Introduction
The development of tissue engineering has aided the
identification of novel approaches in the repair and treatment of
bone damage. The bone tissue engineering process for repair
involves the growth of seed cells using scaffold materials,
frequently in the presence of cytokines, to generate
tissue-engineered bone, and its potential use in clinical settings
has been investigated (1,2). At present, the development of
orthopaedic tissue engineering scaffolds is progressing rapidly,
with the production of porous tantalum, degradable magnesium,
hydroxyapatite and high polymer materials (3). Scaffolds provide seed cells a spatial
structure for adhesion, proliferation and differentiation, and
regulate tissue regeneration and reconstruction (1,4,5).
Growth factors are biologically active cytokines
that stimulate the growth of cells. The binding of growth factors
to cell membrane receptors is a specific, high-affinity process
that regulates the growth and development of cells. Growth factors
are used to promote the bone formation ability of an implant via
surface modifications without altering the overall physical
properties of the implant, thereby regulating the bone-repair
microenvironment. For example, bone morphogenetic protein 2 (BMP-2)
is frequently used for surface modification studies in implants and
is used primarily in the treatment of fresh fractures, bone defects
and avascular necrosis of the femoral head (6). It was demonstrated that BMP-2
promoted the proliferation, migration and osteogenic
differentiation of mesenchymal stem cells (MSC), and induced bone
formation in vivo (7,8). In
addition, vascular endothelial growth factor (VEGF), an important
regulator of vascular development and angiogenesis, serves a
crucial role in skeletal development. Poh et al (9) immobilised VEGF on the surface of
titanium alloys. VEGF-modified implants increased the survival and
proliferation of endothelial cells, and promoted the
differentiation of human MSCs into endothelial cells, aiding
angiogenesis and the formation of novel bone tissue (10,11).
These studies demonstrated that the use of growth factors to modify
the surface of the implant promoted the osteointegration of the
implant material.
Transforming growth factor β1 (TGF-β1) is the most
abundant cytokine in bone cells (12). Osteoblasts secrete large quantities
of TGF-β1, which serves an important role in the process of bone
turnover (13). A member of the
TGF-β superfamily, TGF-β1, promotes the proliferation and
osteogenic differentiation of bone cells (14,15).
Additionally, it exhibits a notable chemotactic effect on human
osteoblasts; this effect is particularly evident at low
concentrations of TGF-β1 (16). It
was demonstrated that TGF-β1 promoted the absorption of
osteoclasts, and that novel bone formation was stimulated by in
vivo injection of TGF-β1. Furthermore, it was revealed that
TGF-β1 released by osteoclasts stimulated novel bone formation and
reduced the extent of subsequent bone resorption (17). TGF-β1 may exhibit therapeutic
potential in wound healing (18).
Previously, Chen et al (19) applied TGF-β1 to porous titanium
loaded with gelatine microspheres, and observed that TGF-β1
promoted the adhesion, proliferation and differentiation of MG63
osteosarcoma cells. Lamberg et al (20) demonstrated that the localised
delivery of TGF-β1 enhanced the stability of titanium implants and
promoted attachment. TGF-β1 has received increasing attention
regarding the modification of scaffold materials.
TGF-β1 is involved in a series of physiological and
biochemical processes, due to the complexity of its regulation of
cell biological activity and the levels of protein phosphorylation
observed in numerous associated signalling pathways. TGF-β
signalling pathways typically involve TGF-β receptor-mediated
suppressor of mothers against decapentaplegic (Smad)-dependent or
-independent signalling (21). The
former promotes the proliferation, chemotaxis and differentiation
of bone cells, and reduces the secretion of receptor activator of
nuclear factor κ-B ligand/osteoprotegerin via TGF-β-Smad signalling
to inhibit osteoclast differentiation (22). The latter affects osteoblasts via
mitogen-activated protein kinase (MAPK) kinase (MKK)-p38MAPK or
MKK-extracellular signal-regulated kinase 1/2 signalling (23). TGF-β suppresses Runt-related
transcription factor 2 (Runx2) to inhibit the differentiation of
osteoblasts (24). Furthermore, an
association between the TGF-β family and the phosphatidylinositol
3-kinase/protein kinase B (PI3K/AKT) signalling pathway has been
reported (25). The PI3K/AKT
signalling pathway has been identified as important in the
survival, growth, proliferation and differentiation of cells
(21–23). A recent study reported that
inhibition of the PI3K/AKT signalling pathway promoted osteoblast
injury (26). Additionally,
PI3K/AKT activity promotes cell survival via the downstream mTOR
pathway and participates in the metabolism, proliferation and
angiogenesis of cells (27,28).
At present, the effects of TGF-β1 signalling on the
migration and mineralisation of human osteoblasts remain unclear.
In the present study, the effects of TGF-β1 on osteoblast migration
and mineralisation were investigated and the role of the
PI3K/AKT/mTOR/S6K1 signalling pathway was determined.
Materials and methods
Cell culture
Human foetal osteoblast hFOB1.19 cells (American
Type Culture Collection, Manassas, VA, USA) were incubated at 37°C
with 5% CO2 and cultured in Dulbecco's Modified Eagle's
medium (DMEM; HyClone; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% fetal bovine serum (HyClone; GE Healthcare
Life Sciences), 100 µg/ml streptomycin and 100 IU/ml penicillin.
Cells of passages 3–6 were selected for experiments.
Cell proliferation assay
hFOB1.19 osteoblasts (5×103 cells/well)
were cultured in a 96-well plate for 24 h at 37°C. Then, the medium
was replaced and supplemented with TGF-β1 (0, 0.5, 1, 2, 5 and 10
ng/ml; PeproTech China, Suzhou, China). Following incubation at
37°C for 1, 3 or 5 days, 10 µl Cell Counting Kit-8 (CCK-8) reagent
(Dojindo Molecular Technologies, Inc., Kumamoto, Japan) was added
to wells. Following incubation for a further 2 h at 37°C, the
absorbance at 450 nm was detected (650 nm reference) using an
iMark™ microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Cell cycle measurement by
fluorescence-activated cell sorting
hFOB1.19 human osteoblasts (5×104
cells/well) were cultured in a 6-well plate for 24 h at 37°C. Then,
the medium was replaced and supplemented with TGF-β1 (0, 0.5, 1, 2,
5 and 10 ng/ml) for 24 h at 37°C. The cells were subsequently
washed with PBS, removed by 0.25% trypsin/EDTA solution for 5 min
at 37°C and centrifuged at 282 × g for 5 min at room temperature.
Subsequently, cells were washed twice with PBS and fixed in 70%
ethanol for ≥1 h at −20°C. They were washed twice to remove the
ethanol, and propidium iodide/RNase Staining Buffer (BD
Biosciences, Franklin Lakes, NJ, US) was added to the cells.
Following incubation for 15 min at 37°C, cells were analyzed using
a flow cytometer (BD FACSDiva software; version 7.0; BD
Biosciences) within 1 h. The DNA content of cells was measured by
flow cytometry to determine the cell cycle stage of cells.
Alkaline phosphatase (ALP)
staining
ALP staining was performed to evaluate
osteoblastogenic activity (29).
hFOB1.19 osteoblasts (5×104 cells/well) were cultured in
a 6-well plate and treated with TGF-β1 (0, 0.5, 1, 2, 5 and 10
ng/ml), in the presence or absence of a PI3K/AKT inhibitor (10 µM
LY294002; PeproTech, Inc., Rocky Hill, NJ, USA) or an inhibitor of
mammalian target of rapamycin/S6 kinase 1 (mTOR/S6K1; 100 nM
rapamycin) (30). Following
incubation for 7 days at 37°C, osteoblasts were washed with PBS two
times, fixed in 4% paraformaldehyde (PFA) for 10 min at room
temperature and washed with deionised water twice. Cells were
incubated with a 5-bromo-4-chloro-3-indolyl-phosphate/nitro blue
tetrazolium (BCIP/NBT) ALP colour development kit (Beyotime
Institute of Biotechnology, Haimen, China) for 1 h at 37°C and
washed with deionised water two times. Then, images were acquired
using a phase contrast microscope equipped with a charge-coupled
device (CCD) camera (magnification, ×100). Image-Pro Plus 6.0
(Media Cybernetics, Inc., Rockville, MD, USA) was used to evaluate
the percentage of the total stained area.
Alizarin red S staining
Alizarin red S staining was performed to evaluate
the extent of Ca2+ deposition in the extracellular
matrix. hFOB1.19 human osteoblasts (5×104 cells/well)
were cultured in a 6-well plate and treated with TGF-β1 (0.5, 1, 2,
5 and 10 ng/ml), with or without 10 µM LY294002 or 100 nM
rapamycin. Negative control (NC) cells were not treated with the
compounds. Following incubation for 14 days at 37°C, osteoblasts
were rinsed with PBS twice, fixed in 4% PFA for 10 min at room
temperature and washed with deionised water twice. Cells were
incubated with 1% (w/v) Alizarin red at pH 4.4 for 40 min at 37°C
and subsequently washed with deionised water two times. Then,
images were captured using a CCD camera-equipped phase contrast
microscope (magnification, ×100). The Alizarin-positive area was
evaluated as a percentage of the total area using Image-Pro Plus
6.0.
Transwell assay
A migration assay was conducted using 24-well
Transwell inserts (8-µm pores; Costar; Corning, Inc., Corning, NY,
USA). The cells were treated with TGF-β1 (0, 0.5, 1, 2, 5 and 10
ng/ml), with or without 10 µM LY294002 or 100 nM rapamycin. In the
assay, 5×103 hFOB1.19 osteoblast cells cultured with
DMEM containing 0.1% FBS (200 µl) were seeded in the upper
chambers, and DMEM containing 1% FBS (HyClone; GE Healthcare Life
Sciences) (500 µl) was added to the lower chambers. Following
incubation for 24 h at 37°C, the cells on the upper surface of the
membrane were gently removed using a cotton swab. Cells penetrating
to the lower surface of the inserts were fixed with methanol for 10
min at room temperature, stained with 0.1% crystal violet for 10
min at room temperature and counted using a light microscope
(magnification, ×100). Five randomly-selected fields were used to
count the migrating cells in each well. Data are presented as the
mean ± standard error of the mean for five randomly-selected fields
per sample. Assays were independently performed in triplicate.
Scratch-wound assay
hFOB1.19 osteoblasts (5×104 cells/well)
were cultured in a 6-well plate and treated with TGF-β1 (0, 0.5, 1,
2, 5 or 10 ng/ml), with or without 10 µM LY294002 or 100 nM
rapamycin at 37°C with 5% CO2. The cell layer was
wounded using a 200-µl pipette tip. The scratched wells were washed
with PBS three times and cultured in serum-free medium for 24 h at
37°C. The initial wounding and cell movement in the scratched area
were monitored using a CCD camera-equipped phase contrast
microscope (magnification, ×100) to quantify the number of cells
that had migrated into the wounded area within 24 h.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
hFOB1.19 human osteoblasts (5×104
cells/well) were cultured in a 6-well plate and treated with 1
ng/ml TGF-β1, 10 µM LY294002 and/or 100 nM rapamycin. Following
incubation for 14 days, total RNA isolation was performed using an
RNeasy Mini kit (Qiagen, Inc., Valencia, CA, USA), according to the
manufacturer's protocols. The purity and concentration of RNA was
determined via spectrophotometry at 260 nm and 260/280 nm. The
extracted RNA was reverse transcribed using a PrimeScript™ II first
Strand cDNA Synthesis kit (Takara Biotechnology Co., Ltd., Dalian,
China). RT reactions were performed using a RT-qPCR System
(Eppendorf, Hamburg, Germany) with the following thermocycling
conditions: 10 min at 37°C, 30 min at 42°C and 5 min at 95°C The
primers [transcription factor Sp7 (Osterix), Runx2, osteocalcin
(OCN), osteopontin (OPN), matrix metalloproteinase (MMP)-2, MMP-9
and β-actin] were synthesised by Sangon Biotech Co., Ltd.
