Introduction
Colorectal cancer (CRC) is the third most commonly
diagnosed type of cancer worldwide and demonstrates a high
incidence and mortality rate (1),
with the number of cases continuing to rise, particularly in Asian
countries (2). Despite a progressive
improvement in the available treatment regimens, prolonging patient
survival following surgery continues to be predominantly hindered
by post-operative recurrence and metastasis. Therefore, the
development of more efficient treatment regimens and greater
understanding of the precise molecular mechanisms of CRC are
required.
MicroRNAs (miRNAs) are conserved short sequences of
non-coding RNA that are 22–24 nucleotides long and appear to be
involved in a broad range of biological functions, such as cell
invasion, metastasis, proliferation, angiogenesis and apoptosis
(3,4).
miRNAs bind to complementary target mRNA sequences and regulate the
expression of target genes at the post-transcriptional level. In
the last decade, an increasing number of studies have identified
miRNAs as critical regulators of carcinogenesis and tumor
progression, affecting tumor growth, progression, metastasis and
drug resistance (5–7). As a result, an increasing number of
studies have demonstrated the potential of miRNAs as novel
therapeutic targets and diagnostic markers (8,9).
Despite a lack of comprehensive understanding of the
mechanisms underlying miRNA dysregulation in cancer, a number of
studies have demonstrated that the silencing of specific miRNAs and
DNA methylation are strongly associated. For example, in cancer
cells, hypermethylated CpG islands are found within the DNA
sequences encoding miR-127 and miR-124a. Furthermore, miR-127
expression was restored in a human bladder cancer cell line
following treatment with chromatin-modifying agents (10,11) and
restoration of miR-124a expression was observed in a CRC cell line
following genetic disruption of DNA methyltransferases (11). Similarly, miR-34b and miR-34c CpG
island hypermethylation was observed in CRC (12,13). Thus,
the silencing of miRNA expression in cancer appears to be largely
mediated by DNA methylation, particularly the methylation of tumor
suppressor genes.
miR-133b is known to be a muscle-specific miRNA and
has been reported to be involved in the development of skeletal
muscle (14). However, previous
studies have supported a broader expression pattern of miR-133b in
a diverse range of tissues. Furthermore, miR-133b has previously
been identified as a tumor suppressor, with miR-133b deregulation
observed in colorectal, head and neck, squamous cell, lung, and
bladder cancers (15–17). miR-133b regulates various target genes
in these cancer types, such as myeloid cell leukemia sequence 1,
B-cell lymphoma-like 2, fascin homolog 1, c-Met and epidermal
growth factor receptor (16,18–21).
Although previous studies have demonstrated that miR-133b is
significantly downregulated in CRC, the epigenetic mechanisms
associated with aberrant miR-133b expression in CRC remain unclear.
Therefore, the present study aimed to determine the epigenetic
mechanisms involved in the regulation of miR-133b silencing in CRC
cells.
Materials and methods
Cell lines and specimens
Five CRC cell lines, consisting of the HT-29,
HCT-116, SW480, SW620 and LoVo cell lines (American Type Culture
Collection, Rockville, MD, USA), and the healthy human colonic
epithelial HCoEpiC cell line (Sciencell Research Laboratories,
Carlsbad, CA, USA) were used in the present study. The cells were
maintained under the recommended conditions.
In total, 6 human CRC, 2 adjacent non-tumor and 2
healthy colorectal tissue samples were obtained in March 2014 from
patients that were diagnosed with colon adenocarcinoma at the
Department of General Surgery, The Third Xiangya Hospital of
Central South University (Changsha, Hunan, China). The specimens
were obtained subsequent to surgical resection and were immediately
frozen at −80°C until use. Written informed consent was obtained
from all patients. The methodology used in the present study
conformed to the standards set by the Declaration of Helsinki
(22) and was approved by the ethics
committee of The Third Xiangya Hospital of Central South
University.
5-aza-2′-deoxycytidine (5-Aza-CdR) and
4-phenylbutyric acid (PBA) treatment
To evaluate the expression of miR-133b, all cell
lines were treated with 3 µM of the demethylation agent 5-Aza-CdR,
3 mM histone deacetylase (HDAC) inhibitor (PBA), or 3 µM 5-Aza-CdR
(Sigma-Aldrich, St. Louis, MO, USA) and 3 mM PBA (Sigma-Aldrich)
for 24, 48 and 72 h. The cells were seeded 24 h prior to
treatment.
