Introduction
The innermost layer of the intestinal mucosa, the
simple columnar epithelium lining the intestinal tract, confers the
barrier/filter function to the intestinal tissue, thus allowing an
optimum exchange of elements between the body and the external
environment. This exchange takes place either through the
epithelial cells (transcellular permeability), or via the
interepithelial space (paracellular transport), with the latter
mainly being regulated by multi-protein complexes, known as tight
junctions (TJs) (1). The
maintenance of an optimal intestinal permeability is further
supported by two auxiliary elements; first, the gel-like layer on
the luminal surface of the intestinal epithelium, consisting of
mucins and antimicrobial peptides, secreted by the epithelial cells
(2), and serving as a potent
chemical barrier (3); second, the
immune modulation of the intestinal integrity, initially by
microfold and antigen presenting cells (3) and ultimately by lymphocytes and
cytokines (1).
Thermal injury, one of the leading causes of
mortality and morbidity worldwide (4), has long been linked to alterations in
intestinal permeability (5). More
specifically, following severe burn, there is an initially
increased systemic inflammatory and metabolic response, marked by
elevated levels of reactive oxygen species (ROS) and cytokines,
such as tumor necrosis factor α (TNF-α), that eventually affect
secondary organs (6). In the gut,
research has revealed increased intestinal epithelial apoptosis
(7), as well as TJ breakdown
secondary to thermal injury (8),
thus attenuating the intestinal integrity. Furthermore, burn
affects the gut immune homeostasis, and therefore the immune aspect
of the intestinal barrier, as indicated by alterations in T-cell
populations of intestinal origin (9). To add an extra layer of complexity to
the dysregulation of the intestinal permeability following thermal
injury, the aforementioned post-burn barrier alterations have been
linked to dramatic shifts in the bacterial gut microbiome (10). In turn, the composition of the
intestinal microbial population has been related to potent changes
in the gut permeability (11),
which then predisposes to bacterial translocation to
extra-intestinal tissues and subsequent sepsis (10).
In addition to thermal injury, infection of the burn
eschar is another major determinant of the intestinal barrier
dysfunction. Subsequent to burn wound infection, systemic
dissemination of the infectious agent, facilitated by the
immunosuppression that follows the initial post-burn
hyper-inflammatory state (6),
gives rise to sepsis (12). Both
the wound infection itself and the ensuing septic state have, in
turn, been shown to affect the intestinal barrier function. Studies
in animals (13) and humans
(14) indicate that post-burn
infection significantly increases the gut permeability compared to
burn alone. What is more, it has previously been suggested that
when superimposed on burn, wound infection and the resultant sepsis
can prolong the duration of the intestinal hyperpermeability in
rats (15).
Despite adequate resuscitation and current
antibiotic treatment, wound infection and subsequent sepsis
jeopardize the function of secondary organs, thus representing a
compelling clinical challenge (16). In this setting, P.
aeruginosa, one of the most common causative microorganisms,
poses a major threat to burn patients (17). This opportunistic Gram negative
bacterial pathogen displays a wide spectrum of antibiotic
resistance (18). We have
developed a promising strategy against this nosocomial pathogen, by
targeting bacterial virulence regulatory systems. More
specifically, we have developed several potent agents targeting
MvfR (also known as PqsR), one of the quorum sensing (QS) cell-cell
bacterial communication systems in P. aeruginosa. MvfR
controls the expression of multiple virulence factors directly and
indirectly, including the expression of the other QS system
regulators LasR and RhlR (19,20).
We have previously shown that our benzamine benzimidazole (BB)
inhibitors specifically bind to the MvfR protein and that targeting
MvfR with non-ligand based antagonists, effectively halts P.
aeruginosa acute and chronic phenotypes, including the
formation of antibiotic-tolerant/persister cells (21–24).
The BB family of anti-MvfR agents we have developed and tested
in vivo, alleviates P. aeruginosa acute infection,
and averts infection relapse after the cessation of the antibiotic
course (21–24). Importantly, these compounds are
expected to thrive where traditional antibiotics fail; by targeting
virulence functions that are not essential for bacterial growth or
survival, they diminish bacterial infectivity and invasiveness,
without imposing a strong selective pressure on the pathogens, thus
potentially reducing the likelihood to generate resistant strains,
while preserving the beneficial enteric flora. In view of our
success in targeting P. aeruginosa virulence functions in
vivo, the present study describes our effort to target the MvfR
QS in the setting of P. aeruginosa burn-wound infection,
aiming to ameliorate the subsequent intestinal barrier dysfunction,
which we found to be significantly affected in our burn-infection
mouse model.
Materials and methods
Mice
Eight-week-old male C57BL/6 mice were purchased from
the Jackson Laboratories. Mice were maintained in a specific
pathogen-free (SPF) environment at the Massachusetts General
Hospital (MGH; Boston, USA), in a 12-h light 12-h dark photoperiod
at an ambient temperature of 22±1°C, with food and water access
ad libitum.
Ethics statement
Animal protocols were reviewed and approved by the
Institutional Animal Care and Use Committee (IACUC) at the MGH
(protocol no. 2006N000093) and are in strict accordance to the
guidelines of the Committee on Animals of the MGH, Harvard Medical
School (Boston, USA), and the regulations of the Subcommittee on
Research Animal Care of the MGH and the National Institutes of
Health. Animals were euthanized according to the guidelines of the
Animal Veterinary Medical Association. All efforts were made to
minimize suffering.
Bacterial strains and inoculum growth
conditions
UCBPP-PA14 (PA14) is a rifampicin resistant P.
aeruginosa human clinical isolate (Rahme laboratory). The
mvfR mutant is isogenic to UCBPP-PA14 (Rahme laboratory)
(25). Unless otherwise indicated,
bacteria were grown in Luria Bertani (LB) broth, LB agar plates, or
LB agar plates containing 100 µg/ml rifampicin.
Animal experiments
All mice were anesthetized using one 500 µl
intraperitoneal (IP) injection of ketamine (125 mg/kg) and xylazine
(12.5 mg/kg) in normal saline (N/S) and the dorsal fur was
subsequently removed with an electric clipper. A 30% total body
surface area (TBSA) dorsal burn was induced by immersion in 90°C
water for 8 sec, using a polystyrene foam template, as in the
well-established burn model described by Walker and Mason (1968),
with some modifications (26).
Spinal protection from the thermal injury was achieved by a dorsal
subcutaneous injection of 500 µl N/S, prior to the induction of the
burn injury. Fluid resuscitation and pain prevention following burn
were achieved by a 100 µl subcutaneous injection of buprenorphine
in N/S (0.3 mg/ml), in a non-burnt area. Sham animals underwent all
procedures except for the thermal injury.
Immediately after burn, 100 µl of 10 mM
MgSO4 containing approximately 105 colony
forming units (CFUs) of P. aeruginosa clinical isolate PA14
culture, or isogenic mvfR mutant culture, were intradermally
injected at the burn eschar of mice in the burn plus infection (BI)
group. Mice in the sham and burn alone groups received an
equivalent injection of 100 µl phosphate-buffered saline (PBS).
After the experiment, all animals were returned to their cages, to
allow recovery from anesthesia. During this period, all cages were
kept on heating pads to prevent hypothermia. Food and hydrogel on
the cage floor were provided ad libitum.
For the group supplemented with our MvfR-inhibiting
compound [(M64-structure fully described and depicted in the
publication by Starkey et al (24)], mice received four intravenous
(tail vein) injections at 2, 4, 8 and 16 h post-BI, at a dose of 4
mg/kg body weight. Control groups received equivalent doses of
dimethylsulfoxide (DMSO) vehicle.
In vivo intestinal permeability
assay
For the assessment of the intestinal barrier
function, 4 h prior to euthanasia, mice were gavaged with 0.2 ml of
Fluorescein Isothiocyanate-Dextran (FITC-Dextran; 3–5 kDa; cat. no.
