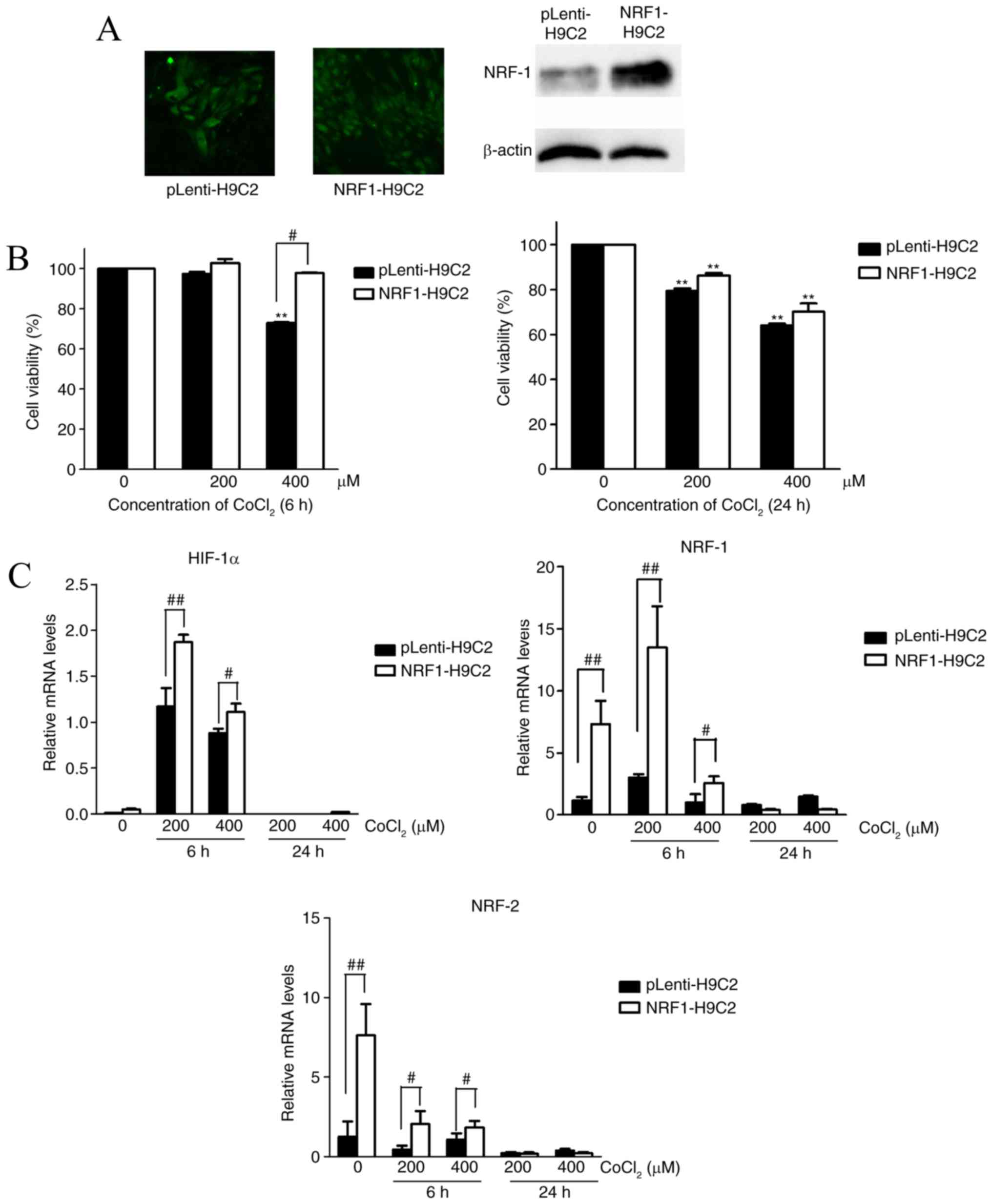
Overexpression of NRF-1 alleviates the
CoCl2-induced decrease in cell viability
To elucidate the role of NRF-1 in hypoxia caused by
CoCl2 in H9C2 cells, H9C2 cells were transfected with
NRF-1 overexpression lentiviral recombinant vector (NRF1-H9C2) or
empty viral vector (pLenti-H9C2). Western blotting revealed that
NRF-1 was upregulated in NRF1-H9C2 cells (Fig. 1A).
Subsequently, NRF1-H9C2 and pLenti-H9C2 cells were
treated with 200 or 400 µM CoCl2 for 6 or 24 h, and
CCK-8 assays were used to detect cell viability. The results
demonstrated that the CoCl2-treated cells for 24 h had
lower survival ability than the untreated cells (Fig. 1B). NRF1-H9C2 cells treated with 400
µM CoCl2 for 6 h demonstrated higher cell viability than
pLenti-H9C2 cells. However, after 24 h treatment, there was no
significant difference between NRF1-H9C2 and pLenti-H9C2 cells.
RT-qPCR was performed to detect the mRNA expression of NRF-1,
NRF-2 and HIF-1α. As described above, CoCl2
can stabilize HIF-1α (33,34). Furthermore, the data demonstrated
that HIF-1α was expressed at low levels in both groups under
normal conditions, whereas treatment with 200 or 400 µM
CoCl2 for 6 h caused HIF-1α mRNA levels to
increase significantly in NRF1-H9C2 and pLenti-H9C2 cells,
suggesting a response to hypoxia. However, it was also identified
that the levels of HIF-1α mRNA in NRF1-H9C2 cells were
higher compared with pLenti-H9C2 cells. One possible reason may be
that NRF-1 possesses a direct regulation on HIF-1a, but
NRF-1 had almost no elevation effect on HIF-1a in
non-CoCl2 treated cells, so another possible mechanism
for the increased levels of HIF-1a in the experiments was
that it was affected by some genes that were activated or
inactivated by CoCl2, and those genes were also
influenced by NRF-1. CoCl2 can simulate hypoxia, which
can also regulate the levels of HIF-1a. Theoretically, the levels
of HIF-1a could be enhanced by CoCl2 in the
experiments of the present study, but it is puzzling that 400 µm
CoCl2 could not promote the levels of HIF-1a as
did 200 µm CoCl2, and that NRF-1 also dropped to a low
levels in 400 µm CoCl2-treated cells. Thus, the increase
of HIF-1a levels caused by CoCl2 existed in
certain conditions: A certain concentration of CoCl2
(200 µm) could cause stress effect of H9C2 cells in a short time (6
h), but when CoCl2 exceeded a certain level (400 µm for
6 h, 200/400 µm for 24 h), it would cause downstream effects on
some genes like NRF-1 and HIF-1a. Therefore NRF-1 and
HIF-1a may have a special regulatory role in the presence of
CoCl2; something which requires further study.
