Introduction
Laryngeal cancer is one of the most common types of
head and neck cancer, and the majority of laryngeal cancers are
squamous cell carcinomas (1). The
incidence rate of laryngeal cancer accounts for 1–5% of all
malignancies, and is third most common head and neck cancer tumor
(2). Laryngeal cancer has enormous
functional and psychological consequences for the patients, in
particular with regard to eating and communication (3). Laryngeal papillomas which are caused
primarily by human papillomavirus (HPV) types 6 and 11 (4) are the most common benign tumors in the
larynx. They are associated with a 3–7% risk of malignant
transformation, in which irradiation and smoking appear to be risk
factors (5). In order to further
improve survival rates, diagnostic and preventive approaches, it is
necessary to understand the mechanisms of laryngeal
carcinogenesis.
The past decade has witnessed major progress in the
understanding of the molecular alteration underlying the
development of head and neck cancers (6,7). For
laryngeal cancer, previous studies have revealed that the use of
DNA microarray is helpful for the elucidation of laryngeal
carcinogenesis (8,9). Colombo et al (10) identified that the differentially
expressed genes (DEGs) in larynx tumors were involved in cellular
processes associated with the cancer phenotype, including the cell
cycle, DNA repair, and signal transduction among which cystatin B
(CSTB) encoded the protein, which has previously been
associated with antimetastatic function. Ma et al (6)reported that numerous molecular
abnormalities encoding for cell cycle control and integrin-mediated
cell adhesion, such as matrix metallopeptidase 12 (MMP12),
keratin 19 (KRT19) and proline-rich protein BstNI subfamily
1 (PRB1) were identified in laryngeal carcinoma cells. In
addition, the findings of Oblak et al (11) demonstrated that overexpression of
suppressor of cytokine signaling 1 (SOCS1) was linked with
reducing the aggressive and metastatic potential in laryngeal
carcinoma due to the inhibition of cancer cell proliferation.
Despite the fact that considerable efforts have been made over the
years, the molecular mechanisms involved in the development and
progression of laryngeal carcinoma have not been completely
demonstrated and require further investigation.
Using the same gene expression profile, DeVoti et
al (12) had demonstrated that
altered expression of numerous genes was associated with cellular
growth and differentiation and that there was a role for altered
innate immunity in recurrent respiratory papillomatosis. In the
present study, microarray analysis was used to identify DEGs in
growing papillomas compared with adjacent laryngeal epithelia as
controls. Comprehensive bioinformatics was used to analyze the
significant pathways and functions and to construct the
protein-protein interaction (PPI) network in addition to
significant modules to identify the critical DEGs. Furthermore,
significant biological pathway regions (sub-pathways) were
predicted. The present study aimed to identify involvement of genes
critical in laryngeal carcinoma and to get an improved
understanding of the molecular circuitry in laryngeal
carcinoma.
Materials and methods
Microarray data and data
preprocessing
The mRNA expression profile of GSE10935 was
downloaded from the National Center of Biotechnology Information
(NCBI) Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo/), which was
deposited by DeVoti et al (12). The platform is GPL96 [HG-U133A]
Affymetrix Human Genome U133A Array and GPL571 [HG-U133A_2]
Affymetrix Human Genome U133A 2.0 Array. In this dataset, there
were 12 specimens from papillomas and 12 specimens from adjacent
clinically normal, laryngeal epithelia from patients with recurrent
respiratory papillomatosis.
The probes were annotated with gene symbols
according to the annotation information in different platforms.
Gene symbols which were common across the GPL96 and GPL571
platforms were retained to the subsequent analysis. Furthermore,
cross slide normalization was performed by using the surrogate
variable analysis (SVA) package (13)
in R (14), and within slide
normalization was performed by using preprocessCore package
(15) in R. The gene expression
matrix of specimens was received.
Screening of DEGs
A t-test (16) in the
limma (17) package in R (14) was used to identify DEGs. Up-regulated
and down-regulated genes were identified between papillomas and
normal controls. The Bonferroni method (18) was applied to perform a multiple
testing correction. The threshold for the DEGs was set as corrected
P-value <0.05 and | log2 fold change (FC) | ≥1. In
addition, the clustering analysis of DEGs was represented by a
heat-map using gplots (19) in R
(14).
Functional and pathway enrichment
analysis of DEGs
In order to facilitate the functional annotation and
pathway analysis, all the DEGs were analyzed using the Database for
Annotation Visualization and Integrated Discovery (DAVID) (20) to perform the Gene Ontology (GO)
(21) and Kyoto Encyclopedia of Genes
and Genomes (KEGG) (22) analysis.
The human genome was used as the background list and human was
chosen as the species. The other default parameters of the tools
were kept unchanged. The GO terms and KEGG pathways that were
enriched by >5 genes and P<0.05 was considered to indicate a
statistically significant difference.
PPI network construction and modules
selection
The PPI network is represented by an undirected
graph with nodes indicating the genes and edges indicating the
mapped interactions of the proteins encoded by the genes (23). In the present study, all DEGs were
imported into Cytoscape plugin to create network visualizations.