RNA extraction and reverse
transcription quantitative-polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the CRC cell lines and
HCoEpiC cells using TRIzol reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA) and then reverse-transcribed into complementary
DNA. HmiRQP0167 primers were obtained from GeneCopoeia, Inc.
(Rockville, MD, USA). The PCR protocol consisted of 35 cycles at
95°C for 3 min, 95°C for 12 sec and 58°C for 30 sec. The relative
expression level of miRNA was normalized to the expression of the
internal control U6 small nuclear RNA using the 2−ΔΔCt
method (23).
Methylation-specific PCR (MSP)
MSP was used to analyze the methylation status of
healthy and CRC tissues, as well as healthy, HT-29 and SW620 cell
lines, as described previously (24).
Genomic DNA was extracted from tissues or cells using the DNA
Extraction Mini Kit (Qiagen GmbH, Hilden, Germany) according to the
manufacturer's instructions. Extracted DNA samples underwent sodium
bisulfite modification using a Universal CpGenome™ DNA Modification
Kit (Merck Millipore, Darmstadt, Germany). MethPrimer software
(available from www.methdb.de) was used to design the
two sets of primers, as follows: methylated miR-133b forward,
3′-TTTATTTAAAATATAAAAATA GCGG-5′ and reverse,
3′-TCACCCAAACTAAAATACAAT AACG-5′; and unmethylated miR-133b
forward, 3′-TTTTTA TTTAAAATATAAAAATAGTGG-5′ and reverse, 3′-ACC
CAAACTAAAATACAATAACACT-5′. MSP was performed using the previously
described negative controls and conditions (25).
Combined bisulfite restriction
analysis (COBRA)
The methylation pattern of miR-133b was analyzed
using COBRA, as previously described (26). Briefly, the process involved sodium
bisulfite treatment, which was followed by PCR, restriction
digestion and quantitation (27). The
following primers were designed using MethPrimer software to
specifically amplify the methylated sequence of miR-133b: Forward,
3′-GTATTT AGTATAAAGAAAGTAA-5′ and reverse, 3′-ACCAATACC
CATAAACAACG-5′.
Cell viability assay
The cells were seeded into 96-well plates at a
density of 1×103 cells/well. Following treatment, cell
viability was evaluated at 0, 24, 48 and 72h using a Cell Counting
Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan), according to
the manufacturer's instructions. Optical density was measured at a
wavelength of 450 nm and all data is based on the results of three
independent experiments with six replicates.
Transwell migration assay
Transwell migration chambers (BioCoat Matrigel
Invasion Chamber; BD Biosciences, Franklin Lakes, NJ, USA) were
used to evaluate cell invasion. A total of 2.5×105
cells/well were starved in serum-free medium (Invitrogen Life
Technologies) for 24 h, then resuspended in serum-free medium. The
cells were added to the upper chamber, while the lower chamber was
filled with basic medium containing 10% fetal bovine serum.
Following incubation for 24 h, the cells that had migrated across
the membrane were fixed with 75% alcohol and stained with crystal
violet (Amresco LLC, Solon, OH, USA) for 20 min. Migrated cells
were then counted using an inverted microscope (TS100; Nikon
Corporation, Tokyo, Japan). All experiments were repeated in
triplicate.
Flow cytometry analysis of the cell
cycle
Flow cytometry was used to analyze cell cycle
distribution, as previously described (27). Briefly, 5-Aza-CdR or PBA-treated cells
were fixed in ice-cold 70% ethanol and stained with propidium
iodide. The cell cycle profiles were assayed using EPICS Elite ESP
flow cytometry (Beckman Coulter, Inc., Brea, CA, USA) at a
wavelength of 488 nm and the data were analyzed using the BD
CellQuest software (BD Biosciences, San Jose, CA, USA).