Overexpression of NRF-1 can reduce
apoptosis trigged by CoCl2
The decrease in H9C2 cell viability induced by
CoCl2 may be due to apoptosis. Therefore, the nuclear
morphology of CoCl2-treated cells using Hoechst 33342
staining was first examined. As shown in Fig. 2A, round and weakly stained cells
were observed prior to treatment. However, CoCl2-treated
cells demonstrated clear pyknosis and strong blue staining. The
number of apoptotic nuclei was significantly lower in NRF1-H9C2
cells compared with that in pLenti-H9C2 cells.
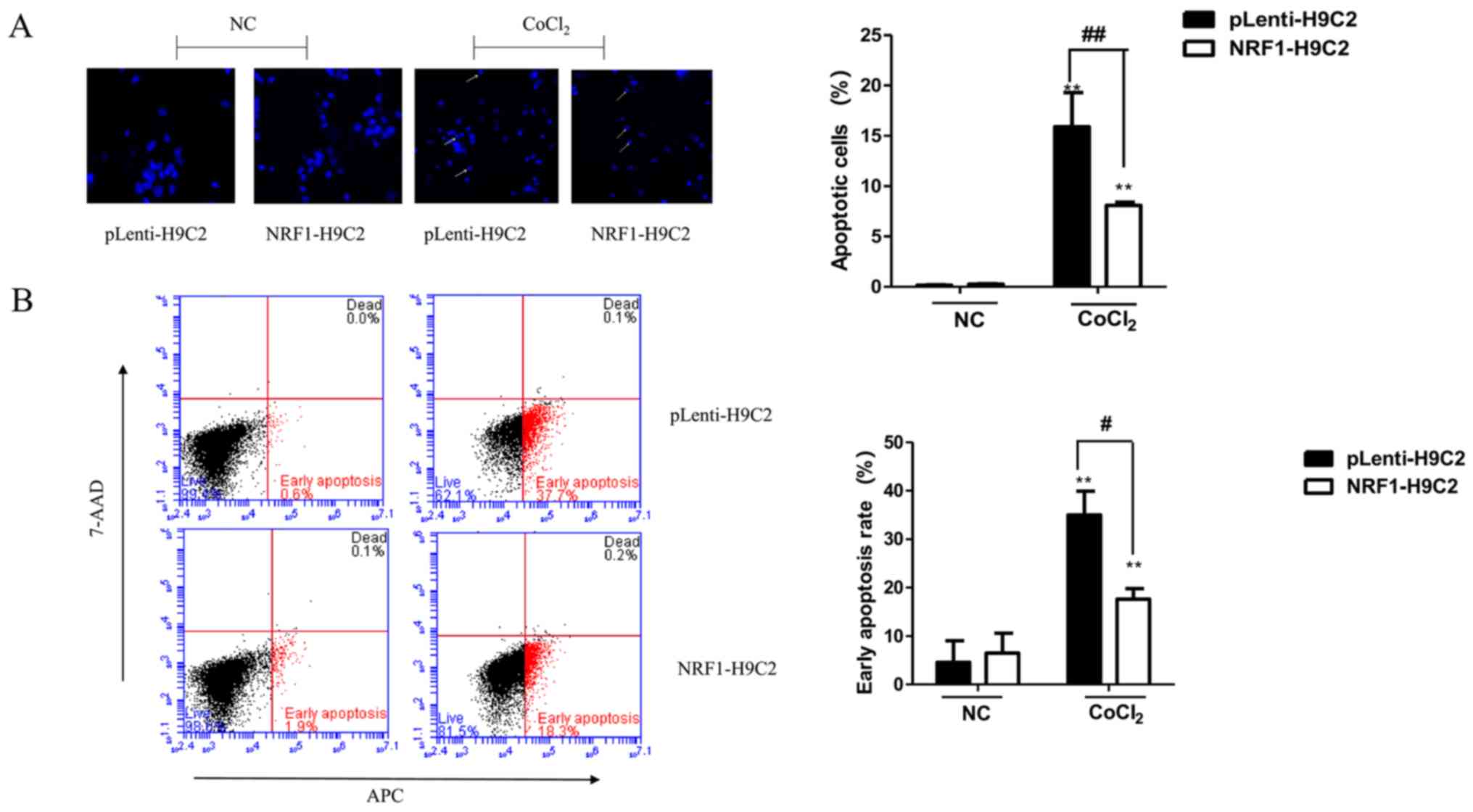 | Figure 2.Effects of CoCl2 on
apoptosis. (A) Changes in nuclear morphology, representative of
apoptosis, in cells induced by CoCl2 (400 µM) for 6 h
were observed by Hoechst 33342 staining in pLenti-H9C2 and
NRF1-H9C2 cells. Cells with intense blue staining in nuclei were
counted. Magnification, ×400. (B) Flow cytometry was used to detect
apoptosis following staining with Annexin V-APC and 7-ADD. (C) mRNA
expression of Bcl-2 and Bax was analyzed by reverse
transcription-quantitative polymerase chain reaction. (D) Protein
expression levels of NRF-1, Bcl-2, Bax and PARP were analyzed by
western blot. **P<0.01 vs. NC; #P<0.05,
##P<0.01 vs. pLenti-H9C2 cells. NC, negative control
(CoCl2 untreated cells); CoCl2, cobalt
chloride; NRF, nuclear respiratory factor; 7-AAD,
7-aminoactinomycin D; APC, allophycocyanin; Bcl-2, B-cell
lymphoma-2; Bax, Bcl-associated X protein; PARP, poly(ADP-ribose)
polymerase 1. |
Subsequently, flow cytometry was performed to
analyze apoptosis in NRF1-H9C2 and pLenti-H9C2 cells. The results
demonstrated that the apoptosis rates of both types of
CoCl2-treated cells were significantly increased,
although NRF1-H9C2 cells exhibited a relatively low rate of
apoptosis (Fig. 2B). The
expression of the Bcl-2 and Bax were also detected (Fig. 2C and D). Notably, in NRF1-H9C2
cells, the expression of the Bcl-2, which encodes an anti-apoptotic
molecule, maintained a higher level compared with that in
pLenti-H9C2 cells. The expression of Bax, which encodes a
pro-apoptotic molecule, was increased in CoCl2-treated
cells; however, there were no obvious differences between NRF1-H9C2
and pLenti-H9C2 cells, which might indicate that NRF-1 has no
inhibitory effect on Bax. The levels of PARP, which is the cleavage
substrate of caspase, were higher in CoCl2-treared
pLenti-H9C2 cells compared with that in NRF1-H9C2 cells. Based on
these findings, the decrease in H9C2 cell viability may be
partially due to the induction of apoptosis by CoCl2,
and the presence of NRF-1 may have an inhibitory effect on
apoptosis.
