Introduction
Stroke, universally acknowledged as a
cerebrovascular accident, may result in lasting brain damage,
long-term disability or even mortality (1,2). A
multitude of biological processes are implicated in ischemic
stroke, including oxygen deprivation, neuronal necrosis and an
intense inflammatory response (3,4).
MicroRNAs (miRNAs), long non-coding RNAs (lncRNAs) and even
circular RNAs (circRNAs) contribute to RNA-mediated networks
(5–8) that regulate notable cellular events
through a variety of complicated mechanisms (9,10).
These networks have been implicated in ischemic stroke in previous
studies (5–10); however, there remain gaps in
current knowledge in this regard, and novel ncRNAs need be mined in
order to provide a better understanding of the precise molecular
mechanisms involved in ischemic stroke.
lncRNAs have been proven to be critical gene
regulators of development and disease (11–13).
lncRNAs may also perform functions through competitively binding to
miRNAs known as competitive endogenous RNAs (14). Washietl et al (15) systematically analyzed the
conservatism of human lncRNA and other six mammalian lncRNA and
identified that ~54% human lncRNA loci may be mapped to that of a
rat. A previous study has demonstrated that significantly
differentially expressed lncRNAs (DELs) may contribute to the
stabilization of mRNA expressions in stroke (7). Stroke-induced lncRNAs may also
interact with chromatin-modifying proteins and modulate genes
associated with ischemic brain damage (16,17).
Furthermore, lncRNA BC088414 was revealed to be involved with
apoptosis-associated genes following hypoxic-ischemic brain damage
(8). Similarly, another study
suggested that lncRNA C2dat1 may modulate
calcium/calmodulin-dependent protein kinase II expression to
promote neuronal survival following cerebral ischemia (10). Although a host of lncRNAs have been
identified by massive parallel sequencing, to date, little is known
on functional RNA molecules and RNA-mediated regulation networks in
ischemic stroke.
The main aim of the present study is to elucidate
the lncRNA-mRNA regulation networks in ischemic stroke induced by
middle cerebral artery occlusion (MCAO) using RNA sequencing
(RNA-seq) in rats.
Materials and methods
MCAO model and tissue preparation
A focal cerebral ischemia model induced by MCAO,
prepared as previously described (18), was prepared using 20 7-week-old
male Sprague-Dawley rats of a specific pathogen-free grade
(weighing 200±20 g), purchased from the experimental animal center
of Anhui Medical University (Anhui, China). The study protocol was
ethically approved by the Committee on the Ethics of Animal
Experiments of Anhui University of Chinese Medicine (approval no.
2012AH-036-03). In brief, the animals were fasted overnight but
allowed ad libitum access to water. They were then
anesthetized with chloral hydrate (350 mg/kg, intraperitoneal
injection). A 4-0 silicon-coated monofilament nylon suture with a
round tip was inserted through an arteriectomy in the common
carotid artery just below the carotid bifurcation and then advanced
into the internal carotid artery ~18 mm distal to the carotid
bifurcation until a mild resistance was felt. Following 2 h of
MCAO, the filament was removed to allow reperfusion. As a control,
control-operated rats underwent identical surgery but did not have
the suture inserted. Four days subsequent to MCAO, the left
hemispheres were collected and immediately frozen in liquid
nitrogen.
RNA-seq
RNA-seq was performed by Ao-Ji Bio-Tech (Shanghai,
China). Briefly, total RNA was extracted using an RNeasy Mini kit
(Qiagen GmbH, Hilden, Germany), according to the manufacturer's
protocol. The RNA quality control was performed using Nanodrop 2000
and Agilent 2100, and mainly depended on the concentration, purity
and integrity of the RNA. Ribosomal RNA was removed from total RNA
using Ribo-Zero rRNA removal beads (Illumina, Inc., San Diego, CA,
USA). Libraries were constructed according to the standard TruSeq
protocol (19). Purified cDNA
libraries were prepared for cluster generation and sequencing on an
Illumina HiSeq 2500 (Illumina, Inc.) according to the
manufacturer's protocol. Subsequently, data analyses were performed
in silico.
lncRNA annotation
Quality control of the RNA-Seq reads was conducted
using FastQC (v0.11.3) (The Babraham Institute, Cambridge, UK).
Reads were trimmed using the software seqtk (github.com/lh3/seqtk) for known Illumina TruSeq
adapter sequences, poor reads and ribosome RNA reads. Trimmed reads
were aligned to the rat genome (Rn6) using Hisat2 (version 2.0.4)
(20). Transcripts were assembled
using Stringtie (v1.3.0) (20,21).
Transcripts constructed from Stringtie were compiled together by
gffcompare (v0.9.8) (20,21). Transcripts detected in at least
five samples (half of the total number) were considered to be bona
fide transcripts. Transcripts, with the exception of those with
just one exon and shorter than 200 base pairs, were further
analyzed for the identification of lncRNAs. Transcripts with class
codes ‘i,’ ‘u,’ and ‘x,’ were considered to be potential novel long
transcripts. Pfam (22), Coding
Potential Calculator (CPC) (23)
and Coding-Non-Coding Index (CNCI) (24) were used to estimate the coding
potential of each novel transcript. Transcripts with a Pfam score
<0, CNCI <0 and CPC non-significant were considered to lack
coding potential. Transcripts were compared with annotation
databases, including NONCODE (v4) (http://www.noncode.org) and Ensembl (25). The matched transcripts were
considered to be known lncRNAs, and others were considered to be
novel lncRNAs. All lncRNAs were quantified using Stringtie.
According to the positional association between lncRNA and mRNA in
the genome, lncRNA may be classified into six types: Bidirectional,
exonic_antisense, exonic_sense, intergenic, intronic_antisense and
intronic_sense (26).
The lncRNA-mRNA coexpression
network
Initially, the DELs and differentially expressed
genes (DEGs) were analyzed using EdgeR (27). For DEGs, log2| [fold change (FC)]
|>1 and P<0.05 were used as the cutoff values. Meanwhile,
log2| (FC) |>1 and false discovery rate (FDR) <0.05 were used
as the threshold for DELs. Hierarchical clustering of DELs was
performed based on mean signals using a Euclidean distance
function. In addition, a volcano plot was generated. The Pearson's
correlation coefficient (PCC) between lncRNAs and mRNAs was
calculated (cutoff value, PCC>0.9, P<0.05) and the
lncRNA-mRNA regulatory network was structured using Cytoscape 2.8.3
(28).
Prediction of target genes and
enrichment analysis
cis- or trans-acting algorithms were
used to predict the potential targets of lncRNAs. The first
algorithm predicted potential target genes of cis-acting
lncRNAs that were physically located within 10 kb upstream or 20 kb
downstream of lncRNAs using liftOver genome browser (genome.ucsc.edu/cgi-bin/hgLiftOver). The
second algorithm predicted potential target genes of
trans-acting lncRNAs based on the lncRNA-mRNA complementary
sequences, and predicted lncRNA-mRNA duplex energy. First, BLASTN
(29) was performed to detect
potential target mRNA sequences with >95% identity and E value
<1×10−5 (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Then,
RNAplex (30) was used to
calculate the complementary energy between lncRNAs and their
potential trans-regulated target genes with
RNAplex-10−30. Gene Ontology (GO) (31) and Kyoto Encyclopedia of Genes and
Genomes (KEGG) (32) enrichment
analyses of the identified potential target genes were performed
using the Database for Annotation, Visualization and Integrated
Discovery (33); and P<0.05 was
considered to indicate a statistically significant difference.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from left hemisphere samples
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and reverse-transcribed using a
Thermo Fisher Scientific RevertAid First Strand cDNA Synthesis kit
(cat. no. K1622; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol at 42°C for 60 min. To further confirm the
expression data from RNA-seq, a cutoff value (FC>2, P<0.05)
was randomly selected for qPCR verification. The expression levels
of six randomly DELs (NONRATT027551.2, MSTRG.1836.1, MSTRG.4344.10,
MSTRG.7720.11, NONRATT005132.2 and MSTRG.20633.3) were assayed
using a SYBRGreen flurophore (Applied Biosystems; Thermo Fisher
Scientific, Inc.) using the PikoReal real-time PCR system (Thermo
Fisher Scientific, Inc.) under the following conditions: Initial
denaturation at 95°C for 30 sec, followed by 40 cycles at 95°C for
30 sec and 60°C for 30 sec, and a final extension step at 4°C for
20 min. FC was determined using the 2−ΔΔCq method
(34). GAPDH mRNA was used as an
internal control. The primers used are listed in Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene | Sequence | Polymerase chain
reaction product length (base pairs) |
|---|
| GAPDH | F:
5′-CCTGGTATGACAACGAATTTG-3′ | 131 |
|
| R:
5′-CAGTGAGGGTCTCTCTCTTCC-3′ |
|
|
NONRATT027551.2 | F: 5′-
GGACCTGGAAGGTGAACAGG-3′ | 118 |
|
| R:
5′-TGAATGGGTGACCAACAGGG-3′ |
|
| MSTRG.1836.1 | F:
5′-CCATTGTCCTTCCATCCCCC-3′ | 85 |
|
| R:
5′-CCACCCTACCAAACTTCCCC-3′ |
|
| MSTRG.4344.10 | F:
5′-GACTTAGGCACAGTGGGTGG-3′ | 119 |
|
| R:
5′-ATGGCAGAGAGCGAATGGAG-3′ |
|
| MSTRG.7720.11 | F:
5′-TCCCTAGAGCAGTCCTCACC-3′ | 97 |
|
| R: 5′-
ATCTCGGGTTCGCCTTTTGT-3′ |
|
|
NONRATT005132.2 | F:
5′-CCTGACTATGGCACGTCCTC-3′ | 152 |
|
| R:
5′-CTGAGTCCAGTGTGCCTGTT-3′ |
|
| MSTRG.20633.3 | F:
5′-CTTTCACTCCGAGAACCCCC-3′ | 117 |
|
| R:
5′-GCAAGCAGGTTGGTTCCTTG-3′ |
|
Statistical analysis
The comparisons between the MCAO group and the
control group were determined using a Student's t-test for the
RT-qPCR results by SPSS 22.0 statistical software (IBM Corp.,
Armonk, NY, USA). P<0.05 was considered to indicate a
statistically significant difference. The PCC between lncRNAs and
mRNAs was calculated using the Hmisc package in R based on the
expression determined using RNA-seq (PCC>0.9, P<0.05). The
correlation analysis between the RT-qPCR results and RNA-seq
results was calculated in Excel 2013 (Microsoft Corporation,
Redmond, WA, USA) with the function of CORR.