Statistical analysis
Data in the present study are expressed as the mean
± standard deviation of a minimum of three independent experiments,
unless otherwise stated. Analyses were conducted using SPSS
software version 17.0 (SPSS, Inc., Chicago, IL, USA). An unpaired
Student's t-test was used for comparisons between two
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
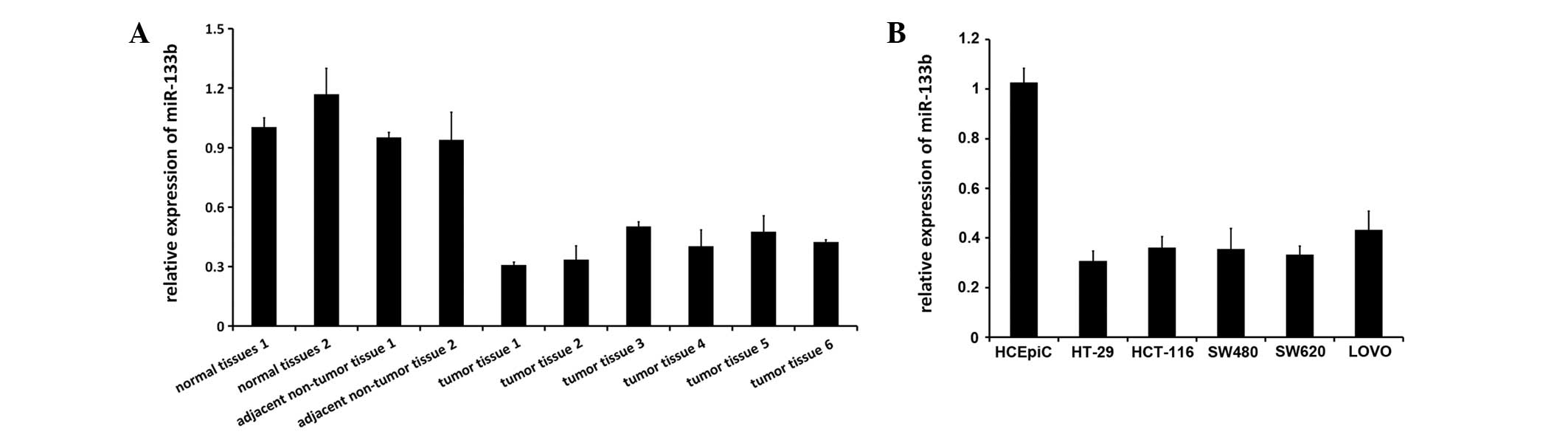
Reduced expression of miR-133b in
CRC
The expression of miR-133b was previously reported
to be decreased in CRC tissues and two colon adenocarcinoma cell
lines (21). Therefore, the present
study used RT-qPCR to determine miR-133b expression levels in
healthy colon tissues, human CRC tissue and adjacent non-tumor
tissue, as well as five CRC cell lines, consisting of the HT-29,
HCT-116, SW480, SW620 and LOVO cell lines, and the healthy HCoEpiC
colon cell line. miR-133b expression was notably downregulated or
silenced in all the investigated CRC tissues and cell lines
(P=0.013), but miR-133b was broadly expressed in all healthy
tissues and HCoEpiC cell lines (Fig. 1A
and B).
Promoter methylation of miR-133b and
restoration of miR-133b expression by demethylation
The present study investigated whether miR-133b
silencing in CRC was caused by CpG methylation in the promoter
region. MSP analysis revealed that miR-133b was frequently
methylated in the investigated CRC tissues and in the SW260 and
HT-29 colon cancer cell lines, and was strongly associated with
expression levels. However, no miR-133b methylation was identified
in the healthy colon tissue samples or healthy colon cell line
(Fig. 2A and B). This finding was
supported by results obtained from the RT-qPCR and COBRA analyses
(Fig. 2C and D), which indicated that
the expression of miR133b was increased in SW620 and HT-29 cells
treated with Aza, PBA and Aza+PBA. CCK-8 was also used to assess
the effects of demethylation agents on SW620 and HT-29 cell
proliferation. It was observed that inhibition of proliferation was
induced by Aza or PBA treatment, whilst a significantly higher
level of inhibition was induced by Aza+PBA treatment (Fig. 2E).