(Shanghai, China) and are presented in Table I. qPCR was conducted using an ABI
QuantStudio™ 7 Flex (Applied Biosystems; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) and SYBR® Green PCR master mix
(Thermo Fisher Scientific, Inc.). Reactions were incubated in a
MicroAmp® 96-well reaction plate (Thermo Fisher
Scientific, Inc.) at 95°C for 10 min, followed by 40 cycles at 95°C
for 15 sec and at 60°C for 1 min. β-actin was used as an internal
standard. Data were analysed using QuantStudio Real-Time PCR
Software (version 2.1; Thermo Fisher Scientific, Inc.). mRNA
expression levels were normalised to β-actin and calculated using
the 2−ΔΔCq method (31). All samples were analysed three
times.
 | Table I.Primer sequences for Osterix, Runx2,
OC, OPN, MMP-2, MMP-9 and β-actin. |
Table I.
Primer sequences for Osterix, Runx2,
OC, OPN, MMP-2, MMP-9 and β-actin.
| Gene | Sequence |
|---|
| Osterix | Forward:
5′-GCGGCAAGGTGTATGGCAAGG-3′ |
|
| Reverse:
5′-GCAGAGCAGGCAGGTGAACTTC-3′ |
| Runx2 | Forward:
5′-AACAGCAGCAGCAGCAGCAG-3′ |
|
| Reverse:
5′-GCACCGAGCACAGGAAGTTGG-3′ |
| OCN | Forward:
5′-CAGGCGCTACCTGTATCAATGGC-3′ |
|
| Reverse:
5′-GCCGATGTGGTCAGCCAACTC-3′ |
| OPN | Forward:
5′-AGCGAGGAGTTGAATGGTGCATAC-3′ |
|
| Reverse:
5′-AATCTGGACTGCTTGTGGCTGTG-3′ |
| MMP-2 | Forward:
5′-GCCTCTCCTGACATTGACCTTGG-3′ |
|
| Reverse:
5′-CACCACGGATCTGAGCGATGC-3′ |
| MMP-9 | Forward:
5′-TCCTGGTGCTCCTGGTGCTG-3′ |
|
| Reverse:
5′-CTGCCTGTCGGTGAGATTGGTTC-3′ |
| β-actin | Forward:
5′-AGCCATGTACGTTGCTATCCA-3′ |
|
| Reverse:
5′-ACCGGAGTCCATCACGATG-3′ |
Western blot analysis
Osteoblasts were inoculated into 10-cm culture
dishes. Following culture for 3 days at 37°C with 5%
CO2, cells were treated with 1 ng/ml TGF-β1 or
untreated. The cells were incubated for 24 h and then washed three
times with 4°C PBS for phosphatase inhibition. A mixture of the
PBS, protease inhibitor and radioimmunoprecipitation assay lysate
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was used to extract
total protein from the cells. Total protein concentration was
determined using a Bradford assay kit (Sangon Biotech Co., Ltd.).
All samples were mixed with 5X loading buffer and boiled in water
at 100°C for 10 min. A total of 15 µg protein was loaded in each
lane. Proteins were separated by 2% SDS-PAGE for 2 h and
transferred to 0.2-µm polyvinylidene difluoride (PVDF) membranes
(EMD Millipore, Billicera, MA, USA) at 80 V for 1 h. Following
blocking for 1.5 h at room temperature in 3% bovine serum albumin
(Sigma-Aldrich; Merck KGaA) in PBS, membranes were cut according to
the molecular weight of the pre-stained marker protein and
incubated overnight at 4°C with the following primary antibodies
(all 1:1,000; Cell Signaling Technology, Inc., Danvers, MA, USA):
PI3K (cat. no. D32A5); phosphorylated (p)-PI3K (cat. no. 4228T);
AKT (cat. no. C67E7); p-AKT (cat. no. D9E); mTOR (cat. no. 7C10);
p-mTOR (cat. no. D9C2) and GAPDH (cat. no. D16H11). The PVDF
membranes were washed three times in TBS-Tween 20 (1×TBS, 0.1%
Tween 20) for 10 min/wash. Membranes were then incubated with a
secondary antibody (anti-rabbit immunoglobulin G, horseradish
peroxidase-labeled; cat no. 7074; 1:7,500; Cell Signaling
Technology, Inc.) for 45 min at room temperature and washed three
times with TBS-T (10 min/wash). Protein bands were visualised via
enhanced chemiluminescence (Thermo Fisher Scientific, Inc.) and
analysed via GIS gel image analysis system photography (Tanon
Science and Technology Co., Ltd., Shanghai, China).
Statistical analyses
All experiments were repeated at least three times.
Unless otherwise stated, all data were presented as the mean ±
standard deviation. Differences between groups were determined by
performing analyses of variance followed by the Least Significant
Difference multiple comparisons test. Data were analysed using SPSS
version 20.0 for Windows (IBM Corp., Armonk, NY, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
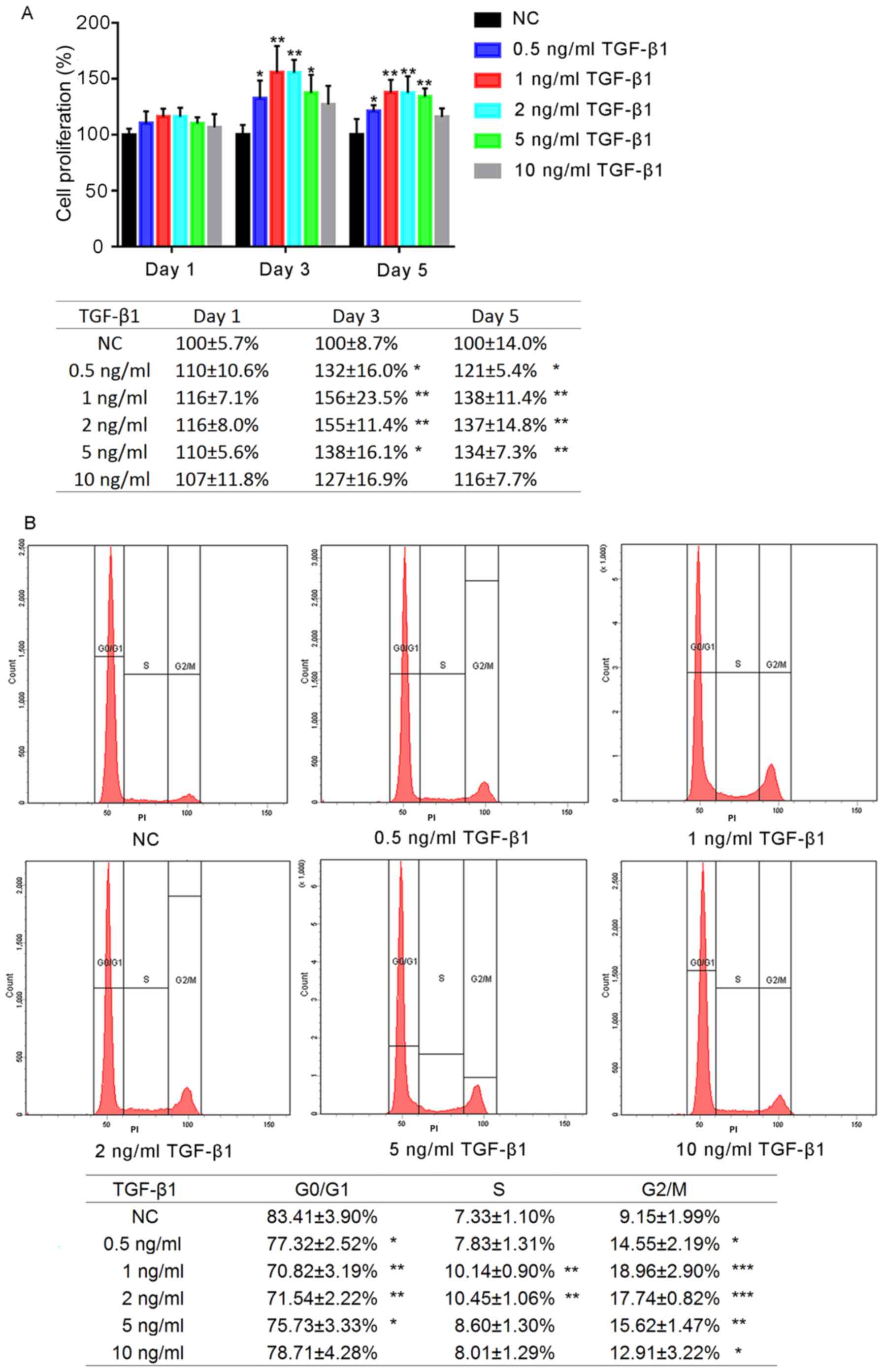
Effects of TGF-β1 on the proliferation
of hFOB1.19 human osteoblast cells
The effects of a range of concentrations of TGF-β1
on the proliferation of hFOB1.19 human osteoblasts were determined
after 1, 3 and 5 days of treatment (Fig. 1A). No significant differences were
observed between the experimental groups following 1 day of
treatment; however, by the third day of treatment, the
proliferation of cells treated with TGF-β1 (0.5, 1, 2 and 5 ng/ml)
was significantly increased compared with the negative control
(NC). Additionally, by day 5, the proliferation of cells in the
0.5, 1, 2 and 5 ng/ml TGF-β1 groups was significantly increased
compared with the NC group (P<0.05); however, no significant
differences was reported following treatment for 5 days with 10
ng/ml TGF-β1 (P>0.05). Proliferation was most markedly increased
in the 1 and 2 ng/ml TGF-β1 groups; no significant difference was
observed between the 1 and 2 ng/ml TGF-β1-treatment groups
(P>0.05).
Effects on hFOB1.19 osteoblast cell
cycle following TGF-β1 treatment
The effects of various concentrations of TGF-β1 on
the hFOB1.19 cell cycle were analysed using flow cytometry. It was
revealed that the proportion of osteoblasts in G0/G1 phase
following treatment for 24 h with TGF-β1 (0.5, 1, 2 and 5 ng/ml)
was significantly decreased compared with the control, whereas that
in G2/M phase was significantly increased (P<0.05; Fig. 1B). The increased percentage of G2/M
cells in the 1 ng/ml group was most notable. Treatment with 10
ng/ml TGF-β1 did not significantly alter the proportion of cells in
G0/G1 and S phase (P>0.05).
