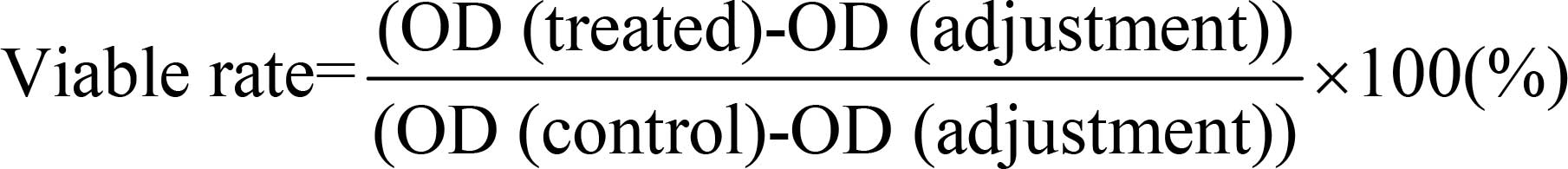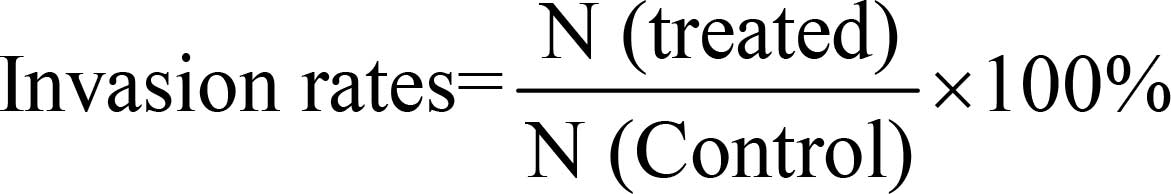Introduction
Osteosarcoma is a bone malignancy that predominantly
affects children and adolescents, and exhibits high invasion and
metastasis rates. It has been reported that the five-year survival
rate of patients who suffer from this disease remains at only 20%
due to a high rate of systemic spread at the early phase and the
strong chemotherapy resistance of osteosarcoma (1). Although adriamycin (ADM) is an effective
benchmark agent for the management of osteosarcoma, it also results
in harmful side-effects including toxicity and chemoresistance that
substantially affect the quality of life of patients (2–4).
Therefore, novel therapeutic approaches and drugs must be sought
for the treatment of osteosarcoma.
Natural products which have potential antitumor
activities have become a focus of attention for study in previous
years (5,6). Cannabinoids, the active components
naturally derived from the marijuana plant Cannabis sativa
L., have been reported as potential antitumor drugs based on their
ability to limit inflammation, cell proliferation and cell survival
(7). To date, several cannabinoids
have been identified and characterized, including
Δ(9)-tetrahydrocannabinol (THC), cannabidiol, cannabinol (CBN) and
anandamide, as well as synthetic cannabinoids, including
WIN-55,212-2, JWH-133 and (R)-methanandamide (7). In the early 1970s, THC and CBN were
shown to inhibit tumor growth in Lewis lung carcinoma (8). Subsequently, cannabinoids were found to
induce apoptosis and inhibit the proliferation of various cancer
cells, including those of glioma (9)
and lymphoma (10), and prostate
(11–13), breast (14), skin (15) and pancreatic cancer (13,16).
Previous studies have speculated about the possible
mechanisms responsible for the physiological and behavioral effects
of cannabinoids. There is a view that cannabinoids exert their
diversified activities via the activation of G-protein-coupled
receptors which are normally bound by endocannabinoids (8). Two types of receptors, CB1 and CB2, have
been cloned and characterized from mammalian tissues. CB1 and CB2
are primarily expressed in the brain and immune system,
respectively (17,18). Another viewpoint states that
cannabinoids suppress tumor cell growth and induce apoptosis by
modulating different cell signaling pathways and the expression of
associated molecules, such as downregulating phosphoinositide
3-kinase (PI3K), protein kinases, extracellular signal-regulated
kinase (ERK) signaling pathways and cell division cycle gene 2
(Cdc2), coupled with an activation of the proapoptotic function of
Bad protein and small guanosine triphosphatase (9,19,20). An effect on the stress-regulated
protein p8 and its downstream targets has also been identified as a
mechanism of the antitumor action of THC (21).
At present, it is believed that the development of
novel combinational therapies may contribute to the enhancement of
the therapeutic effect on tumors. In line with this hypothesis,
trials have been conducted to investigate the synergistic antitumor
effect of multiple drugs. It has been reported that the combined
administration of cannabinoids and temozolomide (TMA) exert a
strong antitumor action in glioblastoma multiforme (22). Combinations of thymoquinone and
diosgenin have also shown potent antiproliferative and apoptotic
effects on squamous cell carcinoma (20). Therefore, the present study aimed to
investigate the antitumor activity of cannabinoid WIN-55,212-2 and
its synergistic effects with ADM against osteosarcoma cells in
vitro. In addition, the present study explores the possible
mechanisms of these antitumor actions, in order to set the basis
for novel strategies for the management of osteosarcoma.
Materials and methods
Cell culture and reagents
The MG-63 human osteosarcoma cell line was obtained
from Nanjing KeyGen Biotechnology, Co., Ltd. (Nanjing, China). The
human umbilical vein endothelial cells (HUVECs) were obtained from
the Institute of Biochemistry and Cell Biology, Shanghai Institutes
for Biological Sciences, Chinese Academy of Sciences (Shanghai,
China). The two cell lines were cultured in RPMI-1640 medium
containing 10% fetal bovine serum (FBS) (Gibco-BRL, Gaithersburg,
MD, USA) at 37°C in an incubator with 5% CO2.
Cannabinoid WIN-55,212-2 was purchased from Sigma-Aldrich (St.
Louis, MO, USA) and ADM was purchased from Pharmacia (Milan,
Italy).
Cell proliferation assay
The cell proliferation assay was performed using a
Cell Counting kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) according to the manufacturer's instructions.
Briefly, exponentially growing MG-63 cells (6×103
cells/well, 100 µl) were seeded into 96-well plates and cultured
overnight using the conditions described above for the cell
culture. Five experimental groups were established: ‘Cannabinoid,’
‘ADM,’ ‘Cannabinoid + ADM,’ ‘Control’ and ‘Blank.’ The adherent
cells in ‘Cannabinoid’ and ‘ADM’ were exposed to 100 µl per well of
20 µM cannabinoid WIN-55,212-2 and 20 µM ADM, respectively,
dissolved in RPMI-1640 fresh medium (Gibco-BRL). Cells in
‘Cannabinoid + ADM’ were exposed to a mixture of the above
cannabinoid and ADM solutions at a ratio of 1:1 of cannabinoid:ADM
(100 µl/well). Fresh medium without drugs (100 µl/well) were added
into ‘Control’ as the control group. The wells for ‘Blank’ which
contained medium without either drugs or cells (100 µl/well) served
as the zero adjustment. The plates were then incubated for 23 h.