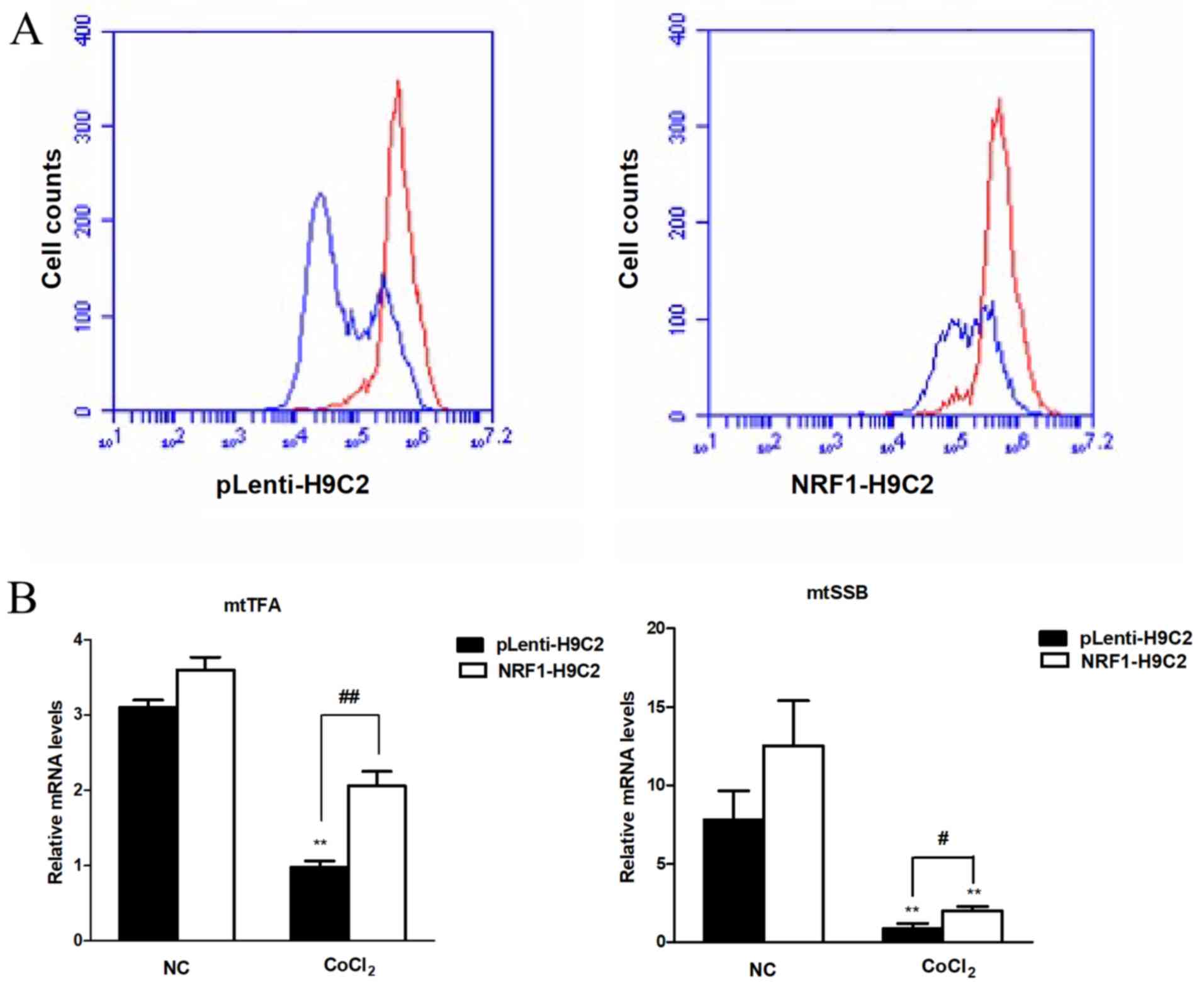
Effects of NRF-1 on MMP in
CoCl2-treated H9C2 cells
Multiple studies have demonstrated that MMP is an
important index of apoptosis (45–48),
MMP plays an important role in mitochondrial function and decline
of MMP caused by stress can lead to mitochondrial dysfunction. In
Fig. 3A, the red curve represented
the untreated cells and the blue curve represented the
CoCl2-treated cells, which suggested the reduction of
MMP level. In normal conditions, MMP in both groups maintained in
high levels. However, changes in MMP level in both groups were
observed following treatment with 400 µM CoCl2 for 6 h,
which exhibited the decline of MMP level, revealing the damaging
effects of CoCl2 on mitochondrial function. The results
of the present study also demonstrated that MMP levels in
pLenti-H9C2 cells was lower compared with NRF1-H9C2 cells. A
previous study identified that apoptosis was accompanied by the
decline of MMP (45), data from
the present study demonstrated that the overexpression of NRF-1
could alleviate the decrease of MMP, suggesting a protective effect
of NRF-1 on mitochondria. In addition, mtTFA and
mtSSB mRNA levels were significantly altered following
treatment with CoCl2, with the mRNA levels remaining
higher in NRF1-H9C2 cells than in pLenti-H9C2 cells. These data
indicated that overexpression of NRF-1 could alleviate the decrease
in MMP, potentially by affecting the expression levels of
mitochondria-associated genes, such as mtTFA and
mtSSB (Fig. 3B).
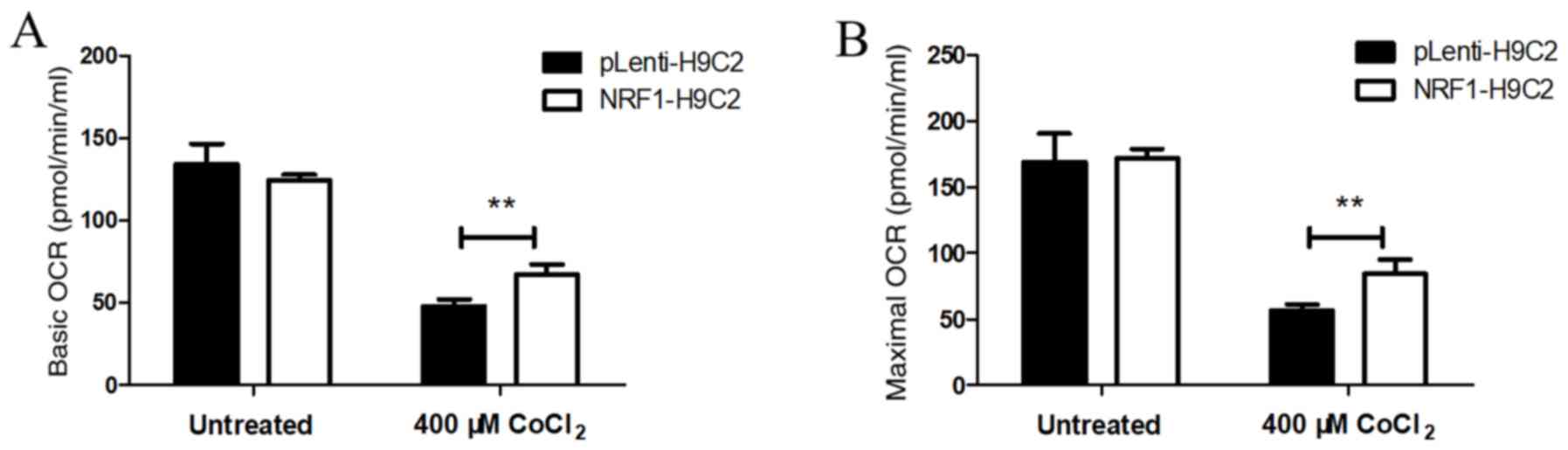
Overexpression of NRF-1 increases the
oxygen consumption in CoCl2-treated H9C2 cells
OCRs were used as an index of mitochondrial
respiratory capacity (49,50). Under normal conditions, OCRs were
similar in NRF1-H9C2 and pLenti-H9C2 cells, suggesting that there
were no differences in mitochondrial respiratory capacity between
the two groups (Fig. 4A and B)
under normal conditions. Following treatment of cells with 400 µM
CoCl2 for 6 h, the basic and maximum OCRs were decreased
in both NRF1-H9C2 and pLenti-H9C2 cells, although the OCR in
NRF1-H9C2 cells was relatively higher than that in pLenti-H9c2
cells. The above results indicated that NRF-1 may enhance the
mitochondrial respiratory capacity to resist
CoCl2-dependent impairment in H9C2 cells.