FD4; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) in PBS, so that
a dose of 440 mg/kg body weight was achieved. 18–20 h post BI, mice
were euthanized. Aseptic cardiac puncture was performed to obtain
blood samples. The collected blood was kept on ice and then
centrifuged at 21,000 × g for 10 min. The serum was removed and was
used to assess the FITC levels with fluorescent spectrophotometry
(excitation, 480 nm and emission, 520 nm). Mice used to assess the
intestinal permeability changes over time, as shown in Fig. 1, were euthanized prior to, or at 4,
10 and 18 h post BI.
Tissue harvesting
Immediately after euthanasia, the mesenteric lymph
nodes (MLNs), ileum and colon were aseptically harvested through
midline laparotomy. The intestine samples were flushed three times
with sterile PBS. The samples were then snap frozen in liquid
nitrogen or stored in RNA-Later and were then frozen at −80°C
(Qiagen Ltd., Manchester, UK) for future analysis.
Stool samples
Stool pellets were collected at 0, 4, 10 and 20 h
post infection. The samples were homogenized in sterile PBS and
were processed as below for fecal lipocalin-2 (Lcn-2)
measurements.
CFU assessment
The MLN tissue was used to assess bacterial
translocation from the intestinal lumen. MLNs were homogenized in
sterile PBS using metallic beads and were serially diluted
1/10-1/1000 and plated on LB agar plates.
Distal ileum was used to assess the PA14
dissemination to the intestinal lumen. The ileal tissue was
homogenized in sterile PBS using metallic beads and was serially
diluted 1/10-1/1000 and plated on LB agar plates containing 100
µg/ml rifampicin.
Systemic blood obtained via cardiac puncture, as
described above, was used to assess bacteremia and was serially
diluted 1/10-1/1000 in sterile PBS and plated on LB agar
plates.
Following inoculation, all plates were incubated at
37°C and CFUs were quantified after 24 h.
TJ assays
Immunofluorescence
Samples of distal ileum were fixed with 4%
paraformaldehyde. Sections then were cut and mounted on microscope
slides. After deparaffinization and antigen retrieval (Antigen
Retrieval reagent; R&D Systems, Minneapolis, MN, USA), tissue
sections were immersed in PBS/0.1 tween for 10 min and were blocked
by Normal Goat Serum. They were then incubated with primary
antibody rabbit polyclonal anti-claudin-1 [ab15098; RRID:
AB_301644; final concentration, 1:100; Abcam, Cambridge, MA, USA
(27)] overnight in a humid
chamber at 4°C. The sections were washed three times with PBS, and
secondary antibody goat anti-rabbit [ab150077; RRID: AB_2630356;
final concentration: 1:500; (28)]
and DAPI (both Abcam, Cambridge, MA, USA) were added. This was
followed by a 1-h incubation at room temperature. The sections were
washed three times with PBS, dried and mounted and images were
collected using a confocal fluorescent microscope (magnification,
×400) (Nikon A1; Nikon Corp., Tokyo, Japan).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The distal ileum was dissected and stored in
RNA-Later at −80°C as described above. RNA was isolated using the
RNeay Plus Mini kit (250) (Qiagen Ltd.), according to the
manufacturer's protocol. cDNA was generated using the iScript TM
Reverse Transcription Supermix for RT-qPCR (Bio-Rad Laboratories,
Inc., Hercules, CA, USA), according to the manufacturer's
instructions. The sequences of the primers used were Claudin 1
forward, 5′-AGTCTTCGACTCCTTGCTGA-3′ and reverse,
5′-ACAAAGATTGCGATCAGCCC-3. Expression of mRNA was normalized with
the reference gene TATA-box Binding Protein (TBP) mRNA expression
(29). RT-qPCR was performed using
the Brilliant II SYBR-Green qPCR Master Mix with Low ROX (Agilent
600830; Agilent Technologies; Santa Clara, CA, USA). The total
volume per reaction was 12 ul, consisting of 6 µl SYBR-Green qPCR
Master Mix with Low ROX, 0.6 µl forward primer (5 µM), 0.6 reverse
primer (5 µM), 3.8 µl distilled water amd 1 µl cDNA sample
(pre-diluted to 1/5). The cycling conditions were as it follows:
95°C for 10 min (1 cycle); 95°C for 30 min (40 cycles); 58°C for 1
min (40 cycles); 72°C for 1 min (40 cycles).
Intestinal inflammation
assessment
Serum and distal ileal TNF-α, and fecal Lcn-2 were
quantified using the mouse TNF-α ELISA Ready-SET-Go kit
(eBioscience; San Diego, CA, USA) and the Mouse Lipocalin-2/NGAL
DuoSet ELISA (R&D Systems) respectively, as per the
manufacturer's instructions.
Endotoxin quantification
Serum lipopolysaccharide (LPS) levels were
determined using a commercially available Limulus Amebocyte Lysate
(LAL) assay kit (ToxinSensor™ Chromogenic LAL Endotoxin Assay kit;
GenScript, Piscataway, NJ, USA), following the manufacturer's
protocol. Absorbance for each sample was measured at 545 nm and the
LPS concentration was determined as per the manufacturer's
instructions (LPS concentration =0.2618 × absorbance at 545 nm
−0.0012).
Statistical analysis
Triplicate samples were used for all assays and all
experiments were repeated at least three times, using 5 mice in
each group. The statistical significance among groups was
determined using one-way analysis of variance (ANOVA), with
multiple post-hoc comparisons using Tukey's test, or two-way ANOVA
analysis, using Bonferroni post-hoc test, as indicated (Graphpad
Software, La Jolla, CA, USA). A P<0.05 was considered
statistically significant.
Results
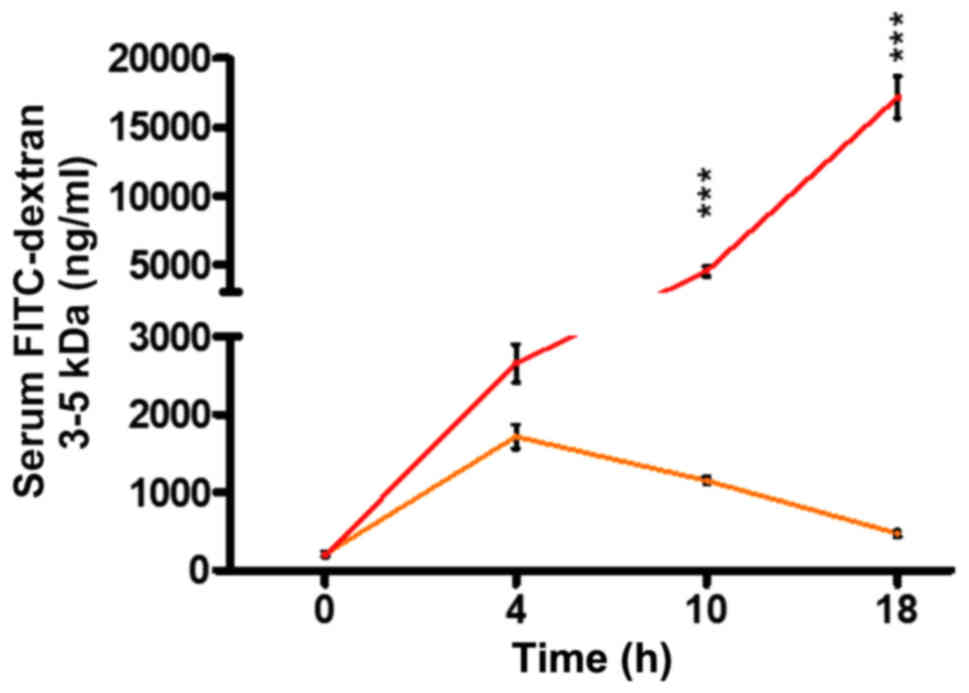
P. aeruginosa infection dramatically
increases the intestinal permeability following thermal injury
FITC-dextran has long and broadly been used as a
reliable marker of intestinal permeability in mice (30,31).