The source of the interaction network databases were the Human
Protein Reference Database (HPRD) (24) (http://www.hprd.org/) and the Biological General
Repository for Interaction Datasets (BioGRID) (25) (http://thebiogrid.org/) database. Then the resulting
PPI network was subjected to module analysis with the Plugin MCODE
(26) with the default parameters
(Degree cutoff ≥2, Node score cutoff ≥2, K-core ≥2, and Max
depth=100).
Risk sub-pathway analysis
iSubpathwayMiner (http://cran.r-project.org/web/packages/iSubpathwayMiner/)
is an R platform for graph-based construction and analysis of KEGG
pathways (27). For a given KEGG
pathway, the sub-pathways were obtained by searching all possible
paths between start-points (membrane receptors or their ligands)
and end-points (transcriptional factors or their immediate targets)
in the adjacency matrix generated by node relationships (28).
Disease associated risk sub-pathway creation and
presentation were based on iSubpathway Miner (28). The genes which had higher degrees in
the PPI network enriched in the sub-pathways were identified as the
critical genes in papilloma.
Results
Data preprocessing and DEGs
screening
After normalization, 22,283 probe sets mapped to
12,496 gene symbols under GPL96 platform. Total 22,277 probe sets
mapped to 12,495 gene symbols under GPL571 platform. Finally,
12,495 common gene symbols were identified and the gene expression
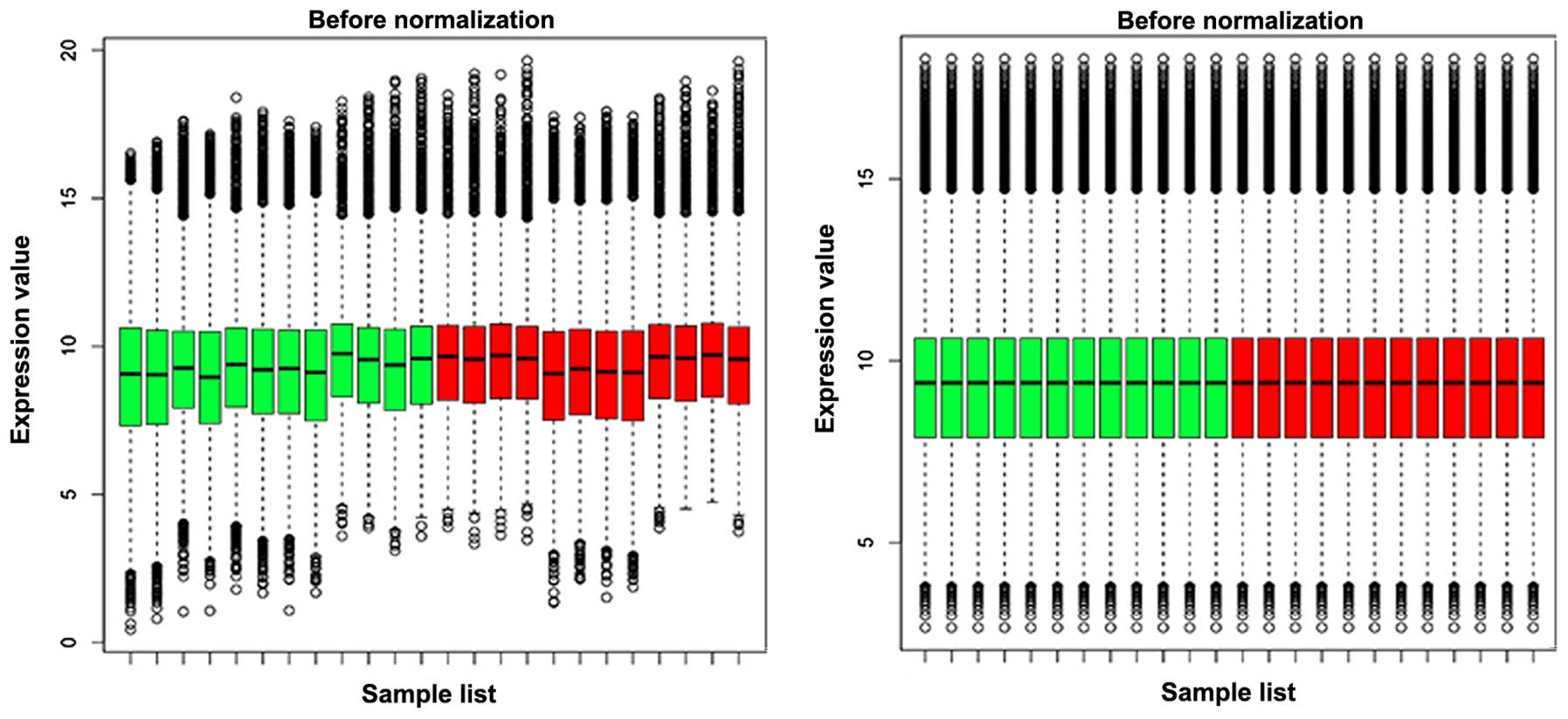
matrix of specimens was received. The gene expression profile
following normalization is presented in Fig. 1. The black line in the box tends
towards the same straight line, indicating a good degree of
standardization.