Discussion
Hypoxia is a common cause of pathological damage to
organs, including the heart, where brief and rapid hypoxia can
cause irreversible damage. The heart is a unique organ that pumps
blood into various tissues, and compared with other parts of the
body, hypoxia in the heart results in serious consequences
(7,8). Cardiomyocytes are the structural and
functional units of the heart (51,52).
Because adult cardiomyocytes lose their ability to proliferate
(52–54), H9C2 cells, a proliferating
cardiomyocyte cell line, has often used to observe the effect of
hypoxia on cardiomyocytes. Many types of hypoxia can be attributed
to the loss of oxygen caused by inadequate tissue perfusion or a
drop in systemic oxygen partial pressure in the arterial blood
(55). As described above,
CoCl2 is a well-known hypoxia-mimetic chemical reagent
that is widely used in many hypoxia studies (56–59).
In the present study, H9C2 cardiomyocytes were exposed to two
different concentrations of CoCl2, which resulted in a
decrease in cell viability consistent with a previous study in
which CoCl2 was reported to behave as a cytotoxic agent
(60,61). The present data also demonstrated
the protective effects of NRF-1 on CoCl2-treated H9C2
cells, an effect that had not been previously reported. Although
multiple studies have reported that NRF-1 has protective effects on
cells under stress conditions (62–64),
these findings led us to explore the role of NRF-1, and
particularly the protective mechanisms of NRF-1 in
CoCl2-treated cells.
NRF-1 is involved in the integration of nuclear and
mitochondrial genes (20,22); however, few studies have evaluated
the protective effects of NRF-1 on CoCl2-induced cell
damage (65). NRF-2, together with
NRF-1, serves a key role in the integration of nuclear and
mitochondrial genomes by directly or indirectly regulating the
expression of nuclear-encoded genes (66–68).
Various studies have focused on NRF-2, which has been described as
a ‘multi-organ protector’ and can protect against many diseases in
a number of organs (69),
including chronic obstructive pulmonary disease, nephrotoxicity
from ischemia-reperfusion injury and cisplatin and cardiomyopathy
induced by smoking (70).
Furthermore, inhibition of NRF-2 can render cells susceptible to
apoptosis (71,72) and has been shown to weaken
endoplasmic reticulum stress-induced apoptosis in some experiments
(73,74). Additionally, NRF-2 is a crucial
regulator of the response to cobalt and can be activated in
response to CoCl2-mediated oxidative stress (75). In the present study, both NRF-2 and
NRF-1 increased in CoCl2-treated H9C2 cells, a previous
study demonstrated that there are four antioxidant-responsive
elements in the promoter region of NRF-1 that can be
regulated by NRF-2, leading to an increase of NRF-1 protein levels
(76); thus, whether the results
of the present study indicated that there exists an interaction
between NRF-1 and NRF-2, or whether their increase does not depend
on each other in CoCl2-treated cells but is a mutual
independence, requires more precise data to elucidate. The findings
of the present study also indicated that the reduced proliferation
of CoCl2-treated H9C2 cells was partially due to the
occurrence of apoptosis, and overexpression of NRF-1 alleviated
CoCl2-induced apoptosis; this pro-survival effect was
consistent with other studies (63,76),
suggesting that NRF-1 has an anti-apoptotic role, similar to NRF-2
(70).
Bcl-2 and Bax are two important apoptotic factors
that are part of the Bcl-2 family, although they are classified
into different groups based on their functions and structures:
Bcl-2 is an anti-apoptotic molecule, whereas Bax is a pro-apoptotic
molecule, an entry point for the intrinsic apoptotic signaling
pathway (77), and can be
activated by various stimuli, ultimately leading to cell death
(77–80). Bax activates caspase-9 (81), regulates the unfolded protein
response by interacting with inositol requiring 1-α (82) and also modulates mitochondrial
function (83). Bax and Bcl-2 are
highly homologous and can form heterodimers (84). A number of studies have suggested
that Bcl-2 is an anti-apoptotic protein (85–88).
Therefore, the opposing roles of Bcl-2 and Bax are important in
studies of apoptosis. Levels of Bax were increased in
CoCl2-treated cells, and Bcl-2 levels reduced in
CoCl2-treated pLenti-H9C2. Notably, no differences were
observed in Bax expression between pLenti-H9C2 and NRF1-H9C2 cells,
suggesting that the inhibitory effects of NRF-1 on apoptosis may be
associated with the transcriptional activation of Bcl-2.
Furthermore, expression levels of the cleaved PARP were lower in
NRF1-H9C2 cells compared with the pLenti vector control following
treatment with CoCl2. These protective effects indicated
that NRF-1 may mediate resistance to apoptosis. Additionally, when
the incubation time was increased to 24 h, the cytoprotective
effects of NRF-1 were not observed, possibly because the notable
decrease in NRF-1 removed the defense against
CoCl2 damage.
The main mechanisms of apoptosis are the
mitochondrial pathway and the death receptor pathway.
Stress-induced mitochondrial dysfunction can severely affect cell
survival and result in apoptotic cell death (89–93).