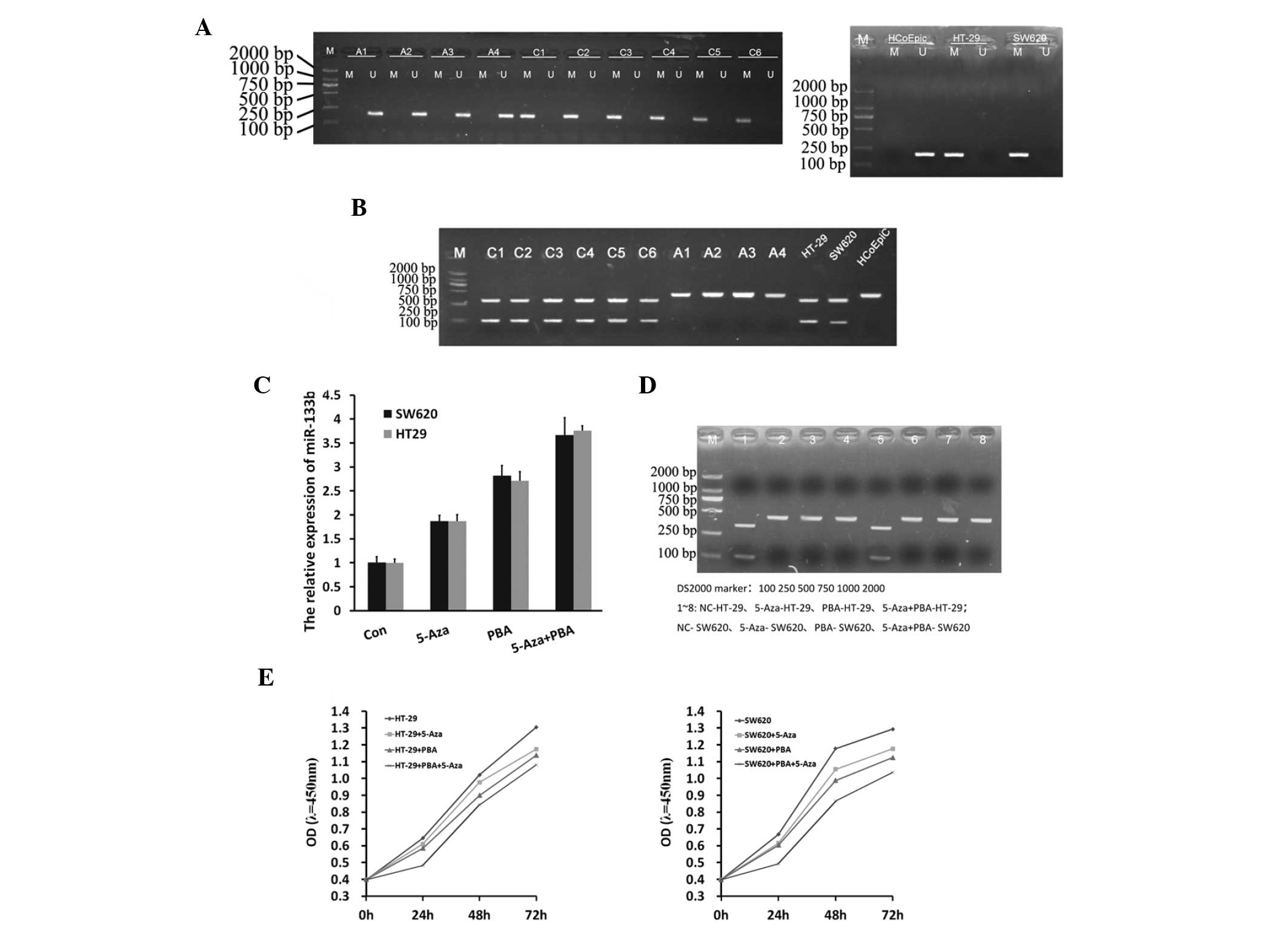 | Figure 2.miR-133b methylation in CRC tissue
samples and cell lines, and reactivation of miR-133b by
demethylation. (A and B) Methylation-specific PCR, revealing
frequent methylation of miR-133b in CRC tissue samples and the CRC
HT-29 and SW60 cell lines, and healthy human colonic HCoEpiC cell
lines. A1-A2, Healthy colon tissue; A3-A4, adjacent non-tumor
tissue; C1-C6, human CRC tissue. (C) Reverse
transcription-quantitative PCR and (D) combined bisulfite
restriction analysis demonstrating that pharmacological
demethylation can restore miR-133b expression in CRC cells. (E)
Survival curve of CRC cells following treatment with 5-Aza/PBA,
determined by using CCK-8. CRC, colorectal cancer; PCR, polymerase
chain reaction; M, methylated; U, unmethylated; miR, microRNA; con,
control; 5-Aza, 5-aza-2′-deoxycytidine; PBA, 4-phenylbutyric acid;