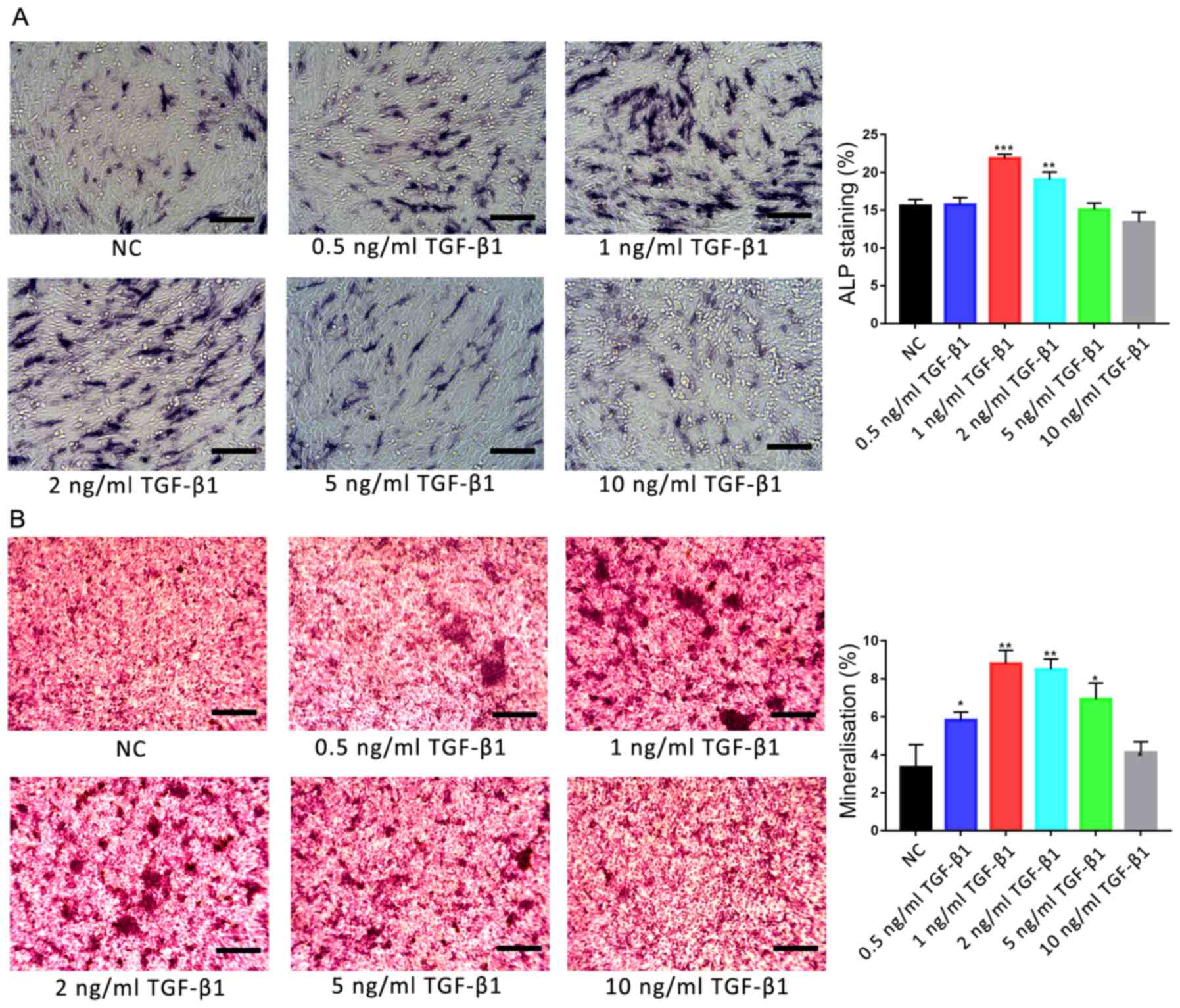
Effects of TGF-β1 on osteogenic
induction and the motility of hFOB1.19 cells
Osteoblasts were treated with various concentrations
of TGF-β1 (0, 0.5, 1, 2, 5 and 10 ng/ml), and the osteogenic
activities and motility of cells were investigated using ALP and
Alizarin red S staining, and scratch-wound and Transwell assays,
respectively. ALP staining revealed that only 1 and 2 ng/ml TGF-β1
significantly promoted ALP activity in osteoblasts compared with
the control group (P<0.01; Fig.
2A). Additionally, Alizarin red S staining revealed that the
degree of mineralisation was significantly increased following
treatment with 1 and 2 ng/ml TGF-β1 compared with the control group
(P<0.01; Fig. 2B). Furthermore,
the degree of mineralisation was also significantly increased
following treatment with 0.5 and 5 ng/ml TGF-β1 (P<0.05); no
significant difference was reported between the 10 ng/ml
TGF-β1-treated and control groups.
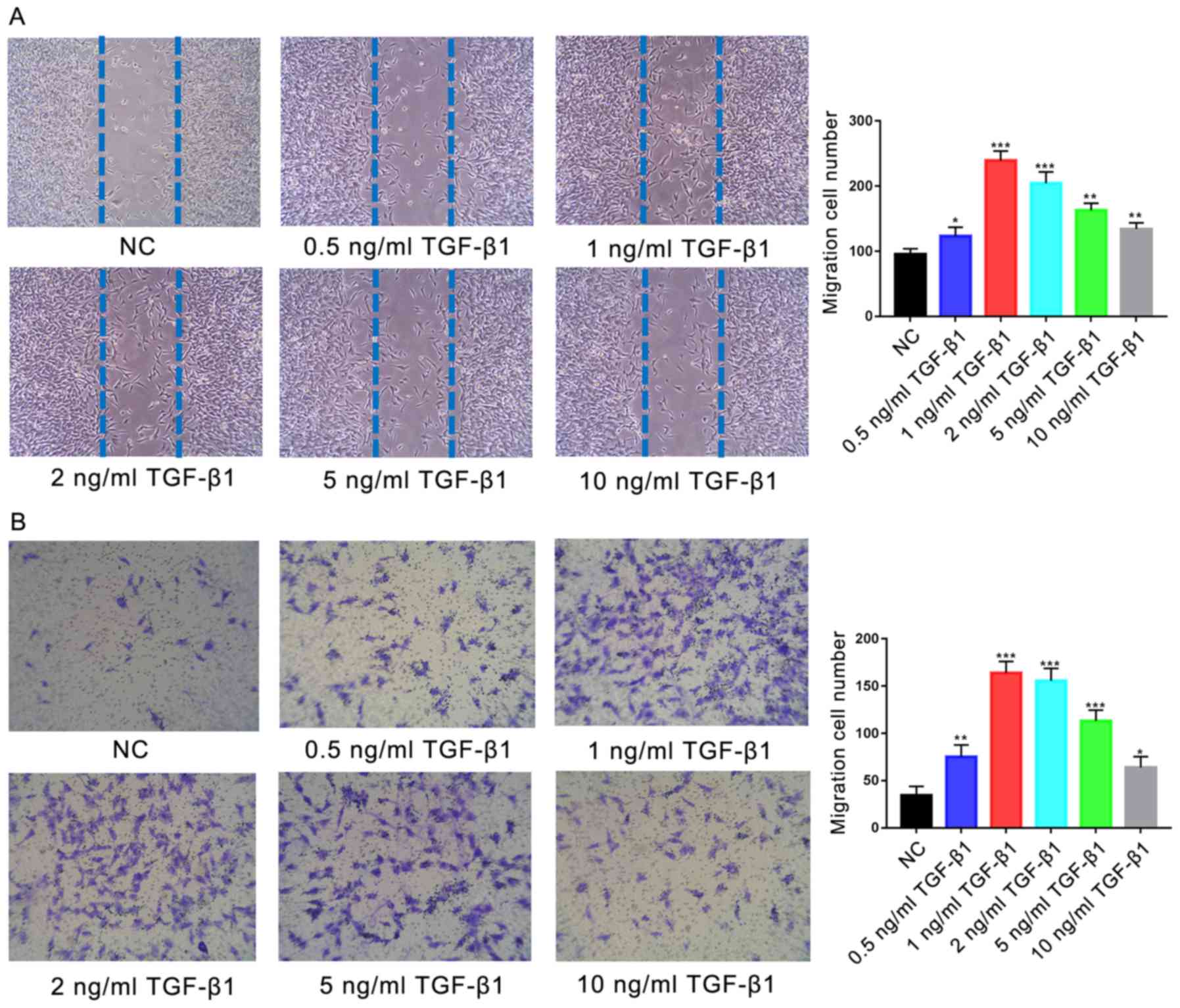
Treatment with TGF-β1 (0.5, 1, 2, 5 or 10 ng/ml)
resulted in significantly increased migration of cells in the
scratch-wound assay at all concentrations compared with the control
(P<0.05; Fig. 3A); the greatest
effects were observed in the 1 and 2 ng/ml TGF-β1-treatment groups
(P<0.001). Similarly, significantly increased migration was
observed during the Transwell assay following treatment with all
concentrations of TGF-β1 compared with the control (P<0.05;
Fig. 3B), with the most
significant increases observed in the 1 and 2 ng/ml
TGF-β1-treatment groups (P<0.001). No significant difference was
reported between the migration of cells treated with 1 or 2 ng/ml
TGF-β1 (P>0.05).
Effects of TGF-β1-mediated PI3K/AKT
signalling on ALP activity in osteoblasts
The effects of 1 ng/ml TGF-β1 treatment on the
activity of ALP in hFOB1.19 osteoblasts was investigated; ALP
activity generates high concentrations of phosphate at mineral
deposition sites during bone formation (5) and is a characteristic marker of
active osteoblasts. ALP-positive osteoblasts are stained purple by
BCIP/NBT (Fig. 4A). ALP staining
revealed that treatment with 1 ng/ml TGF-β1 for 7 days
significantly promoted ALP activity in human osteoblasts; however,
significant reductions in ALP activity compared with the control
were observed following treatment with the PI3K/AKT inhibitor
LY294002 (10 µM), in the presence or absence of TGF-β1
(P<0.001).
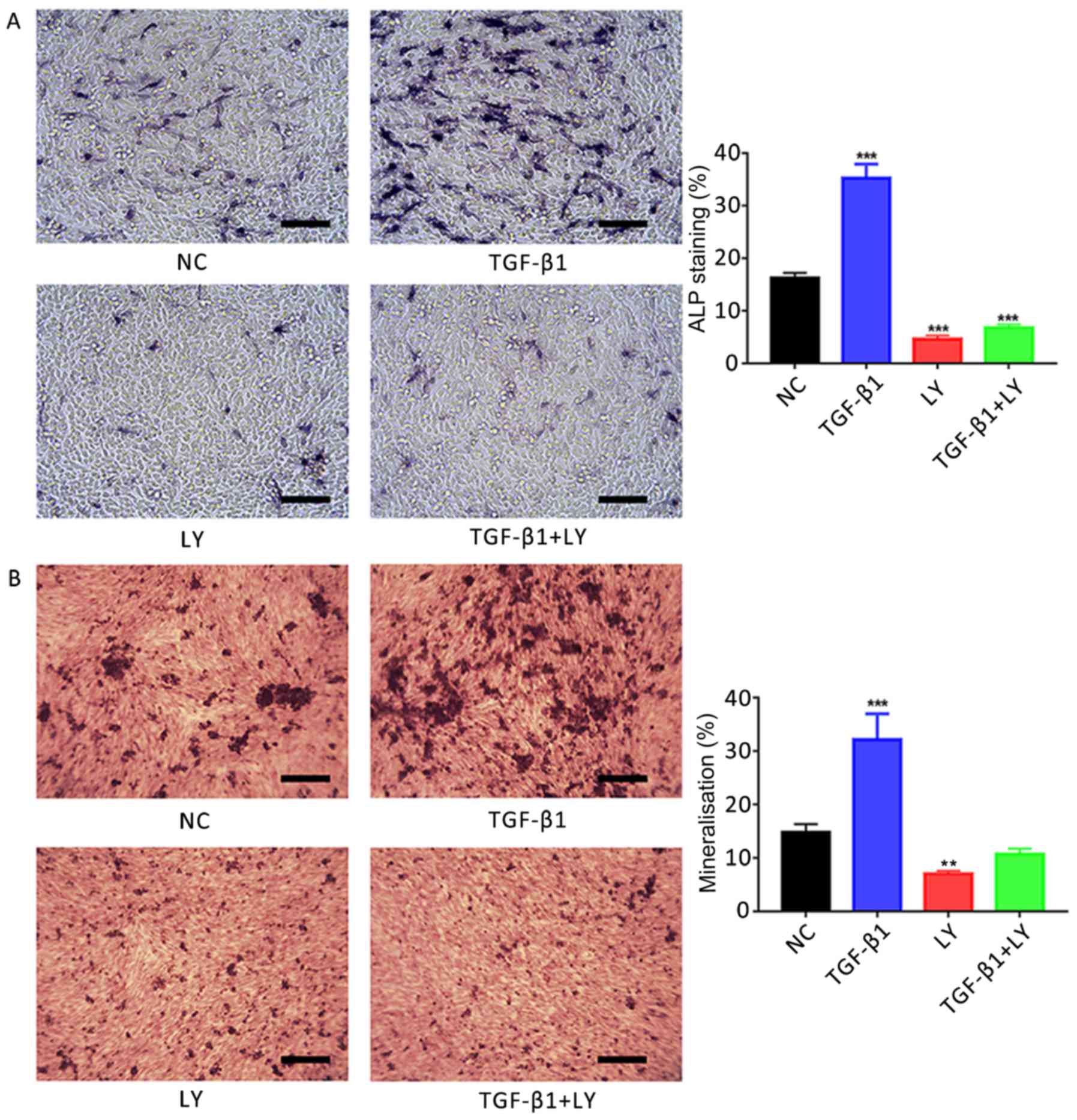 | Figure 4.Effects of TGF-β1 on osteogenic
induction are dependent upon PI3K/AKT signalling. (A) Effects of
treatment for 7 days with 1 ng/ml TGF-β1 and/or 10 µM LY, a
PI3K/AKT inhibitor, on the activity of ALP in hFOB1.19 human
osteoblasts. ALP-positive cells were stained with
5-bromo-4-chloro-3-indolyl-phosphate/nitro blue tetrazolium. (B)
Ca2+ deposits following treatment of osteoblasts for 14 days with 1
ng/ml TGF-β1 and/or 10 µM LY, identified by Alizarin red S
staining. Scale bar, 200 µm. Data are presented as the mean ±
standard deviation. **P<0.01 and ***P<0.001 vs. NC. AKT,
protein kinase B; ALP, alkaline phosphatase; LY, LY294002; NC,
negative control; PI3K, phosphatidylinositol 3-kinase; TGF-β1,
transforming growth factor β1. |
Effects of TGF-β1-mediated PI3K/AKT
signalling on osteoblast mineralisation
Mineralisation of the extracellular matrix serves an
important role in the formation of bone (5). In the present study, it was
demonstrated using Alizarin red S staining that the treatment of
hFOB.19 osteoblasts with 1 ng/ml TGF-β1 for 14 days significantly
promoted the mineralisation of extracellular matrices compared with
the control (Fig. 4B). Conversely,
significant reductions in matrix Ca2+ deposits were
reported following the inhibition of the PI3K/AKT signalling
pathway via treatment with 10 µM LY294002 compared with the NC
group (P<0.01).