CCK-8 (10 µl) was subsequently added to each well and the cells
were further incubated at 37°C for 1 h. Absorbance at 450 nm was
measured using a microplate reader (Infinite® M200 PRO, Tecan GmbH,
Grödig, Austria). The viable rate of cells in each group was
calculated using the following equation:
In which optical density (OD) (control) was obtained
from ‘Control,’ OD (treated) was obtained from the groups with
cells in the presence of drugs, and OD (adjustment) was obtained
from ‘Blank.’ The assay was performed in triplicate.
Scratch assay
The cell migration was assessed by scratch assay.
MG-63 cells were seeded at a density of 5×105 cells per
well in a 6-well plate and grown overnight to confluence on the
plates. A single scrape was made in the confluent monolayer using a
sterile pipette tip. The monolayer was washed with PBS [Sangon
Biotech (Shanghai) Co., Ltd., Shanghai, China] and 2 ml serum-free
medium containing 20 µM cannabinoid WIN-55,212-2, 20 µM ADM or
‘Cannabinoid + ADM’ (Gibco-BRL) was added. Fresh medium without any
drugs was used as the control. Serial photographic images of the
same scraped section were captured at 0, 12, 24 and 36 h using a
Leica TCS 4D microscope (Leica Camera AG, Wetzlar, Germany).
Invasion assay
The invasive capacity of MG-63 cells was measured
using a Transwell® chamber (Corning, NY, USA). All reagents and the
Transwell® chamber were pre-incubated at 37°C. The suspension of
exponentially growing MG-63 cells (2×105 cells/ml) was
placed into the upper compartment in serum-free medium.
Subsequently, 20 µM cannabinoid WIN-55,212-2, 20 µM ADM or
‘Cannabinoid + ADM’ were separately added into the upper
compartments. The complete medium containing 10% FBS was placed
into the lower compartment and served as chemoattractant. After 48
h of incubation at 37°C with 5% CO2, non-invading cells
were removed using cotton swabs. Invading cells were fixed with 4%
paraformaldehyde (Sigma-Aldrich) for 40 min, stained with 0.1%
crystal violet (Sigma-Aldrich) for 30 min and counted in six random
image fields by microscopy (TCS 4D; Leica Camera AG). The invasion
rates were calculated as follows:
In which N (control) was the number of invading
cells in the control group and N (treated) was the number of
invading cells in the groups with cannabinoid and/or ADM
treatment.
Angiogenesis assays in vitro
The angiogenic properties of MG-63 cell-conditioned
medium were assessed using the angiogenesis assay with HUVECs in
vitro. Exponentially growing MG-63 cells (1×105
cells/ml) were seeded in a 24-well plate and incubated with 20 µM
cannabinoid WIN-55,212-2, 20 µM ADM or ‘Cannabinoid + ADM’ for 24 h
at 37°C with 5% CO2. Conditioned medium was obtained by
collecting the supernatant following centrifugation at 1,000 × g
for 10 min. Exponentially growing HUVECs were cultured in the mixed
medium containing equal amounts of conditioned and serum-free
medium, and then seeded into 12-well plates precoated with
Matrigel™ at a density of 5×104 cells/ml. Plates were
incubated at 37°C with 5% CO2 for 8 h. The tube
formation of HUVECs was observed by microscopy (TCS 4D; Leica
Camera AG) and counted in five random image fields. The assay was
performed in triplicate.
Total RNA extraction and reverse
transcription quantitative polymerase chain reaction (RT-qPCR)
Following treatment with 20 µM cannabinoid
(WIN-55,212-2), 20 µM ADM or ‘Cannabinoid + ADM’ and incubation for
24 h at 37°C with 5% CO2, MG-63 cells were harvested.
MG-63 cells without treatment served as the control group. The
total RNA was extracted from each sample of 1×106 cells
using TRIzol® (Invitrogen Life Technologies, Carlsbad, CA, USA)
following the manufacturer's instructions. M-MLV reverse
transcriptase (Takara Bio Inc., Otsu, Japan) and oligo(dT) were
used to synthesize the first-strand cDNA from the total RNA. The
mixture was then incubated at 37°C for 2 h, terminated by heating
at 95°C for 5 min and stored at −20°C. The transcript levels of
Notch-1, matrix metalloproteinase-2 (MMP-2) and vascular
endothelial growth factor (VEGF) were detected by RT-qPCR using a
Light Cycler 2.0 (Roche Diagnostics Deutschland GmbH, Mannheim,
Germany) using SYBR® Green Master Mix (Toyobo, Osaka, Japan). The
RT-PCR program was as follows: 95°C for 10 sec, followed by 40
cycles of 95°C for 10 sec, 56–65°C for 5 sec and 72°C for 10 sec.
The GAPDH gene, which served as the reference gene for the internal
standard, was amplified with forward and reverse primers. The PCR
primers used to detect each gene were synthesized by SBS Genetech
Co., Ltd. (Beijing, China) and the sequences and information are
listed in Table I. The dissociation
curve analysis of amplification products was performed at the end
of each PCR reaction to confirm the specificity of the
amplification. The data were then analyzed by the 2−∆∆Ct
method (23) to obtain the relative
gene expression levels compared with that of the controls.
 | Table I.Primers used in the quantitative
polymerase chain reaction. |
Table I.