OD, optical density; CCK-8, cell counting kit-8. |
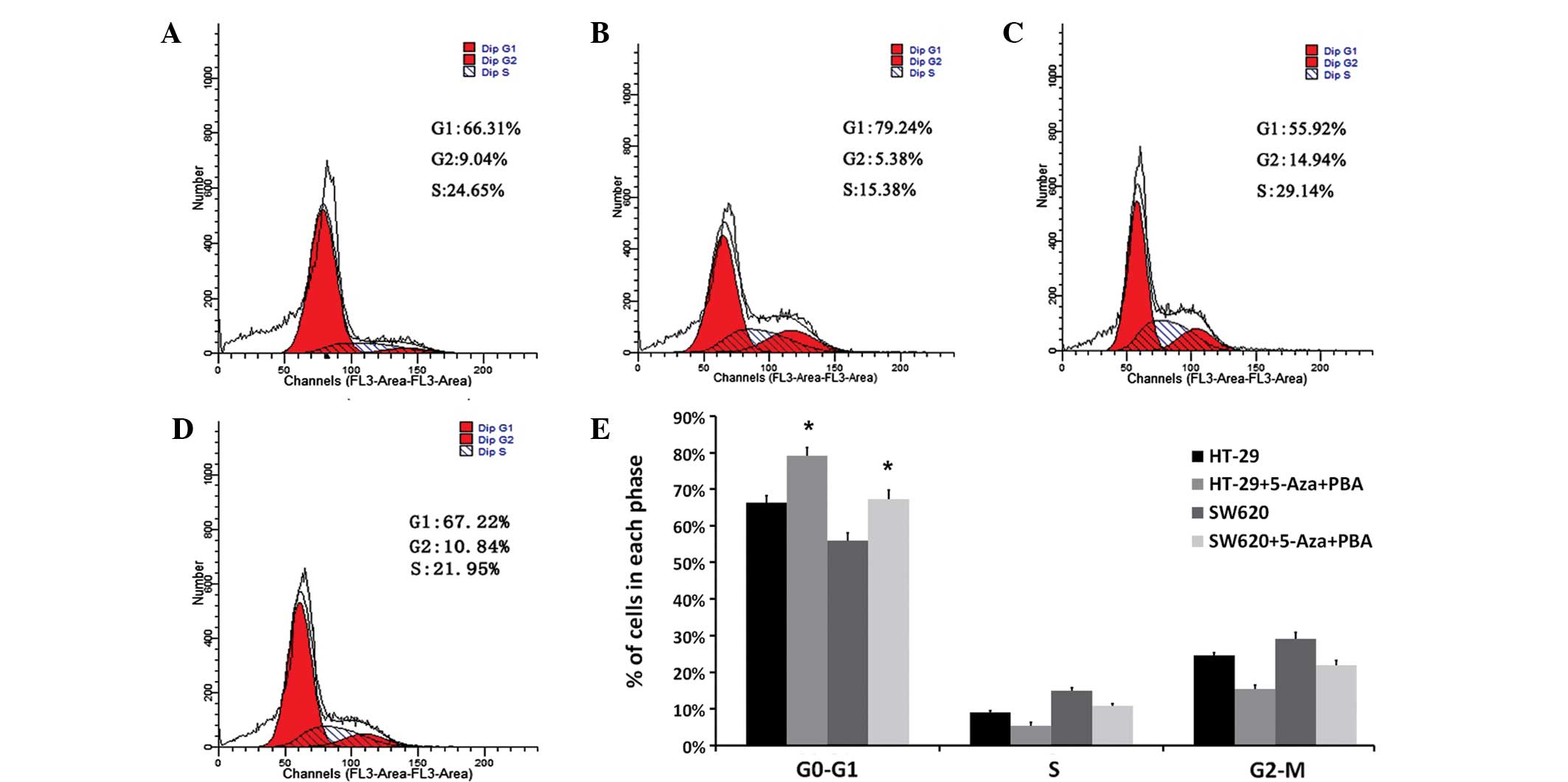
miR-133b demethylation affects cell
cycle arrest and cell invasion in CRC
The present study investigated the effects of
miR-133b expression on the cell cycle and invasive ability of the
HT-29 and SW620 CRC cell lines. Representative results of cell
cycle distribution in control and 5-Aza-CdR/PBA-treated cells are
shown in Fig. 3A–D. Flow cytometry
analysis revealed a statistically significant increase in the
number of 5-Aza-CdR/PBA-treated cells in the G0/G1 phase (13% in
HT-29 cells and 12% in SW620 cells) compared with the number of
control cells in the G0/G1 phase (P<0.05). This was accompanied
by a significant decrease in the number of 5-Aza-CdR/PBA-treated
cells in the S and G2-M phase compared with the control cells
(P=0.027; Fig. 3E).