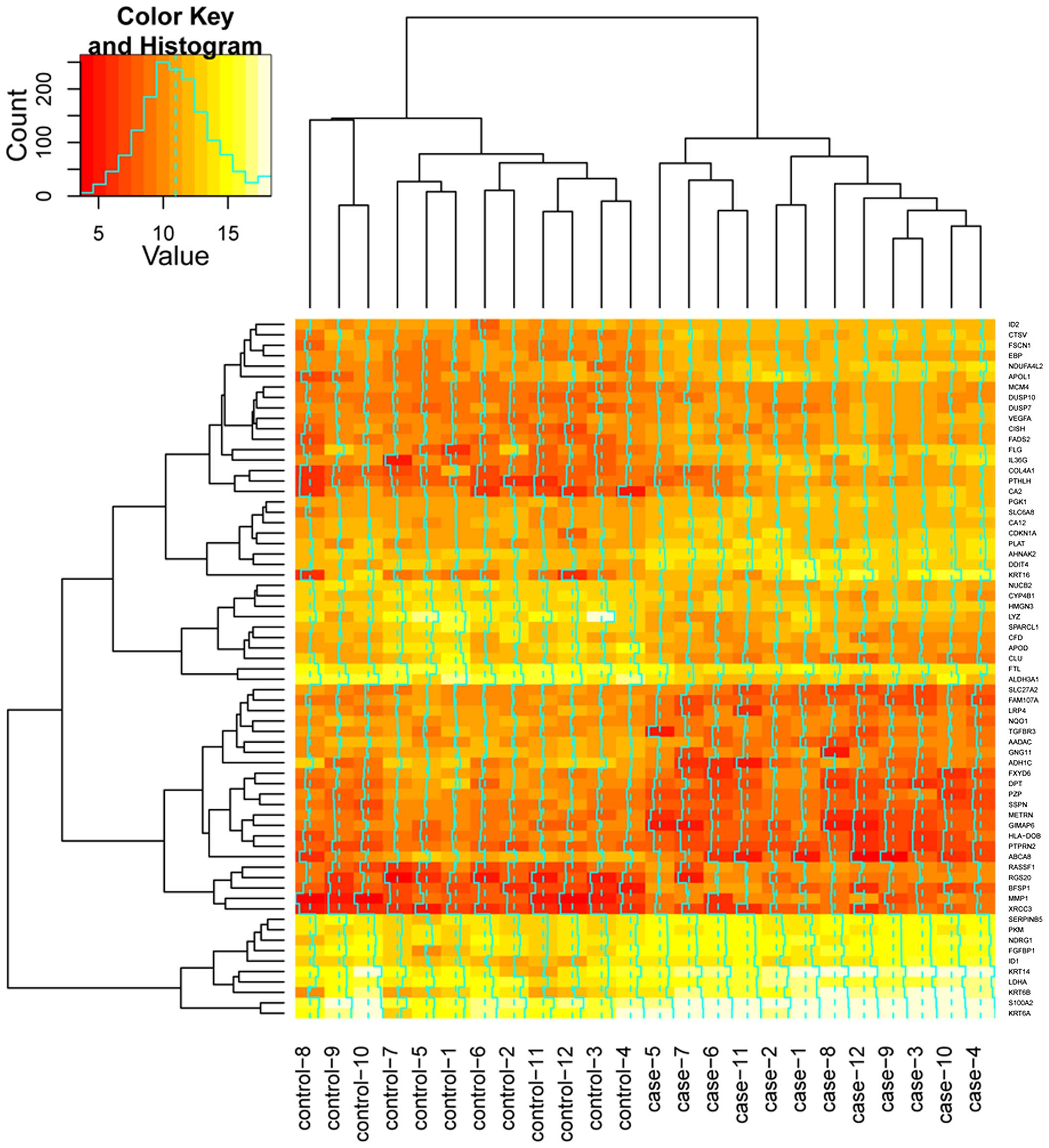
A total of 67 DEGs were identified in papilloma
compared with the normal tissues, including 27 up-regulated genes
and 40 down-regulated genes. A heat-map of the DEGs is presented in
Fig. 2. The results demonstrated that
the DEGs expression pattern could be used to distinguish the
papillomas samples from normal controls.
Functional analysis of DEGs
GO and pathway analysis indicated that up-regulated
DEGs and down-regulated DEGs in papillomas samples were
significantly enriched in different GO terms and KEGG pathways. A
total of 17 GO terms were enriched, for example, ectoderm
development, epithelium development, epithelial cell
differentiation, extracellular space, and intermediate filament
cytoskeleton (Table I). In addition,
2 KEGG pathways were significantly enriched, including bladder
cancer and glycolysis/gluconeogenesis.
 | Table I.The enriched gene ontology terms of
differentially expressed genes. |
Table I.