Effects of TGF-β1-mediated PI3K/AKT
signalling on the motility of osteoblasts
The role of TGF-β1 in the migration of osteoblasts
was investigated using a scratch-wound assay. It was revealed that
the number of migrating cells following treatment with 1 ng/ml
TGF-β1 (364±11 cells) was significantly increased compared with the
control (176±7 cells; Fig. 5A).
Conversely, treatment with 10 µM LY294002 (106±8 cells) or LY294002
plus TGF-β1 (101±6 cells) significantly decreased the migration of
cells compared with the control (P<0.01).
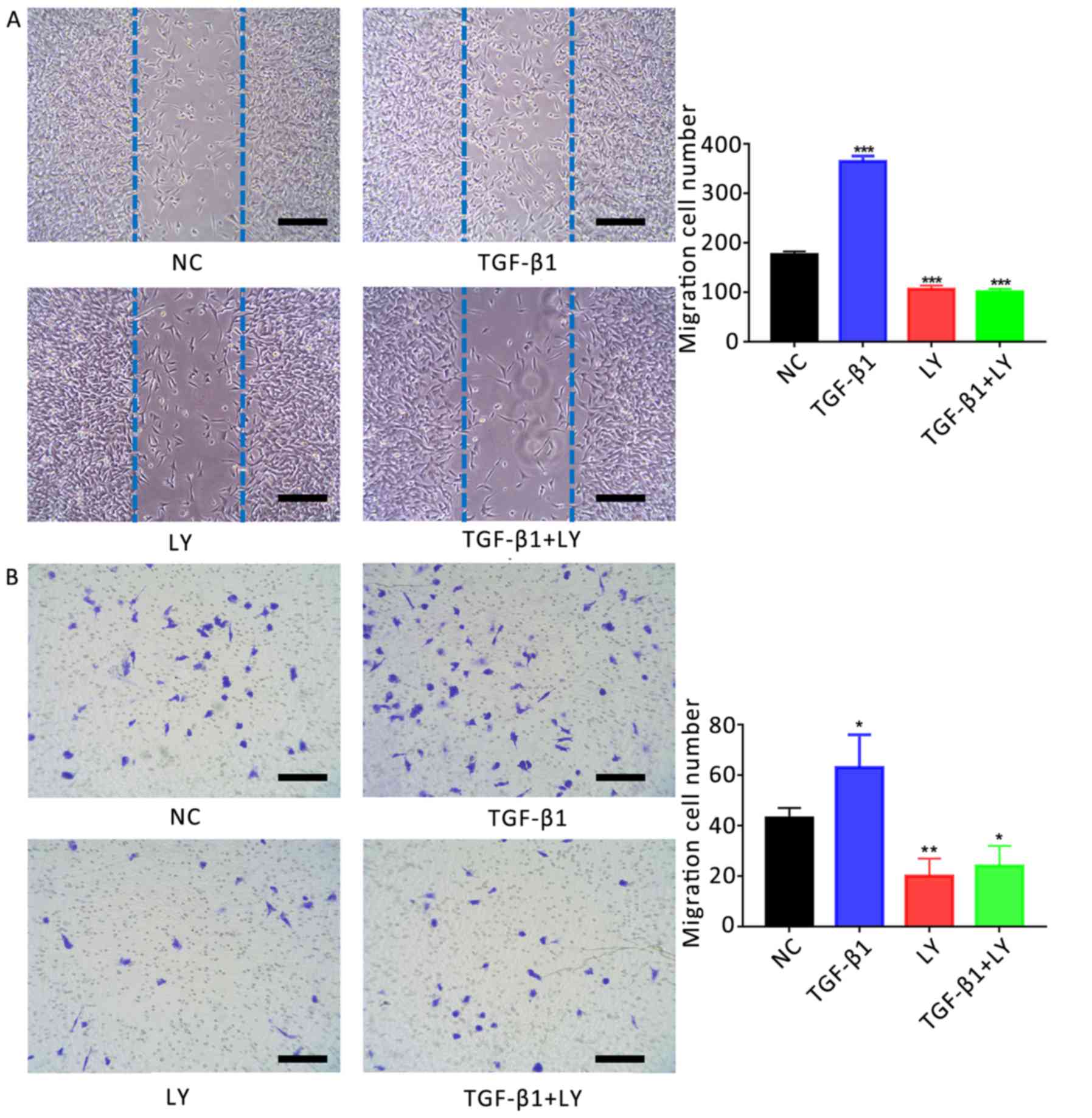 | Figure 5.Effects of TGF-β1 on the motility of
human osteoblasts are dependent upon PI3K/AKT signalling. (A)
Migration of hFOB1.19 human osteoblasts treated for 24 h with 1
ng/ml TGF-β1 and/or 10 µM LY, a PI3K/AKT inhibitor, as determined
by a scratch-wound assay. The number of cells entering the wound
indicated the migration of cells. (B) Migration of osteoblasts
treated for 24 h with 1 ng/ml TGF-β1 and/or 10 µM LY, as determined
by a Transwell assay. Scale bar, 200 µm. Data are presented as the
mean ± standard deviation. *P<0.05, **P<0.01 and
***P<0.001 vs. NC. AKT, protein kinase B; LY, LY294002; NC,
negative control; PI3K, phosphatidylinositol 3-kinase; TGF-β1,
transforming growth factor β1. |
The migration of osteoblasts was also investigated
using a Transwell assay. The number of migrating cells following
treatment with 1 ng/ml TGF-β1 (63±13 cells) was significantly
increased compared with the NC (43±4 cells; Fig. 5B). Conversely, treatment with 10 µM
LY294002 with (20±7 cells) or without TGF-β1 (24±8 cells)
significantly reduced the number of migrating cells compared with
the control (P<0.05).
PI3K/AKT signalling mediates the
TGF-β1-induced expression of genes associated with osteogenesis and
migration
The mechanisms underlying the effects of TGF-β1 and
PI3K/AKT signalling on the function of osteoblasts were
investigated by determining the mRNA expression of genes associated
with the osteogenesis, migration and chemotaxis of cells (Runx2,
Osterix, OPN, OCN, MMP-2 and MMP-9). Cells were separated into four
groups: The NC; treatment with 1 ng/ml TGF-β1; treatment with 10 µM
LY294002 combined with TGF-β1, and LY294002 treatment alone. It was
demonstrated that TGF-β1 treatment significantly upregulated the
expression of osteogenesis-associated (Runx2, Osterix, OPN and OCN)
and migration-associated genes (MMP-2 and MMP-9) compared with the
NC (Fig. 6). Conversely,
inhibition of the PI3K/AKT signalling pathway significantly
downregulated the expression of the aforementioned genes. The
results indicated that TGF-β1 promoted the migration and
mineralisation of osteoblasts in a PI3K/AKT-dependent manner.
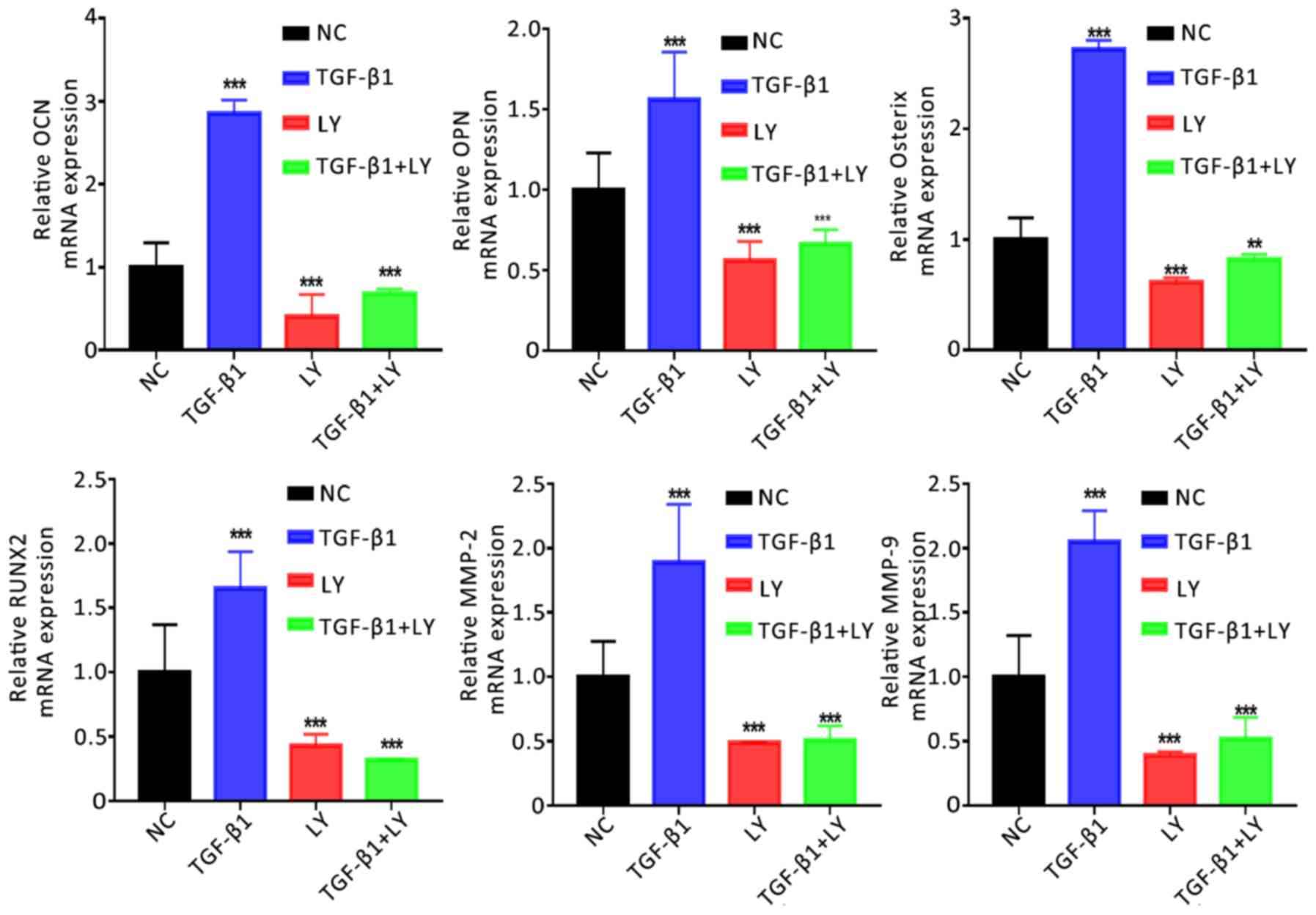 | Figure 6.Effects of TGF-β1 and inhibition of
the PI3K/AKT pathway on the expression of osteogenic and
migration-associated genes. Expression of osteogenic (OCN, OPN,
Osterix and RUNX2) and migration-associated (MMP2 and MMP9) genes
following treatment for 14 days with 1 ng/ml TGF-β1 and/or 10 µM
LY, a PI3K/AKT inhibitor, as determined by reverse
transcription-quantitative polymerase chain reaction. Data are
presented as the mean ± standard deviation. **P<0.01,
***P<0.001 vs. NC. AKT, protein kinase B; LY, LY294002; MMP,
matrix metalloproteinase; NC, negative control; OCN, osteocalcin;
OPN, osteopontin; Osterix, transcription factor Sp7; PI3K,
phosphatidylinositol 3-kinase; RUNX2, Runt-related transcription
factor 2; TGF-β1, transforming growth factor β1. |
Mechanisms underlying the
PI3K/AKT/mTOR/S6K1 signalling pathway-dependent effects of TGF-β1
on hFOB1.19 osteoblasts
The involvement of mTOR/S6K1 signalling downstream
of PI3K/AKT in the osteogenesis and migration of osteoblasts was
investigated. Cells were separated into three treatment groups: 1
ng/ml TGF-β1; 1 ng/ml TGF-β1 plus 10 µM LY294002, and 1 ng/ml
TGF-β1 plus 100 nM rapamycin (an inhibitor of mTOR/S6K1
signalling). As presented in Fig.