NRF-1 was originally identified in a study of the promoter activity
of cytochrome c (94,95),
an electron transport vehicle that can shuttle between different
complexes; this feature has become an important target for the
study of transcriptional regulation of various
mitochondria-associated genes (96). Further in-depth studies have
revealed that there are NRF-1 recognition sites in the promoter
regions of several nuclear genes required for the expression of
respiratory genes (97–101). Thus, the anti-apoptotic and
pro-survival effects of NRF-1 on CoCl2-treated H9C2
cells may be associated with the maintenance of mitochondrial
function. Previous studies have demonstrated that MMP has a
critical role in the maintenance of mitochondrial function
(102,103). Disruption of MMP can cause
cytochrome c to enter the cytosol, inducing apoptosis
(104–106). The present findings indicated
that overexpression of NRF-1 reduced the depolarization of MMP
caused by 400 µM CoCl2. In addition, the present results
demonstrated that NRF-1 may enhance the expression of the
mtTFA gene, consistent with previous studies (99). mtTFA is important for the
initiation and transcription of mitochondria-encoded genes
(107–109). These data suggest that in
response to injury caused by CoCl2 and to maintain
adequate mitochondrial functionality, NRF-1 can increase the
expression of mitochondria-associated genes, mtTFA and mtSSB,
thereby regulating mtDNA transcription and replication (110–112), and protecting the cell from
stress.
MMP can effectively promote the synthesis of ATP by
generating electrochemical gradients. It was speculated that
disruption of MMP may inhibit the cellular mitochondrial
respiratory capacity. In the present study, mitochondrial
respiration was investigated by detecting the OCR of
CoCl2-treated NRF1-H9C2 and pLenti-H9C2 cells.
Mitochondrial respiratory potential has been measured as an index
of oxidative stress-mediated mitochondrial dysfunction in previous
studies (49,113). In the present study, there was a
significant decrease in mitochondrial respiration in
CoCl2-treated cells, and overexpression of NRF-1
subsequently enhanced mitochondrial respiration. Thus, these
findings revealed that NRF-1 may be involved in the regulation of
mitochondrial function.
In summary, NRF-1 acted as an important
anti-apoptotic molecule in CoCl2-treated H9C2 cells,
potentially by reducing the decrease in MMP and increasing
mitochondrial respiration to strengthen cellular adaptability in
the context of CoCl2-induced hypoxia.
Acknowledgements
The authors thank the members of the Institute of
Medical Sciences of Ningxia Hui Autonomous Region for providing
excellent technical assistance.
Funding
This project was supported by the National Natural
Science Foundation of China (grant no. 81360042).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
NN, WZ, ZHL and RS contributed to writing the
manuscript and revision. RS and WZ designed and revised the
experiments. NN and ZHL wrote the manuscript and performed cell
experiments. NN, HLS and SMX performed detection of energy
metabolism. ZHL and MXZ prepared the images. MXZ performed the
packaging of the virus. JHY performed flow cytometry assay.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that there are no competing
interests.
References
|
1
|
Ratcliffe P, Koivunen P, Myllyharju J,
Ragoussis J, Bovée JV, Batinic-Haberle I, Vinatier C, Trichet V,
Robriquet F, Oliver L and Gardie B: Update on hypoxia-inducible
factors and hydroxylases in oxygen regulatory pathways: From
physiology to therapeutics. Hypoxia (Auckl). 5:11–20. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Romero JI, Holubiec MI, Tornatore TL,
Rivière S, Hanschmann EM, Kölliker-Frers RA, Tau J, Blanco E,
Galeano P, Rodríguez de Fonseca F, et al: Neuronal damage induced
by perinatal asphyxia is attenuated by postinjury glutaredoxin-2
administration. Oxid Med Cell Longev 2017. 41624652017.
|
|
3
|
Keel M and Trentz O: Pathophysiology of
polytrauma. Injury. 36:691–709. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grifka RG: Cyanotic congenital heart
disease with increased pulmonary blood flow. Pediatr Clin North Am.
46:405–425. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ostadal B, Ostadalova I and Dhalla NS:
Development of cardiac sensitivity to oxygen deficiency:
Comparative and ontogenetic aspects. Physiol Rev. 79:635–659. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bär H, Kreuzer J, Cojoc A and Jahn L:
Upregulation of embryonic transcription factors in right
ventricular hypertrophy. Basic Res Cardiol. 98:285–294. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Murray CJ and Lopez AD: Alternative
projections of mortality and disability by cause 1990–2020: Global
burden of disease study. Lancet. 349:1498–1504. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yach D, Hawkes C, Gould CL and Hofman KJ:
The global burden of chronic diseases: Overcoming impediments to
prevention and control. JAMA. 291:2616–2622. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Akhmedov AT, Rybin V and Marín-García J:
Mitochondrial oxidative metabolism and uncoupling proteins in the
failing heart. Heart Fail Rev. 20:227–249. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ingwall JS: ATP and the Heart: An
Overview. Springer; US: 2002, View Article : Google Scholar
|
|
11
|
Ong SB and Hausenloy DJ: Mitochondrial
morphology and cardiovascular disease. Cardiovasc Res. 88:16–29.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ventura-Clapier R, Garnier A, Veksler V
and Joubert F: Bioenergetics of the failing heart. Biochim Biophys
Acta 1813. 1360–1372. 2011.
|
|
13
|
Hausenloy DJ and Ruiz-Meana M: Not just
the powerhouse of the cell: Emerging roles for mitochondria in the
heart. Cardiovasc Res. 88:5–6. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Soubannier V and Mcbride HM: Positioning
mitochondrial plasticity within cellular signaling cascades.
Biochim Biophys Acta 1793. 154–170. 2009.
|
|
15
|
Dominic EA, Ramezani A, Anker SD, Verma M,
Mehta N and Rao M: Mitochondrial cytopathies and cardiovascular
disease. Heart. 100:611–618. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Saraste M: Oxidative phosphorylation at
the fin de siècle. Science. 283:1488–1493. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bianca B and Montagna E: The advances and
new technologies for the study of mitochondrial diseases. Einstein
(Sao Paulo). 14:291–293. 2016.(In English, Portuguese). View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Björkholm P, Harish A, Hagström E, Ernst
AM and Andersson SG: Mitochondrial genomes are retained by
selective constraints on protein targeting. Proc Natl Acad Sci USA.
112:10154–10161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
El-Hattab AW and Fernando S: Mitochondrial
Cardiomyopathies. Front Cardiovasc Med. 3:252016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Evans MJ and Scarpulla RC: NRF-1: A
trans-activator of nuclear-encoded respiratory genes in animal
cells. Genes Dev. 4:1023–1034. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huo L and Scarpulla RC: Mitochondrial DNA
instability and peri-implantation lethality associated with
targeted disruption of nuclear respiratory factor 1 in mice. Mol
Cell Biol. 21:644–654. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Scarpulla RC: Nuclear respiratory factors
and the pathways of nuclear-mitochondrial interaction. Trends
Cardiovasc Med. 6:39–45. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Scarpulla RC: Nuclear control of
respiratory gene expression in mammalian cells. J Cell Biochem.