The enriched gene ontology terms of
differentially expressed genes.
| Category | Term | P-value | Genes |
|---|
| BP | Ectoderm
development | 1.21E-04 | PTHLH, KRT6A,
KRT6B, KRT16, FLG, KRT14, AHNAK2 |
| BP | Epithelium
development | 0.001897 | PTHLH, FLG, VEGFA,
KRT14, AHNAK2, CA2 |
| BP | Epithelial cell
differentiation | 0.002007 | PTHLH, FLG, VEGFA,
KRT14, AHNAK2 |
| BP | Epidermis
development | 0.005772 | PTHLH, KRT16, FLG,
KRT14, AHNAK2 |
| BP | Regulation of cell
proliferation | 0.010959 | PTHLH, CDKN1A,
KRT6A, ID2, VEGFA, CLU, TGFBR3, FGFBP1, DPT |
| BP | Positive regulation
of cell proliferation | 0.037925 | LDHA, ADH1C, FADS2,
NQO1, ALDH3A1, CYP4B1, FTL |
| BP | Oxidation
reduction | 0.037925 | LDHA, ADH1C, FADS2,
NQO1, ALDH3A1, CYP4B1, FTL |
| BP | Negative regulation
of molecular function | 0.041489 | PTHLH, CDKN1A, ID2,
ID1, NQO1 |
| CC | Extracellular
region part | 2.57E-09 | PLAT, PZP, COL4A1,
SPARCL1, CLU, LYZ, FXYD6, MMP1, PTHLH, METRN, APOL1, APOD,
SERPINB5, VEGFA, NUCB2, TGFBR3, CA2, CFD, FGFBP1, DPT |
| CC | Extracellular
space | 5.20E-08 | PLAT, PZP, CLU,
LYZ, FXYD6, PTHLH, METRN, APOL1, APOD, SERPINB5, VEGFA, NUCB2,
TGFBR3, CA2, CFD, FGFBP1 |
| CC | Extracellular
region | 2.02E-04 | PLAT, PZP, COL4A1,
SPARCL1, CLU, LYZ, FXYD6, MMP1, PTHLH, METRN, APOL1, APOD,
SERPINB5, VEGFA, NUCB2, TGFBR3, CA2, CFD, FGFBP1, DPT |
| CC | Intermediate