7, inhibition of the mTOR/S6K1 signalling pathway with
rapamycin significantly decreased the activity of ALP in
osteoblasts and Ca2+ deposition compared with the TGF-β1
group (P<0.001). Similar effects were observed on the motility
of cells following treatment with LY294002 or rapamycin compared
with TGF-β1 treatment alone (Fig.
8). Additionally, treatment with LY294002 or rapamycin
significantly downregulated the mRNA expression of migration- and
osteogenesis-associated genes compared with TGF-β1 treatment alone
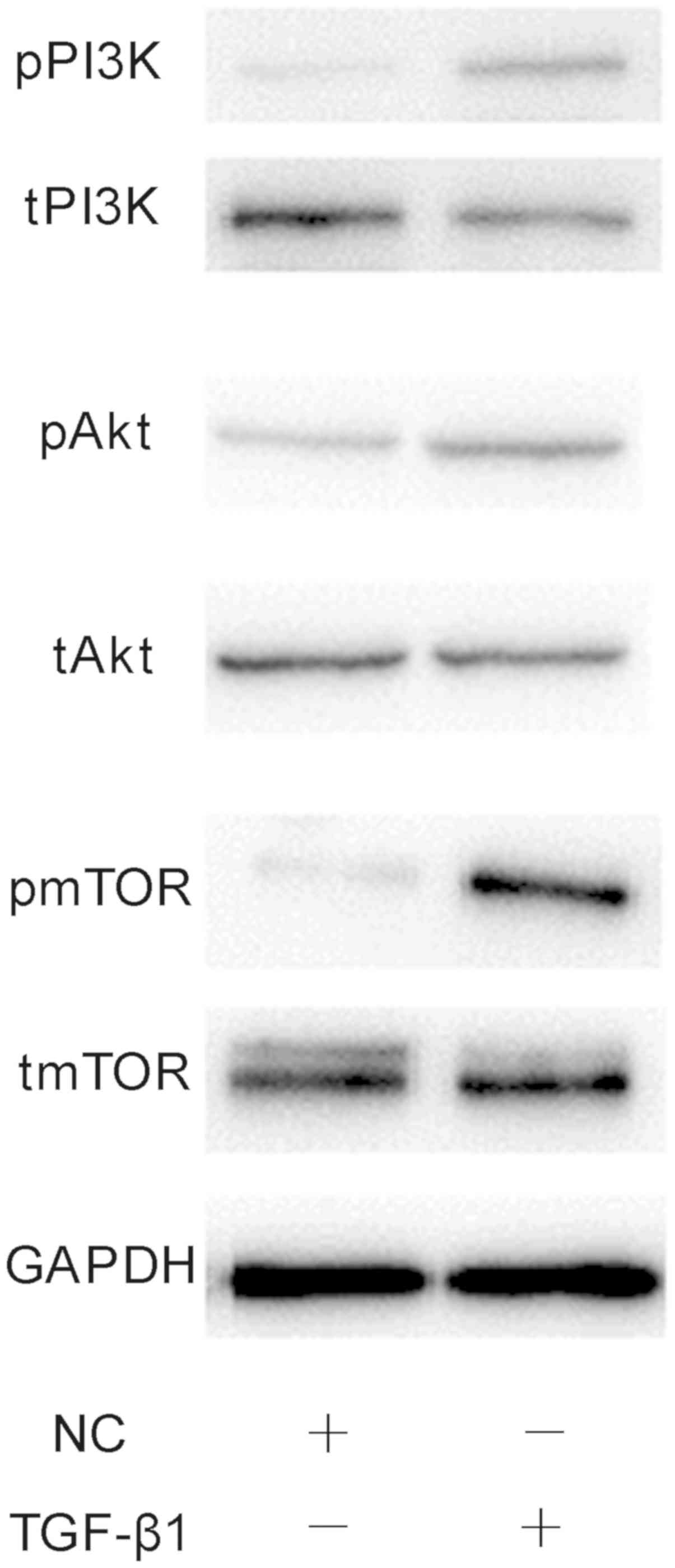
(Fig. 9). Finally, western
blotting was performed to investigate the effects of 1 ng/ml TGF-β1
on PI3K/AKT/mTOR signalling. It was demonstrated that TGF-β1
markedly increased the phosphorylation of PI3K, AKT and mTOR
compared with the NC group (Fig.
10). The results suggested that the effects of TGF-β1 on the
osteogenesis and mineralisation osteoblast were mediated via
mTOR/S6K1 signalling downstream of the PI3K/AKT signalling
pathway.
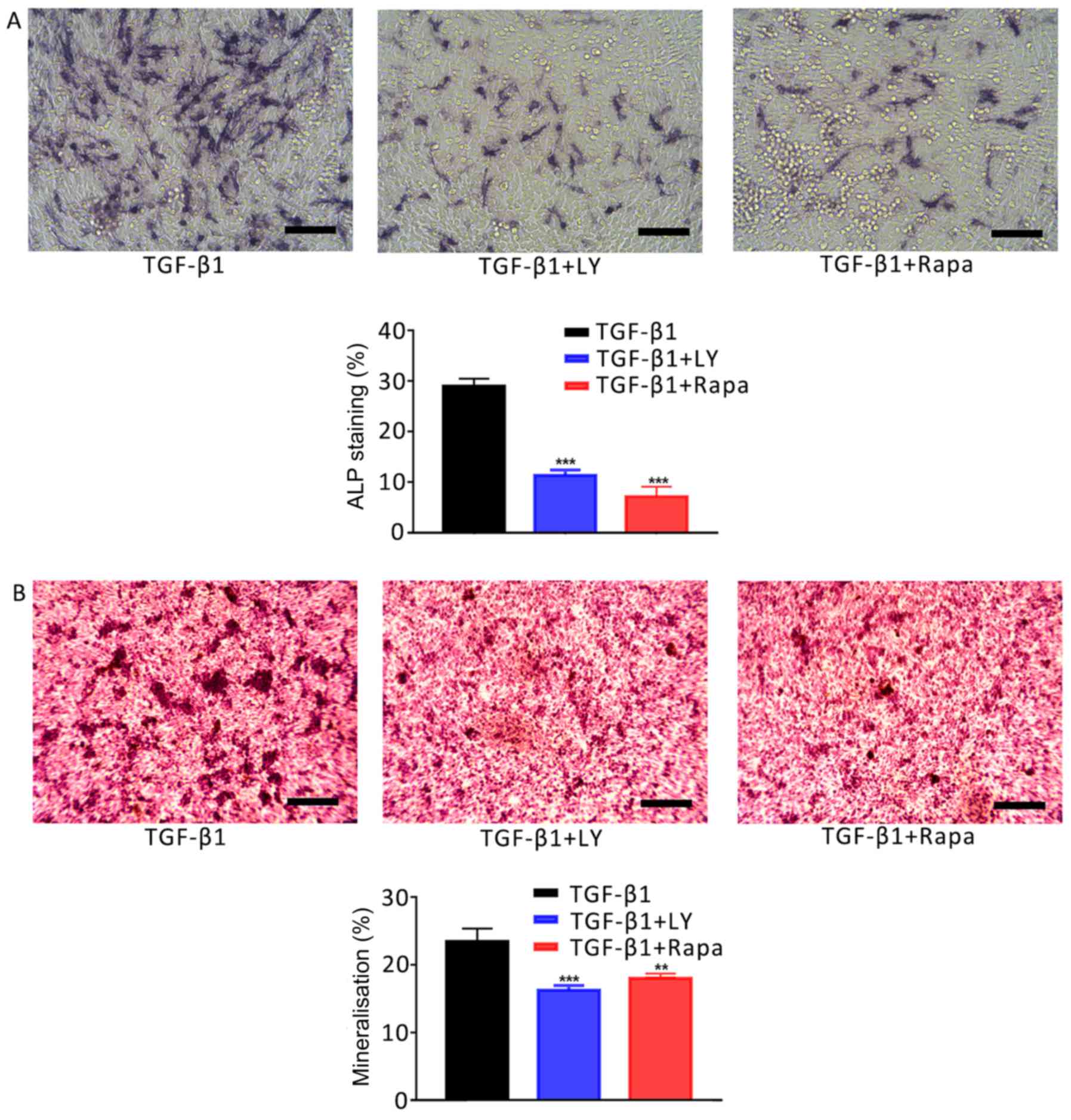 | Figure 7.Effects of the inhibition of PI3K/AKT
and mTOR/S6K1 signalling on TGF-β1-mediated osteogenic induction.
(A) Effects of treatment for 7 days with 1 ng/ml TGF-β1 plus 10 µM
LY, a PI3K/AKT inhibitor, or 100 nM Rapa, an mTOR/S6K1 inhibitor,
on the activity of ALP in hFOB1.19 human osteoblasts. ALP-positive
cells were stained with 5-bromo-4-chloro-3-indolyl-phosphate/nitro
blue tetrazolium. (B) Ca2+ deposits following treatment of
osteoblasts for 14 days with 1 ng/ml TGF-β1 plus 10 µM LY or 100 nM
Rapa, as identified by Alizarin red S staining. Scale bar, 200 µm.