Primers used in the quantitative
polymerase chain reaction.
| Target gene | Accession
number | Primer | Sequence | Annealing length
(bp) | Temperature
(°C) |
|---|
| Notch-1 | NM_017617.3 | F |
5′-TCAGCGGGATCCACTGTGAG-3′ | 104 | 65 |
|
|
| R |
5′-ACACAGGCAGGTGAACGAGTTG-3′ |
|
|
| MMP-2 | NM_004530.4 | F |
5′-CAGGAATGAATACTGGATCTACT-3′ | 173 | 65 |
|
|
| R |
5′-GCTCCAGTTAAAGGCGGCATCCAC-3′ |
|
|
| VEGF | NM_001025370.1 | F |
5′-GAGCCTTGCCTTGCTGCTCTAC-3′ | 148 | 50 |
|
|
| R |
5′-CACCAGGGTCTCGATTGGATG-3′ |
|
|
| GAPDH | NM_002046.3 | F |
5′-GCACCGTCAAGGCTGAGAAC-3′ | 138 | 58 |
|
|
| R |
5′-TGGTGAAGACGCCAGTGGA-3′ |
|
|
Western blot analysis
The expression levels of Notch-1, MMP-2 and VEGF
proteins in MG-63 cells were detected by western blot analysis, and
β-actin was used as the loading control. As described for the total
RNA extraction, MG-63 cells with or without drug treatments were
lysed in ice-cold lysing buffer consisting of 50 mM Trizma base (pH
7.4; Sigma-Aldrich), 1% Triton X-100, 150 mM NaCl, 1% sodium
deoxycholate, 0.1% sodium dodecyl sulfate, 2 mM sodium
pyrophosphate, 25 mM β-glycerophosphate, 1 mM EDTA, 0.5 µg/ml
leupeptin and 1 mM sodium vanadate. All reagents were purchased
from Sigma-Aldrich. Cell lysates were centrifuged at 13,000 × g for
5 min at 4°C. The supernatant was collected and the protein
expressions were measured as follows. The whole cell extracts (50
µg/lane) were separated on 12% SDS-PAGE gels and then transferred
to polyvinylidene difluoride membranes. Subsequent to blocking in
Tris-buffered saline with 5% (w/v) non-fat dried milk, the
membranes were probed with a primary mouse monoclonal anti-human
antibodies for Notch-1, MMP-2 or VEGF (Abcam, Cambridge, UK)
diluted in blocking buffer to a concentration of 1:1,000 at 4°C
overnight. Subsequent to washing three times with Tris-buffered
saline with Tween (TBST), membranes were incubated with diluted
secondary HRP-labeled goat anti-mouse IgG antibody (Abcam), at a
concentration of 1:5,000, conjugated with horseradish peroxidase
and detected by enhanced chemiluminescence reagent [Sangon Biotech
(Shanghai) Co., Ltd.]. Photographic images of the bands were taken
and analyzed by National Institutes of Health Image software
(Bethesda, MD, USA).
Statistical analysis
Statistical analysis was performed with a Student's
t-test using SPSS 20.0 (IBM SPSS, Armonk, NY, USA). All data
are presented in the form of the mean ± standard deviation.
Qualitative data were analyzed using the χ2 test.
Differences between the treatment and control groups were
considered significant at P<0.05.
Results
Cannabinoid and ADM inhibit the
proliferation of MG-63 cells
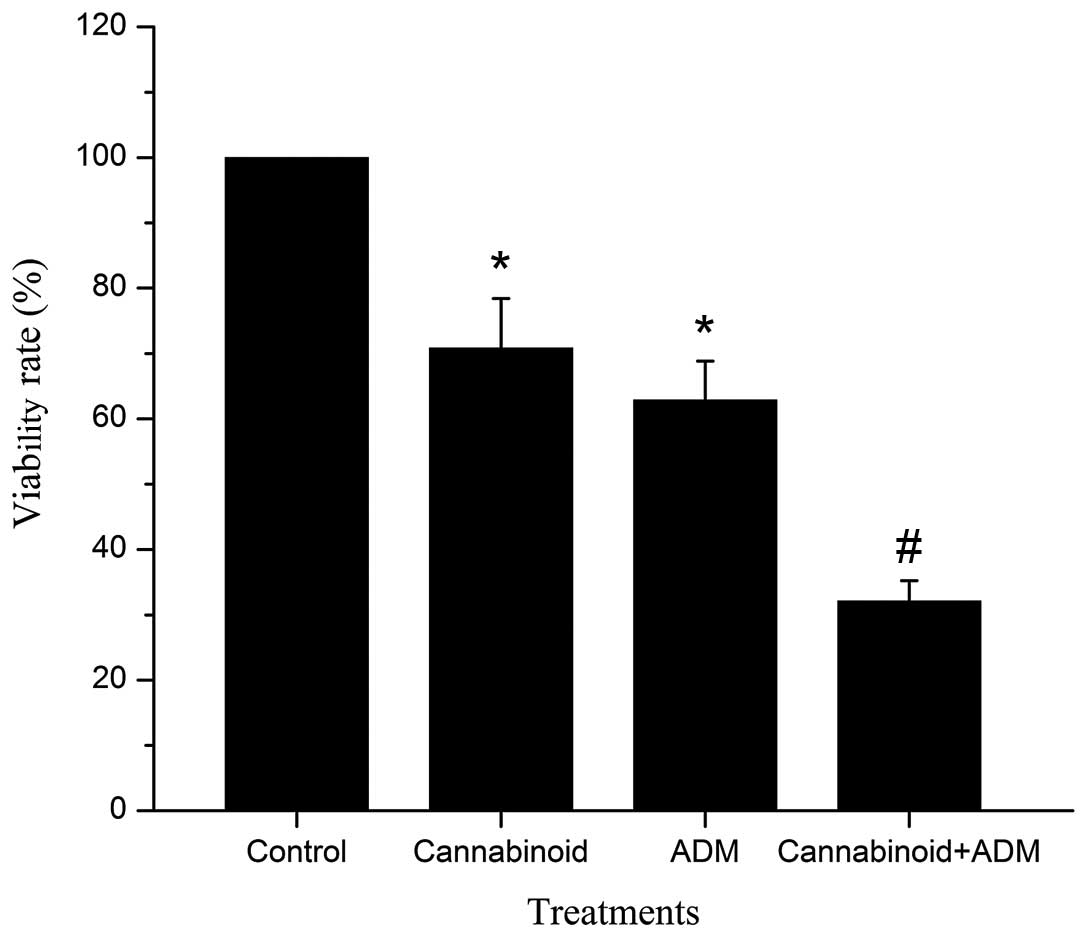
Cannabinoid and ADM were tested individually and as
a combined treatment to ascertain whether these agents inhibited
the proliferation of MG-63 cells. The groups, ‘Cannabinoid,’ ‘ADM,’
and ‘Cannabinoid + ADM,’ in which cells were incubated with 20 µM
cannabinoid (WIN-55,212-2), 20µM ADM and or the two agents for 24
h, were shown to effectively inhibit the growth of MG-63 cells. The
viability rates of cells in the ‘Cannabinoid,’ ‘ADM’ and
‘Cannabinoid + ADM’ treatment groups were 70.86±7.55, 62.87±5.98
and 32.12±3.13%, respectively, which were all significantly reduced
(P<0.05) compared with those in ‘Control’ (100%). Notably, the
viability rate of cells in the ‘Cannabinoid + ADM’ group was
significantly lower (P<0.05) than those in ‘Cannabinoid’ and
‘ADM’ groups (Fig. 1).