filament | 8.58E-04 | KRT6A, KRT6B,
KRT16, FLG, KRT14, BFSP1 |
| CC | Intermediate
filament cytoskeleton | 9.46E-04 | KRT6A, KRT6B,
KRT16, FLG, KRT14, BFSP1 |
| CC | Endoplasmic
reticulum | 0.038809 | AADAC, EBP, NUCB2,
TGFBR3, FADS2, SLC27A2, HLA-DOB, ALDH3A1, CYP4B1 |
| CC | Proteinaceous
extracellular matrix | 0.040784 | COL4A1, SPARCL1,
VEGFA, MMP1, DPT |
| MF | Structural
constituent of cytoskeleton | 2.31E-04 | KRT6A, KRT6B,
KRT16, KRT14, BFSP1 |
| MF | Structural molecule
activity | 0.043459 | KRT6A, KRT6B,
COL4A1, KRT16, FLG, KRT14, BFSP1 |
PPI network construction and modules
selection
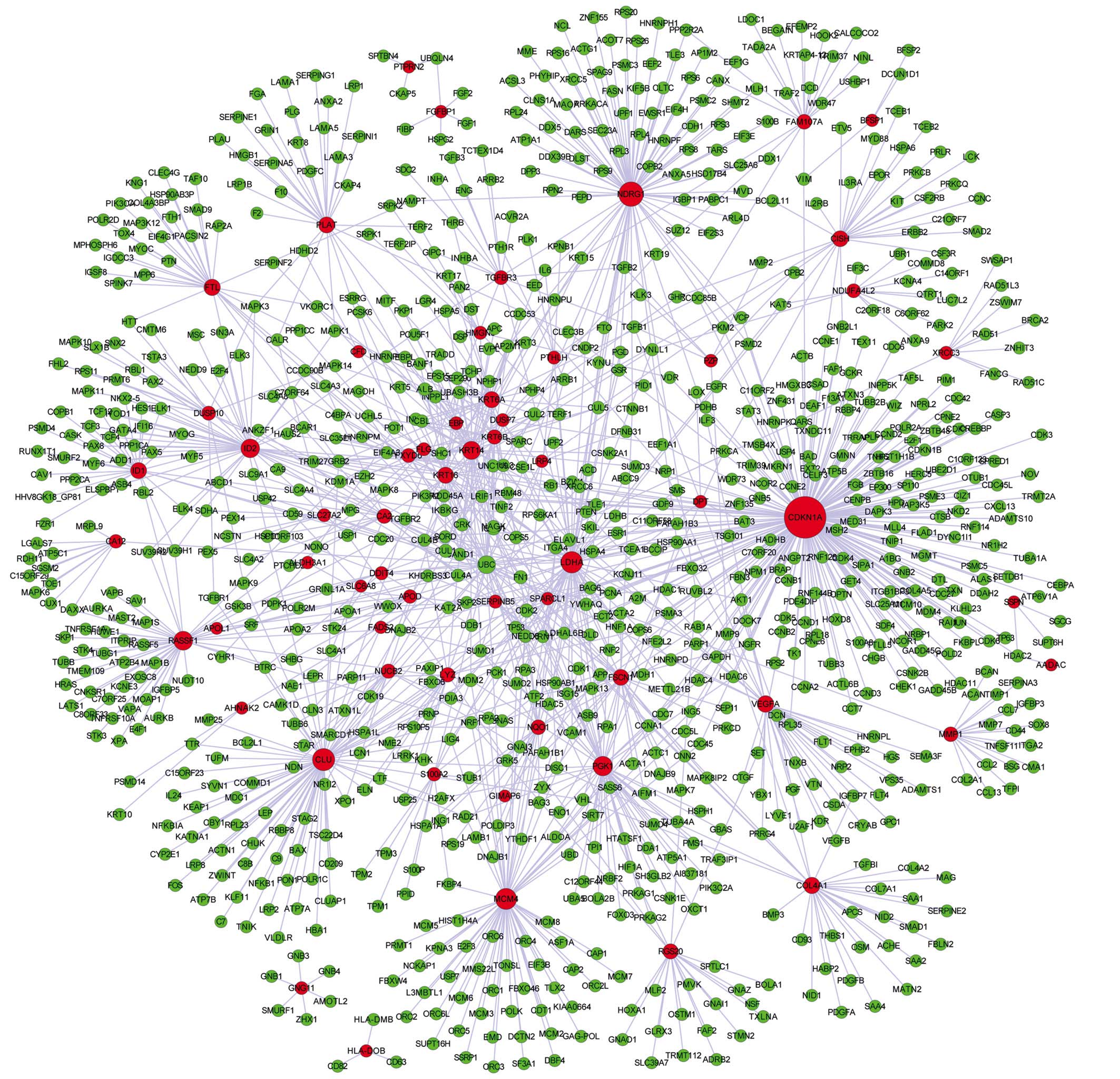
The PPI network included 984 nodes and 1,314
interactions (Fig. 3). The results
demonstrated that a total of 34 genes had a node degree of >10,
such as cyclin-dependent kinase inhibitor 1A (CDKN1A)
(degree=208), N-myc downstream regulated 1 (NDRG1)
(degree=87), lactate dehydrogenase A (LDHA) (degree=66), and
Ras association (RalGDS/AF-6) domain family member 1
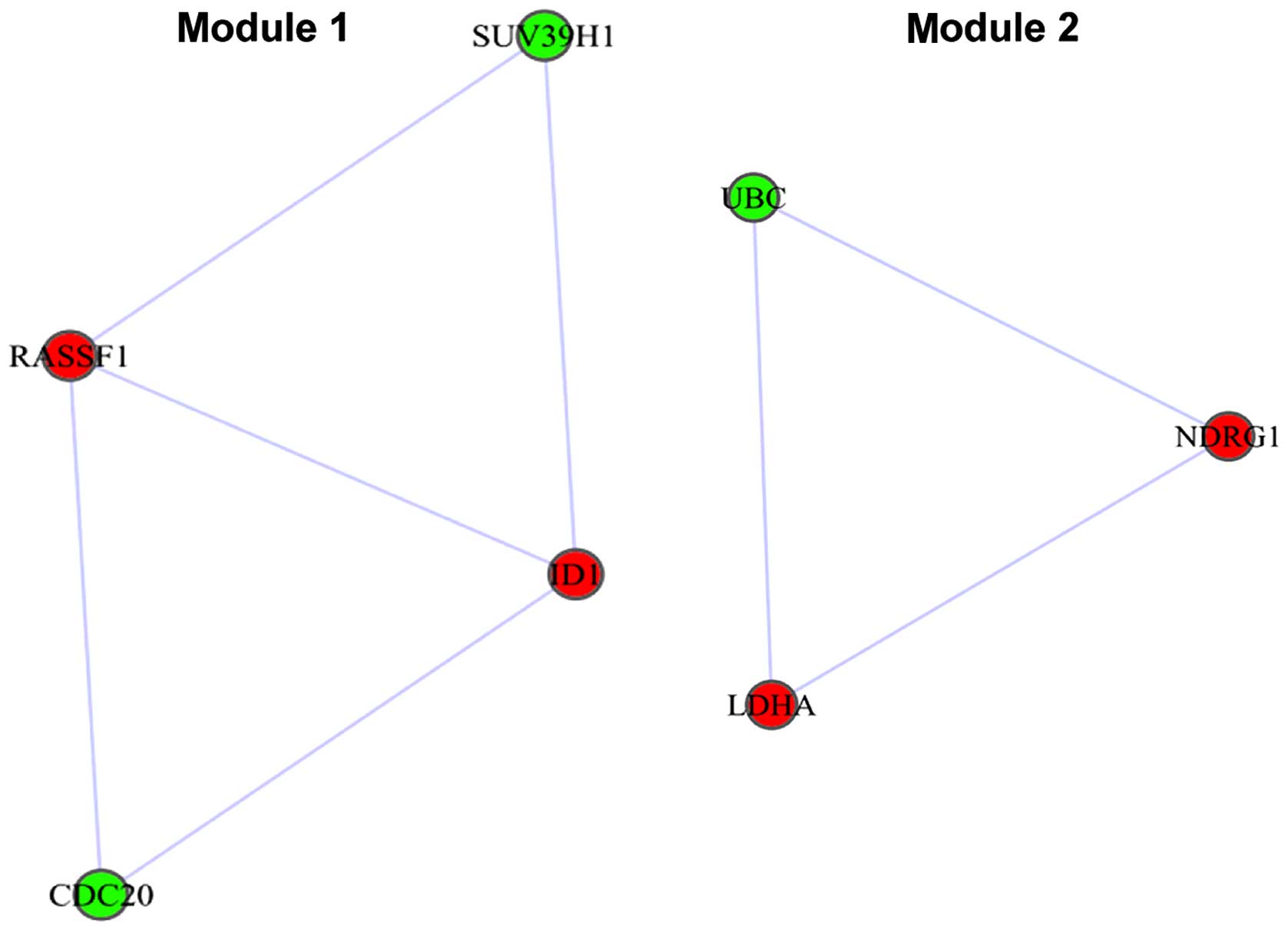
(RASSF1) (degree=48). In addition, 2 modules were identified
from the PPI network (Fig. 4). Module
1 had 2 DEGs, RASSF1 and inhibitor of DNA binding 1
(ID1), and another 2 genes, suppressor of variegation 3–9
homolog 1 (SUV39H1) and cell division cycle 20
(CDC20). Besides, module 2 had 2 DEGs, NDRG1 and
LDHA, and another gene, ubiquitin C (UBC).