In our study, we used this macromolecule to determine the
contribution of P. aeruginosa in the functional status of
the intestinal barrier in the context of thermal injury. First,
using a burn mouse model, we assessed the intestinal permeability
of mice following burn, or burn plus P. aeruginosa infection
(burn-infection, BI), at several time points, by assessing
FITC-dextran 3–5 kDa flux from the intestinal lumen to the systemic
circulation. Fig. 1 shows that in
our model, gut permeability increases over time, reaching a peak of
1,714 ng/ml at 4 h following the induction of thermal injury, and
then gradually returns to almost sham levels (472 ng/ml) by 18 h.
However, mice that underwent post-burn infection with the P.
aeruginosa clinical isolate PA14, exhibited a dramatically
increased intestinal permeability compared to burn alone, thus,
allowing larger volumes of FITC to flow out of the intestine (4,539
ng/ml in BI vs. 1,151 ng/ml in burn alone at 10 h following injury;
P<0.001). Furthermore, BI mice exhibited a prolonged rise in gut
permeability levels over time, with FITC concentration reaching
17,166 ng/ml by 18 h, indicating the strong impact of infection on
the intestinal barrier dysfunction.
Inhibition of the P
aeruginosa QS transcription factor
MvfR alleviates the deranged intestinal integrity following
burn-site infection
Inhibition of MvfR activity in vivo has
previously been linked to decreased virulence (24). To determine whether the in
vivo inactivation of MvfR can also ameliorate the intestinal
barrier dysfunction promoted by PA14 burn-site infection, we sought
to investigate the FITC-dextran 3–5 kDa flux changes when using one
of our original well-established MvfR antagonist, M64 (22–24).
We assessed the FITC flux out of the intestine at 18–20 h, when the
burn impact on gut permeability largely returned to the sham levels
(as indicated by animals euthanized at 0 h, ie prior to burn, or
BI), while strong impact of infection on the intestinal barrier
dysfunction is observed (Fig. 1).
Fig. 2A shows that FITC flow from
the intestinal lumen to the systemic circulation was increased by
PA14 in the BI group, compared to burn alone (P<0.001). Notably,
FITC concentration in the serum was significantly decreased in mice
treated with the anti-MvfR compound M64 (10,348 ng/ml), compared to
non-treated animals (17,166 ng/ml; P<0.05). Thus, MvfR
antagonist administration significantly protected the intestinal
integrity in the setting of PA14 burn-site infection, rendering the
gut barrier less permeable (Fig.
2A). The MvfR contribution on the intestinal integrity was
validated in mice infected with the isogenic mvfR mutant,
which we have previously reported that exerts reduced virulence and
mortality in burnt and infected mice (19,24).
Mice infected with the mvfR mutant exhibited considerably
reduced FITC flux outside the intestinal lumen, compared to that
promoted by the isogenic parental strain (P<0.001) (data not
shown; available upon request).
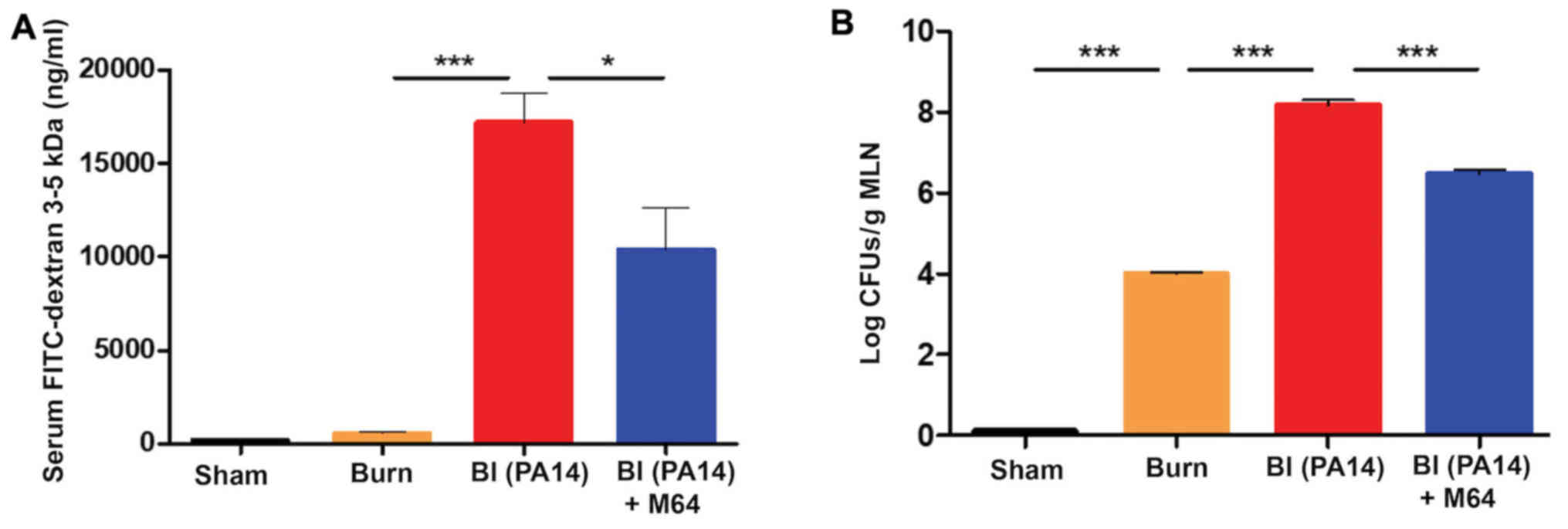 | Figure 2.MvfR antagonists reduce the flow from
the intestinal lumen to the systemic circulation, as well as the
bacterial translocation to the MLNs. (A) FITC-dextran 3–5 kDa flow,
and (B) bacterial translocation from the intestinal lumen to the
systemic circulation at 18 h following burn and infection are
considerably elevated in the BI group, while MvfR antagonist
administration significantly reduces both FITC and bacterial flux
out of the gut. FITC levels were assessed in the serum with
fluorescent spectrophotometry (excitation, 480 nm and emission, 520
nm). LB agar plates were used for the bacterial CFUs assessment in
the MLNs. Black bars, sham; Orange bars, burn; Red bars, PA14
burn-site infection (intradermal administration of 105
CFUs/animal); Blue bars, MvfR antagonist intravenous administration
at 2, 4, 8 and 16 h following burn and infection. Data show the
average +/- SEM (n=5). Statistical significance was assessed using
one-way ANOVA + Tukey's post-hoc test. FITC-dextran, Fluorescein
Isothiocyanate-Dextran; MLNs, mesenteric lymph nodes; BI, burn plus
infection; CFU, colony forming units; LB, Luria Bertani; ANOVA,
one-way analysis of variance. *P<0.05; ***P<0.001. |
Intestinal hyperpermeability after burn injury has
previously been correlated with increased bacterial translocation
from the intestinal lumen to the MLNs (10). Therefore, we interrogated the
impediment of this translocation after MvfR antagonist treatment,
by assessing the bacterial CFUs in the MLNs. Fig. 2B shows that burn alone is
responsible for an increase in bacterial translocation from the
intestine compared to sham (P<0.001), while Pseudomonas
burn-site infection further deregulates the function of the already
defective intestinal barrier, thus doubling the number of bacterial
colonies in the MLNs (P<0.0001). On the contrary, the MvfR
antagonist demonstrates a marked reduction in bacterial MLN
translocation compared to the BI group, with the number of CFUs
being 1.5 log lower in the animals treated with the antagonist
(P<0.001). These results corroborate our findings regarding the
MvfR role in the functional status of the intestinal barrier, and
further indicate that inhibition of MvfR confers significant
protection to the intestinal integrity (Fig. 2B).
MvfR silencing improves morphological
intestinal barrier features in BI mice
Key regulators of the intestinal barrier function
are the TJ protein families. Burn-mediated dysregulation of these
multiple-protein complexes that orchestrate the paracellular gut
permeability, leads to barrier defects (8). Claudins, a family of transmembrane
junctional proteins, seem to be crucial modulators of the
intestinal barrier integrity (32). More specifically, some claudin
family members are considered to be protective ‘tightening’
proteins, while others mostly contribute to intestinal permeability
functions (33). We sought to
evaluate the qualitative and quantitative changes of one of the
‘tightening’ claudin proteins, claudin-1, by exposure-matched
confocal microscopy images and RT-qPCR analysis, respectively.