97:673–683. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Virbasius CA, Virbasius JV and Scarpulla
RC: NRF-1, an activator involved in nuclear-mitochondrial
interactions, utilizes a new DNA-binding domain conserved in a
family of developmental regulators. Genes Dev. 7:2431–2445. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Clayton DA: Transcription and replication
of animal mitochondrial DNAs. Int Rev Cytol. 141:217–232. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choi YS, Kim S, Kyu LH, Lee KU and Pak YK:
In vitro methylation of nuclear respiratory factor-1 binding site
suppresses the promoter activity of mitochondrial transcription
factor A. Biochem Biophy Res Commun. 314:118–122. 2004. View Article : Google Scholar
|
|
27
|
Piantadosi CA and Suliman HB:
Mitochondrial transcription factor A induction by redox activation
of nuclear respiratory factor 1. J Biol Chem. 281:324–333. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang L, Bao Y, Liu Y and Li J:
Downregulation of nuclear respiratory factor-1 contributes to
mitochondrial events induced by benzo(a)pyrene. Environ Toxicol.
29:780–787. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suliman HB, Sweeney TE, Withers CM and
Piantadosi CA: Co-regulation of nuclear respiratory factor-1 by
NFkappaB and CREB links LPS-induced inflammation to mitochondrial
biogenesis. J Cell Sci. 123:2565–2575. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Piantadosi CA and Suliman HB:
Transcriptional regulation of SDHa flavoprotein by nuclear
respiratory factor-1 prevents pseudo-hypoxia in aerobic cardiac
cells. J Biol Chem. 283:10967–10977. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang GL and Semenza GL: Characterization
of hypoxia-inducible factor 1 and regulation of DNA binding
activity by hypoxia. J Biol Chem. 268:21513–21518. 1993.PubMed/NCBI
|
|
32
|
Wang GL and Semenza GL: Desferrioxamine
induces erythropoietin gene expression and hypoxia-inducible factor
1 DNA-binding activity: Implications for models of hypoxia signal
transduction. Blood. 82:3610–3615. 1993.PubMed/NCBI
|
|
33
|
Hervouet E, Pecina P, Demont J, Vojtísková
A, Simonnet H, Houstek J and Godinot C: Inhibition of cytochrome c
oxidase subunit 4 precursor processing by the hypoxia mimic cobalt
chloride. Biochem Biophys Res Commun. 344:1086–1093. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang YB, Wang X, Meister EA, Gong KR, Yan
SC, Lu GW, Ji XM and Shao G: The effects of CoCl2 on
HIF-1α protein under experimental conditions of autoprogressive
hypoxia using mouse models. Int J Mol Sci. 15:10999–11012. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jaakkola P, Mole DR, Tian YM, Wilson MI,
Gielbert J, Gaskell SJ, von Kriegsheim A, Hebestreit HF, Mukherji
M, Schofield CJ, et al: Targeting of HIF-alpha to the von
Hippel-Lindau ubiquitylation complex by O2-regulated
prolyl hydroxylation. Science. 292:468–472. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kaelin WG Jr: The von Hippel-Lindau tumor
suppressor gene and kidney cancer. Clin Cancer Res. 10:6290S–6295S.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Taylor MS: Characterization and
comparative analysis of the EGLN gene family. Gene. 275:125–132.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bae S, Jeong HJ, Cha HJ, Kim K, Choi YM,
An IS, Koh HJ, Lim DJ, Lee SJ and An S: The hypoxia-mimetic agent
cobalt chloride induces cell cycle arrest and alters gene
expression in U266 multiple myeloma cells. Int J Mol Med.
30:1180–1186. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shrivastava K, Ram MS, Bansal A, Singh SS
and Ilavazhagan G: Cobalt supplementation promotes hypoxic
tolerance and facilitates acclimatization to hypobaric hypoxia in
rat brain. High Alt Med Biol. 9:63–75. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shrivastava K, Shukla D, Bansal A, Sairam
M, Banerjee PK and Ilavazhagan G: Neuroprotective effect of cobalt
chloride on hypobaric hypoxia-induced oxidative stress. Neurochem
Int. 52:368–375. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Araya J, Maruyama M, Inoue A, Fujita T,
Kawahara J, Sassa K, Hayashi R, Kawagishi Y, Yamashita N, Sugiyama
E and Kobayashi M: Inhibition of proteasome activity is involved in
cobalt-induced apoptosis of human alveolar macrophages. Am J
Physiol Lung Cell Mol Physiol. 283:L849–L858. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mecklenburgh KI, Walmsley SR, Cowburn AS,
Wiesener M, Reed BJ, Upton PD, Deighton J, Greening AP and Chilvers
ER: Involvement of a ferroprotein sensor in hypoxia-mediated
inhibition of neutrophil apoptosis. Blood. 100:3008–3016. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tello D, Balsa E, Acosta-Iborra B,
Fuertes-Yebra E, Elorza A, Ordóñez Á, Corral-Escariz M, Soro I,
López-Bernardo E, Perales-Clemente E, et al: Induction of the
mitochondrial NDUFA4L2 protein by HIF-1α decreases oxygen
consumption by inhibiting Complex I activity. Cell Metab.
14:768–779. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tischlerova V, Kello M, Budovska M and
Mojzis J: Indole phytoalexin derivatives induce
mitochondrial-mediated apoptosis in human colorectal carcinoma
cells. World J Gastroenterol. 23:4341–4353. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Venkatarame G, owda Saralamma V, Lee HJ,
Hong GE, Park HS, Yumnam S, Raha S, Lee WS, Kim EH, Sung NJ, Lee
SJ, et al: Korean Scutellaria baicalensis Georgi flavonoid
extract induces mitochondrially mediated apoptosis in human gastric
cancer AGS cells. Oncol Lett. 14:607–614. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang L, Gao S, Jiang W, Luo C, Xu M,
Bohlin L, Rosendahl M and Huang W: Antioxidative dietary compounds
modulate gene expression associated with apoptosis, DNA repair,
inhibition of cell proliferation and migration. Int J Mol Sci.
15:16226–16245. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang Y, Yu RY and He QY: Proteomic
analysis of anticancer TCMs targeted at mitochondria. Evid Based
Complement Alternat Med 2015. 5392602015.
|
|
49
|
Kogot-Levin A, Saada A, Leibowitz G,
Soiferman D, Douiev L, Raz I and Weksler-Zangen S: Upregulation of
mitochondrial content in cytochrome c oxidase deficient
fibroblasts. PLoS One. 11:e01654172016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
van der Windt GJ, Everts B, Chang CH,
Curtis JD, Freitas TC, Amiel E, Pearce EJ and Pearce EL:
Mitochondrial respiratory capacity is a critical regulator Of CD8+
T cell memory development. Immunity. 36:68–78. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Paradis AN, Gay MS and Zhang L:
Binucleation of cardiomyocytes: The transition from a proliferative
to a terminally differentiated state. Drug Discovery Today.
19:602–609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li F, Wang X, Capasso JM and Gerdes AM:
Rapid transition of cardiac myocytes from hyperplasia to
hypertrophy during postnatal development. J Mol Cell Cardiol.