Disease risk sub-pathway analysis
A total of 9 disease risk sub-pathways were
identified as presented in Table II,
such as glycolysis/gluconeogenesis, nitrogen metabolism, tyrosine
metabolism, and phenylalanine metabolism. It should be noted that
phosphoglycerate kinase 1 (PGK1), carbonic anhydrase II
(CA2), and carbonic anhydrase XII (CA12) which were
enriched in different sub-pathways including
glycolysis/gluconeogenesis and nitrogen metabolism also had a
higher node degree of >10. Moreover, aldehyde dehydrogenase 3
family, member A1 (ALDH3A1) was enriched in different
pathways, including tyrosine metabolism and phenylalanine
metabolism.
| Table II.The statistically significant
sub-pathways identified of differentially expressed genes. |
Table II.
The statistically significant
sub-pathways identified of differentially expressed genes.
| Pathway | Genes | FDR |
|---|
|
Glycolysis/Gluconeogenesis | PGK1, PKM,
ALDH3A1 | 0.000937 |
| Nitrogen
metabolism | CA2, CA12 | 0.005907 |
| Metabolism of
xenobiotics by cytochrome P450 | CYP4B1,
ALDH3A1 | 0.005907 |
| Steroid
biosynthesis | EBP | 0.027276 |
| Tyrosine
metabolism | ALDH3A1 | 0.027276 |
| Phenylalanine
metabolism | ALDH3A1 | 0.040641 |
| Drug
metabolism-cytochrome P450 | ALDH3A1,
CYP4B1 | 0.040641 |
| Fatty acid
metabolism | SLC27A2 | 0.040641 |
| Histidine
metabolism | ALDH3A1 | 0.042535 |
Discussion
Advanced laryngeal cancer is an aggressive disease,
which has a low rate of treatment efficacy and a high rate of
recurrence (29). An improved
understanding of the pathogenesis of laryngeal cancer should open
therapeutic possibilities. In the present study, DEGs in laryngeal
papillomas compared with normal controls were analyzed. The
analysis of DEGs revealed that the DEGs were enriched in different
GO terms and pathways, such as epithelial cell differentiation and
the glycolysis/gluconeogenesis pathway. Besides, RASSF1 and
ID1 were enriched in module 1 that extracted from the PPI
network of DEGs. Furthermore, PGK1, CA2 and
CA12 which had higher node degrees in the PPI network were
identified to be involved in different disease risk sub-pathways,
including glycolysis/gluconeogenesis and nitrogen metabolism.
RASSF1 encodes a protein similar to the RAS
effector proteins and may be a potential tumor suppressor (30). Agathanggelou et al (31) have previously demonstrated that
RASSF1 serves an important role in cell cycle regulation and
apoptosis as well as microtubule stability and RAS-association
domain family 1, isoform A gene (RASSF1A) inhibits tumor
growth in both in vitro and in vivo systems.
Furthermore, the findings of Sun et al (32) revealed that gene silencing through
promoter hypermethylation of CpG islands is linked to the
inactivation of tumor suppressor genes, such as RASSF1 in
head and neck cancers. By contrast, ID1 is a helix-loop-helix (HLH)
protein which inhibits the DNA binding and transcriptional
activation ability of HLH family proteins with which it interacts
(33). Coskunpinar et al
(34) quantified the expression of
tumorigenesis associated genes in laryngeal cancer and observed
that a set of 16 genes had altered expression including ID1.
Furthermore, Wang et al (35)
reported that ID1 could interact with CDC20 and
RASSF1A during early mitosis and then led to enhanced CDC20
activity. In line with the previous study, the present study
identified that RASSF1A which was a hub protein in the PPI
network could interact with ID1 in module 1 (Fig. 4), indicating that RASSF1A may
interact with ID1 influencing cell cycle regulation and
inhibiting tumor growth in laryngeal cancer.
In addition, the results of the present study showed
that PGK1 were involved in the risk sub-pathway of
glycolysis/gluconeogenesis. Glycolysis has been shown to be
enhanced in almost all cancers, termed the ‘Warburg effect’
(36). Gatenby et al (37) demonstrated that tumor cells might
sustain high glycolysis not only for biosynthetic precursor
production and energy, but also for the eradication of adjacent
normal cells within organ parenchyma. Besides, PGK1 is a glycolytic
enzyme and may also act as a polymerase alpha cofactor (primer
recognition protein) (38). Ning
et al (39) reported that
PGK1 in the glycolysis/gluconeogenesis pathway was
significantly differentially expressed between laser
micro-dissected malignant compared with benign clinical samples of
prostate tissue. Moreover, Sun et al (32) observed that PGK1 were
up-regulated in the tumor xenograft that originated from
transketolase-like 1 (TKTL1) -expressing O11 cells which was
from head and neck squamous cell carcinoma. Thus, PGK1 may
serve an essential role in the glycolysis/gluconeogenesis pathway
in promoting laryngeal cancer progression.