Fig. 3Ab shows a marked decrease
in staining for claudin-1 in the BI group, compared to sham animals
(Fig. 3Aa), with a less even
distribution at the areas of cell-cell contact and eliminated
delineation of the cell periphery. On the contrary, MvfR inhibition
attenuates this effect, as can be appreciated in Fig. 3Ac, where claudin-1 staining
exhibits a more organized appearance at the periphery of the cells,
with a more uniform localization at the sites of cell-cell
interaction. Quantification of these results is shown in Fig. 3Ad, and it correlates well with our
RT-qPCR data, as shown in Fig. 3B.
PA14 burn wound infection significantly reduces claudin-1 mRNA
levels, as compared to the sham and burn groups (P<0.01).
Conversely, mice treated with MvfR antagonist display an augmented
claudin-1 mRNA expression, in comparison to the BI group
(P<0.05). These data indicate a considerable improvement in the
morphology of the intestinal paracellular transport following MvfR
silencing. Hence, the role of MvfR in the fate of this key
intestinal permeability regulator becomes apparent in the setting
of Pseudomonas burn wound infection.
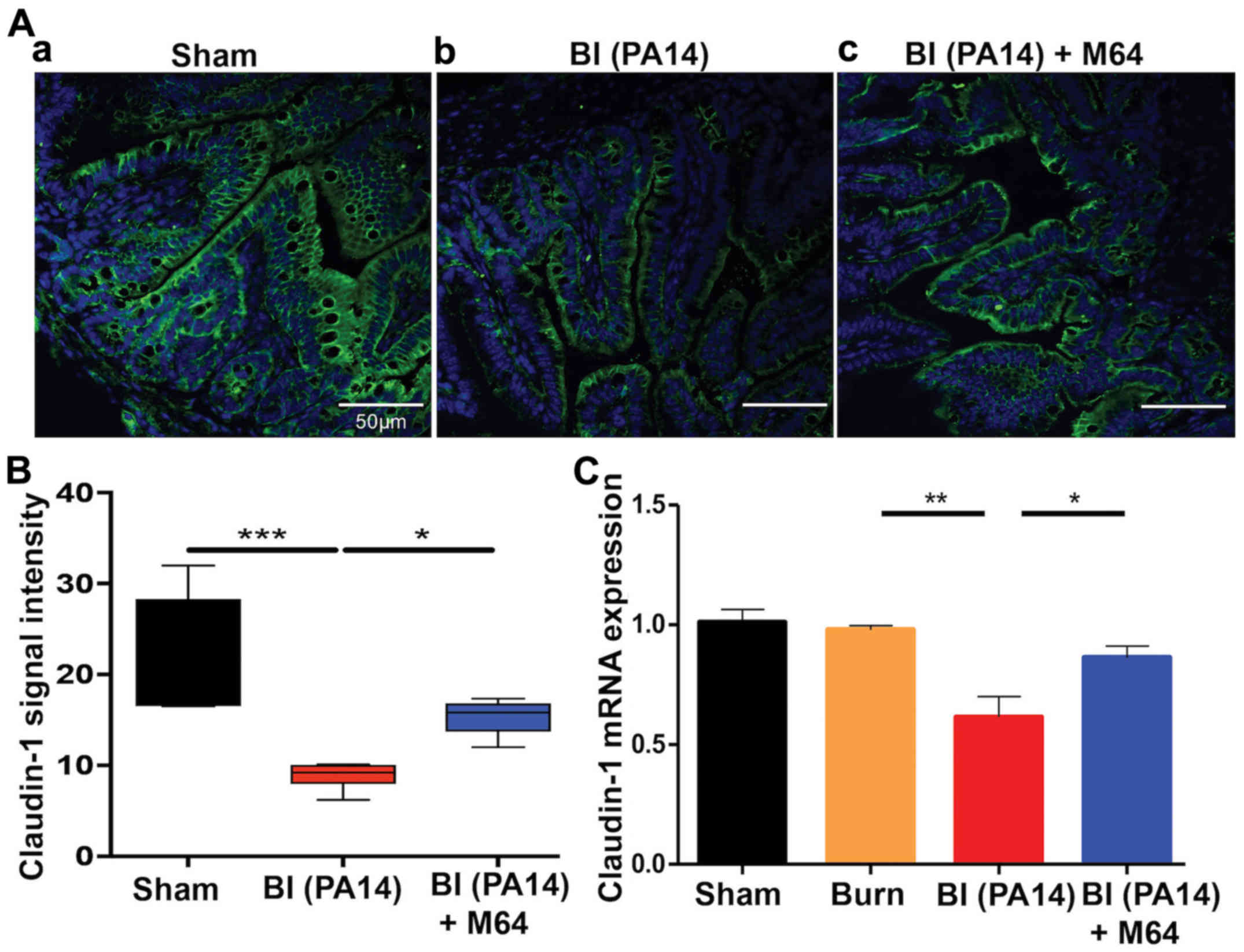 | Figure 3.MvfR antagonists ameliorate the tight
junction downregulation in BI mice. (A) Confocal microscopy images
of distal ileum immunostaining for claudin-1 at 18 h following BI;
(Aa). Representative image from sham animal depicting a uniform,
organized pattern of claudin-1 staining at the periphery of the
cells; (Ab). PA14 burn-site infection dysregulates the smooth,
continuous distribution of claudin-1 and eliminates the cell
periphery delineation; (Ac). Treatment with MvfR antagonist
exhibits a more even localization of claudin-1 at the sites of
cell-cell interaction, with less areas of claudin-1 breakdown. Size
bar, 50 µm. (B) Quantification of confocal microscopy images. (C)
RT-qPCR analysis of claudin-1 mRNA expression levels at 18 h post
BI validates that PA14 burn wound infection downregulates claudin-1
expression, while MvfR inhibition alleviates this effect.
Normalization of mRNA expression was performed for the reference
gene TBP. Black bar, sham; Orange bar, burn; Red bar, PA14
burn-site infection (intradermal administration of 105
CFUs/animal); Blue bar, MvfR antagonist intravenous administration
at 2, 4, 8 and 16 h following burn and infection. Data show the
average +/- SEM (n=5). Statistical significance was assessed using
one-way ANOVA + Tukey's post-hoc test. TBP, TATA-box Binding
Protein; BI, burn plus infection; CFU, colony forming units;
RT-qPCR, reverse transcription-quantitative polymerase chain
reaction. *P<0.05; **P<0.01; ***P<0.001. |
MvfR inhibition alleviates intestinal
inflammation in BI mice
Following deregulation of the intestinal barrier
integrity, microbial paracellular transport out of the lumen cues
an inflammatory response from the intestinal mucosa (34). Similarly, mucosal inflammation is
known to increase TJ permeability, further deranging the
paracellular transport (35), thus
leading to a vicious cycle of defective intestinal integrity. We
therefore sought to determine changes in the levels of intestinal
inflammation following BI, with and without MvfR inhibition.
Fig. 4A demonstrates a sharp rise
of the ileal TNF-α in the BI group (the mean TNF-α level was 58
pg/mg of ileal tissue compared to 11 pg/mg in the burn group;
P<0.001), with a significant reduction achieved after treatment
with the MvfR antagonist, going down to a mean TNF-α concentration
of 38 pg/mg (P<0.01). Similarly, we investigated the changes in
fecal Lcn-2, an acute phase protein that has proven to be a highly
reliable marker of the inflammatory status in the intestinal lumen
(36). Fecal Lcn-2 levels
displayed a gradual time-related rise in both the BI and the
antagonist groups (Fig. 4B), with
the levels of this protein being significantly lower in the burn
alone group at all three time points (4, 10 and 18 h following
insult). Interestingly, there is a clearly disproportionate
elevation of Lcn-2 levels in the BI animals over time, compared to
mice treated with our MvfR antagonist. More specifically, while the
two groups do not significantly differ at 4 h, there is a
progressive difference at 10 (mean for the BI group was 13 ng/mg;
mean for the antagonist group was 10 ng/mg; P<0.01) and 18 h
(mean for the BI group was 19 ng/mg; mean for the antagonist group
was 15 ng/mg; P<0.001). These variations in fecal Lcn-2 levels
further support our observation that P. aeruginosa burn-site
infection promotes, while MvfR inhibition significantly diminishes
inflammation within the intestinal lumen, as shown in Fig. 4B. Taken together, our data support
the contribution of MvfR in the elevated levels of intestinal
inflammation post BI, as well as the significance of the inhibition
of this QS system in this setting.