Results
lncRNA-sequencing data analysis
The present study characterized the lncRNA landscape
and expression by performing deep RNA-seq experiments on three
control and three MCAO tissue samples. Subsequent to the seqtk
quality assessment of sequencing, >33 million total original
reads for each sample were obtained, and the proportion of bases
with quality values >20 was >94%. These results indicated
that the quality of the sequencing results was acceptable (Table II). Subsequent to filtering out
the adaptor sequence and low quality reads, the percentage of clean
reads within the raw reads accounted for 94% of the total sequences
in two groups. Hisat2 software was used to map the obtained clean
reads to the Rattus norvegicus reference genome. As
presented in Table II, ~97% of
the trimmed reads were mapped onto the reference genome. In total,
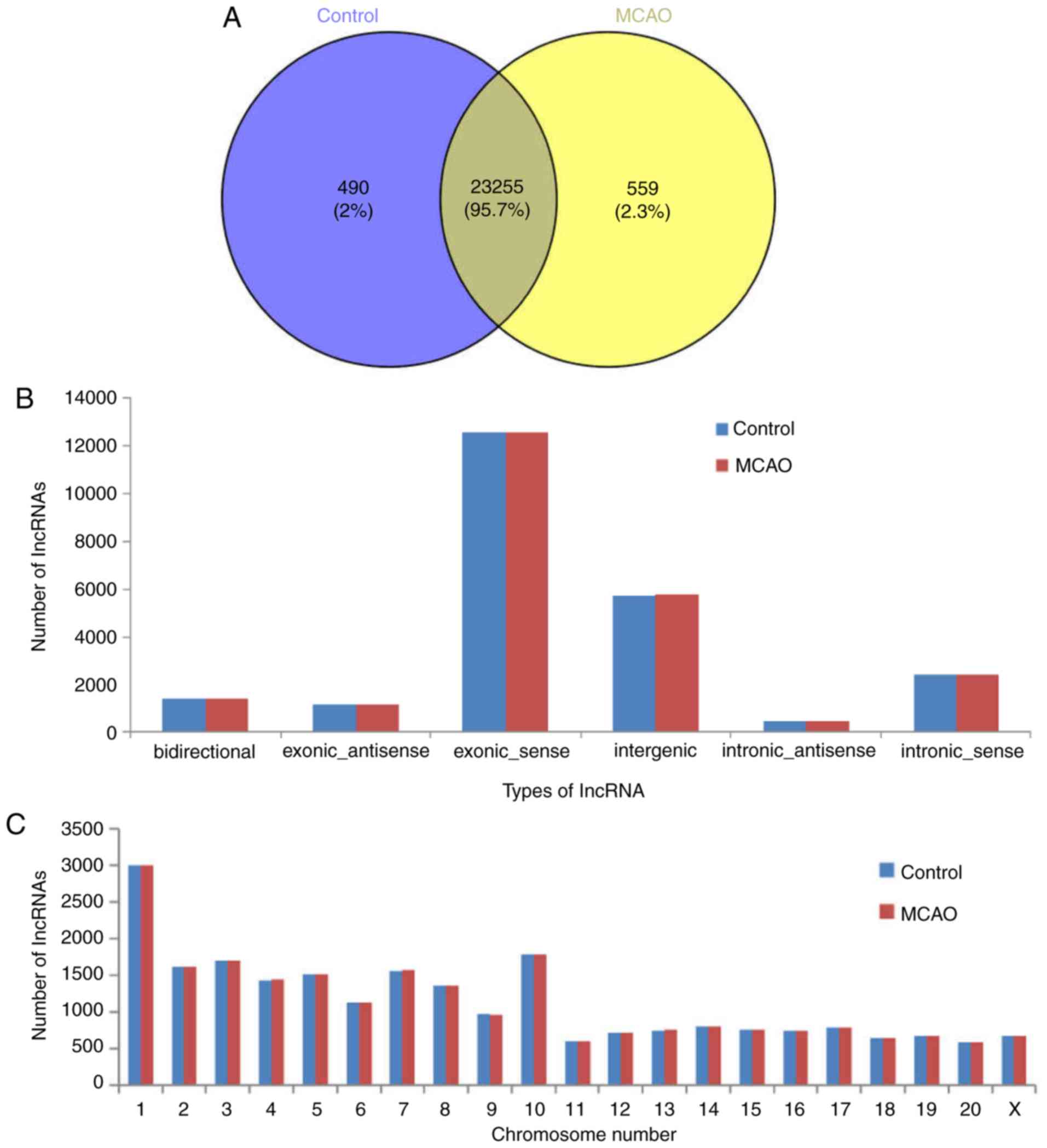
24,304 lncRNAs were screened from six samples, and there were
23,255 shared lncRNAs detected in the MCAO and control groups
(Fig. 1A). The majority of the
identified lncRNAs were transcribed from protein-coding exons;
others were from introns and intergenic regions (Fig. 1B). In addition, the present study
analyzed the distribution of the identified lncRNAs on the rat
chromosomes; 24,304 lncRNA transcripts were identified in all
chromosomes, and chromosome 1 included the most lncRNAs (Fig. 1C).
| Table II.Results of the RNA sequencing. |
Table II.
Results of the RNA sequencing.
| Sample ID | Raw reads | Clean reads | Clean ratio
(%) | rRNA trimmed | Mapped reads | Mapped ratio |
|---|
| MCAO 1 | 155190870 | 147282901 | 94.90 | 147214274 | 131290423 | 0.824026529 |
| MCAO 2 | 144450130 | 136930064 | 94.79 | 136768708 | 120311195 | 0.812346697 |
| MCAO 3 | 151411986 | 142862439 | 94.35 | 142724911 | 125540743 | 0.804354762 |
| Control 1 | 169303916 | 160544960 | 94.83 | 160466568 | 142289885 | 0.819832697 |
| Control 2 | 136533930 | 129672892 | 94.97 | 129609605 | 116072061 | 0.828973282 |
| Control 3 | 124878376 | 118432971 | 94.84 | 118349299 | 105506286 | 0.821661742 |
Identification of DEGs and DELs
EdgeR was used to filter DEGs and DELs and
differentiate their expression between the control and MCAO groups.
A total of 1,007 DEGs (|FC|>2, P<0.05) were identified,
including 785 upregulated genes and 222 downregulated genes.