Furthermore, these findings revealed that CA2
and CA12 which had higher node degrees in PPI network were
associated with nitrogen metabolism sub-pathway. Glutamine is the
major source of nitrogen metabolism for nucleotide and amino acid
synthesis, although many cells can metabolize glutamine in excess
of their nitrogen requirement (40).
Tennant et al (41)demonstrated that glutamine was an
essential nutrient for many tumor cells and glutamine depletion may
contribute to the effectiveness of the drug in acute lymphoblastic
leukaemia. By contrast, CA2 and CA12 are two of several isozymes of
carbonic anhydrase which can catalyze reversible hydration of
carbon dioxide (42). Dasgupta et
al (43) discovered that
CA2 overexpressed in head and neck cancer cell line. In
addition, Parkkila et al (44)
showed that transmembrane CA12 were highly expressed in
certain tumors, and may be implicated in the acidification of the
extracellular enviroment surrounding cancer cells, thus creating a
microenvironment beneficial to tumor growth and spread. Besides,
Lascorz et al (45) found that
CA2 and CA12 were enriched in the KEGG pathway
nitrogen metabolism and the GO category carbonate dehydratase
activity in microarray studies on colorectal carcinogenesis. In
this context, CA2 and CA12 may serve crucial roles in
nitrogen metabolism beneficial to laryngeal cancer progression.
In summary, the present study identified several key
genes (RASSF1, PGK1, CA2 and CA12) that
participated in different pathways (glycolysis/gluconeogenesis, and
nitrogen metabolism) were involved in the mechanism of laryngeal
cancer. RASSF1A may inhibit tumor growth in laryngeal cancer
via interacting with ID1 involving in the cell cycle
regulation. Besides, PGK1 may serve an essential role in the
glycolysis/gluconeogenesis pathway in promoting laryngeal cancer
progression. In addition, CA2 and CA12 may be crucial
for nitrogen metabolism beneficial to laryngeal cancer progression.
These efforts may provide an improved understanding of molecular
mechanisms underlying the carcinogenesis of laryngeal cancer.
Further investigation of the genes identified in this study and
prospective studies to determine their potential clinical value are
needed.
Acknowledgements
The present study was granted by National Basic
Research Program of China grants 2002 (grant no.
0040205401078).
References
|
1
|
Licitra L, Bernier J, Grandi C, Locati L,
Merlano M, Gatta G and Lefebvre JL: Cancer of the larynx. Crit Rev
Oncol Hematol. 47:65–80. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen H, Zhou L, Dou T, Wan G, Tang H and
Tian J: BMI1′S maintenance of the proliferative capacity of
laryngeal cancer stem cells. Head Neck. 33:1115–1125. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferlito A, Rogers SN, Shaha AR, Bradley PJ
and Rinaldo A: Quality of life in head and neck cancer. Acta
Otolaryngol. 123:5–7. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Torrente MC, Rodrigo JP, Haigentz M Jr,
Dikkers FG, Rinaldo A, Takes RP, Olofsson J and Ferlito A: Human
papillomavirus infections in laryngeal cancer. Head Neck.
33:581–586. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hobbs CG and Birchall MA: Human
papillomavirus infection in the etiology of laryngeal carcinoma.
Curr Opin Otolaryngol Head Neck Surg. 12:88–92. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ma LJ, Li W, Zhang X, Huang DH, Zhang H,
Xiao JY and Tian YQ: Differential gene expression profiling of
laryngeal squamous cell carcinoma by laser capture microdissection
and complementary DNA microarrays. Arch Med Res. 40:114–123. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nakanishi H, Taccioli C, Palatini J,
Fernandez-Cymering C, Cui R, Kim T, Volinia S and Croce C: Loss of
miR-125b-1 contributes to head and neck cancer development by
dysregulating TACSTD2 and MAPK pathway. Oncogene. 33:702–712. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Järvinen A, Autio R, Haapa-Paananen S,
Wolf M, Saarela M, Grénman R, Leivo I, Kallioniemi O, Mäkitie AA
and Monni O: Identification of target genes in laryngeal squamous
cell carcinoma by high-resolution copy number and gene expression
microarray analyses. Oncogene. 25:6997–7008. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fountzilas E, Markou K, Vlachtsis K,
Nikolaou A, Arapantoni-Dadioti P, Ntoula E, Tassopoulos G, Bobos M,
Konstantinopoulos P, Fountzilas G and Spentzos D: Identification
and validation of gene expression models that predict clinical
outcome in patients with early-stage laryngeal cancer. Ann Oncol.
23:2146–2153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Colombo J, Fachel AA, De Freitas Calmon M,
Cury PM, Fukuyama EE, Tajara EH, Cordeiro JA, Verjovski-Almeida S,
Reis EM and Rahal P: Gene expression profiling reveals molecular
marker candidates of laryngeal squamous cell carcinoma. Oncol Rep.