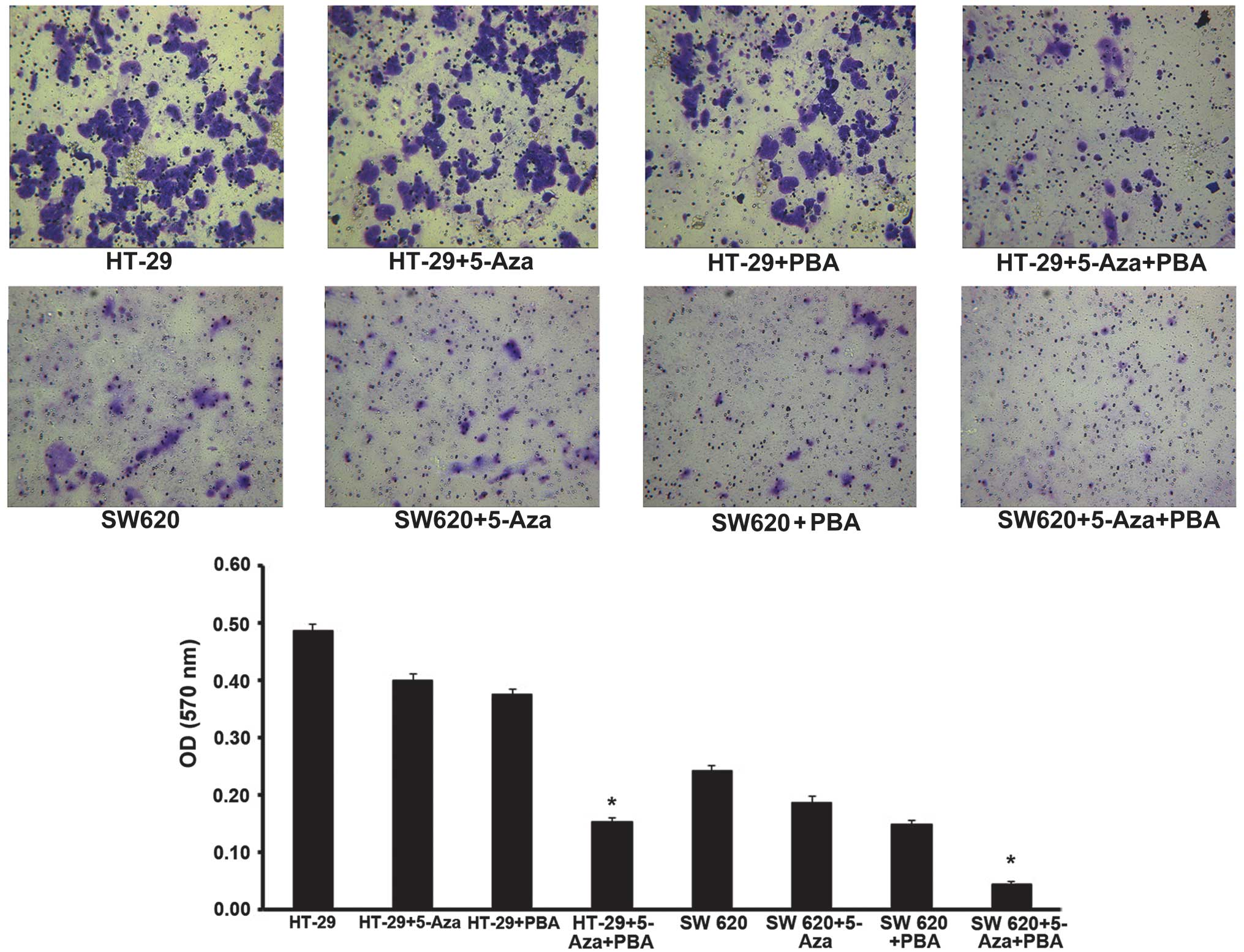
The effect of miR-133b methylation on tumor
metastasis was evaluated by performing a cell invasion assay.
Analysis of the Transwell invasion chamber assay data indicated
that the invasive ability of cells in the miR-133b demethylation
group treated with 5-Aza-CdR/PBA was significantly inhibited
compared with the control group (P<0.05; Fig. 4).
The aforementioned results indicate that miR-133b
methylation reduced CRC cell cycle progression and the invasiveness
of CRC tumor cells.