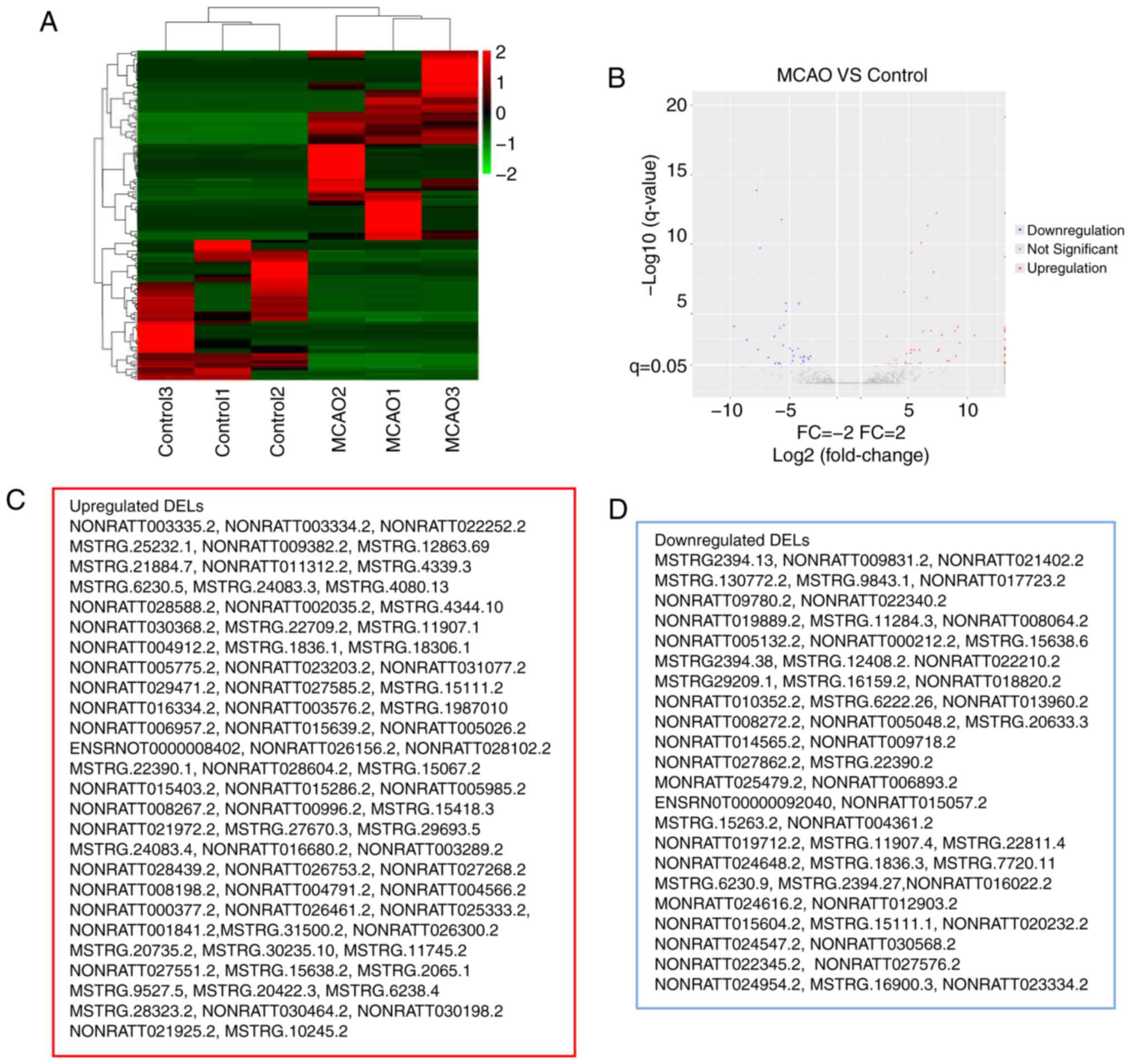
Similarly, as presented in Fig. 2,
134 DELs (|FC|>2, FDR<0.05) were identified in the MCAO group
(Fig. 2A and B), including 77
upregulated and 57 downregulated DELs (Fig. 2C and D). In the present study, it
was revealed that the FC values of certain DELs were equal to
positive infinity and negative infinity, meaning that these lncRNAs
are switched-on or off with MCAO. Essentially, positive or negative
infinity indicates zero expression of the lncRNA in normal or MCAO
groups. It was speculated that this may be associated with the
abundance of lncRNAs and the sensitivity to RNA-seq. The top five
upregulated DELs were NONRATT027551.2, MSTRG.1836.1, MSTRG.4344.10,
NONRATT028102.2 and MSTRG.31500.2; the top five downregulated DELs
were MSTRG.7720.11, NONRATT005132.2, MSTRG.20633.3, NONRATT020232.2
and MSTRG.1836.3.
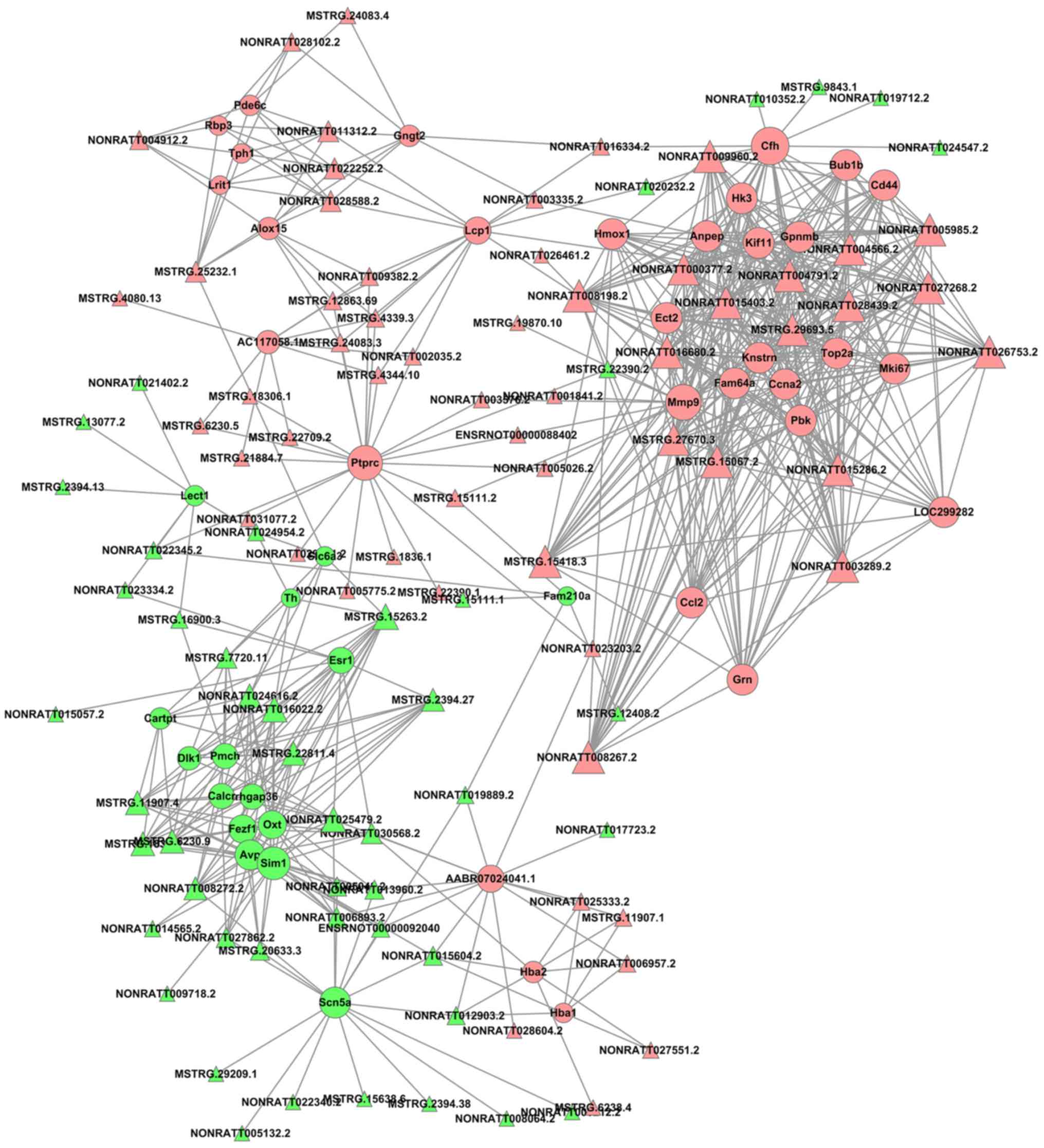
lncRNA-mRNA network
The cutoff correlation r-values (|PCC|>0.9) and
P-values (P<0.05) were selected to structure a lncRNA-mRNA
co-expression network between DEGs and DELs. As Fig. 3 presents, 46 DEGs, 104 DELs and 664
edges were filtered out using Cytoscape to construct the
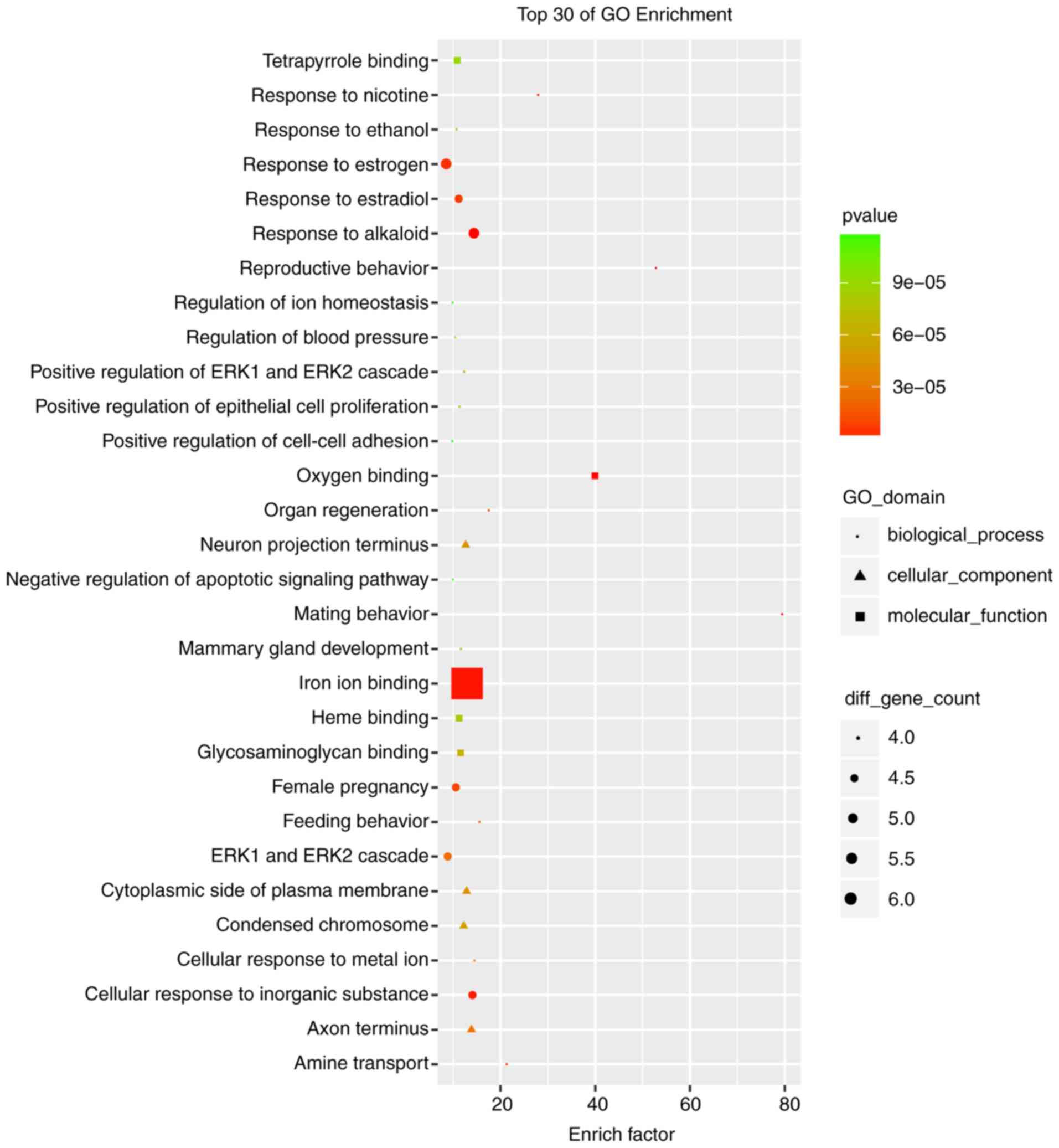
co-expression network. The co-expression-associated top 30 GO terms
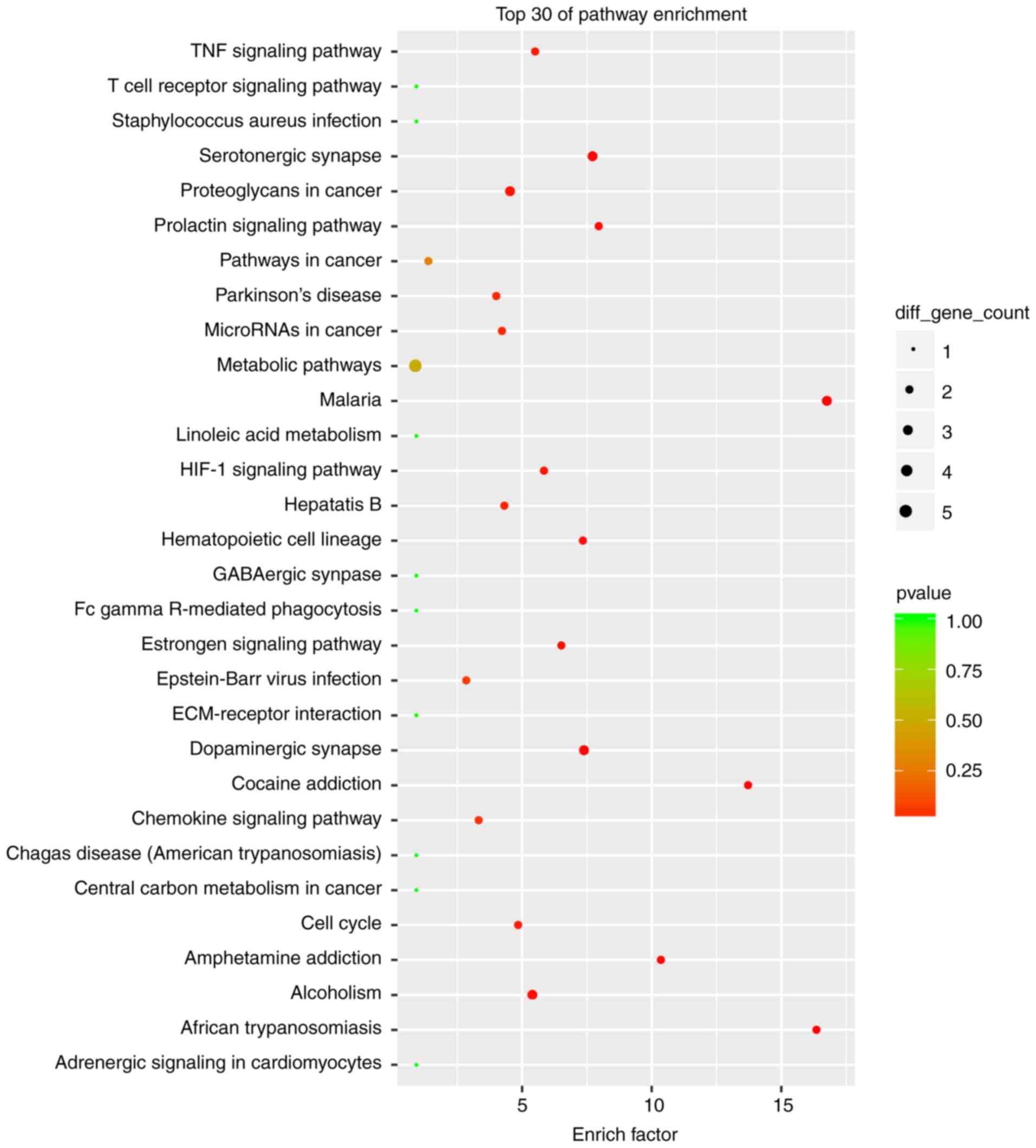
and pathway terms enrichment analyses presented in Figs. 4 and 5 suggest that these DELs were associated
with the cellular response to inorganic substances, alkaloids,
estradiols, reactive oxygen species, metal ions and oxidative
stress. In particular, the heme oxygenase 1 (HO-1) gene