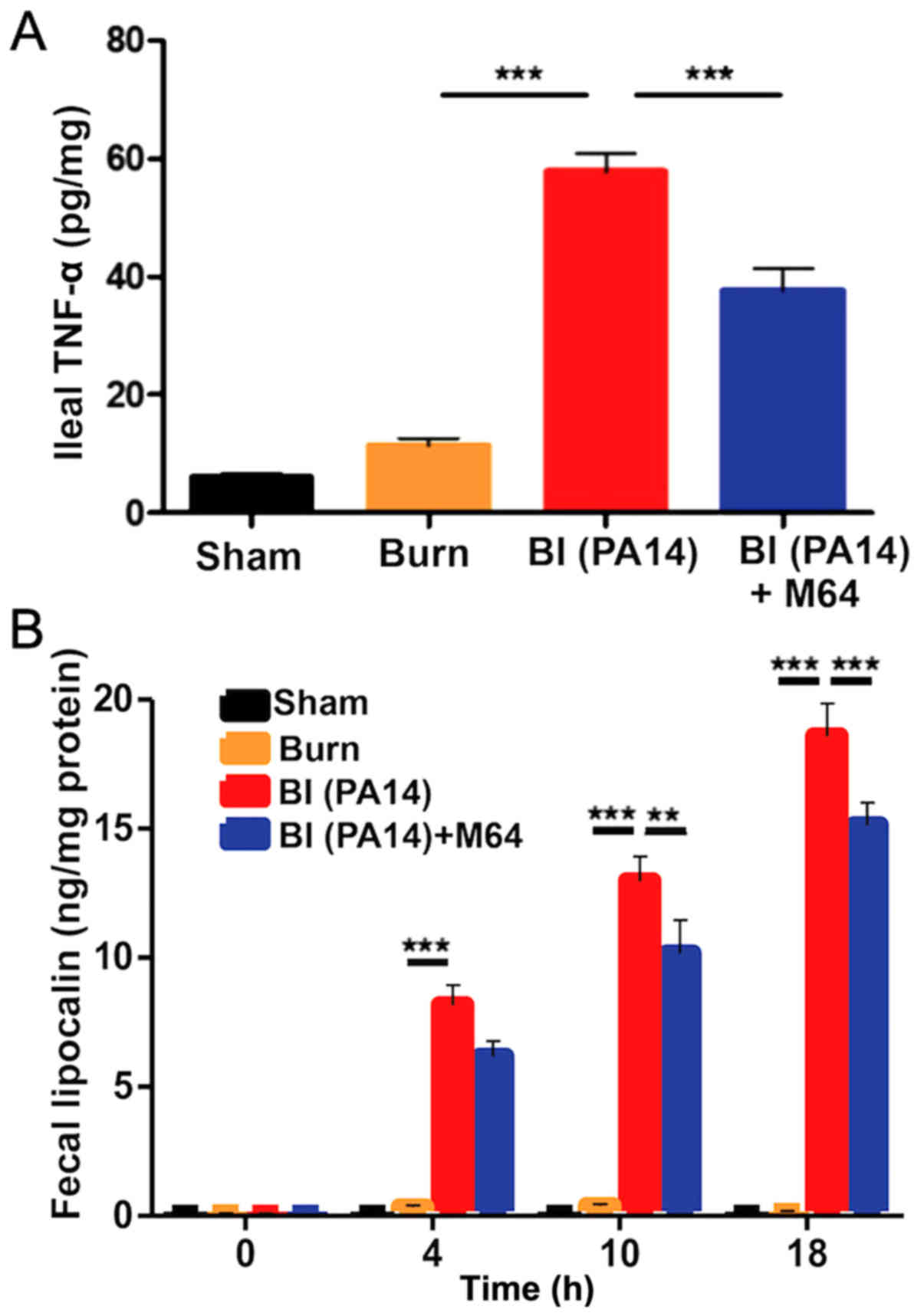 | Figure 4.The role of MvfR in augmenting the
intestinal inflammation post BI. (A) shows TNF-α concentration from
distal ileal tissue at 18 h post BI. Animals that received MvfR
antagonist exhibit significantly reduced TNF-α levels in the ileum.
(B) shows Lcn-2 concentrations from animal feces at 4, 10 and 18 h
following BI. Lcn-2 levels increase with time in both BI and MvfR
antagonist groups, however there is a disproportionate rise in the
Lcn-2 levels compared to the MvfR inhibition group at 18 h. TNF-α
and Lcn-2 levels were assessed using ELISA. Black bars, sham;
Orange bars, burn; Red bars, PA14 burn-site infection (intradermal
administration of 105 CFUs/animal); Blue bars, MvfR
antagonist intravenous administration at 2, 4, 8 and 16 h following
burn and infection. Data show the average +/- SEM (n=5).
Statistical significance was assessed using one-way ANOVA + Tukey's
post-hoc test in Fig. 5A and B,
and two-way ANOVA + Bonferroni correction in Fig. 5C. TNF-α, tumor necrosis factor α;
Lcn-2, lipocalin-2; BI, burn plus infection; ANOVA, one-way
analysis of variance; CFU, colony formation unit.
***P<0.001. |
MvfR inhibition mitigates the post-BI
augmented systemic inflammation and bacterial systemic
dissemination
The surge of the systemic inflammatory response to
burn has been linked to circulating damage-associated molecular
patterns (DAMPs) following thermal injury (37). Besides this, P. aeruginosa
burn wound infection in rodent models has long been recognized to
progress from wound colonization to hematogenous dissemination
(38,39). Consistent with these data, in our
study, superimposition of PA14 infection on thermal injury led to a
significant rise of the systemic inflammatory response, as well as
P. aeruginosa systemic dissemination. More specifically, all
three systemic inflammatory indicators we studied, serum endotoxin
levels, bacterial load in the systemic circulation, and TNF-α
levels in the serum, were substantially elevated in the BI group
compared to burn alone (all P-values were <0.001) (Fig. 5A-C). MvfR antagonist administration
led to a significant reduction in both endotoxin levels (P<0.01)
and bacterial CFUs number (P<0.001) (Fig. 5A and B). TNF-α reduction following
MvfR inhibition failed to reach statistical significance (Fig. 5C). Nevertheless, TNF-α
concentration still follows the same downward trend in the
antagonist group, as observed in the case of endotoxin levels and
bacterial load in the systemic circulation. The above findings were
accompanied by a substantial rise in visceral PA14 load in the BI
group compared to burn alone (P<0.001), as shown by the number
of colonies in the ileal tissue in Fig. 5D. Notably, MvfR inhibition
demonstrated a marked reduction of the PA14 CFUs in the ileum
(P<0.001), indicating that inactivation of MvfR confers a
reduced systemic bacterial dissemination, as expected. Taken
together, these results suggest that this QS system is highly
important for the rise of systemic inflammation and P.
aeruginosa systemic dissemination following burn wound
infection.