Cannabinoid and ADM inhibit the
migration and invasion of MG-63 cells
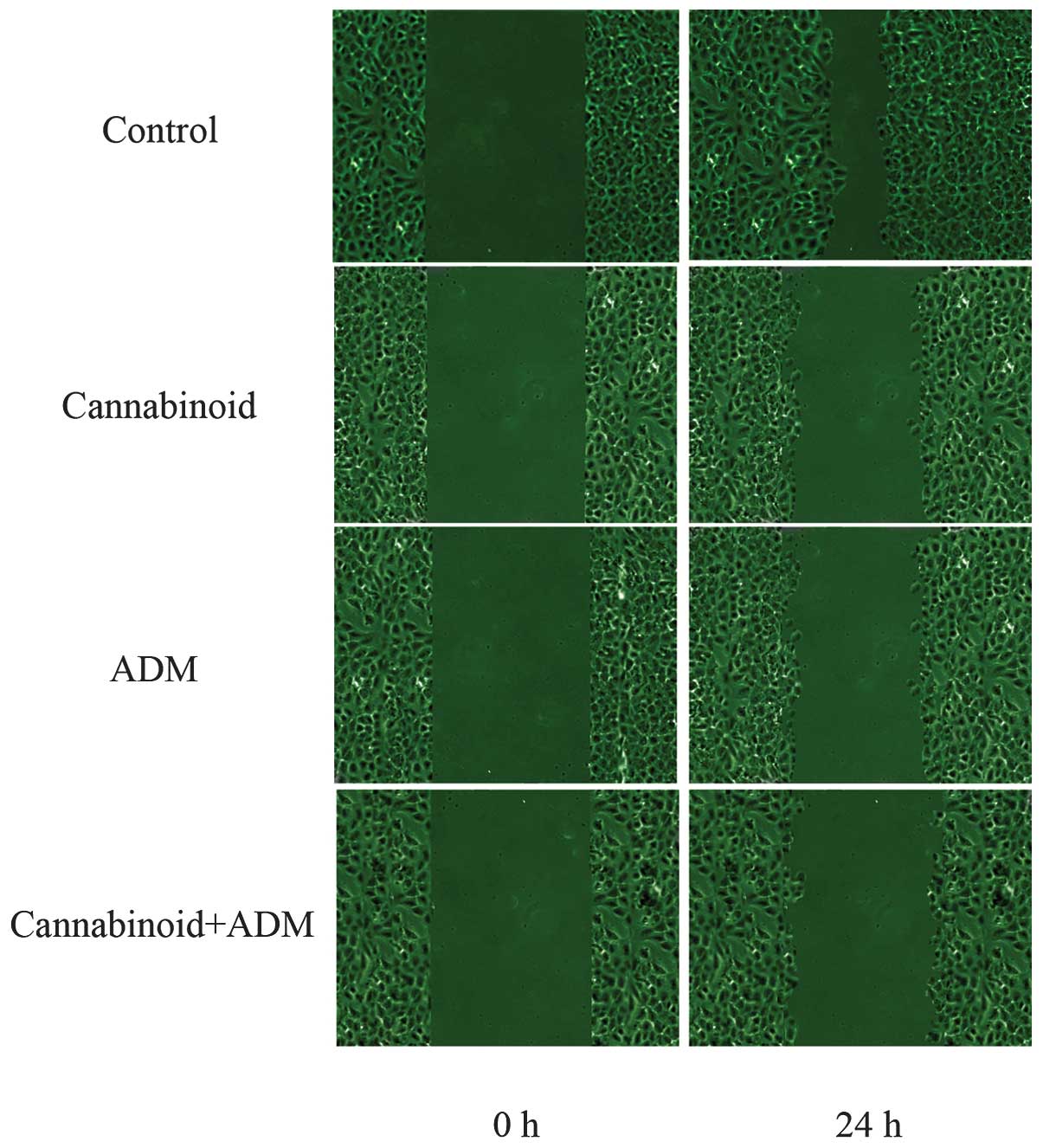
MG-63 cells were treated with 20 µM cannabinoid
(WIN-55,212-2) and ADM alone or combined, and cell migration was
measured by scratch assay. The results demonstrated that the
individual and combined treatments of cannabinoid and ADM had
time-dependent inhibition effects on the migration of cells. This
inhibition was enhanced as time progressed. Compared with the
control treatment, the number of cells that had migrated after 24 h
to heal the wound was fewer in the cells with cannabinoid and/or
ADM treatment, and the scraped sections in these treatment groups
were also wider (Fig. 2). Notably,
these phenomena were observed most markedly in ‘Cannabinoid + ADM’.