participates in many of these functions. A multitude of pathways
were implicated, including metabolic pathways, chemokine signaling
pathways, malaria, Parkinson's disease and the cell cycle. Notably,
the BUB1 mitotic checkpoint serine/threonine kinase B (BUB1B) and
C-C motif chemokine ligand 2 (CCL2) genes were associated with the
cell cycle.
Regulatory analysis of DELs
A total of 91 cis-regulatory genes of 94
DELs, including 55 upregulated lncRNAs in the MCAO group were
identified; 14 of the 91 cis-regulatory genes exhibited
differential expression. A total of 13 of the
DEL/cis-regulatory gene pairs had positive correlations as
follows: NONRATT021925.2 (Rho GDP dissociation inhibitor β),
NONRATT004791.2 (G protein subunit γ transducing 2),
NONRATT015286.2 (periostin), MSTRG.30235.10 (LRR binding FLII
interacting protein 1), NONRATT015403.2 (IQ motif containing GTPase
activating protein 3), NONRATT008267.2 (kinesin family member 14),
NONRATT009960.2 (non-SMC condensin I complex subunit G),
NONRATT011312.2 (retinol binding protein 3), NONRATT005985.2 (DNA
topoisomerase IIα), NONRATT016680.2 (ENSRNOG00000053081),
NONRATT013960.2 (galanin receptor 1), NONRATT016022.2 (CART
prepropeptide) and NONRATT024616.2 (δ like non-canonical Notch
ligand 1) (Table III).
Additionally, 90 trans-regulatory genes of lncRNAs were
filtered by BLASTN and RNAplex, with a negative correlation
identified between ENSRNOT00000092040 and Ccl9 (Table IV).
| Table III.Differentially expressed lncRNAs
mechanisms involved in cis-regulatory elements. |
Table III.
Differentially expressed lncRNAs
mechanisms involved in cis-regulatory elements.