Discussion
In previous years, an increasing number of studies
have identified the dysregulation of miRNA expression in a broad
range of cancer types (3,5,6,8,11,12). Epigenetic mechanisms were found to be
the key mediators underlying the downregulation of miRNA
expression. For example, miR-126 was downregulated in human cancer
cells and was activated by inhibitors of DNA methylation and
histone deacetylation (28). In
addition, the activation of miR-512-5p by epigenetic treatment
resulted in the apoptosis of gastric cancer cells (29). Similarly, miR-34b and miR-34c are
novel tumor suppressors that are frequently silenced by DNA
methylation in gastric cancer (12).
In addition, the hypermethylation of miRNA genes that is frequently
observed in CRC, as well as the association between the
clinicopathological features and methylation status, provides
evidence for the hypothesis that epigenetic instability is an
important event in tumorigenesis, particularly in CRC. The present
findings revealed miRNA methylation is involved in an epigenetic
field defect and may be a target of epigenetic therapy in
cancer.
Considering the high occurrence, mortality and
relapse rates of CRC, there is an acute requirement for the
identification of novel prognostic factors and development of
therapeutic strategies. miR-133b is known to be a muscle-specific
miRNA (30) and has recently been
reported to act as a tumor suppressor that is dysregulated in
colorectal, head and neck, squamous cell, lung, and bladder cancer
(20,31). Aberrant miR-133b expression has been
identified in CRC cancer tissues (24), with the overexpression of miR-133b in
CRC cells observed to induce apoptosis and G1 phase cell cycle
arrest (20). However, the epigenetic
mechanisms associated with aberrant miR-133b expression remain
unclear.
The present study identified that the expression of
miR-133b was significantly decreased in CRC cells with miR-133b
promoter hypermethylation, indicating that miR-133b was silenced by
epigenetic mechanisms. Furthermore, inhibitors of DNA methylation
and HDAC reduced the extent of DNA methylation and activated the
expression of silenced miR-133b in human CRC cells. In addition,
functional studies have revealed that miR-133b methylation
interfered with CRC cell apoptosis, cell cycle progression and cell
invasion. The results of the present study are consistent with
those of previous studies which revealed that miR-133b inhibits
cell growth and invasion in bladder (32), non-small-cell lung (33) and prostate (34) cancer.
In conclusion, the present study observed a high
frequency of miR-133b hypermethylation in CRC cells and tissue
samples, indicating that methylation of the miR-133b gene may be a
useful diagnostic marker for patients with CRC. However, future
in vivo studies are required to investigate the
tumor-suppressive role of miR-133b in CRC.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sung JJ, Lau JY, Goh KL and Leung WKAsia
Pacific Working Group on Colorectal Cancer: Increasing incidence of
colorectal cancer in Asia: Implications for screening. Lancet
Oncol. 6:871–876. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Manikandan J, Aarthi JJ, Kumar SD and
Pushparaj PN: Oncomirs: The potential role of non-coding microRNAs
in understanding cancer. Bioinformation. 2:330–334. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meltzer PS: Cancer genomics: Small RNAs
with big impacts. Nature. 435:745–746. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fang K, Qian F and Chen YQ: MicroRNAs as
regulators in normal hematopoietic and leukemia stem cells: Current
concepts and clinical implications. Curr Mol Med. 12:536–546. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Saito Y, Liang G, Egger G, et al: Specific
activation of microRNA-127 with downregulation of the
proto-oncogene BCL6 by chromatin-modifying drugs in human cancer
cells. Cancer Cell. 9:435–443. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lujambio A, Ropero S, Ballestar E, et al:
Genetic unmasking of an epigenetically silenced microRNA in human
cancer cells. Cancer Res. 67:1424–1429. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Suzuki H, Yamamoto E, Nojima M, et al:
Methylation-associated silencing of microRNA-34b/c in gastric
cancer and its involvement in an epigenetic field defect.
Carcinogenesis. 31:2066–2073. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Toyota M, Suzuki H, Sasaki Y, et al:
Epigenetic silencing of microRNA-34b/c and B-cell translocation
gene 4 is associated with CpG island methylation in colorectal
cancer. Cancer Res. 68:4123–4132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Koutsoulidou A, Mastroyiannopoulos NP,
Furling D, Uney JB and Phylactou LA: Expression of miR-1, miR-133a,
miR-133b and miR-206 increases during development of human skeletal
muscle. BMC Dev Biol. 11:342011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu J, Yang T, Li X, et al: Alteration of
serum miR-206 and miR-133b is associated with lung carcinogenesis
induced by 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone. Toxicol
Appl Pharmacol. 267:238–246. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamamoto H, Kohashi K, Fujita A and Oda Y:
Fascin-1 overexpression and miR-133b downregulation in the
progression of gastrointestinal stromal tumor. Mod Pathol.