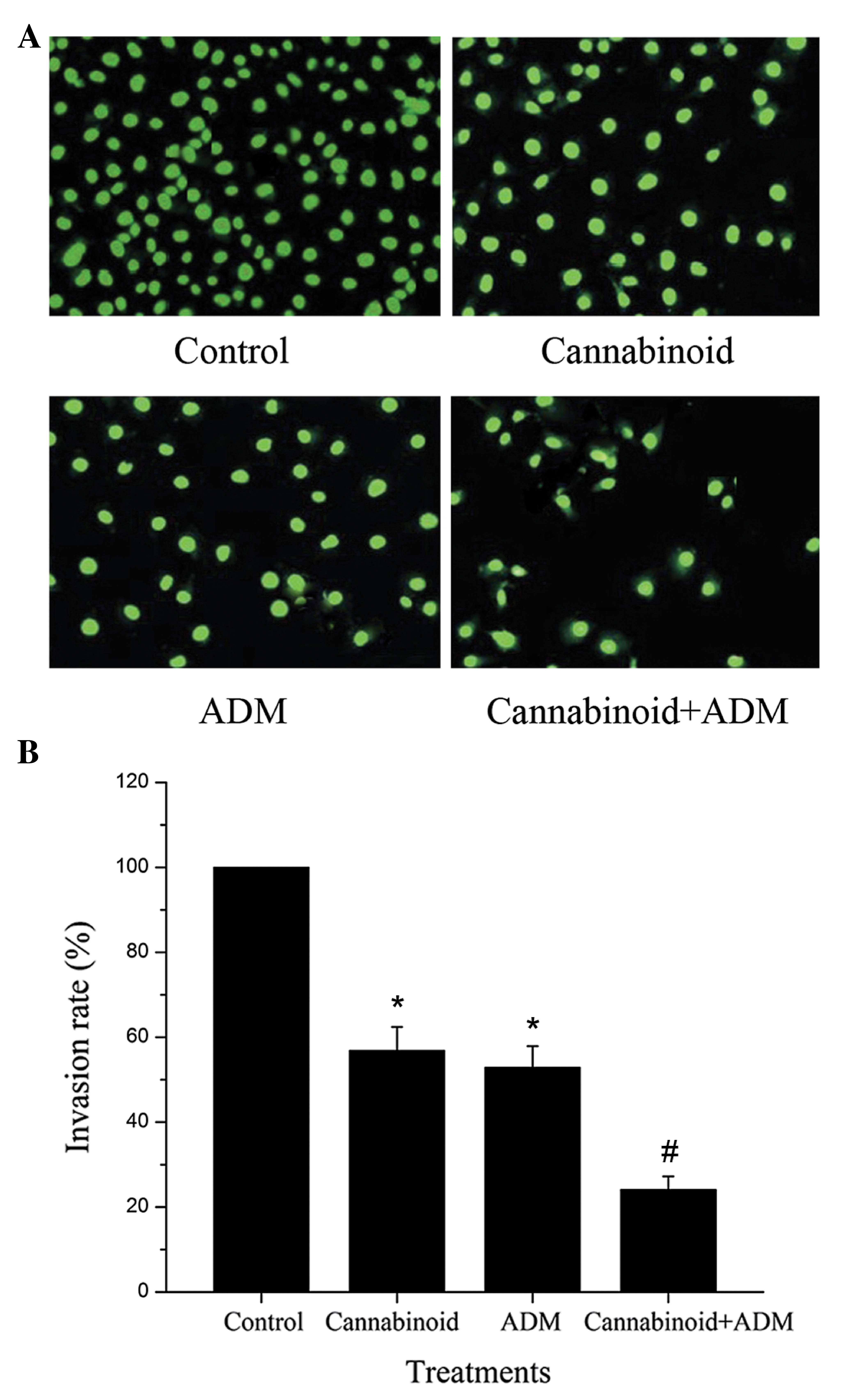
In the cell invasion assay, as shown in Fig. 3A, the number of invading cells in the
treatment groups with cannabinoid and ADM alone or combined was
reduced compared with the control group, and the number of invading
cells in ‘Cannabinoid + ADM’ was the lowest in all the three
treatment groups. Consistently, the invasion rate of cells in
‘Cannabinoid + ADM’ was significantly lower than those in the
groups with the individual treatments of either cannabinoid or ADM
(Fig. 3B).
Cannabinoid and ADM inhibit the
angiogenic activity of HUVECs treated with MG-63 cell-conditioned
medium
The angiogenic properties of MG-63 cell-conditioned
medium were assessed using the angiogenesis assay with HUVECs in
vitro. The results showed that tube formation of HUVECs was
observed with the drug-free MG-63 cell-conditioned medium. However,
tube formation was significantly reduced when the HUVECs were
cultured with the cannabinoid and ADM alone, or the combined
conditioned medium. The combined treatment reduced the angiogenesis
to the greatest extent (Fig. 4).
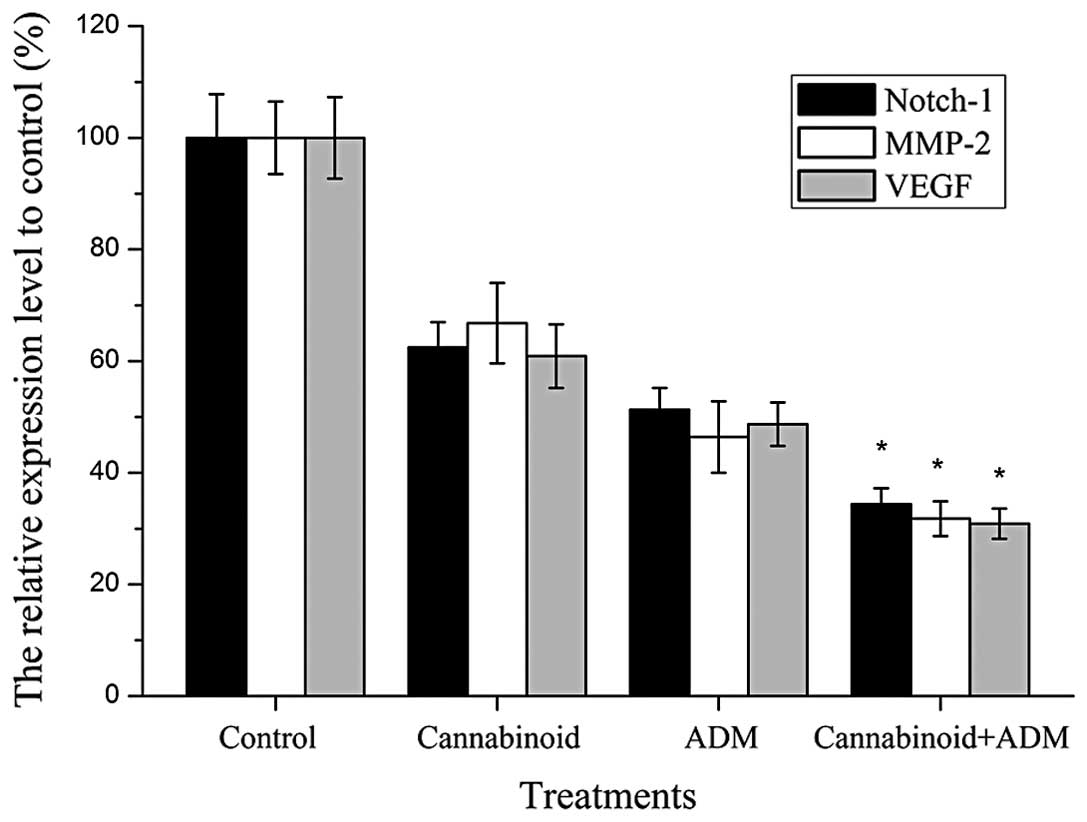
Notch-1, MMP-2 and VEGF expression
levels were downregulated by cannabinoid and ADM
The expression levels of Notch-1, MMP-2 and VEGF
genes were measured by RT-qPCR (Fig.