| lncRNA ID | log2FC | Q value |
Up/downregulated | Type | Gene ID | Gene name | log2FC | P-value |
Up/downregulated |
|---|
|
NONRATT017723.2 | −3.68 | 0.0125 | Down | cis |
ENSRNOG00000000633 | Rhobtb1 | 0.85 | 0.1148 | – |
|
NONRATT008064.2 | −6.73 | 0.0145 | Down | cis |
ENSRNOG00000001183 | Hnf1a | 1.25 | 0.7907 | – |
|
NONRATT000377.2 | +∞ | 0.0492 | Up | cis |
ENSRNOG00000001492 | Slc8a2 | 0.13 | 0.9091 | – |
|
NONRATT006957.2 | 6.69 | 0.0000 | Up | cis |
ENSRNOG00000001982 | Cblb | 0.60 | 0.1402 | – |
|
NONRATT009831.2 | −5.72 | 0.0425 | Down | cis |
ENSRNOG00000002137 | Aasdh | 0.21 | 0.7048 | – |
|
NONRATT009718.2 | −5.48 | 0.0020 | Down | cis |
ENSRNOG00000002146 | Pkd2 | 0.29 | 0.5035 | – |
|
NONRATT009780.2 | −3.94 | 0.0473 | Down | cis |
ENSRNOG00000002208 | Shroom3 | 0.83 | 0.1301 | – |
|
NONRATT005026.2 | 6.14 | 0.0000 | Up | cis |
ENSRNOG00000002607 | Sox9 | 0.25 | 0.5566 | – |
|
NONRATT014565.2 | −2.91 | 0.0492 | Down | cis |
ENSRNOG00000002864 | Nacc1 | 0.26 | 0.5744 | – |
|
NONRATT005048.2 | -∞ | 0.0432 | Down | cis |
ENSRNOG00000003144 | Gprc5c | 0.48 | 0.5195 | – |
|
NONRATT024547.2 | -∞ | 0.0082 | Down | cis |
ENSRNOG00000003955 | Spata7 | 0.26 | 0.6325 | – |
|
NONRATT026753.2 | 3.08 | 0.0450 | Up | cis |
ENSRNOG00000003993 | Thap2 | 0.26 | 0.7629 | – |
|
NONRATT005132.2 | −8.71 | 0.0471 | Down | cis |
ENSRNOG00000004049 | Baiap2 | −0.04 | 0.8211 | – |
|
NONRATT026300.2 | +∞ | 0.0469 | Up | cis |
ENSRNOG00000004628 | Dazap2 | 0.13 | 0.8554 | – |
|
NONRATT026156.2 | 7.44 | 0.0000 | Up | cis |
ENSRNOG00000005332 | Csdc2 | −0.58 | 0.1263 | – |
|
NONRATT005775.2 | 3.69 | 0.0453 | Up | cis |
ENSRNOG00000005538 | Psmd11 | −0.03 | 0.7616 | – |
|
NONRATT023334.2 | −4.85 | 0.0448 | Down | cis |
ENSRNOG00000005711 | Ptprd | −0.21 | 0.4658 | – |
|
NONRATT021925.2 | +∞ | 0.0000 | Up | cis |
ENSRNOG00000005809 | Arhgdib | 1.81 | 0.0074 | Up |
|
NONRATT004791.2 | 4.02 | 0.0218 | Up | cis |
ENSRNOG00000006108 | Gngt2 | 4.95 | 0.0000 | Up |
| MSTRG.22811.4 | -∞ | 0.0388 | Down | cis |
ENSRNOG00000006966 | Nfia | 0.21 | 0.6590 | – |
|
NONRATT025479.2 | −4.14 | 0.0000 | Down | cis |
ENSRNOG00000007610 | Gdf11 | 0.08 | 0.9402 | – |
|
NONRATT020232.2 | −7.72 | 0.0000 | Down | cis |
ENSRNOG00000007804 | C1galt1 | 0.45 | 0.2729 | – |
|
NONRATT008198.2 | 4.83 | 0.0305 | Up | cis |
ENSRNOG00000007887 | Elk4 | 0.76 | 0.2762 | – |
|
NONRATT022252.2 | +∞ | 0.0305 | Up | cis |
ENSRNOG00000008099 | Galnt12 | 0.85 | 0.2496 | – |
|
NONRATT024954.2 | −3.98 | 0.0471 | Down | cis |
ENSRNOG00000008155 | Dus4l | −0.17 | 0.7340 | – |
|
NONRATT028439.2 | 5.06 | 0.0430 | Up | cis |
ENSRNOG00000008187 | Ubash3b | −0.21 | 0.4764 | – |
|
NONRATT022210.2 | −4.24 | 0.0492 | Down | cis |
ENSRNOG00000008237 | Unc13b | 0.30 | 0.6027 | – |
|
NONRATT027551.2 | 9.08 | 0.0119 | Up | cis |
ENSRNOG00000008709 | Arhgap32 | −0.24 | 0.5090 | – |
|
NONRATT027576.2 | -∞ | 0.0061 | Down | cis |
ENSRNOG00000008757 | Tmem218 | 0.16 | 0.8511 | – |
|
NONRATT021402.2 | -∞ | 0.0291 | Down | cis |
ENSRNOG00000009156 | Tra2a | 0.47 | 0.2267 | – |
|
NONRATT021972.2 | +∞ | 0.0335 | Up | cis |
ENSRNOG00000009338 | Kras | −0.11 | 0.6201 | – |
|
NONRATT022345.2 | −5.23 | 0.0000 | Down | cis |
ENSRNOG00000009795 | Nfib | 0.04 | 0.9239 | – |
| MSTRG.16900.3 | −4.70 | 0.0248 | Down | cis |
ENSRNOG00000009882 | Ppp3ca | 0.07 | 1.0000 | – |
| MSTRG.19870.10 | +∞ | 0.0000 | Up | cis |
ENSRNOG00000010993 | Dpm1 | −0.14 | 0.5966 | – |
|
NONRATT008272.2 | −3.96 | 0.0431 | Down | cis |
ENSRNOG00000011063 | Dennd1b | 0.20 | 0.7240 | – |
| MSTRG.10245.2 | 6.31 | 0.0002 | Up | cis |
ENSRNOG00000011704 | Fbxo34 | −0.37 | 0.2745 | – |
|
NONRATT001841.2 | 3.41 | 0.0430 | Up | cis |
ENSRNOG00000012110 | Col17a1 | −1.52 | 0.0483 | Down |
|
NONRATT002035.2 | 7.89 | 0.0003 | Up | cis |
ENSRNOG00000012324 | Soga3 | −0.04 | 0.7913 | – |
| MSTRG.22390.2 | -∞ | 0.0000 | Down | cis |
ENSRNOG00000012634 | Fbxo10 | −0.41 | 0.1989 | – |
| MSTRG.22390.1 | +∞ | 0.0000 | Up | cis |
ENSRNOG00000012634 | Fbxo10 | −0.41 | 0.1989 | – |
|
NONRATT015286.2 | 5.30 | 0.0425 | Up | cis |
ENSRNOG00000012660 | Postn | 3.66 | 0.0004 | Up |
| MSTRG.1836.3 | −7.60 | 0.0041 | Down | cis |
ENSRNOG00000012716 | Chd2 | 0.06 | 0.9929 | – |
| MSTRG.1836.1 | 9.24 | 0.0001 | Up | cis |
ENSRNOG00000012716 | Chd2 | 0.06 | 0.9929 | – |
|
NONRATT016334.2 | +∞ | 0.0000 | Up | cis |
ENSRNOG00000012734 | Dcun1d1 | −0.20 | 0.5140 | – |
|
NONRATT015057.2 | −4.70 | 0.0049 | Down | cis |
ENSRNOG00000012799 | Prkaa1 | −0.28 | 0.3193 | – |
|
NONRATT000212.2 | −6.12 | 0.0378 | Down | cis |
ENSRNOG00000013194 | Rps6ka2 | −0.14 | 0.5704 | – |
|
NONRATT030198.2 | 5.34 | 0.0000 | Up | cis |
ENSRNOG00000013213 | Epha4 | 0.35 | 0.5640 | – |
|
NONRATT010352.2 | −6.23 | 0.0004 | Down | cis |
ENSRNOG00000013353 | Tmem260 | 0.07 | 0.9952 | – |
|
NONRATT029471.2 | 4.91 | 0.0082 | Up | cis |
ENSRNOG00000013557 | Lancl1 | −0.17 | 0.5373 | – |
|
NONRATT028588.2 | 6.05 | 0.0378 | Up | cis |
ENSRNOG00000013829 | Chrna3 | 1.17 | 0.2523 | – |
|
NONRATT023203.2 | +∞ | 0.0007 | Up | cis |
ENSRNOG00000013956 | Rnf38 | 0.17 | 0.7841 | – |
| MSTRG.29693.5 | +∞ | 0.0484 | Up | cis |
ENSRNOG00000013991 | Creg2 | −0.69 | 0.2339 | – |
| MSTRG.12408.2 | -∞ | 0.0041 | Down | cis |
ENSRNOG00000014007 | Gfod1 | 0.04 | 0.9704 | – |
|
NONRATT004566.2 | 3.44 | 0.0440 | Up | cis |
ENSRNOG00000015002 | Abhd15 | 0.76 | 0.3266 | – |
| MSTRG.15067.2 | +∞ | 0.0090 | Up | cis |
ENSRNOG00000015334 | Fcho2 | 0.49 | 0.2280 | – |
|
NONRATT003289.2 | 7.56 | 0.0052 | Up | cis |
ENSRNOG00000015717 | Ptpre | 0.21 | 0.6977 | – |
|
NONRATT013960.2 | −3.81 | 0.0248 | Down | cis |
ENSRNOG00000016654 | Galr1 | −3.38 | 0.0001 | Down |
|
NONRATT028604.2 | 4.71 | 0.0000 | Up | cis |
ENSRNOG00000017193 | Lingo1 | −0.52 | 0.2454 | – |
|
NONRATT016022.2 | −5.74 | 0.0001 | Down | cis |
ENSRNOG00000017712 | Cartpt | −3.83 | 0.0001 | Down |
|
NONRATT015604.2 | −5.60 | 0.0000 | Down | cis |
ENSRNOG00000018166 | Prkab2 | 0.00 | 0.8495 | – |
|
NONRATT024616.2 | −3.79 | 0.0143 | Down | cis |
ENSRNOG00000019584 | Dlk1 | −4.21 | 0.0000 | Down |
| MSTRG.30235.10 | 8.50 | 0.0275 | Up | cis |
ENSRNOG00000019892 | Lrrfip1 | 1.14 | 0.0479 | Up |
|
NONRATT026461.2 | +∞ | 0.0118 | Up | cis |
ENSRNOG00000020230 | Pias4 | 0.17 | 0.8050 | – |
|
NONRATT018820.2 | -∞ | 0.0002 | Down | cis |
ENSRNOG00000020337 | Sla2 | 0.52 | 0.5953 | – |
|
NONRATT004912.2 | 6.00 | 0.0041 | Up | cis |
ENSRNOG00000020658 | Aarsd1 | 0.05 | 0.9927 | – |
|
NONRATT019712.2 | −6.20 | 0.0446 | Down | cis |
ENSRNOG00000021262 | Slc23a2 | 0.09 | 0.9742 | – |
|
NONRATT027268.2 | 5.27 | 0.0071 | Up | cis |
ENSRNOG00000022570 | Pus7l | 0.51 | 0.5096 | – |
| MSTRG.12863.69 | +∞ | 0.0304 | Up | cis |
ENSRNOG00000023661 | Celf2 | 0.30 | 0.5495 | – |
|
NONRATT030368.2 | 5.55 | 0.0043 | Up | cis |
ENSRNOG00000025527 | Mtcl1 | 0.16 | 0.8136 | – |
|
NONRATT027862.2 | −3.75 | 0.0304 | Down | cis |
ENSRNOG00000027145 | Rora | −0.16 | 0.5963 | – |
|
NONRATT015403.2 | 4.83 | 0.0409 | Up | cis |
ENSRNOG00000027894 | Iqgap3 | 2.03 | 0.0137 | Up |
|
NONRATT003576.2 | +∞ | 0.0028 | Up | cis |
ENSRNOG00000028017 | Tmem109 | 0.36 | 0.4097 | – |
|
NONRATT004361.2 | −5.86 | 0.0409 | Down | cis |
ENSRNOG00000028341 | Alkbh5 | 0.00 | 0.8591 | – |
|
NONRATT012903.2 | −5.26 | 0.0000 | Down | cis |