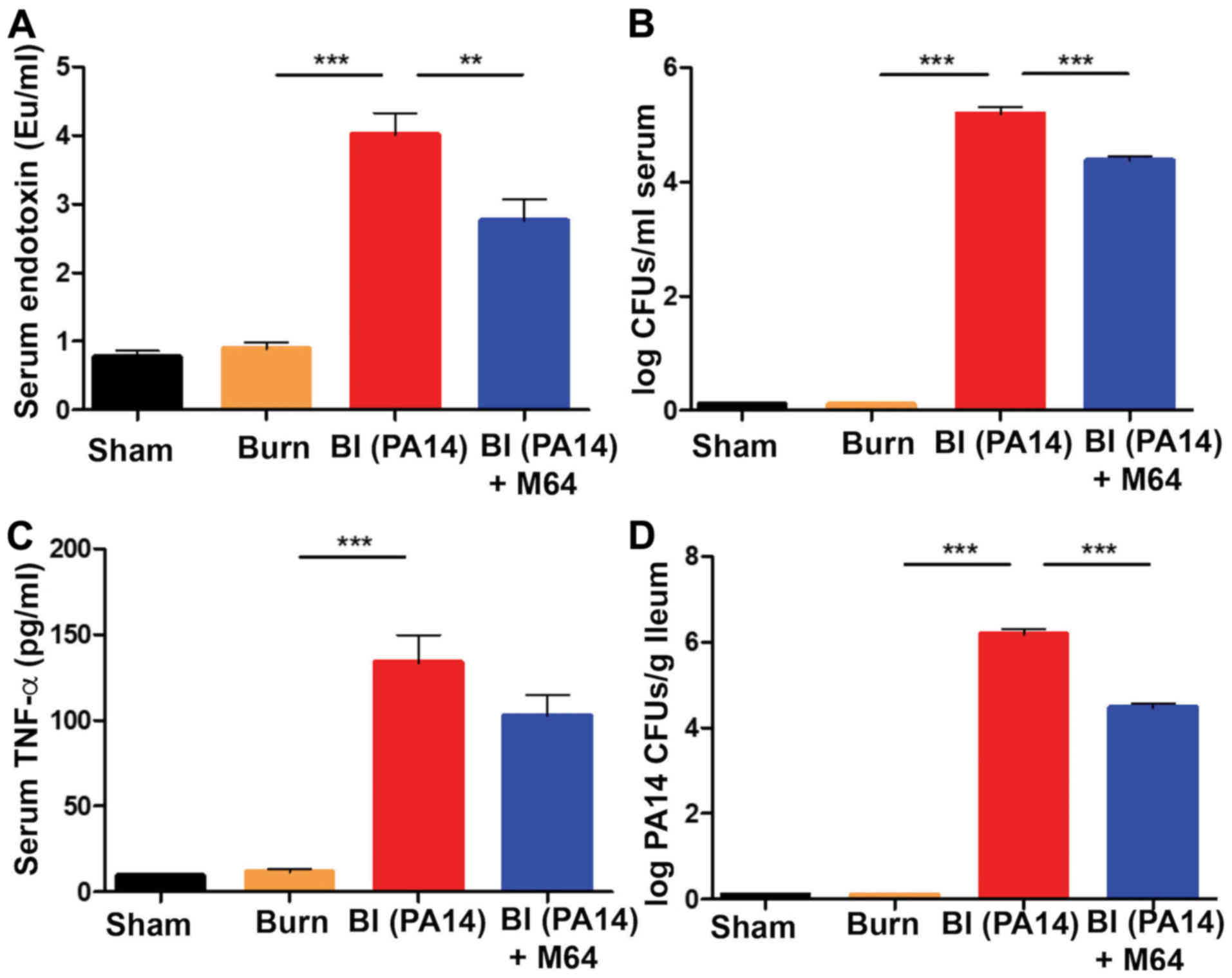 | Figure 5.MvfR inactivation ameliorates the
post-BI elevated systemic inflammation and PA14 systemic
dissemination. (A) Systemic serum endotoxin levels, (B) bacterial
CFUs number, and (C) TNF-α concentration respectively, at 18 h
following BI. A marked elevation of all three systemic inflammation
indicators is demonstrated in the BI group. MvfR inhibition
achieves a significant decrease of endotoxin levels and CFUs number
in the serum. TNF-α concentration also follows the same downward
trend in the MvfR inhibition group, even though it does not reach
statistical significance. (D) Treatment with MvfR antagonist
results in a significant decrease in PA14 CFUs in the ileum,
compared to the BI group, thus representing the respective trends
in PA14 dissemination from the burn wound to the intestine.
Endotoxin levels were determined using a LAL assay; TNF-α was
assessed using ELISA; LB agar plates were used for the bacterial
CFUs assessment in the serum; LB agar plates containing 100 µg/ml
rifampicin were used for the PA14 CFUs assessment in the ileal
tissue. Black bars, sham; Orange bars, burn; Red bars, PA14
burn-site infection (intradermal administration of 105
CFUs/animal); Blue bars, MvfR antagonist intravenous administration
at 2, 4, 8 and 16 h following burn and infection. Data show the
average +/- SEM (n=5). Statistical significance was assessed using
one-way ANOVA + Tukey's post-hoc test. EU, endotoxin units; CFU,
colony formation unit; TNF-α, tumor necrosis factor α; LAL, Limulus
Amebocyte Lysate; ANOVA, analysis of variance. **P<0.01;
***P<0.001. |
Discussion
Despite advances in post-burn resuscitation, P.
aeruginosa burn wound infection, often resistant to the typical
antibiotic regimens, remains a constraining challenge following
thermal injury. Identification of specific P. aeruginosa
traits that contribute to the observed increase in mortality and
morbidity following BI will expedite the development of novel, more
effective therapeutic approaches. Previous studies have
demonstrated that MvfR is a key regulator of multiple P.
aeruginosa virulence factors (19,20,24).
Our team has formulated a promising strategy against this
opportunistic nosocomial pathogen, by developing and utilizing
several potent anti-MvfR agents (21,22,24).
We have previously reported that these robust MvfR-targeting agents
successfully impede P. aeruginosa acute and chronic
phenotypes, both in vitro and in vivo, including the
mortality in thermally injured and infected mice and the ability of
this pathogen to cause relapsing and persistent infection (21–24).
The current study provides novel insights in the understanding of
the MvfR role in the context of intestinal barrier dysregulation
following P. aeruginosa burn wound infection. Indeed, the
data presented here suggest that MvfR functions, in the context of
BI, significantly contribute to the derangement of functional and
morphological traits of the intestinal barrier. We also provide
evidence that MvfR QS plays an important role in the post-BI
modulation of intestinal and systemic inflammation, as well as in
the bacterial systemic dissemination. In agreement with these
findings, we have previously shown that P. aeruginosa
dissemination in the blood stream is decreased as a result of MvfR
regulon inhibition using anthranilic acid analogs (40). Importantly, we herein provide
strong evidence that inhibition of MvfR QS with one of our potent
anti-MvfR BB agents can ameliorate the aforementioned intestinal
and systemic alterations following thermal injury and
infection.
We initially investigated the time-related
variations in intestinal integrity of mice following a 30% dorsal
burn, or after BI. Through evaluation of FITC-dextran flux out of
the intestine at different time points, we observed that the
increase of intestinal permeability exhibits different patterns
depending on the type of the insult. A previous study demonstrated
that burn wound infection with P. aeruginosa (PSA
59-1244) led to an extended period of intestinal barrier
dysfunction, compared to burn alone in Wistar rats (15). This report is consistent with our
finding that while intestinal permeability returns to almost
pre-insult levels at 18 h following burn alone, BI leads to
prolonged barrier dysfunction. Interestingly, our study further
indicates that BI leads to a dramatic exacerbation of the gut
hyperpermeable state compared to burn alone. This is in contrast to
the findings of Jones et al (15) who reported that bacterial
translocation was comparable in the burn and BI groups. This
difference could possibly be attributed to the type of animals
used, mice vs. rats, or to the different P. aeruginosa
strain used in our study. Indeed, the findings of a later study
(13), utilizing a different
strain of P. aeruginosa (P037, Fisher Immunotype-1),
indicate that BI prompts a further increase in the intestinal
permeability compared to burn alone, and are thus in unison with
our present results. Moreover, a clinical study assessing the gut
barrier function following burn, also suggests that thermally
injured and infected patients exhibited higher levels of intestinal
permeability (14).