5). The results showed that the mRNA expression levels of these
genes in MG-63 cells with cannabinoid and/or ADM treatment reduced
significantly (P<0.05) compared with those in the control group
without any drug treatment. Furthermore, the expression levels of
Notch-1, MMP-2 and VEGF in cells with the combined treatment of
cannabinoid and ADM were significantly lower (P<0.05) compared
with cells that received individual treatment of either cannabinoid
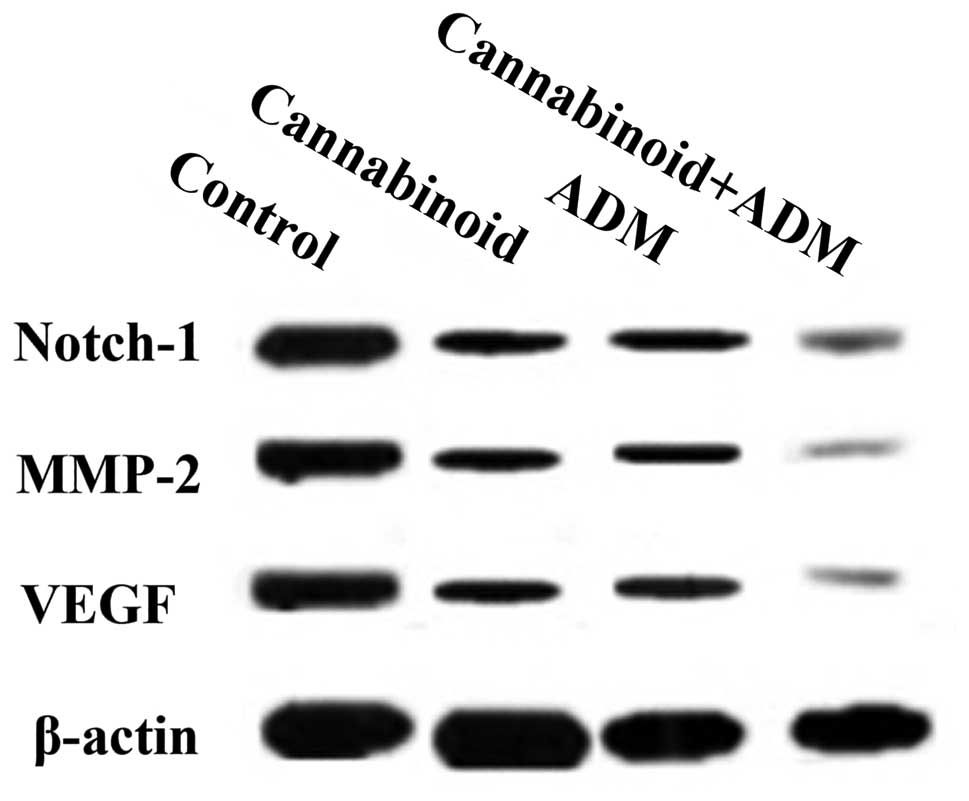
or ADM. Western blotting further confirmed these downregulation
effects (Fig. 6).
Discussion
Despite its potent antitumor activity, ADM, the
conventional medicine against osteosarcoma, has side-effects.
Therefore, there is a requirement to search for novel therapeutic
drugs and approaches aimed at improving the poor prognosis of
patients with osteosarcoma. The results obtained in the present
study demonstrated that WIN-55,212-2, the synthetic cannabinoid
from the main active component of marijuana, reduced the growth of
osteosarcoma MG-63 cells. This is consistent with reports of the
suppressive effects of cannabinoids on lung, prostate, breast, skin
and pancreatic cancer cells (8–16,24). These data together indicate that
cannabinoids may have potential as antitumor drugs for osteosarcoma
therapy. Notably, the present data also demonstrated that the
combined administration of ADM and cannabinoid exerted an enhanced
antiproliferation effect on MG-63 cells. Similar synergistic
effects of drugs were also observed with cannabinoids and TMA
against glioblastoma multiforme (22). Cannabinoids have been shown to be
devoid of the strong side-effects associated with other
chemotherapeutic agents (25,26) and no overt toxic effects of
cannabinoids in patients have been reported in clinical trials for
various applications (8,27). Thus, the synergistic effects of
cannabinoid and ADM in the present study indicated a novel strategy
that may possibly reduce the side-effects and also enhance the
antitumor activity of ADM, while requiring a lower dosage.
Metastasis has been shown to be the main cause of
mortality in patients with osteosarcoma, which must be suppressed
for improved prognosis (28). The
present results demonstrated that cannabinoid (WIN-55,212-2) and/or
ADM could inhibit the migration and invasion of MG-63 cells. This
was consistent with the defined role of ADM in the migration of
osteosarcoma cells (3). Suppression
of metastasis by THC was also reported in severe combined
immunodeficient mice (29). The
results implied that cannabinoid may enhance the antimetastastic
effect of ADM for osteosarcoma, since the combined treatment with
cannabinoid and ADM inhibited the migration and invasion of MG-63
cells to a significantly higher extent than the individual
treatment with either of these two agents.
In terms of the mechanism of the antimetastatic
activities, it was shown that the expression levels of Notch-1 and
MMP-2 were significantly downregulated following the cannabinoid
(WIN-55,212-2) and/or ADM treatments in the present study. Previous
studies have demonstrated that the Notch signaling pathway is
critical in cell proliferation and apoptosis (30,31). Among
the Notch genes, Notch-1 has been reported to be involved in the
migration and invasion of cancer cells (32) and to exhibit crosstalk with nuclear
factor κB (NF-κB), another important regulatory pathway in the
processes of tumor cell invasion and metastasis (33–37). The
MMPs, a family of associated enzymes that degrade the extracellular
matrix, are also considered to be important in facilitating tumor
invasion (38). The level of active
MMP-2 is considered to be a cancer metastasis indicator (39). A previous study also revealed that the
invasion and metastasis of human breast cancer cells is inhibited
by the knockdown of Notch-1, coupled with the inactivation of MMP
expression (32). The present results
showed the downregulation of Notch-1 and MMP-2 in MG-63 cells
following treatments with cannabinoid and ADM alone or in
combination, indicating that cannabinoid and/or ADM may inhibit the
metastasis and invasion of osteosarcoma cells via the suppression
of the Notch signaling pathway and MMPs. Similar observations were
reported in human cervical cancer cells following cannabinoid
treatment, in which the expression level of MMP-2 was downregulated
(40).