ENSRNOG00000031706 | AABR07027388.1 | −0.85 | 0.1761 | – |
| MSTRG.15111.2 | +∞ | 0.0000 | Up | cis |
ENSRNOG00000032735 | Srek1 | 0.18 | 0.7203 | – |
| MSTRG.15111.1 | −7.42 | 0.0000 | Down | cis |
ENSRNOG00000032735 | Srek1 | 0.18 | 0.7203 | – |
|
NONRATT028102.2 | 10.21 | 0.0000 | Up | cis |
ENSRNOG00000033809 | Mlh1 | 0.76 | 0.1272 | – |
|
NONRATT019889.2 | −4.60 | 0.0113 | Down | cis |
ENSRNOG00000034031 | Vstm2l | −0.45 | 0.3771 | – |
|
NONRATT008267.2 | +∞ | 0.0039 | Up | cis |
ENSRNOG00000037211 | Kif14 | 5.80 | 0.0005 | Up |
|
NONRATT009960.2 | 4.06 | 0.0430 | Up | cis |
ENSRNOG00000038572 | Ncapg | 3.28 | 0.0054 | Up |
| MSTRG.21884.7 | +∞ | 0.0492 | Up | cis |
ENSRNOG00000042458 | Stau2 | −0.20 | 0.5104 | – |
|
NONRATT030464.2 | 5.30 | 0.0042 | Up | cis |
ENSRNOG00000046053 | Nudt10 | −0.59 | 0.1573 | – |
| MSTRG.15418.3 | +∞ | 0.0448 | Up | cis |
ENSRNOG00000048993 | Metazoa_SRP | 0.03 | 0.9881 | – |
| MSTRG.28323.2 | +∞ | 0.0492 | Up | cis |
ENSRNOG00000049584 | AABR07070555.1 | −0.13 | 0.8591 | – |
| MSTRG.4080.13 | +∞ | 0.0002 | Up | cis |
ENSRNOG00000049768 | Adcy9 | −0.04 | 0.7770 | – |
|
NONRATT025333.2 | 6.61 | 0.0000 | Up | cis |
ENSRNOG00000051719 | AABR07065498.1 | 0.11 | 0.9304 | – |
|
NONRATT011312.2 | +∞ | 0.0479 | Up | cis |
ENSRNOG00000051911 | Rbp3 | 9.84 | 0.0000 | Up |
|
NONRATT024648.2 | -∞ | 0.0492 | Down | cis |
ENSRNOG00000052540 | SNORD113 | 0.45 | 0.6631 | – |
|
NONRATT005985.2 | 4.53 | 0.0490 | Up | cis |
ENSRNOG00000053047 | Top2a | 5.28 | 0.0000 | Up |
|
NONRATT016680.2 | 7.62 | 0.0041 | Up | cis |
ENSRNOG00000053081 |
| 3.59 | 0.0011 | Up |
|
NONRATT009382.2 | 8.35 | 0.0257 | Up | cis |
ENSRNOG00000056826 | Arap2 | 0.21 | 0.6941 | – |
| MSTRG.15263.2 | -∞ | 0.0041 | Down | cis |
ENSRNOG00000060329 | Emb | −0.06 | 0.7939 | – |
|
NONRATT015639.2 | 3.25 | 0.0005 | Up | cis |
ENSRNOG00000061058 | Csde1 | 0.02 | 0.8739 | – |
|
NONRATT027585.2 | 7.19 | 0.0000 | Up | cis |
ENSRNOG00000061656 | SNORD14 | 0.17 | 0.9160 | – |
| Table IV.Differentially expressed lncRNA
mechanisms involved in trans-regulatory elements. |
Table IV.
Differentially expressed lncRNA
mechanisms involved in trans-regulatory elements.
| lncRNA ID | log2FC | Q value |
Up/downregulated | Type | Gene name | log2FC | P-value |
Up/downregulated |
|---|
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ccl9 | 5.19 | 0.0017 | Up |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Homer3 | 1.00 | 0.0611 | – |
|
ENSRNOT00000088402 | +∞ | 0.0471 | Up | Trans | Aurkb | 2.35 | 0.0723 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Galns | 1.03 | 0.0724 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Slc39a1 | 0.92 | 0.0739 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | AC099384.2 | +∞ | 0.1052 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Hsd3b7 | 0.83 | 0.1369 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Klrb1b | 1.98 | 0.1413 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Me2 | 0.63 | 0.1558 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Pnma2 | −0.51 | 0.1564 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Cass4 | 1.24 | 0.1599 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Klhl23 | −0.44 | 0.2192 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Elac1 | −0.49 | 0.2260 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Clcf1 | 1.26 | 0.2519 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Fzd6 | 0.80 | 0.2590 | – |
|
NONRATT009960.2 | 4.06 | 0.0430 | Up | Trans | Lcorl | −0.44 | 0.3199 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Gpr37l1 | 0.40 | 0.3297 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Mmab | −0.35 | 0.3391 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Spon1 | −0.28 | 0.3414 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ifngr2 | 0.44 | 0.3660 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Foxred2 | −0.35 | 0.3732 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Prr22 | −0.67 | 0.3889 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Sppl2a | 0.40 | 0.3894 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Haus5 | 0.55 | 0.4019 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Cpvl | 0.80 | 0.4041 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Fam120b | −0.24 | 0.4061 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Fbxo3 | −0.21 | 0.4206 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ypel4 | −0.24 | 0.4477 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Klhl26 | −0.20 | 0.4480 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Sync | 0.43 | 0.5076 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Lhfpl5 | −0.31 | 0.5231 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Mrpl52 | 0.44 | 0.5306 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Cldn15 | −0.39 | 0.5315 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Mrps35 | −0.18 | 0.5443 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Zfp382 | −0.17 | 0.5640 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ube2k | −0.14 | 0.5697 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Drg1 | −0.12 | 0.5755 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | P2rx5 | −0.36 | 0.5770 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Xpnpep3 | 0.25 | 0.5933 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | RGD1311345 | 0.28 | 0.5950 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Acer2 | 0.32 | 0.6034 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Urgcp | 0.28 | 0.6182 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Megf8 | −0.10 | 0.6223 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Lrtm2 | −0.14 | 0.6301 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | RGD1562299 | 0.26 | 0.6325 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Rpl9 | 0.38 | 0.6347 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Slc25a44 | −0.10 | 0.6348 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ssmem1 | −0.28 | 0.6350 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Stt3a | 0.24 | 0.6468 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Spire1 | −0.09 | 0.6631 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | RGD1561777 | −0.13 | 0.6846 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Luzp1 | −0.09 | 0.6941 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Acot2 | 0.26 | 0.7000 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Fitm2 | −0.07 | 0.7111 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Aox4 | 0.32 | 0.7374 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Golga1 | −0.04 | 0.7775 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Lefty2 | 1.23 | 0.7977 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Zdhhc24 | −0.04 | 0.8075 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Zfp329 | −0.03 | 0.8205 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Tmem101 | −0.03 | 0.8236 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Dhh | 0.28 | 0.8353 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ncr3 | 0.57 | 0.8354 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ppp1r15b | 0.14 | 0.8423 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Zfyve27 | −0.01 | 0.8439 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ints7 | 0.15 | 0.8457 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Dus1l | 0.14 | 0.8545 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ahsa2 | 0.15 | 0.8576 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Aif1l | 0.15 | 0.8579 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Rsg1 | 0.16 | 0.8645 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Fuom | 0.15 | 0.8883 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Cwf19l1 | 0.12 | 0.8894 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Paqr7 | 0.01 | 0.8942 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Coa5 | 0.02 | 0.8985 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Cdc14b | 0.13 | 0.8989 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Crebl2 | −0.01 | 0.9081 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | RGD1564541 | 0.10 | 0.9210 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Slc15a1 | −0.03 | 0.9211 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Ppm1k | 0.03 | 0.9299 | – |