In agreement with the role of the MvfR-regulon in
P. aeruginosa virulence, our results provide evidence that
MvfR has a significant contribution to the intestinal permeability
alterations following the dual BI insult. We herein demonstrate for
the first time a robust relation between the inhibition of a
crucial P. aeruginosa QS system, and the regulation of the
intestinal integrity following thermal injury and burn wound
infection. Our results provide strong evidence that MvfR silencing
leads to a less prominent induction of gut barrier damage and lower
levels of systemic inflammation. Therefore, these data further
suggest a key role for MvfR in the regulation of intestinal
permeability following burn and P. aeruginosa wound
infection.
A plethora of mechanisms may be involved in the gut
barrier function changes observed post BI. Alterations in
morphological and functional elements of the barrier have been
implicated. Pathological intestinal epithelial cell shedding and
subsequent disruption of the intestinal barrier have previously
been demonstrated in hyperinflammatory states (41), while systemic endotoxin seems to
play a crucial role in the observed gut injury (42). Similarly, we found higher levels of
systemic endotoxin to occur in association with augmented
intestinal hyperpermeability following BI, with MvfR inhibition
diminishing both of these findings. Additionally, disrupted TJ
integrity following burn has been demonstrated in animal models
(8,43,44).
In our present study, claudin-1 was found to be significantly
dysregulated following BI, while MvfR inhibition attenuated this
effect. What is more, cytokine levels such as interferon-γ (INF-γ)
and TNF-α have been implicated in the regulation of paracellular
permeability in the gut (45,46).
These data, along with our finding of elevated TNF-α levels in the
BI group suggest that this cytokine may play a causative role in
the deterioration of the intestinal barrier function. Finally,
microbial dysbiosis in the gut following burn has recently been
shown to precipitate the derangement of the intestinal permeability
(10).
Whatever the mechanism may be, collectively, our
study reveals an intriguing association between P.
aeruginosa QS inhibition and post-BI intestinal barrier
function. This finding highlights the importance of MvfR in this
setting and offers the potential to enrich the therapeutic
armamentarium against P. aeruginosa burn-site infections
with novel and effective virulence-targeting antimicrobial
compounds that we continue to improve, and that could either be
used in combination with antibiotics or as monotherapy. This would
be particularly important in the context of thermal injuries, where
infections with multi-drug resistant P. aeruginosa strains
are very frequent. Follow-up studies could interrogate whether our
compounds attenuate the deranged intestinal function by exerting
additional effects on the host, or on the commensal bacteria
populations residing the gut, thus directly modulating the
molecular components of intestinal dysfunction. Implementation of
clinical investigations to translate our animal-study findings to
patients will be an important future direction of our research
efforts.
Acknowledgements
Not applicable.
Funding
The present study was supported by NIAID grants
AI063433 and AI105902 and was supported by a Shriners Postdoctoral
Research Fellowship from the Shriners Hospitals for Children,
Boston (grant no. 84313).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FA undertook the acquisition and
analysis/interpretation of data, critical revision and final
approval of the manuscript. MA performed analysis/interpretation of
data, manuscript preparation, critical revision and final approval
of the manuscript. MHG was responsible for the acquisition and
analysis/interpretation of data, critical revision and final
approval of the manuscript. MN also undertook the acquisition and
analysis/interpretation of data, critical revision and final
approval of the manuscript. RAH made substantial contribution to
study conception and design, critical revision and final approval
of the manuscript, and LGR also made substantial contribution to
study conception and design, critical revision and gave final
approval of the manuscript.
Ethics approval and consent to
participate
Animal protocols were reviewed and approved by the
IACUC at the MGH (protocol no. 2006N000093) and are in strict
accordance to the guidelines of the Committee on Animals of the
MGH, Harvard Medical School, and the regulations of the
Subcommittee on Research Animal Care of the MGH and the National
Institutes of Health. Animals were euthanized according to the
guidelines of the Animal Veterinary Medical Association. All
efforts were made to minimize suffering.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
QS
|
quorum sensing
|
|
FITC-Dextran
|
Fluorescein Isothiocyanate-Dextran
|
|
MLNs
|
mesenteric lymph nodes
|
|
TJs
|
tight junctions
|
|
ROS
|
reactive oxygen species
|
|
TNF-α
|
tumor necrosis factor α
|
|
BB
|
benzamine benzimidazole
|
|
SPF
|
specific pathogen-free
|
|
MGH
|
Massachusetts General Hospital
|
|
IACUC
|
Institutional Animal Care and Use
Committee
|
|
PA14
|
UCBPP-PA14
|
|
LB
|
Luria Bertani
|
|
IP
|
Intraperitoneal
|
|
TBSA
|
total body surface area
|
|
N/S
|
normal saline
|
|
CFUs
|
colony forming units
|
|
BI
|
burn plus infection
|
|
PBS
|
phosphate-buffered saline
|
|
DMSO
|
dimethylsulfoxide
|
|
Lcn-2
|
lipocalin-2
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
TBP
|
TATA-box Binding Protein
|
|
LPS
|
lipopolysaccharide
|
|
LAL
|
Limulus Amebocyte Lysate
|
|
ANOVA
|
analysis of variance
|
|
DAMPs
|
damage-associated molecular
patterns
|
|
INF-γ
|
interferon-γ
|
References
|
1
|
Groschwitz KR and Hogan SP: Intestinal
barrier function: Molecular regulation and disease pathogenesis. J
Allergy Clin Immunol. 124:3–20. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Muniz LR, Knosp C and Yeretssian G:
Intestinal antimicrobial peptides during homeostasis, infection,
and disease. Front Immunol. 3:3102012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Van Spaendonk H, Ceuleers H, Witters L,
Patteet E, Joossens J, Augustyns K, Lambeir AM, De Meester I, De
Man JG and De Winter BY: Regulation of intestinal permeability: The
role of proteases. World J Gastroenterol. 23:2106–2123. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Forjuoh SN: Burns in low- and
middle-income countries: A review of available literature on
descriptive epidemiology, risk factors, treatment, and prevention.
Burns. 32:529–537. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ryan CM, Yarmush ML, Burke JF and Tompkins
RG: Increased gut permeability early after burns correlates with
the extent of burn injury. Crit Care Med. 20:1508–1512. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nielson CB, Duethman NC, Howard JM,
Moncure M and Wood JG: Burns: Pathophysiology of systemic
complications and current management. J Burn Care Res.
38:e469–ee481. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wolf SE, Ikeda H, Matin S, Debroy MA,
Rajaraman S, Herndon DN and Thompson JC: Cutaneous burn increases
apoptosis in the gut epithelium of mice. J Am Coll Surg. 188:10–16.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Costantini TW, Loomis WH, Putnam JG,
Drusinsky D, Deree J, Choi S, Wolf P, Baird A, Eliceiri B, Bansal V
and Coimbra R: Burn-induced gut barrier injury is attenuated by
phosphodiesterase inhibition: Effects on tight junction structural
proteins. Shock. 31:416–422. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fazal N, Shelip A and Alzahrani AJ:
Burn-injury affects gut-associated lymphoid tissues derived CD4+ T
cells. Results Immunol. 3:85–94. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Earley ZM, Akhtar S, Green SJ, Naqib A,
Khan O, Cannon AR, Hammer AM, Morris NL, Li X, Eberhardt JM, et al:
Burn injury alters the intestinal microbiome and increases gut
permeability and bacterial translocation. PLoS One.
10:e01299962015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Noble EE, Hsu TM and Kanoski SE: Gut to
brain dysbiosis: Mechanisms linking western diet consumption, the
microbiome, and cognitive impairment. Front Behav Neurosci.
11:92017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li N, Hu X, Liu Y, Wang Y, Wang Y, Liu J,
Cai W, Bai X, Zhu X, Han J and Hu D: Systemic inflammatory
responses and multiple organ dysfunction syndrome following skin
burn wound and Pseudomonas aeruginosa infection in mice. Shock.
40:152–159. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ryan CM, Bailey SH, Carter EA, Schoenfeld
DA and Tompkins RG: Additive effects of thermal injury and
infection on gut permeability. Arch Surg. 129:325–328. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ziegler TR, Smith RJ, O'Dwyer ST, Demling
RH and Wilmore DW: Increased intestinal permeability associated
with infection in burn patients. Arch Surg. 123:1313–1319. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jones WG II, Barber AE, Minei JP, Fahey TJ
III, Shires GT III and Shires GT: Differential pathophysiology of
bacterial translocation after thermal injury and sepsis. Ann Surg.