In the present study, cannabinoid WIN-55,212-2 was
also shown to suppress the angiogenic activity of MG-63 cells and
to enhance the antiangiogenic effect of ADM as well. Similar
suppressive effects of cannabinoid were also observed against
gliomas in vitro (41) and
human colon carcinoma xenografts in nude mice (42). The present results supported the view
that cannabinoid exhibits highly effective antiangiogenic action
and has a synergetic effect in combination with ADM. Additionally,
previous studies revealed that VEGF is vital for tumor-associated
microvascular invasion as an angiogenic factor (43,44) and
its expression was shown to be associated with the distant
metastasis of tumor cells (45,46). The
present study also showed the inhibition of VEGF expression in
MG-63 cells following cannabinoid and/or ADM treatment. Thus, the
present results indicated that cannabinoid and/or ADM may exert
their antiangiogenic and antimetastatic activities partially
through the downregulation of VEGF expression. Furthermore,
combination therapy ‘Cannabinoid + ADM’ was shown to be more
effective in the downregulation of Notch-1, MMP-2 and VEGF
expression levels compared with treatments of cannabinoid or ADM
alone, which further supports the findings of synergistic antitumor
effects of the two drugs against osteosarcoma.
In conclusion, the present study indicated that
cannabinoid WIN-55,212-2 is antiproliferative, antimetastatic and
antiangiogenic against MG-63 cells in vitro, and presented
evidence that cannabinoid WIN-55,212-2 may result in synergistic
antitumor action in combination with ADM against osteosarcoma. The
present data also indicated that the mechanism of cannabinoid
and/or ADM antitumor activity may be based on the inactivation of
the Notch signaling pathway and the downregulation of MMP-2 and
VEGF, which may be useful for the development and exploration of
potent, nontoxic and novel therapeutic strategies for osteosarcoma.
However, further in-depth studies are required to investigate the
precise and comprehensive molecular mechanisms of these antitumor
activities, and whether this coadministration of cannabinoid and
ADM could be effectively applied in practice.
References
|
1
|
Bielack SS, KempfBielack B, Delling G,
Exner GU, Flege S, Helmke K, Kotz R, SalzerKuntschik M, Werner M,
Winkelmann W, et al: Prognostic factors in high-grade osteosarcoma
of the extremities or trunk: An analysis of 1,702 patients treated
on neoadjuvant cooperative osteosarcoma study group protocols. J
Clin Oncol. 20:776–790. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Friedman MA and Carter SK: The therapy of
osteogenic sarcoma: Current status and thoughts for the future. J
Surg Oncol. 4:482–510. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jaffe N: Recent advances in the
chemotherapy of metastatic osteogenic sarcoma. Cancer.
30:1627–1631. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cortes EP, Holland JF, Wang JJ, Sinks LF,
Blom J, Senn H, Bank A and Glidewell O: Amputation and adriamycin
in primary osteosarcoma. N Engl J Med. 291:998–1000. 1974.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Clark AM: Natural products as a resource
for new drugs. Pharm Res. 13:1133–1144. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Potterat O and Hamburger M: Drug discovery
and development with plant-derived compounds. Prog Drug Res.
65:47–118. 2008.
|
|
7
|
Sarfaraz S, Adhami VM, Syed DN, Afaq F and
Mukhtar H: Cannabinoids for cancer treatment: Progress and promise.
Cancer Res. 68:339–342. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guzmán M: Cannabinoids: Potential
anticancer agents. Nat Rev Cancer. 3:745–755. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
EllertMiklaszewska A, Kaminska B and
Konarska L: Cannabinoids down-regulate PI3K/Akt and Erk signalling
pathways and activate proapoptotic function of Bad protein. Cell
Signal. 17:25–37. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Flygare J, Gustafsson K, Kimby E,
Christensson B and Sander B: Cannabinoid receptor ligands mediate
growth inhibition and cell death in mantle cell lymphoma. Febs
Lett. 579:6885–6889. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ruiz L, Miguel A and Diaz-Laviada I: Delta
(9)-tetrahydrocannabinol induces apoptosis in human prostate PC-3
cells via a receptor-independent mechanism. FEBS Lett. 458:400–404.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nithipatikom K, Endsley MP, Isbell MA,
Falck JR, Iwamoto Y, Hillard CJ and Campbell WB:
2-arachidonoylglycerol: A novel inhibitor of androgen-independent
prostate cancer cell invasion. Cancer Res. 64:8826–8830. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Carracedo A, Gironella M, Lorente M,
Garcia S, Guzmán M, Velasco G and Iovanna JL: Cannabinoids induce
apoptosis of pancreatic tumor cells via endoplasmic reticulum
stress-related genes. Cancer Res. 66:6748–6755. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ligresti A, Moriello AS, Starowicz K,
Matias I, Pisanti S, De Petrocellis L, Laezza C, Portella G,
Bifulco M and Di Marzo V: Antitumor activity of plant cannabinoids
with emphasis on the effect of cannabidiol on human breast
carcinoma. J Pharmacol Exp Ther. 318:1375–1387. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Blázquez C, Carracedo A, Barrado L, Real
PJ, Fernández-Luna JL, Velasco G, Malumbres M and Guzmán M:
Cannabinoid receptors as novel targets for the treatment of
melanoma. Faseb J. 20:2633–2635. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fogli S, Nieri P, Chicca A, Adinolfi B,
Mariotti V, Iacopetti P, Breschi MC and Pellegrini S: Cannabinoid
derivatives induce cell death in pancreatic MIA PaCa-2 cells via a
receptor-independent mechanism. Febs Lett. 580:1733–1739. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Howlett AC, Barth F, Bonner TI, Cabral G,
Casellas P, Devane WA, Felder CC, Herkenham M, Mackie K, Martin BR,
et al: International union of pharmacology. XXVII. Classification
of cannabinoid receptors. Pharmacol Rev. 54:161–202. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fernández-Ruiz J, Romero J, Velasco G,
Tolón RM, Ramos JA and Guzmán M: Cannabinoid CB2 receptor: A new
target for controlling neural cell survival? Trends Pharmacol Sci.
28:39–45. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Caffarel MM, Sarrió D, Palacios J, Guzman
M and Sánchez C: Delta9-tetrahydrocannabinol inhibits cell cycle
progression in human breast cancer cells through Cdc2 regulation.