28:1737–1746. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ahuja P, Sdek P and Maclellan WR: Cardiac
myocyte cell cycle control in development, disease, and
regeneration. Physiol Rev. 87:521–544. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Burrell JH, Boyn AM, Kumarasamy V, Hsieh
A, Head SI and Lumbers ER: Growth and maturation of cardiac
myocytes in fetal sheep in the second half of gestation. Anat Rec A
Discov Mol Cell Evol Biol. 274:952–961. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ostádal B, Ostádalová I, Kolár F,
Charvátová Z and Netuka I: Ontogenetic development of cardiac
tolerance to oxygen deprivation-possible mechanisms. Physiol Res.
58 Suppl 2:S1–S12. 2009.
|
|
56
|
Dai M, Cui P, Yu M, Han J, Li H and Xiu R:
Melatonin modulates the expression of VEGF and HIF-1 alpha induced
by CoCl2 in cultured cancer cells. J Pineal Res.
44:121–126. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Jiang Y, Satoh K, Watanabe S, Kusama K and
Sakagami H: Inhibition of chlorogenic acid-induced cytotoxicity by
CoCl2. Anticancer Res. 21:3349–3353. 2001.PubMed/NCBI
|
|
58
|
Jung JY, Mo HC, Yang KH, Jeong YJ, Yoo HG,
Choi NK, Oh WM, Oh HK, Kim SH, Lee JH, et al: Inhibition by
epigallocatechin gallate of CoCl2-induced apoptosis in
rat PC12 cells. Life Sci. 80:1355–1363. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen R, Xu J, She Y, Jiang T, Zhou S, Shi
H and Li C: Necrostatin-1 protects C2C12 myotubes from
CoCl2-induced hypoxia. Int J Mol Med. 41:2565–2572.
2018.PubMed/NCBI
|
|
60
|
Chen R, Jiang T, She Y, Xu J, Li C, Zhou
S, Shen H, Shi H and Liu S: Effects of cobalt chloride, a
hypoxia-mimetic agent, on autophagy and atrophy in skeletal C2C12
myotubes. Biomed Res Int 2017. 70975802017.
|
|
61
|
Rovetta F, Stacchiotti A, Faggi F,
Catalani S, Apostoli P, Fanzani A and Aleo MF: Cobalt triggers
necrotic cell death and atrophy in skeletal C2C12 myotubes. Toxicol
Appl Pharmacol. 271:196–205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Bartz RR, Suliman HB and Piantadosi CA:
Redox mechanisms of cardiomyocyte mitochondrial protection. Front
Physiol. 6:2912015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chen L, Kwong M, Lu R, Ginzinger D, Lee C,
Leung L and Chan JY: Nrf1 is critical for redox balance and
survival of liver cells during development. Mol Cell Biol.
23:4673–4686. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Doerks T, Copley RR, Schultz J, Ponting CP
and Bork P: Systematic identification of novel protein domain
families associated with nuclear functions. Genome Res. 12:47–56.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Saxena S, Shukla D, Saxena S, Khan YA,
Singh M, Bansal A, Sairam M and Jain SK: Hypoxia preconditioning by
cobalt chloride enhances endurance performance and protects
skeletal muscles from exercise-induced oxidative damage in rats.
Acta Physiol (Oxf). 200:249–263. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Priya A, Johar K and Wong-Riley M: Nuclear
respiratory factor 2 regulates the expression of the same NMDA
receptor subunit genes as NRF-1: Both factors act by a concurrent
and parallel mechanism to couple energy metabolism and synaptic
transmission. Biochim Biophys Acta 1833. 48–58. 2013.
|
|
67
|
Priya A, Johar K, Nair B and Wong-Riley
MT: Nuclear respiratory factor 2 regulates the transcription of
AMPA receptor subunit GluA2 (Gria2). Biochim Biophys Acta 1843.
3018–3028. 2014.
|
|
68
|
Vercauteren K, Gleyzer N and Scarpulla RC:
PGC-1-related coactivator complexes with HCF-1 and NRF-2beta in
mediating NRF-2(GABP)-dependent respiratory gene expression. J Biol
Chem. 283:12102–12111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Lee JM, Li J, Johnson DA, Stein TD, Kraft
AD, Calkins MJ, Jakel RJ and Johnson JA: Nrf2, a multi-organ
protector? FASEB J. 19:1061–1066. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Liby KT and Sporn MB: Synthetic oleanane
triterpenoids: Multifunctional drugs with a broad range of
applications for prevention and treatment of chronic disease.
Pharmacol Rev. 64:972–1003. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Arlt A, Sebens S, Krebs S, Geismann C,
Grossmann M, Kruse ML, Schreiber S and Schäfer H: Inhibition of the
Nrf2 transcription factor by the alkaloid trigonelline renders
pancreatic cancer cells more susceptible to apoptosis through
decreased proteasomal gene expression and proteasome activity.
Oncogene. 32:4825–4835. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Arlt A, Bauer I, Schafmayer C, Tepel J,
Müerköster SS, Brosch M, Röder C, Kalthoff H, Hampe J, Moyer MP, et
al: Increased proteasome subunit protein expression and proteasome
activity in colon cancer relate to an enhanced activation of
nuclear factor E2-related factor 2 (Nrf2). Oncogene. 28:3983–3996.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Calvert JW, Elston M, Nicholson CK,
Gundewar S, Jha S, Elrod JW, Ramachandran A and Lefer DJ: Genetic
and pharmacologic hydrogen sulfide therapy attenuates
ischemia-induced heart failure in mice. Circulation. 122:11–19.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Malhotra D, Thimmulappa R, Vij N,
Navas-Acien A, Sussan T, Merali S, Zhang L, Kelsen SG, Myers A,
Wise R, et al: Heightened endoplasmic reticulum stress in the lungs
of patients with chronic obstructive pulmonary disease: The role of
Nrf2-regulated proteasomal activity. Am J Respir Crit Care Med.
180:1196–1207. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Permenter MG, Dennis WE, Sutto TE, Jackson
DA, Lewis JA and Stallings JD: Exposure to cobalt causes
transcriptomic and proteomic changes in two rat liver derived cell
lines. PLoS One. 8:e837512013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Piantadosi CA, Carraway MS, Babiker A and
Suliman HB: Heme oxygenase-1 regulates cardiac mitochondrial
biogenesis via Nrf2-mediated transcriptional control of nuclear
respiratory factor-1. Circ Res. 103:1232–1240. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Liu Z, Ding Y, Ye N, Wild C, Chen H and
Zhou J: Direct activation of Bax protein for cancer therapy. Med
Res Rev. 36:313–341. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Brenner D and Mak TW: Mitochondrial cell
death effectors. Curr Opin Cell Biol. 21:871–877. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Pagliari LJ, Kuwana T, Bonzon C, Newmeyer
DD, Tu S, Beere HM and Green DR: The multidomain proapoptotic
molecules Bax and Bak are directly activated by heat. Proc Natl
Acad Sci USA. 102:17975–17980. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Yin C, Knudson CM, Korsmeyer SJ and Van
DT: Bax suppresses tumorigenesis and stimulates apoptosis in vivo.