214:24–30. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yan S, Tsurumi A, Que YA, Ryan CM,
Bandyopadhaya A, Morgan AA, Flaherty PJ, Tompkins RG and Rahme LG:
Prediction of multiple infections after severe burn trauma: A
prospective cohort study. Ann Surg. 261:781–792. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Alp E, Coruh A, Gunay GK, Yontar Y and
Doganay M: Risk factors for nosocomial infection and mortality in
burn patients: 10 years of experience at a university hospital. J
Burn Care Res. 33:379–385. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lister PD, Wolter DJ and Hanson ND:
Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and
complex regulation of chromosomally encoded resistance mechanisms.
Clin Microbiol Rev. 22:582–610. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Déziel E, Gopalan S, Tampakaki AP, Lépine
F, Padfield KE, Saucier M, Xiao G and Rahme LG: The contribution of
MvfR to Pseudomonas aeruginosa pathogenesis and quorum sensing
circuitry regulation: Multiple quorum sensing-regulated genes are
modulated without affecting lasRI, rhlRI or the production of
N-acyl-L-homoserine lactones. Mol Microbiol. 55:998–1014. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Maura D, Hazan R, Kitao T, Ballok AE and
Rahme LG: Evidence for direct control of virulence and defense gene
circuits by the Pseudomonas aeruginosa quorum sensing regulator,
MvfR. Sci Rep. 6:340832016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Allegretta G, Maurer CK, Eberhard J, Maura
D, Hartmann RW, Rahme L and Empting M: In-depth profiling of
MvfR-regulated small molecules in Pseudomonas aeruginosa after
quorum sensing inhibitor treatment. Front Microbiol. 8:9242017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maura D, Drees SL, Bandyopadhaya A, Kitao
T, Negri M, Starkey M, Lesic B, Milot S, Déziel E, Zahler R, et al:
Polypharmacology approaches against the Pseudomonas aeruginosa MvfR
regulon and their application in blocking virulence and antibiotic
tolerance. ACS Chem Biol. 12:1435–1443. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maura D and Rahme LG: Pharmacological
inhibition of the Pseudomonas aeruginosa MvfR Quorum-sensing system
interferes with biofilm formation and potentiates
antibiotic-mediated biofilm disruption. Antimicrob Agents
Chemother. 61(pii): e01362–17. 2017.PubMed/NCBI
|
|
24
|
Starkey M, Lepine F, Maura D,
Bandyopadhaya A, Lesic B, He J, Kitao T, Righi V, Milot S, Tzika A
and Rahme L: Identification of anti-virulence compounds that
disrupt quorum-sensing regulated acute and persistent
pathogenicity. PLoS Pathog. 10:e10043212014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rahme LG, Stevens EJ, Wolfort SF, Shao J,
Tompkins RG and Ausubel FM: Common virulence factors for bacterial
pathogenicity in plants and animals. Science. 268:1899–1902. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Walker HL and Mason AD Jr: A standard
animal burn. J Trauma. 8:1049–1051. 1968. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pasternak AJ, Hamonic GM, Van Kessel A and
Wilson HL: Postnatal regulation of MAMDC4 in the porcine intestinal
epithelium is influenced by bacterial colonization. Physiol Rep.
4(pii): e130182016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu L, Zhai Q, Tian F, Liu X, Wang G, Zhao
J, Zhang H, Narbad A and Chen W: Potential of Lactobacillus
plantarum CCFM639 in protecting against aluminum toxicity mediated
by intestinal barrier function and oxidative stress. Nutrients.
8(pii): E7832016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cani PD, Bibiloni R, Knauf C, Waget A,
Neyrinck AM, Delzenne NM and Burcelin R: Changes in gut microbiota
control metabolic endotoxemia-induced inflammation in high-fat
diet-induced obesity and diabetes in mice. Diabetes. 57:1470–1481.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Volynets V, Reichold A, Bardos G, Rings A,
Bleich A and Bischoff SC: Assessment of the intestinal barrier with
five different permeability tests in healthy C57BL/6J and BALB/cJ
mice. Dig Dis Sci. 61:737–746. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gunzel D and Yu AS: Claudins and the
modulation of tight junction permeability. Physiol Rev. 93:525–569.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Markov AG, Veshnyakova A, Fromm M, Amasheh
M and Amasheh S: Segmental expression of claudin proteins
correlates with tight junction barrier properties in rat intestine.
J Comp Physiol B. 180:591–598. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Michielan A and D'Inca R: Intestinal
permeability in inflammatory bowel disease: Pathogenesis, clinical
evaluation, and therapy of leaky gut. Mediators Inflamm.
2015:6281572015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bruewer M, Luegering A, Kucharzik T,
Parkos CA, Madara JL, Hopkins AM and Nusrat A: Proinflammatory
cytokines disrupt epithelial barrier function by
apoptosis-independent mechanisms. J Immunol. 171:6164–6172. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chassaing B, Srinivasan G, Delgado MA,
Young AN, Gewirtz AT and Vijay-Kumar M: Fecal lipocalin 2, a
sensitive and broadly dynamic non-invasive biomarker for intestinal
inflammation. PLoS One. 7:e443282012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Q, Raoof M, Chen Y, Sumi Y, Sursal
T, Junger W, Brohi K, Itagaki K and Hauser CJ: Circulating
mitochondrial DAMPs cause inflammatory responses to injury. Nature.
464:104–107. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Teplitz C, Davis D, Mason AD Jr and
Moncrief JA: Pseudomonas burn wound sepsis. I pathogenesis of
experimental Pseudomonas burn wound sepsis. J Surg Res. 4:200–216.
1964. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Teplitz C, Davis D, Walker HL, Raulston
GL, Mason AD Jr and Moncrief JA: Pseudomonas burn wound sepsis. Ii
hematogenous infection at the junction of the burn wound and the
unburned hypodermis. J Surg Res. 4:217–222. 1964. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lesic B, Lépine F, Déziel E, Zhang J,
Zhang Q, Padfield K, Castonguay MH, Milot S, Stachel S, Tzika AA,
et al: Inhibitors of pathogen intercellular signals as selective
anti-infective compounds. PLoS Pathog. 3:1229–1239. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Williams JM, Duckworth CA, Burkitt MD,
Watson AJ, Campbell BJ and Pritchard DM: Epithelial cell shedding
and barrier function: A matter of life and death at the small
intestinal villus tip. Vet Pathol. 52:445–455. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Williams JM, Duckworth CA, Watson AJ, Frey
MR, Miguel JC, Burkitt MD, Sutton R, Hughes KR, Hall LJ, Caamaño
JH, et al: A mouse model of pathological small intestinal
epithelial cell apoptosis and shedding induced by systemic
administration of lipopolysaccharide. Dis Model Mech. 6:1388–1399.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cannon AR, Akhtar S, Hammer AM, Morris NL,
Javorski MJ, Li X, Kennedy RH, Gamelli RL and Choudhry MA: Effects
of mesalamine treatment on gut barrier integrity after burn injury.
J Burn Care Res. 37:283–292. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Samonte VA, Goto M, Ravindranath TM, Fazal
N, Holloway VM, Goyal A, Gamelli RL and Sayeed MM: Exacerbation of
intestinal permeability in rats after a two-hit injury: Burn and
Enterococcus faecalis infection. Crit Care Med. 32:2267–2273. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Capaldo CT and Nusrat A: Cytokine
regulation of tight junctions. Biochim Biophys Acta. 1788:864–871.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ma TY, Iwamoto GK, Hoa NT, Akotia V,
Pedram A, Boivin MA and Said HM: TNF-alpha-induced increase in
intestinal epithelial tight junction permeability requires NF-kappa
B activation. Am J Physiol Gastrointest Liver Physiol.
286:G367–G376. 2004. View Article : Google Scholar : PubMed/NCBI
|