|
NONRATT025479.2 | −4.14 | 0.0000 | Down | Cis and trans | Gdf11 | 0.08 | 0.9402 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Fam20b | 0.04 | 0.9436 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Anapc11 | 0.09 | 0.9536 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Tmem79 | −0.04 | 0.9595 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Cwc25 | 0.06 | 0.9692 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Blvrb | 0.07 | 0.9859 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Stk4 | 0.08 | 0.9880 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Rbm20 | 0.08 | 0.9896 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Tbc1d10b | 0.06 | 0.9927 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Mapkbp1 | 0.07 | 0.9986 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Psma8 | 1.00 | 1.0000 | – |
|
ENSRNOT00000092040 | −3.35 | 0.0204 | Down | Trans | Rbbp8nl | 0.37 | 1.0000 | – |
Validation of expression of DELs by
RT-qPCR
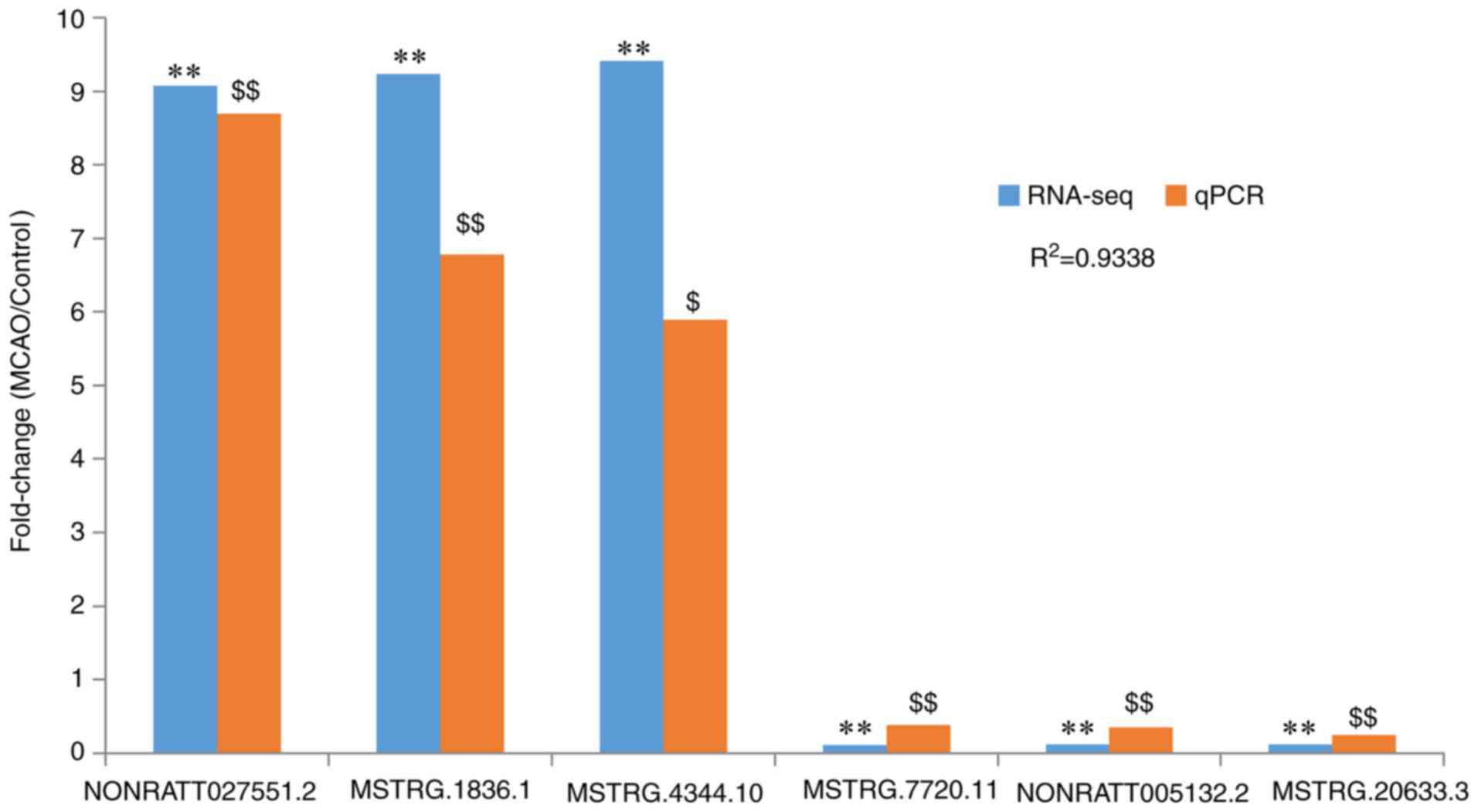
From the data in Fig.
6, NONRATT027551.2, MSTRG.1836.1 and MSTRG.4344.10 were
identified to be significantly upregulated in the MCAO group
compared with the control (P<0.05), consistent with the RNA-seq
data, while MSTRG.7720.11, NONRATT005132.2 and MSTRG.20633.3 were
significantly downregulated in the MCAO group compared with the
control (P<0.01), also consistent with the RNA-seq data. These
results, revealing that the RNA-seq results were consistent with
the RT-qPCR results, verified that the RNA-seq results were
reliable (Fig. 6).
Discussion
A host of lncRNAs have been indicated to be involved
in ischemic stroke by microarray or RNA-seq studies (35,36).
Metastasis associated lung adenocarcinoma transcript 1 was
identified to have a function in ischemic stroke through inhibiting
endothelial cell death and inflammation (36,37).
Additionally, the upregulation of H19 imprinted maternally
expressed transcript may induce apoptosis and necrosis in cerebral
ischemic reperfusion injury (38–40).
In the present study, a total of 77 upregulated and 57
downregulated DELs (|FC|>2, P<0.05) were identified through
reliable RNA-seq and validated using RT-qPCR in an ischemic stroke
group induced by MCAO compared with a control group.
HO-1-mediated neurogenesis has been demonstrated to
be enhanced in ischemic stroke in mice (41). HO-1 has been revealed to promote
angiogenesis following cerebral ischemic reperfusion injury in rats
(42). GO enrichment analysis
suggested that HO-1 was associated with responses to alkaloids,
cellular responses to oxidative stress and responses to reactive
oxygen species. BUB1B has been reported to promote tumor
proliferation in glioblastoma (43,44).
Similarly, BUB1B has been implicated in tumor growth and the
progression of prostate cancer (45), and overexpressed BUB1B has been
demonstrated to be involved in lung adenocarcinoma in humans
(46). The KEGG enrichment
analysis in the present study indicated that BUB1B was associated
with the cell cycle. It has previously been reported that
upregulated CCL2 is associated with protection from stroke induced
by hypoxic preconditioning (47),
and the knockdown of CCL2 was used to successfully reverse the drug
resistance of tumor cells in gastric cancer (48). In the KEGG enrichment analysis
performed in the present study, CCL2 was additionally associated
with the cell cycle. Furthermore, based on the data presented in
Fig. 3, HO-1, BUB1B and CCL2 may
be regulated by a number of novel lncRNAs, including
NONRATT008267.2, NONRATT015286.2, NONRATT004791.2, MSTRG.15067.2,
NONRATT003289.2, NONRATT004566.2, NONRATT005985.2, NONRATT008198.2,
NONRATT028439.2, NONRATT026753.2, NONRATT027268.2, MSTRG.15418.3,
NONRATT016680.2, NONRATT015403.2, MSTRG.29693.5, NONRATT009960.2,
MSTRG.27670.3 and NONRATT000377.2. A previous study has suggested
that the knockdown of DNA topoisomerase IIα (Top2a) may suppress
proliferation and invasion of colon cancer cells (49); based on the present regulatory
analysis of DELs, Top2a, as a cis-regulatory gene of
NONRATT005985.2, may have a vital function in ischemic stroke.
Overall, the analyzed data provide novel DELs and an lncRNA-mRNA
regulatory network that may provide a better understanding of
ischemic stroke induced by MCAO.
Acknowledgements
The authors would like to thank Mr. Qiang Fan (Ao-Ji
Bio-tech Co., Ltd., Shanghai, China) for help with data
analysis.
Funding
The present study was financially supported by the
National Key Research and Development Plan (grant nos.
2017YFC1701600 and 2017YFC1701601), the National Natural Science
Foundation of China (grant nos. 81473387, 81503291 and 81703805),
the Anhui Provincial Natural Science Foundation of China (grant no.
1508085QH191) and the Key Project of the National Science Fund of
Anhui Province (grant no. KJ2013A169).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XD and DP conceived and designed the study. XD and
QB performed the experiments. LH, CP, LX and HP analyzed the data
and drafted the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by The Animal
Experiments Ethics Committee of The Anhui University of Chinese
Medicine (Hefei, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Writing Group Members, Mozaffarian D,
Benjamin EJ, Go AS, Arnett DK, Blaha MJ, Cushman M, Das SR, de
Ferranti S, Després JP, et al: Heart disease and stroke
statistics-2016 update: A report from the american heart
association. Circulation. 133:e38–e360. 2016.PubMed/NCBI
|
|
2
|
Tewari D, Majumdar D, Vallabhaneni S and
Bera AK: Aspirin induces cell death by directly modulating
mitochondrial voltage-dependent anion channel (VDAC). Sci Rep.
7:451842017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mitsios N, Gaffney J, Kumar P, Krupinski
J, Kumar S and Slevin M: Pathophysiology of acute ischaemic stroke:
An analysis of common signalling mechanisms and identification of
new molecular targets. Pathobiology. 73:159–175. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Deb P, Sharma S and Hassan KM:
Pathophysiologic mechanisms of acute ischemic stroke: An overview
with emphasis on therapeutic significance beyond thrombolysis.
Pathophysiology. 17:197–218. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dharap A, Bowen K, Place R, Li LC and
Vemuganti R: Transient focal ischemia induces extensive temporal
changes in rat cerebral microRNAome. J Cereb Blood Flow Metab.