Data are presented as the mean ± standard deviation. **P<0.01
and ***P<0.001 vs. TGF-β1. AKT, protein kinase B; ALP, alkaline
phosphatase; LY, LY294002; mTOR, mammalian target of rapamycin; NC,
negative control; PI3K, phosphatidylinositol 3-kinase; Rapa,
rapamycin; S6K1, S6 kinase 1; TGF-β1, transforming growth factor
β1. |
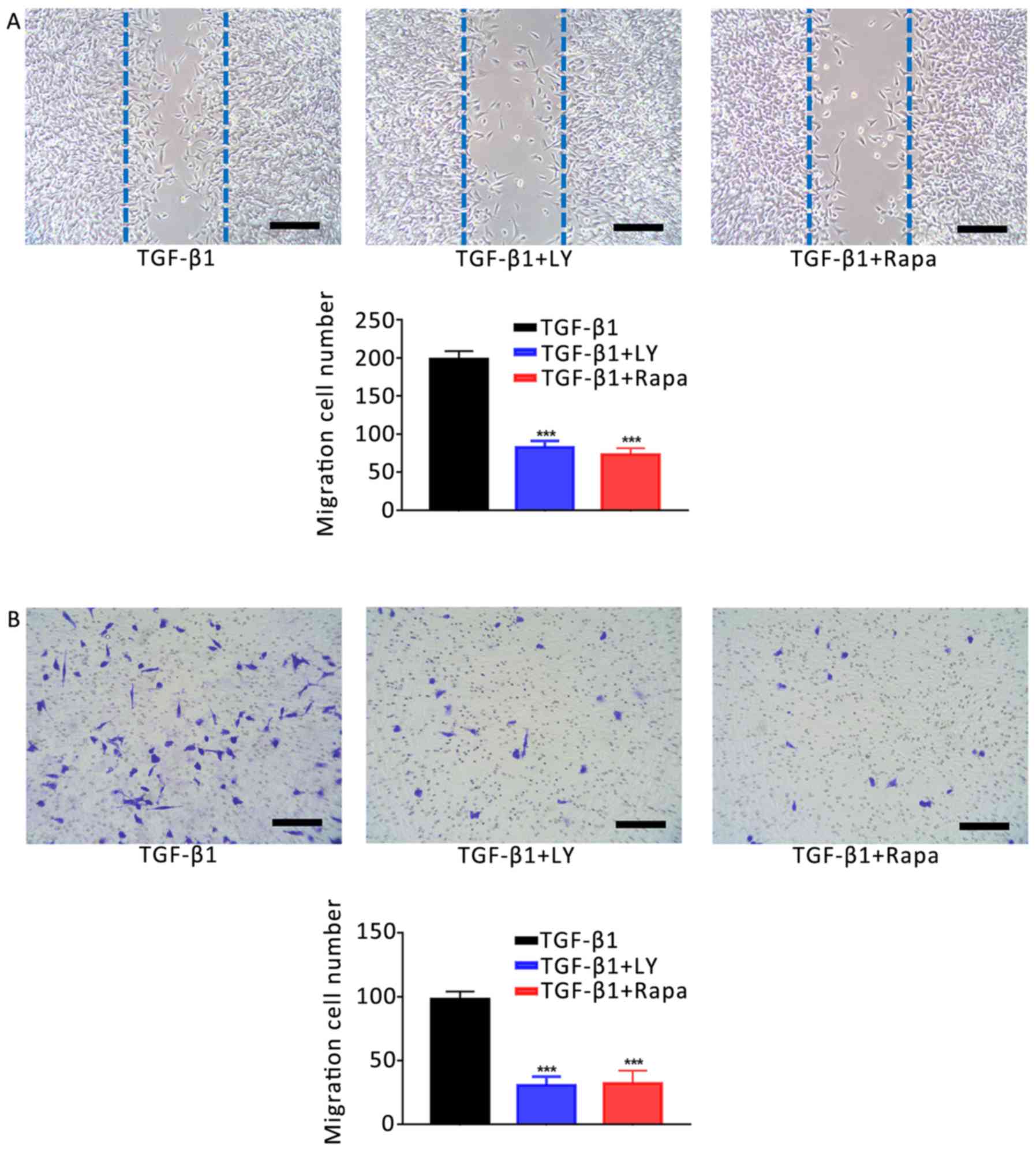 | Figure 8.Effects of the inhibition of PI3K/AKT
and mTOR/S6K1 signalling on the TGF-β1-induced increase in
osteoblast motility. (A) Migration of hFOB1.19 human osteoblasts
treated for 24 h with 1 ng/ml TGF-β1 plus 10 µM LY, a PI3K/AKT
inhibitor, or 100 nM Rapa, an mTOR/S6K1 inhibitor; as determined by
a scratch-wound assay. The number of cells entering the wound
indicated the migration of cells. (B) Migration of osteoblasts
treated for 24 h with 1 ng/ml TGF-β1 plus 10 µM LY or 100 nM Rapa,
as determined by a Transwell assay. Scale bar, 200 µm. Data are
presented as the mean ± standard deviation. ***P<0.001 vs.
TGF-β1. AKT, protein kinase B; LY, LY294002; mTOR, mammalian target
of rapamycin; NC, negative control; PI3K, phosphatidylinositol
3-kinase; Rapa, rapamycin; S6K1, S6 kinase 1; TGF-β1, transforming
growth factor β1. |
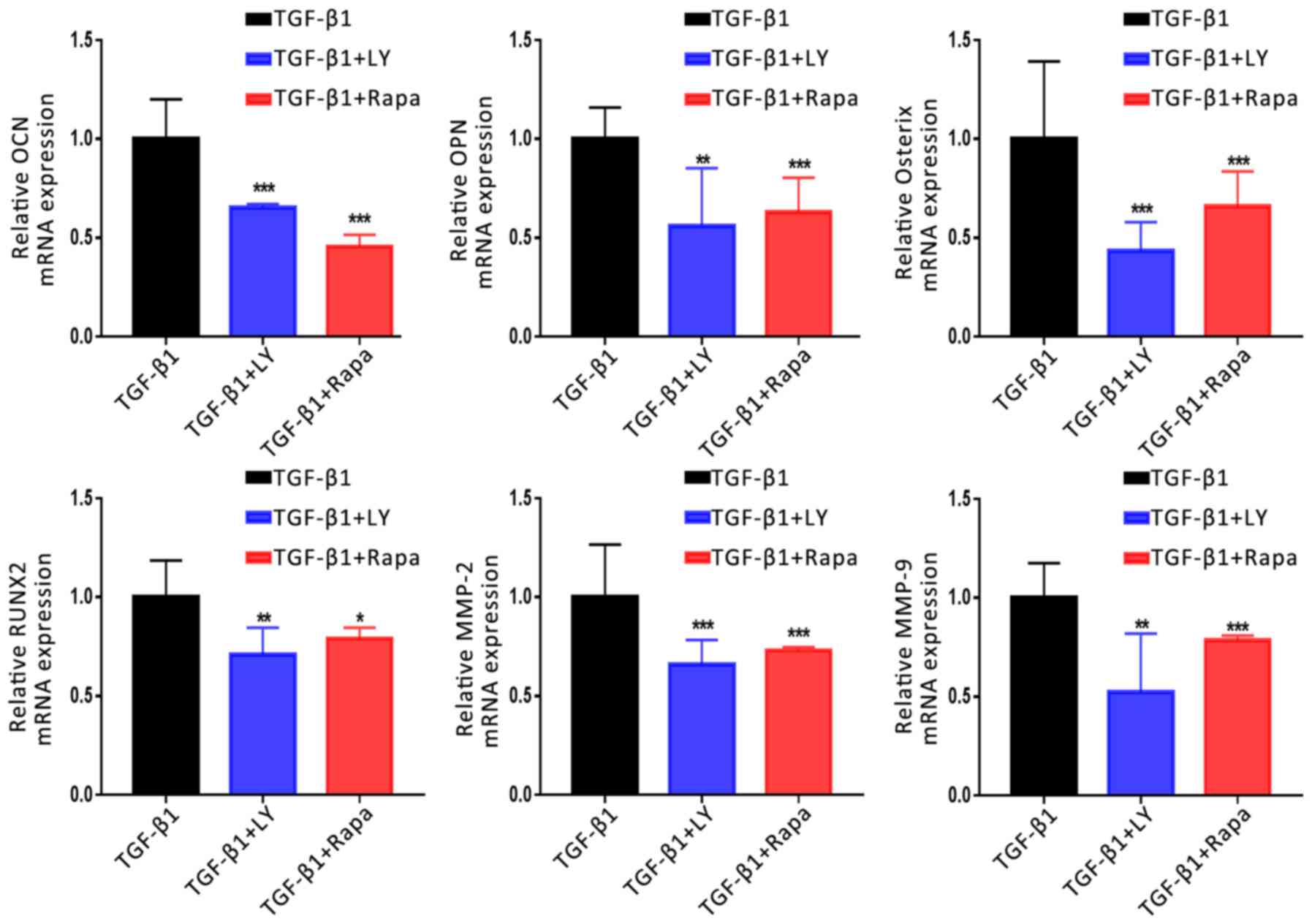 | Figure 9.Effects of the inhibition of PI3K/AKT
and mTOR/S6K1 signalling on the TGF-β1-induced upregulation of
osteogenic and migration-associated genes. Expression of osteogenic
(OCN, OPN, Osterix and RUNX2) and migration-associated (MMP2 and
MMP9) genes following treatment for 14 days with 1 ng/ml TGF-β1
plus 10 µM LY, a PI3K/AKT inhibitor, or 100 nM Rapa, an mTOR/S6K1
inhibitor, as determined by reverse transcription-quantitative
polymerase chain reaction. Data are presented as the mean ±
standard deviation. *P<0.05, **P<0.01, ***P<0.001 vs.
TGF-β1. AKT, protein kinase B; LY, LY294002; MMP, matrix
metalloproteinase; mTOR, mammalian target of rapamycin; NC,
negative control; OCN, osteocalcin; OPN, osteopontin; Osterix,
transcription factor Sp7; PI3K, phosphatidylinositol 3-kinase;
Rapa, rapamycin; RUNX2, Runt-related transcription factor 2; S6K1,
S6 kinase 1; TGF-β1, transforming growth factor β1. |
Discussion
Bone tissue engineering is frequently used in the
repair and regeneration process of bone injury (32). Further investigation is required to
identify novel cytokines, and develop materials and seed cells for
improved engineering. Cytokines serve important roles in immune
responses, and the proliferation, chemotaxis and differentiation of
cells (33,34). TGF-β1 was hypothesized to be a
potential candidate for the treatment of bone injury; previous
studies reported that TGF-β1 was a key regulator of bone
remodelling and regeneration (35), and stimulated the proliferation of
osteoblasts and regulated the function of osteoclasts (36).
Unlike pathways including the MAPK, Wnt, Notch and
PI3K signalling pathways, TGF-β1 regulates the biological activity
of osteoblasts via a variety of mechanisms; this may explain the
complexity of its effects on cells (23,37).
The cellular concentration of TGF-β1 affects the activity of cells;
for example, low concentrations promote cell proliferation, whereas
opposing effects are observed with high concentrations (38). In the present study, hFOB1.19
osteoblasts were treated with various concentrations of TGF-β1, and
the effects on proliferation, osteoinduction and migration were
investigated. CCK-8 and flow cytometry assays revealed that the
proliferation of osteoblasts was most notably increased following
treatment with 1 and 2 ng/ml TGF-β1, compared with 0.5, 5 and 10
ng/ml; however, no significant difference was reported between 1
and 2 ng/ml. Therefore, the proliferation of osteoblasts was
affected by the concentration of TGF-β1 in a non-linear manner.
Additionally, the effects of this concentration gradient on
osteogenic induction and migration were determined. ALP and
Alizarin red S staining revealed that 1 and 2 ng/ml TGF-β1 markedly
increased the activity of ALP in osteoblasts and levels of
Ca2+ deposition, compared with 0.5 and >5 mg/ml
TGF-β1. Furthermore, scratch-wound and Transwell assays revealed
that TGF-β1 most notably increased the motility of osteoblasts at 1
and 2 ng/ml. These findings indicated that a basal concentration of
1 ng/ml may be optimal for inducing the biological effects of
TGF-β1 on human osteoblasts.
The mineralisation of osteoblasts is important for
regeneration during bone repair; improvements in osteogenesis and
mineralisation promote bone repair (5). It has been reported that TGF-β1
induces osteoblast differentiation (39,40).
Cell differentiation mediated by TGF-β1 may be regulated by
Smad-dependent and -independent pathways (41); however, the effects and involvement
of Smad-independent signalling pathways in the TGF-β1-mediated
differentiation of osteoblasts require further investigation. PI3K
is an important signalling molecule for numerous cellular
activities (41,42). PI3K catalyses the phosphorylation
of the D3 hydroxyl of phosphatidylinositol to produce
phosphatidylinositol-3,4,5-trisphosphate, which binds to
phosphoinositide-dependent kinase and the intracellular pleckstrin
homology domain of AKT (42).