Cancer Res. 66:6615–6621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Takeda S, Okajima S, Miyoshi H, Yoshida K,
Okamoto Y, Okada T, Amamoto T, Watanabe K, Omiecinski CJ and
Aramaki H: Cannabidiolic acid, a major cannabinoid in fiber-type
cannabis, is an inhibitor of MDA-MB-231 breast cancer cell
migration. Toxicol Lett. 214:314–319. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Carracedo A, Lorente M, Egia A, Blázquez
C, García S, Giroux V, Malicet C, Villuendas R, Gironella M,
González-Feria L, et al: The stress-regulated protein p8 mediates
cannabinoid-induced apoptosis of tumor cells. Cancer Cell.
9:301–312. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Torres S, Lorente M, Rodríguez-Fornés F,
Hernández-Tiedra S, Salazar M, García-Taboada E, Barcia J, Guzmán M
and Velasco G: A combined preclinical therapy of cannabinoids and
temozolomide against glioma. Mol Cancer Ther. 10:90–103. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C (T) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sarfaraz S, Afaq F, Adhami VM, Malik A and
Mukhtar H: Cannabinoid receptor agonist-induced apoptosis of human
prostate cancer cells LNCaP proceeds through sustained activation
of ERK1/2 leading to G1 cell cycle arrest. J Biol Chem.
281:39480–39491. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mackie K: Cannabinoid receptors as
therapeutic targets. Annu. Rev Pharmacol Toxicol. 46:101–122. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pertwee RG: Emerging strategies for
exploiting cannabinoid receptor agonists as medicines. Br J
Pharmacol. 156:397–411. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hall W, Christie M and Currow D:
Cannabinoids and cancer: Causation, remediation and palliation. The
lancet oncology. 6:35–42. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang YH, Han XD, Qiu Y, Xiong J, Yu Y,
Wang B, Zhu ZZ, Qian BP, Chen YX, Wang SF, et al: Increased
expression of insulin-like growth factor-1 receptor is correlated
with tumor metastasis and prognosis in patients with osteosarcoma.
J Surg Oncol. 105:235–243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Preet A, Ganju R and Groopman J:
Delta9-Tetrahydrocannabinol inhibits epithelial growth
factor-induced lung cancer cell migration in vitro as well as its
growth and metastasis in vivo. Oncogene. 27:339–346. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Berx G, Nollet F and Van Roy F:
Dysregulation of the E-cadherin/catenin complex by irreversible
mutations in human carcinomas. Cell Adhes Commun. 6:171–184. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
ArtavanisTsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang J, Fu L, Gu F and Ma Y: Notch1 is
involved in migration and invasion of human breast cancer cells.
Oncol Rep. 26:1295–1303. 2011.PubMed/NCBI
|
|
33
|
Curran S and Murray G: Matrix
metalloproteinases: Molecular aspects of their roles in tumour
invasion and metastasis. Eur J Cancer. 36:1621–1630. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
John A and Tuszynski G: The role of matrix
metalloproteinases in tumor angiogenesis and tumor metastasis.
Pathology oncology research. 7:14–23. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nagakawa Y, Aoki T, Kasuya K, Tsuchida A
and Koyanagi Y: Histologic features of venous invasion, expression
of vascular endothelial growth factor and matrix
metalloproteinase-2 and matrix metalloproteinase-9 and the relation
with liver metastasis in pancreatic cancer. Pancreas. 24:169–178.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xiong HQ, Abbruzzese JL, Lin E, Wang L,
Zheng L and Xie K: NF-kappaaB activity blockade impairs the
angiogenic potential of human pancreatic cancer cells. Int J
Cancer. 108:181–188. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dong Z, Bonfil RD, Chinni S, Deng X,
Trindade Filho JC, Bernardo M, Vaishampayan U, Che M, Sloane BF,
Sheng S, et al: Matrix metalloproteinase activity and osteoclasts
in experimental prostate cancer bone metastasis tissue. Am J
Pathol. 166:1173–1186. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang Z, Banerjee S, Li Y, Rahman KM, Zhang
Y and Sarkar FH: Down-regulation of notch-1 inhibits invasion by
inactivation of nuclear factor-kappaB, vascular endothelial growth
factor and matrix metalloproteinase-9 in pancreatic cancer cells.
Cancer Res. 66:2778–2784. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jezierska A and Motyl T: Matrix
metalloproteinase-2 involvement in breast cancer progression: A
mini-review. Med Sci Monit. 15:RA32–RA40. 2009.PubMed/NCBI
|
|
40
|
Ramer R and Hinz B: Inhibition of cancer
cell invasion by cannabinoids via increased expression of tissue
inhibitor of matrix metalloproteinases-1. J Natl Cancer Inst.
100:59–69. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
MachadoRocha FC, Dos Dos Santos Júnior JG,
Stefano SC and da Silveira DX: Systematic review of the literature
on clinical and experimental trials on the antitumor effects of
cannabinoids in gliomas. J Neurooncol. 116:11–24. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kogan NM, Blázquez C, Alvarez L, Gallily
R, Schlesinger M, Guzmán M and Mechoulam R: A cannabinoid quinone
inhibits angiogenesis by targeting vascular endothelial cells.
Molecular Pharmacology. 70:51–59. 2006.PubMed/NCBI
|
|
43
|
Joo YE, Sohn YH, Lee WS, Park CH, Choi SK,
Rew JS, Park CS and Kim SJ: Expression of vascular endothelial
growth factor and p53 in pancreatic carcinomas. Korean J Intern
Med. 17:153–159. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zeng H, Datta K, Neid M, Li J, Parangi S
and Mukhopadhyay D: Requirement of different signaling pathways
mediated by insulin-like growth factor-I receptor for
proliferation, invasion and VPF/VEGF expression in a pancreatic
carcinoma cell line. Biochem Biophys Res Commun. 302:46–55. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fujioka S, Sclabas GM, Schmidt C, Niu J,
Frederick WA, Dong QG, Abbruzzese JL, Evans DB, Baker C and Chiao
PJ: Inhibition of constitutive NF-kappa B activity by I kappa B
alpha M suppresses tumorigenesis. Oncogene. 22:1365–1370. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wey JS, Fan F, Gray MJ, Bauer TW, McCarty
MF, Somcio R, Liu W, Evans DB, Wu Y, Hicklin DJ, et al: Vascular
endothelial growth factor receptor-1 promotes migration and
invasion in pancreatic carcinoma cell lines. Cancer. 104:427–438.
2005. View Article : Google Scholar : PubMed/NCBI
|