Nature. 385:637–640. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Li P, Nijhawan D, Budihardjo I,
Srinivasula SM, Ahmad M, Alnemri ES and Wang X: Cytochrome c
and dATP-dependent formation of Apaf-1/caspase-9 complex initiates
an apoptotic protease cascade. Cell. 91:479–489. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Rampino N, Yamamoto H, Ionov Y, Li Y,
Sawai H, Reed JC and Perucho M: Somatic frameshift mutations in the
BAX gene in colon cancers of the microsatellite mutator phenotype.
Science. 275:967–969. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Karbowski M, Norris KL, Cleland MM, Jeong
SY and Youle RJ: Role of Bax and Bak in mitochondrial
morphogenesis. Nature. 443:658–662. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Walensky LD and Gavathiotis E: BAX
unleashed: The biochemical transformation of an inactive cytosolic
monomer into a toxic mitochondrial pore. Trends Biochem Sci.
36:642–652. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Adams JM and Cory S: Bcl-2-regulated
apoptosis: Mechanism and therapeutic potential. Curr Opin Immunol.
19:488–496. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Hata AN, Engelman JA and Faber AC: The
BCL2 family: Key mediators of the apoptotic response to targeted
anticancer therapeutics. Cancer Discov. 5:475–487. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Kale J, Liu Q, Leber B and Andrews DW:
Shedding light on apoptosis at subcellular membranes. Cell.
151:1179–1184. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wood WG, Igbavboa U, Muller WE and Eckert
GP: Statins, Bcl-2 and apoptosis: Cell death or cell protection?
Mol Neurobiol. 48:308–314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Gogvadze V and Orrenius S: Mitochondrial
regulation of apoptotic cell death. Chem Biol Interact. 163:4–14.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Jeong SY and Seol DW: The role of
mitochondria in apoptosis. BMB Rep. 41:11–22. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Park JB, Chang H and Kim KW: Expression of
Fas ligand and apoptosis of disc cells in herniated lumbar disc
tissue. Spine (Phila Pa 1976). 26:618–621. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Park JB, Kim KW, Han CW and Chang H:
Expression of Fas receptor on disc cells in herniated lumbar disc
tissue. Spine (Phila Pa 1976). 26:142–146. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Parsons MJ and Green DR: Mitochondria in
cell death. Essays Biochem. 47:99–114. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Evans MJ and Scarpulla RC: Interaction of
nuclear factors with multiple sites in the somatic cytochrome
c promoter. Characterization of upstream NRF-1, ATF, and
intron Sp1 recognition sequences. J Biol Chem. 264:14361–14368.
1989.PubMed/NCBI
|
|
95
|
Evans MJ and Scarpulla RC: Both upstream
and intron sequence elements are required for elevated expression
of the rat somatic cytochrome c gene in COS-1 cells. Mol
Cell Biol. 8:35–41. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Scarpulla RC and Wu R: Nonallelic members
of the cytochrome c multigene family of the rat may arise
through different messenger RNAs. Cell. 32:473–482. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Chau CM, Evans MJ and Scarpulla RC:
Nuclear respiratory factor 1 activation sites in genes encoding the
gamma-subunit of ATP synthase, eukaryotic initiation factor 2
alpha, and tyrosine aminotransferase. Specific interaction of
purified NRF-1 with multiple target genes. J Biol Chem.
267:6999–7006. 1992.PubMed/NCBI
|
|
98
|
Kelly DP and Scarpulla RC: Transcriptional
regulatory circuits controlling mitochondrial biogenesis and
function. Genes Dev. 18:357–368. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Scarpulla RC: Nuclear activators and
coactivators in mammalian mitochondrial biogenesis. Biochim Biophys
Acta 1576. 1–14. 2002.
|
|
100
|
Scarpulla RC: Transcriptional activators
and coactivators in the nuclear control of mitochondrial function
in mammalian cells. Gene. 286:81–89. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Scarpulla RC: Nuclear control of
respiratory chain expression by nuclear respiratory factors and
PGC-1-related coactivator. Ann NY Acad Sci 1147. 321–334. 2008.
View Article : Google Scholar
|
|
102
|
Perry SW, Norman JP, Barbieri J, Brown EB
and Gelbard HA: Mitochondrial membrane potential probes and the
proton gradient: A practical usage guide. Biotechniques. 50:98–115.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Zhang BB, Wang DG, Guo FF and Xuan C:
Mitochondrial membrane potential and reactive oxygen species in
cancer stem cells. Fam Cancer. 14:19–23. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Henry-Mowatt J, Dive C, Martinou JC and
James D: Role of mitochondrial membrane permeabilization in
apoptosis and cancer. Oncogene. 23:2850–2860. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Lakhani SA, Masud A, Kuida K, Porter GA
Jr, Booth CJ, Mehal WZ, Inayat I and Flavell RA: Caspases 3 and 7:
Key mediators of mitochondrial events of apoptosis. Science.
311:847–851. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Ly JD, Grubb DR and Lawen A: The
mitochondrial membrane potential (deltapsi(m)) in apoptosis; an
update. Apoptosis. 8:115–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Campbell CT, Kolesar JE and Kaufman BA:
Mitochondrial transcription factor A regulates mitochondrial
transcription initiation, DNA packaging, and genome copy number.
Biochim Biophys Acta 1819. 921–929. 2012.
|
|
108
|
Fisher RP and Clayton DA: Purification and
characterization of human mitochondrial transcription factor 1. Mol
Cell Biol. 8:3496–3509. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Parisi MA and Clayton DA: Similarity of
human mitochondrial transcription factor 1 to high mobility group
proteins. Science. 252:965–969. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Maier D, Farr CL, Poeck B, Alahari A,
Vogel M, Fischer S, Kaguni LS and Schneuwly S: Mitochondrial
single-stranded DNA-binding protein is required for mitochondrial
DNA replication and development in Drosophila melanogaster.
Mol Biol Cell. 12:821–830. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Takamatsu C, Umeda S, Ohsato T, Ohno T,
Abe Y, Fukuoh A, Shinagawa H, Hamasaki N and Kang D: Regulation of
mitochondrial D-loops by transcription factor A and single-stranded
DNA-binding protein. EMBO Rep. 3:451–456. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Van Dyck E, Foury F, Stillman B and Brill
SJ: A single-stranded DNA binding protein required for
mitochondrial DNA replication in S. cerevisiae is homologous to E.
coli SSB. EMBO J. 11:3421–3430. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Mali VR, Pan G, Deshpande M, Thandavarayan
RA, Xu J, Yang XP and Palaniyandi SS: Cardiac mitochondrial
respiratory dysfunction and tissue damage in chronic hyperglycemia
correlate with reduced aldehyde dehydrogenase-2 activity. PLoS One.
11:e01631582016. View Article : Google Scholar : PubMed/NCBI
|