Activation of AKT regulates the proliferation, differentiation,
apoptosis and migration of cells (43). TGF-β1 induces various biological
effects on cells via PI3K/AKT signalling (44). The activity of ALP and the
synthesis of extracellular matrix calcium ion deposits are
important indicators of osteoblast ossification and mineralisation.
In the present study, the effects of TGF-β1 on the ossification and
mineralisation of osteoblasts were evaluated by Alizarin red and
ALP staining. It was demonstrated that the expression of ALP and
the number of mineralised nodules were significantly increased
following treatment with 1 ng/ml TGF-β1; however, treatment with
the PI3K/AKT blocker LY294002 induced opposing effects, regardless
of TGF-β1 treatment. The results indicated that the effects of
TGF-β1 on ALP activity and the mineralisation of osteoblasts may
involve the PI3K/AKT signalling pathway.
The migration of osteoblasts to a damaged area
serves an important role in the repair of bone injury, as it
promotes bone regeneration (45,46).
A previous study reported that TGF-β1 upregulated the expression of
chemokine C-X-C motif ligand 16 in osteoblasts to stimulate the
migration of cells (47). In the
present study, the effects of TGF-β1 on the migration of
osteoblasts were evaluated. It was demonstrated that cells treated
with 1 ng/ml TGF-β1 exhibited significantly increased motility
compared with the NC. Conversely, inhibition of the PI3K/AKT
signalling pathway reduced the migration of osteoblasts, indicating
that the effects of TGF-β1 on the motility of osteoblasts involved
PI3K/AKT.
To further investigate the effects of TGF-β1 on the
ossification and migration of osteoblasts, the mRNA expression of
associated genes was investigated, including Runx2, OPN, OCN,
Osterix, MMP-2 and MMP-9. Runx2 is a transcription factor specific
to osteoblast differentiation, and is the earliest and most
specific marker gene during bone formation (48,49).
It has been reported that the Runx2 gene activates the
transcription and expression of OCN, OPN, bone sialoprotein and
type I collagen by binding to the cis-acting elements of
osteoblasts (50). OPN serves an
important role in the mineralisation and absorption of bone matrix
(51). OCN is an important
component of the extracellular matrix and is regarded as a specific
marker of the bone turnover process (52); when OCN is in an inactive state, it
directly affects the formation, absorption and mineralisation of
bone (53). The zinc finger
protein Osterix is considered to be an important regulator of
osteoblast differentiation and maturation (54). The MMP family serves roles in the
invasion and metastasis of cells; MMP-2 and MMP-9 are involved in
bone remodelling and regeneration (55). In the present study, the expression
of osteogenic (Runx2, OPN, OCN and Osterix) and
migration-associated genes (MMP2 and MMP9) was significantly
upregulated following treatment with 1 ng/ml TGF-β1. In addition,
by blocking the PI3K/AKT signalling pathway, the expression of
these genes was downregulated. These findings indicated that TGF-β1
promoted the osteogenesis and migration of osteoblasts via the
upregulation of associated genes in a PI3K/AKT-dependent manner;
however, in addition to TGF-β1, there may be other signals
regulating the PI3K/AKT pathway in osteoblasts, as treatment with
LY294002 significantly downregulated gene expression compared with
the NC.
mTOR is a target of rapamycin and is downstream of
the PI3K/AKT signalling pathway. Activated AKT directly
phosphorylates the Ser2448 site of mTOR (56); S6K1 is one of the main targets
downstream of mTOR that is induced following the activation of mTOR
(57). mTOR/S6K1 signalling serves
important roles in the biological processes of various cells,
including DNA replication, mRNA translation, and the synthesis of
proteins and lipids to regulate the growth, survival, migration,
and differentiation of cells (58,59).
Luo et al (30) reported
that blocking the AKT signalling pathway decreased mTOR/S6K1
signalling, and reduced the ability of osteocytes to differentiate
and proliferate. In the present study, treatment with TGF-β1 was
combined with an mTOR/S6K1 inhibitor, rapamycin, to investigate the
involvement of mTOR/S6K1 in the effects of TGF-β1. Similar to
PI3K/AKT blockade, treatment with rapamycin significantly decreased
ALP activity, the number of mineralised nodules and the motility of
osteoblasts compared with TGF-β1 treatment alone. Furthermore,
analysis of mRNA expression in osteoblasts revealed that rapamycin
suppressed the TGF-β1-induced upregulation of osteogenic and
migration-associated genes. The results indicated that the
PI3K/AKT-mediated effects of TGF-β1 on osteoblast osteogenesis and
mineralisation also involved the mTOR/S6K1 pathway downstream of
PI3K/AKT (Fig. 11).
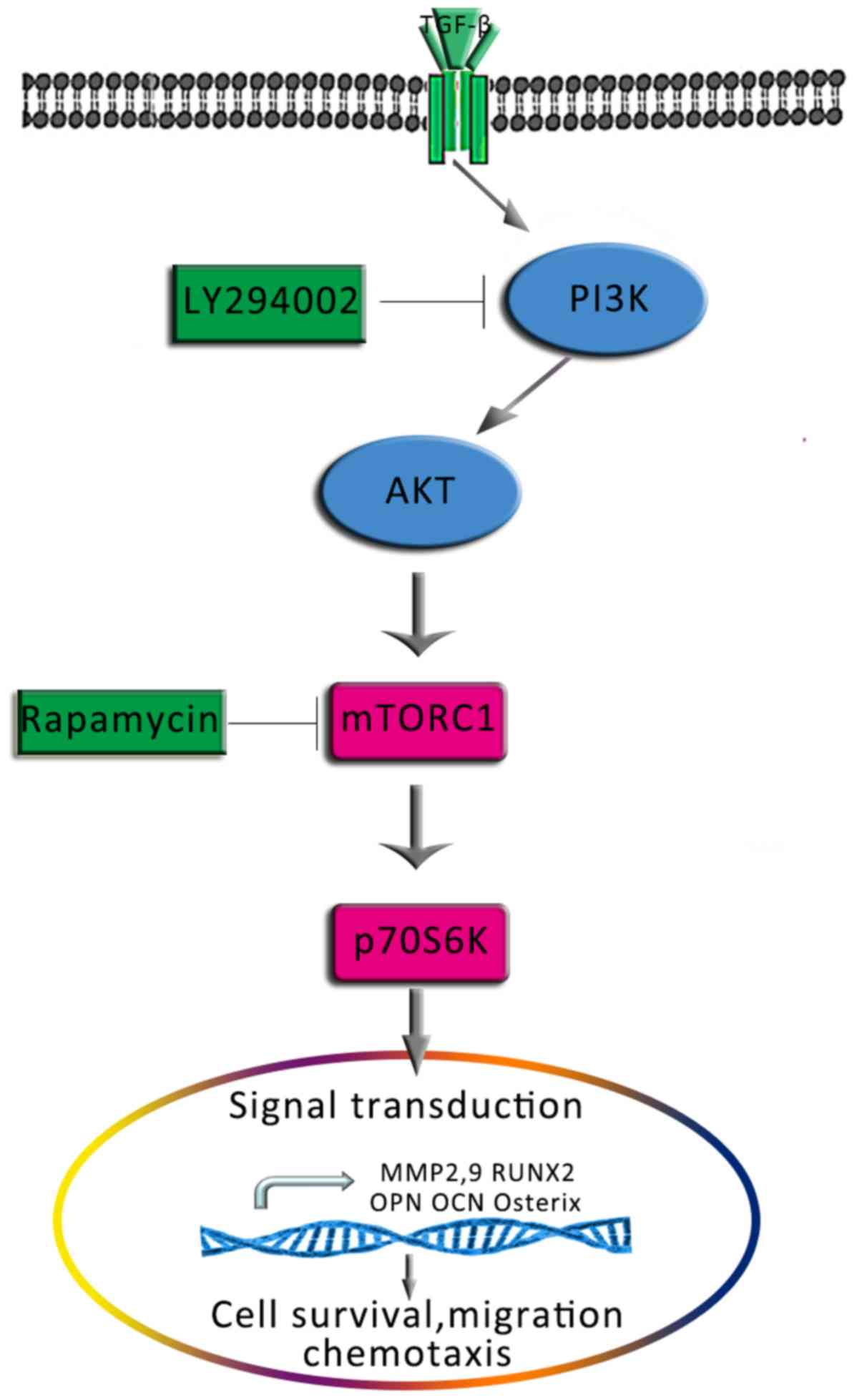 | Figure 11.Proposed schematic depicting the
signalling pathway via which TGF-β1 promotes the osteogenesis and
migration of osteoblasts. AKT, protein kinase B; MMP, matrix
metalloproteinase; mTOR, mammalian target of rapamycin; OCN,
osteocalcin; OPN, osteopontin; Osterix, transcription factor Sp7;
p70S6K, p70-S6 kinase 1; PI3K, phosphatidylinositol 3-kinase;
Runx2, Runt-related transcription factor 2; TGF-β1, transforming
growth factor β1. |
In conclusion, it was demonstrated that TGF-β1
promoted the survival, osteogenic differentiation and migration of
human hFOB1.19 osteoblasts. Additionally, it was revealed that
TGF-β1 induced these effects via the PI3K/AKT/mTOR/S6K1 signalling
pathway. These findings indicated a potential role for TGF-β1 in
the field of bone tissue engineering and provided novel insight
into the mechanisms for application in bone injury repair in the
future.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
class general financial grant from the China Postdoctoral Science
Foundation (grant nos. 2017M620099 and 171478), the National
Natural Scientific Foundation of China (grant no. 81672139), the
Science and Technology Research Foundation of Liaoning Provincial
Department of Education (grant no. L2015034), the Scientific
Research Foundation for the PhD, Liaoning Provincial Department of
Science and Technology (grant no. 201601305), and the Scientific
Research Foundation for Overseas Students, Ministry of Human
Resources and Social Security of the People's Republic of China
(grant no. 2016-176).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
XZ and ZZ contributed to the conception and design
of the study, acquired and analyzed the data and drafted the
manuscript. DZ contributed to the conception and design of the
study, and revised the article critically for important
intellectual content. BL, BW, WY, JL, XY, FC, GZ, YZ and YL
contributed to the design of the study, revised the article and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patients consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wei X, Zhao D, Wang B, Wang W, Kang K, Xie
H, Liu B, Zhang X, Zhang J and Yang Z: Tantalum coating of porous
carbon scaffold supplemented with autologous bone marrow stromal
stem cells for bone regeneration in vitro and in vivo. Exp Biol Med
(Maywood). 241:592–602. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Su N, Gao PL, Wang K, Wang JY, Zhong Y and
Luo Y: Fibrous scaffolds potentiate the paracrine function of
mesenchymal stem cells: A new dimension in cell-material
interaction. Biomaterials. 141:74–85. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xia Y, Sun J, Zhao L, Zhang F, Liang XJ,
Guo Y, Weir MD, Reynolds MA, Gu N and Xu HHK: Magnetic field and
nano-scaffolds with stem cells to enhance bone regeneration.
Biomaterials. 183:151–170. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hallam P, Haddad F and Cobb J: Pain in the
well-fixed, aseptic titanium hip replacement. The role of
corrosion. J Bone Joint Surg Br. 86:27–30. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang X, Zu H, Zhao D, Yang K, Tian S, Yu
X, Lu F, Liu B, Yu X, Wang B, et al: Ion channel functional protein
kinase TRPM7 regulates Mg ions to promote the osteoinduction of
human osteoblast via PI3K pathway: In vitro simulation of the
bone-repairing effect of Mg-based alloy implant. Acta Biomater.
63:369–382. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin D, Zuo S, Li L, Wang L and Lian K:
Treatment of neglected femoral neck fractures using the modified
dynamic hip screw with autogenous bone and bone morphogenetic
protein-2 composite materials grafting. Indian J Orthop.