29:675–687. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jeyaseelan K, Lim KY and Armugam A:
MicroRNA expression in the blood and brain of rats subjected to
transient focal ischemia by middle cerebral artery occlusion.
Stroke. 39:959–966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dharap A, Nakka VP and Vemuganti R: Effect
of focal ischemia on long noncoding RNAs. Stroke. 43:2800–2802.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao F, Qu Y, Liu J, Liu H, Zhang L, Feng
Y, Wang H, Gan J, Lu R and Mu D: Microarray profiling and
co-expression network analysis of lncRNAs and mRNAs in neonatal
rats following hypoxic-ischemic brain damage. Sci Rep. 5:138502015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wei N, Xiao L, Xue R, Zhang D, Zhou J, Ren
H, Guo S and Xu J: MicroRNA-9 mediates the cell apoptosis by
targeting Bcl2l11 in ischemic stroke. Mol Neurobiol. 53:6809–6817.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu Q, Deng F, Xing Z, Wu Z, Cen B, Xu S,
Zhao Z, Nepomuceno R, Bhuiyan MI, Sun D, et al: Long non-coding RNA
C2dat1 regulates CaMKIIδ expression to promote neuronal survival
through the NF-κB signaling pathway following cerebral ischemia.
Cell Death Dis. 7:e21732016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qureshi IA and Mehler MF: Emerging roles
of non-coding RNAs in brain evolution, development, plasticity and
disease. Nat Rev Neurosci. 13:528–541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schaukowitch K and Kim TK: Emerging
epigenetic mechanisms of long non-coding RNAs. Neuroscience.
264:25–38. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Briggs JA, Wolvetang EJ, Mattick JS, Rinn
JL and Barry G: Mechanisms of long non-coding RNAs in mammalian
nervous system development, plasticity, disease, and evolution.
Neuron. 88:861–877. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Washietl S, Kellis M and Garber M:
Evolutionary dynamics and tissue specificity of human long
noncoding RNAs in six mammals. Genome Res. 24:616–628. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dharap A, Pokrzywa C and Vemuganti R:
Increased binding of stroke-induced long non-coding RNAs to the
transcriptional corepressors Sin3A and coREST. ASN Neuro.
5:283–289. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mehta SL, Kim T and Vemuganti R: Long
noncoding RNA FosDT promotes ischemic brain injury by interacting
with REST-associated chromatin-modifying proteins. J Neurosci.
35:16443–16449. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Han L, Ji Z, Chen W, Yin D, Xu F, Li S,
Chen F, Zhu G and Peng D: Protective effects of tao-Hong-si-wu
decoction on memory impairment and hippocampal damage in animal
model of vascular dementia. Evid Based Complement Alternat Med.
2015:1958352015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Duan X, Han L, Peng D, Chen W, Peng C,
Xiao L and Bao Q: High throughput mRNA sequencing reveals potential
therapeutic targets of Tao-Hong-Si-Wu decoction in experimental
middle cerebral artery occlusion. Front Pharmacol. 9:15702019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pertea M, Kim D, Pertea GM, Leek JT and
Salzberg SL: Transcript-level expression analysis of RNA-seq
experiments with HISAT, StringTie and Ballgown. Nat Protoc.
11:1650–1667. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pertea M, Pertea GM, Antonescu CM, Chang
TC, Mendell JT and Salzberg SL: StringTie enables improved
reconstruction of a transcriptome from RNA-seq reads. Nat
Biotechnol. 33:290–295. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun L, Zhang Z, Bailey TL, Perkins AC,
Tallack MR, Xu Z and Liu H: Prediction of novel long non-coding
RNAs based on RNA-Seq data of mouse Klf1 knockout study. BMC
Bioinformatics. 13:3312012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kong L, Zhang Y, Ye ZQ, Liu XQ, Zhao SQ,
Wei L and Gao G: CPC: Assess the protein-coding potential of
transcripts using sequence features and support vector machine.
Nucleic Acids Res 35 (Web Server Issue). W345–D349. 2007.
View Article : Google Scholar
|
|
24
|
Sun L, Luo H, Bu D, Zhao G, Yu K, Zhang C,
Liu Y, Chen R and Zhao Y: Utilizing sequence intrinsic composition
to classify protein-coding and long non-coding transcripts. Nucleic
Acids Res. 41:e1662013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zerbino DR, Achuthan P, Akanni W, Amode
MR, Barrell D, Bhai J, Billis K, Cummins C, Gall A, Girón CG, et
al: Ensembl 2018. Nucleic Acids Res 46 D. D754–D761. 2018.
View Article : Google Scholar
|
|
26
|
Knauss JL and Sun T: Regulatory mechanisms
of long noncoding RNAs in vertebrate central nervous system
development and function. Neuroscience. 235:200–214. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nikolayeva O and Robinson MD: edgeR for
differential RNA-seq and ChIP-seq analysis: An application to stem
cell biology. Methods Mol Biol. 1150:45–79. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Altschul SF, Gish W, Miller W, Myers EW
and Lipman DJ: Basic local alignment search tool. J Mol Biol.
215:403–410. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tafer H and Hofacker IL: RNAplex: A fast
tool for RNA-RNA interaction search. Bioinformatics. 24:2657–2663.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gene Ontology Consortium, Blake JA, Dolan
M, Drabkin H, Hill DP, Li N, Sitnikov D, Bridges S, Burgess S, Buza
T, et al: Gene ontology annotations and resources. Nucleic Acids
Res 41 (Database Issue). D530–D535. 2013.
|
|
32
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dykstra-Aiello C, Jickling GC, Ander BP,
Shroff N, Zhan X, Liu D, Hull H, Orantia M, Stamova BS and Sharp
FR: Altered expression of long noncoding RNAs in blood after
ischemic stroke and proximity to putative stroke risk loci. Stroke.
47:2896–2903. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang J, Yuan L, Zhang X, Hamblin MH, Zhu
T, Meng F, Li Y, Chen YE and Yin KJ: Altered long non-coding RNA
transcriptomic profiles in brain microvascular endothelium after
cerebral ischemia. Exp Neurol. 277:162–170. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang X, Tang X, Liu K, Hamblin MH and Yin
KJ: Long noncoding RNA malat1 regulates cerebrovascular pathologies
in ischemic stroke. J Neurosci. 37:1797–1806. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang J, Cao B, Han D, Sun M and Feng J:
Long non-coding RNA H19 induces cerebral ischemia reperfusion
injury via activation of autophagy. Aging Dis. 8:71–84. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tao H, Cao W, Yang JJ, Shi KH, Zhou X, Liu
LP and Li J: Long noncoding RNA H19 controls DUSP5/ERK1/2 axis in
cardiac fibroblast proliferation and fibrosis. Cardiovasc Pathol.
25:381–389. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Puyal J and Clarke PG: Targeting autophagy
to prevent neonatal stroke damage. Autophagy. 5:1060–1061. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nada SE, Tulsulkar J and Shah ZA: Heme
oxygenase 1-mediated neurogenesis is enhanced by Ginkgo
biloba (EGb 761®) after permanent ischemic stroke in
mice. Mol Neurobiol. 49:945–956. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dong B, Zhang Z, Xie K, Yang Y, Shi Y,
Wang C and Yu Y: Hemopexin promotes angiogenesis via up-regulating
HO-1 in rats after cerebral ischemia-reperfusion injury. BMC
Anesthesiol. 18:22018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ma Q, Liu Y, Shang L, Yu J and Qu Q: The
FOXM1/BUB1B signaling pathway is essential for the tumorigenicity
and radioresistance of glioblastoma. Oncol Rep. 38:3367–3375.
2017.PubMed/NCBI
|
|
44
|
Lee E, Pain M, Wang H, Herman JA, Toledo
CM, DeLuca JG, Yong RL, Paddison P and Zhu J: Sensitivity to BUB1B
inhibition defines an alternative classification of glioblastoma.
Cancer Res. 77:5518–5529. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fu X, Chen G, Cai ZD, Wang C, Liu ZZ, Lin
ZY, Wu YD, Liang YX, Han ZD, Liu JC and Zhong WD: Overexpression of
BUB1B contributes to progression of prostate cancer and predicts
poor outcome in patients with prostate cancer. Onco Targets Ther.
9:2211–2220. 2016.PubMed/NCBI
|
|
46
|
Chen H, Lee J, Kljavin NM, Haley B, Daemen
A, Johnson L and Liang Y: Requirement for BUB1B/BUBR1 in tumor
progression of lung adenocarcinoma. Genes Cancer. 6:106–118.
2015.PubMed/NCBI
|
|
47
|
Stowe AM, Wacker BK, Cravens PD, Perfater
JL, Li MK, Hu R, Freie AB, Stüve O and Gidday JM: CCL2 upregulation
triggers hypoxic preconditioning-induced protection from stroke. J
Neuroinflammation. 9:332012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xu W, Wei Q, Han M, Zhou B, Wang H, Zhang
J, Wang Q, Sun J, Feng L, Wang S, et al: CCL2-SQSTM1 positive
feedback loop suppresses autophagy to promote chemoresistance in
gastric cancer. Int J Biol Sci. 14:1054–1066. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang R, Xu J, Zhao J and Bai JH:
Proliferation and invasion of colon cancer cells are suppressed by
knockdown of TOP2A. J Cell Biochem. 119:7256–7263. 2018. View Article : Google Scholar : PubMed/NCBI
|