26:563–571. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao H, Li M, Li L, Yang X, Lan G and
Zhang Y: MiR-133b is down-regulated in human osteosarcoma and
inhibits osteosarcoma cells proliferation, migration and invasion
and promotes apoptosis. PLoS One. 8:e835712013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Crawford M, Batte K, Yu L, et al: MicroRNA
133B targets pro-survival molecules MCL-1 and BCL2L2 in lung
cancer. Biochem Biophys Res Commun. 388:483–489. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Patron JP, Fendler A, Bild M, et al:
MiR-133b targets antiapoptotic genes and enhances death
receptor-induced apoptosis. PLoS One. 7:e353452012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu G, Chen D, Li X, Yang K, Wang H and Wu
W: miR-133b regulates the MET proto-oncogene and inhibits the
growth of colorectal cancer cells in vitro and in vivo. Cancer Biol
Ther. 10:190–197. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiang KM and Li XR: MiR-133b acts as a
tumor suppressor and negatively regulates TBPL1 in colorectal
cancer cells. Asian Pac J Cancer Prev. 15:3767–3772. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
General Assembly of the World Medical
Association, . World Medical Association Declaration of Helsinki:
Ethical principles for medical research involving human subjects. J
Am Coll Dent. 81:14–18. 2014.PubMed/NCBI
|
|
23
|
Izutsu N, Maesawa C, Shibazaki M, et al:
Epigenetic modification is involved in aberrant expression of class
III beta-tubulin, TUBB3, in ovarian cancer cells. Int J Oncol.
32:1227–1235. 2008.PubMed/NCBI
|
|
24
|
Bandrés E, Cubedo E, Agirre X, et al:
Identification by Real-time PCR of 13 mature microRNAs
differentially expressed in colorectal cancer and non-tumoral
tissues. Mol Cancer. 5:292006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ogasawara S, Maesawa C, Yamamoto M, et al:
Disruption of cell-type-specific methylation at the Maspin gene
promoter is frequently involved in undifferentiated thyroid
cancers. Oncogene. 23:1117–1124. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Karpinski P, Szmida E, Misiak B, et al:
Assessment of three epigenotypes in colorectal cancer by combined
bisulfite restriction analysis. Mol Carcinog. 51:1003–1008. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xiang T, Li L, Yin X, et al: The ubiquitin
peptidase UCHL1 induces G0/G1 cell cycle arrest and apoptosis
through stabilizing p53 and is frequently silenced in breast
cancer. PLoS One. 7:e297832012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saito Y, Friedman JM, Chihara Y, Egger G,
Chuang JC and Liang G: Epigenetic therapy upregulates the tumor
suppressor microRNA-126 and its host gene EGFL7 in human cancer
cells. Biochem Biophys Res Commun. 379:726–731. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saito Y, Suzuki H, Tsugawa H, et al:
Chromatin remodeling at Alu repeats by epigenetic treatment
activates silenced microRNA-512-5p with downregulation of Mcl-1 in
human gastric cancer cells. Oncogene. 28:2738–2744. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Duan FT, Qian F, Fang K, et al: miR-133b,
a muscle-specific microRNA, is a novel prognostic marker that
participates in the progression of human colorectal cancer via
regulation of CXCR4 expression. Mol Cancer. 12:1642013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wen D, Li S, Ji F, et al: miR-133b acts as
a tumor suppressor and negatively regulates FGFR1 in gastric
cancer. Tumour Biol. 34:793–803. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou Y, Wu D, Tao J, Qu P, Zhou Z and Hou
J: MicroRNA-133 inhibits cell proliferation, migration and invasion
by targeting epidermal growth factor receptor and its downstream
effector proteins in bladder cancer. Scand J Urol. 47:423–432.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu L, Shao X, Gao W, Zhang Z, Liu P, Wang
R, Huang P, Yin Y and Shu Y: MicroRNA-133b inhibits the growth of
non-small-cell lung cancer by targeting the epidermal growth factor
receptor. FEBS J. 279:3800–3812. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tao J, Wu D, Xu B, Qian W, Li P, Lu Q, Yin
C and Zhang W: microRNA-133 inhibits cell proliferation, migration
and invasion in prostate cancer cells by targeting the epidermal
growth factor receptor. Oncol Rep. 27:1967–1975. 2012.PubMed/NCBI
|