49:342–346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moore NM, Lin NJ, Gallant ND and Becker
ML: Synergistic enhancement of human bone marrow stromal cell
proliferation and osteogenic differentiation on BMP-2-derived and
RGD peptide concentration gradients. Acta Biomater. 7:2091–2100.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Piek E, Sleumer LS, van Someren EP, Heuver
L, de Haan JR, de Grijs I, Gilissen C, Hendriks JM, van
Ravestein-van Os RI, Bauerschmidt S, et al: Osteo-transcriptomics
of human mesenchymal stem cells: Accelerated gene expression and
osteoblast differentiation induced by vitamin D reveals c-MYC as an
enhancer of BMP2-induced osteogenesis. Bone. 46:613–627. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Poh CK, Shi Z, Lim TY, Neoh KG and Wang W:
The effect of VEGF functionalization of titanium on endothelial
cells in vitro. Biomaterials. 31:1578–1585. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Geiger F, Beverungen M, Lorenz H, Wieland
J, Fehr M and Kasten P: Bone substitute effect on vascularization
and bone remodeling after application of phVEGF165 transfected
BMSC. J Funct Biomater. 3:313–326. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu H, Zeng X, Deng C, Shi C, Ai J and Leng
W: Exogenous VEGF introduced by bioceramic composite materials
promotes the restoration of bone defect in rabbits. Biomed
Pharmacother. 98:325–332. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ueland T, Lekva T, Otterdal K, Dahl TB,
Olarescu NC, Jørgensen AP, Fougner KJ, Brixen K, Aukrust P and
Bollerslev J: Increased serum and bone matrix levels of
transforming growth factor {beta}1 in patients with GH deficiency
in response to GH treatment. Eur J Endocrinol. 165:393–400. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bonewald LF and Dallas SL: Role of active
and latent transforming growth factor beta in bone formation. J
Cell Biochem. 55:350–357. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao Y, Li Y, Gao Y, Yuan M, Manthari RK
and Wang J and Wang J: TGF-β1 acts as mediator in fluoride-induced
autophagy in the mouse osteoblast cells. Food Chem Toxicol.
115:26–33. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Siegel PM and Massagué J: Cytostatic and
apoptotic actions of TGF-beta in homeostasis and cancer. Nat Rev
Cancer. 3:807–821. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lind M: Growth factor stimulation of bone
healing. Effects on osteoblasts, osteomies, and implants fixation.
Acta Orthop Scand Suppl. 283:2–37. 1998.PubMed/NCBI
|
|
17
|
Duan X, Liu J, Zheng X, Wang Z, Zhang Y,
Hao Y, Yang T and Deng H: Deficiency of ATP6V1H causes bone loss by
inhibiting bone resorption and bone formation through the TGF-β1
pathway. Theranostics. 6:2183–2195. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marcelli C, Yates AJ and Mundy GR: In vivo
effects of human recombinant transforming growth factor beta on
bone turnover in normal mice. J Bone Miner Res. 5:1087–1096. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen LJ, Chen C, Qiao XY, Yu K, Xie LZ,
Cao J, Liu BL and Yan Y: Effect of porous titanium coated with
IGF-1 and TGF-β1 loaded gelatin microsphere on function
of MG63 cells. Transact Nonferr Metals Soc China. 25:2974–2985.
2015. View Article : Google Scholar
|
|
20
|
Lamberg A, Schmidmaier G, Søballe K and
Elmengaard B: Locally delivered TGF-beta1 and IGF-1 enhance the
fixation of titanium implants: A study in dogs. Acta Orthop.
77:799–805. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng XH and Derynck R: Specificity and
versatility in tgf-beta signaling through Smads. Annu Rev Cell Dev
Biol. 21:659–693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Karst M, Gorny G, Galvin RJ and Oursler
MJ: Roles of stromal cell RANKL, OPG, and M-CSF expression in
biphasic TGF-beta regulation of osteoclast differentiation. J Cell
Physiol. 200:99–106. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen G, Deng C and Li YP: TGF-β and BMP
signaling in osteoblast differentiation and bone formation. Int J
Biol Sci. 8:272–288. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kang JS, Alliston T, Delston R and Derynck
R: Repression of Runx2 function by TGF-beta through recruitment of
class II histone deacetylases by Smad3. EMBO J. 24:2543–2555. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xie F, Ling L, van Dam H, Zhou F and Zhang
L: TGF-β signaling in cancer metastasis. Acta Biochim Biophys Sin
(Shanghai). 50:121–132. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang J, Ma XY, Feng YF, Ma ZS, Ma TC,
Zhang Y, Li X, Wang L and Lei W: Magnesium ions promote the
biological behaviour of rat calvarial osteoblasts by activating the
PI3K/Akt signalling pathway. Biol Trace Elem Res. 179:284–293.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bertacchini J, Heidari N, Mediani L,
Capitani S, Shahjahani M, Ahmadzadeh A and Saki N: Targeting
PI3K/AKT/mTOR network for treatment of leukemia. Cell Mol Life Sci.
72:2337–2347. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Manfredi GI, Dicitore A, Gaudenzi G,
Caraglia M, Persani L and Vitale G: PI3K/Akt/mTOR signaling in
medullary thyroid cancer: A promising molecular target for cancer
therapy. Endocrine. 48:363–370. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Golub EE, Harrison G, Taylor AG, Camper S
and Shapiro IM: The role of alkaline phosphatase in cartilage
mineralization. Bone Miner. 17:273–278. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Luo G, Xu B and Huang Y: Icariside II
promotes the osteogenic differentiation of canine bone marrow
mesenchymal stem cells via the PI3K/AKT/mTOR/S6K1 signaling
pathways. Am J Transl Res. 9:2077–2087. 2017.PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen Q, Zhu C and Thouas GA: Progress and
challenges in biomaterials used for bone tissue engineering:
Bioactive glasses and elastomeric composites. Prog Biomater.
1:22012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen X, Lu J, Ji Y, Hong A and Xie Q:
Cytokines in osteoblast-conditioned medium promote the migration of
breast cancer cells. Tumour Biol. 35:791–798. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hughes FJ, Turner W, Belibasakis G and
Martuscelli G: Effects of growth factors and cytokines on
osteoblast differentiation. Periodontol 2000. 41:48–72. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Janssens K, ten Dijke P, Janssens S and
Van Hul W: Transforming growth factor-beta1 to the bone. Endocr
Rev. 26:743–774. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ota K, Quint P, Ruan M, Pederson L,
Westendorf JJ, Khosla S and Oursler MJ: TGF-β induces Wnt10b in
osteoclasts from female mice to enhance coupling to osteoblasts.
Endocrinology. 154:3745–3752. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kanaan RA and Kanaan LA: Transforming
growth factor beta1, bone connection. Med Sci Monit.
12:RA164–RA169. 2006.PubMed/NCBI
|
|
38
|
Wang X, Dong F, Zhang S, Yang W, Yu W,
Wang Z, Zhang S, Wang J, Ma S, Wu P, et al: TGF-beta1 negatively
regulates the number and function of hematopoietic stem cells. Stem
Cell Reports. 11:274–287. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Matsunobu T, Torigoe K, Ishikawa M, de
Vega S, Kulkarni AB, Iwamoto Y and Yamada Y: Critical roles of the
TGF-beta type I receptor ALK5 in perichondrial formation and
function, cartilage integrity, and osteoblast differentiation
during growth plate development. Dev Biol. 332:325–338. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ramirez-Yañez GO, Hamlet S, Jonarta A,
Seymour GJ and Symons AL: Prostaglandin E2 enhances transforming
growth factor-beta 1 and TGF-beta receptors synthesis: An in vivo
and in vitro study. Prostaglandins Leukot Essent Fatty Acids.
74:183–192. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cantrell DA: Phosphoinositide 3-kinase
signalling pathways. J Cell Sci. 114:1439–1445. 2001.PubMed/NCBI
|
|
42
|
Cantley LC: The phosphoinositide 3-kinase
pathway. Science. 296:1655–1657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Guntur AR and Rosen CJ: The skeleton: A
multi-functional complex organ: New insights into osteoblasts and
their role in bone formation: The central role of PI3Kinase. J
Endocrinol. 211:123–130. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mukherjee A and Rotwein P: Akt promotes
BMP2-mediated osteoblast differentiation and bone development. J
Cell Sci. 122:716–726. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kristensen HB, Andersen TL, Marcussen N,
Rolighed L and Delaisse JM: Osteoblast recruitment routes in human
cancellous bone remodeling. Am J Pathol. 184:778–789. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Maes C, Kobayashi T, Selig MK, Torrekens
S, Roth SI, Mackem S, Carmeliet G and Kronenberg HM: Osteoblast
precursors, but not mature osteoblasts, move into developing and
fractured bones along with invading blood vessels. Dev Cell.
19:329–344. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ota K, Quint P, Weivoda MM, Ruan M,
Pederson L, Westendorf JJ, Khosla S and Oursler MJ: Transforming
growth factor beta 1 induces CXCL16 and leukemia inhibitory factor
expression in osteoclasts to modulate migration of osteoblast
progenitors. Bone. 57:68–75. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fie C, Guo J, Zhao Y, Gu S, Zhao S, Li X
and Chang C: Notch-Hes pathway mediates the impaired osteogenic
differentiation of bone marrow mesenchymal stromal cells from
myelodysplastic syndromes patients through the down-regulation of
Runx2. Am J Transl Res. 7:1939–1951. 2015.PubMed/NCBI
|
|
49
|
Kumar Y, Kapoor I, Khan K, Thacker G, Khan
MP, Shukla N, Kanaujiya JK, Sanyal S, Chattopadhyay N and Trivedi
AK: E3 ubiquitin ligase Fbw7 negatively regulates osteoblast
differentiation by targeting Runx2 for degradation. J Biol Chem.
290:30975–30987. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gersbach CA, Byers BA, Pavlath GK and
García AJ: Runx2/Cbfa1 stimulates transdifferentiation of primary
skeletal myoblasts into a mineralizing osteoblastic phenotype. Exp
Cell Res. 300:406–417. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Creff G, Safi S, Roques J, Michel H,
Jeanson A, Solari PL, Basset C, Simoni E, Vidaud C and Den Auwer C:
Actinide(IV) deposits on bone: Potential role of the
osteopontin-thorium complex. Inorg Chem. 55:29–36. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Schwetz V, Pieber T and Obermayer-Pietsch
B: The endocrine role of the skeleton: Background and clinical
evidence. Eur J Endocrinol. 166:959–967. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lee NK, Sowa H, Hinoi E, Ferron M, Ahn JD,
Confavreux C, Dacquin R, Mee PJ, McKee MD, Jung DY, et al:
Endocrine regulation of energy metabolism by the skeleton. Cell.
130:456–469. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Renn J and Winkler C: Osterix/Sp7
regulates biomineralization of otoliths and bone in medaka (Oryzias
latipes). Matrix Biol. 34:193–204. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Dong Q, Fu L, Zhao Y, Tan S and Wang E:
Derlin-1 overexpression confers poor prognosis in muscle invasive
bladder cancer and contributes to chemoresistance and invasion
through PI3K/AKT and ERK/MMP signaling. Oncotarget. 8:17059–17069.
2017.PubMed/NCBI
|
|
56
|
Huang J and Manning BD: The TSC1-TSC2
complex: A molecular switchboard controlling cell growth. Biochem
J. 412:179–190. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Magnuson B, Ekim B and Fingar D:
Regulation and function of ribosomal protein S6 kinase (S6K) within
mTOR signalling networks. Biochem J. 441:1–21. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Swiech L, Perycz M, Malik A and Jaworski
J: Role of mTOR in physiology and pathology of the nervous system.
Biochim Biophys Acta. 1784:116–132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Inoki K, Ouyang H, Li Y and Guan KL:
Signaling by target of rapamycin proteins in cell growth control.
Microbiol Mol Biol Rev. 69:79–100. 2005. View Article : Google Scholar : PubMed/NCBI
|