21:649–663. 2009.PubMed/NCBI
|
|
11
|
Oblak A and Jerala R: Toll-like receptor 4
activation in cancer progression and therapy. Clin Dev Immunol.
2011:609–579. 2011. View Article : Google Scholar
|
|
12
|
DeVoti JA, Rosenthal DW, Wu R, Abramson
AL, Steinberg BM and Bonagura VR: Immune dysregulation and
tumor-associated gene changes in recurrent respiratory
papillomatosis: A paired microarray analysis. Mol Med. 14:608–617.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Parker HS, Leek JT, Favorov AV, Considine
M, Xia X, Chavan S, Chung CH and Fertig EJ: Preserving biological
heterogeneity with a permuted surrogate variable analysis for
genomics batch correction. Bioinformatics. 30:2757–2763. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ihaka R and Gentleman R: R: A language for
data analysis and graphics. J Comput Graph Stat. 5:299–314. 1996.
View Article : Google Scholar
|
|
15
|
Yuan K, Liang W and Zhang J: A
comprehensive analysis of differentially expressed genes and
pathways in abdominal aortic aneurysm. Mol Med Rep. 12:2707–2714.
2015.PubMed/NCBI
|
|
16
|
Cui X and Churchill GA: Statistical tests
for differential expression in cDNA microarray experiments. Genome
Biol. 4:2102003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Diboun I, Wernisch L, Orengo CA and
Koltzenburg M: Microarray analysis after RNA amplification can
detect pronounced differences in gene expression using limma. BMC
Genomics. 7:2522006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bland JM and Altman DG: Multiple
significance tests: The Bonferroni method. BMJ. 310:1701995.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Belda-Ferre P, Alcaraz LD, Cabrera-Rubio
R, et al: The oral metagenome in health and disease. ISME J.
6:46–56. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
da Huang W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Köhler S, Bauer S, Horn D and Robinson PN:
Walking the interactome for prioritization of candidate disease
genes. Am J Hum Genet. 82:949–958. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Prasad Keshava TS, Goel R, Kandasamy K,
Keerthikumar S, Kumar S, Mathivanan S, Telikicherla D, Raju R,
Shafreen B, Venugopal A, et al: Human protein reference
database-2009 update. Nucleic Acids Res. 37(Database Issue):
D767–D772. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Stark C, Breitkreutz BJ, Chatr-Aryamontri
A, Boucher L, Oughtred R, Livstone MS, Nixon J, Van Auken K, Wang
X, Shi X, et al: The BioGRID interaction database: 2011 update.
Nucleic Acids Res. 39(Database Issue): D698–D704. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bandettini WP, Kellman P, Mancini C,
Booker OJ, Vasu S, Leung SW, Wilson JR, Shanbhag SM, Chen MY and
Arai AE: MultiContrast delayed enhancement (MCODE) improves
detection of subendocardial myocardial infarction by late
gadolinium enhancement cardiovascular magnetic resonance: A
clinical validation study. J Cardiovasc Magn Reson. 14:832012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li C and Li MC: Package
‘iSubpathwayMiner’. 2013.
|
|
28
|
Chen X, Xu J, Huang B, Li J, Wu X, Ma L,
Jia X, Bian X, Tan F, Liu L, et al: A sub-pathway-based approach
for identifying drug response principal network. Bioinformatics.
27:649–654. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ayaz L, Görür A, Yaroğlu HY, Özcan C and
Tamer L: Differential expression of microRNAs in plasma of patients
with laryngeal squamous cell carcinoma: Potential early-detection
markers for laryngeal squamous cell carcinoma. J Cancer Res Clin
Oncol. 139:1499–1506. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tommasi S, Dammann R, Jin SG, Zhang XF,
Avruch J and Pfeifer GP: RASSF3 and NORE1: Identification and
cloning of two human homologues of the putative tumor suppressor
gene RASSF1. Oncogene. 21:2713–2720. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Agathanggelou A, Cooper WN and Latif F:
Role of the Ras-association domain family 1 tumor suppressor gene
in human cancers. Cancer Res. 65:3497–3508. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun W, Liu Y, Glazer CA, Shao C, Bhan S,
Demokan S, Zhao M, Rudek MA, Ha PK and Califano JA: TKTL1 is
activated by promoter hypomethylation and contributes to head and
neck squamous cell carcinoma carcinogenesis through increased
aerobic glycolysis and HIF1alpha stabilization. Clin Cancer Res.
16:857–866. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sakurai D, Yamaguchi A, Tsuchiya N,
Yamamoto K and Tokunaga K: Expression of ID family genes in the
synovia from patients with rheumatoid arthritis. Biochem Biophys
Res Commun. 284:436–442. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Coskunpinar E, Oltulu YM, Orhan KS,
Tiryakioglu NO, Kanliada D and Akbas F: Identification of a
differential expression signature associated with tumorigenesis and
metastasis of laryngeal carcinoma. Gene. 534:183–188. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang X, Di K, Zhang X, Han HY, Wong YC,
Leung SC and Ling MT: Id-1 promotes chromosomal instability through
modification of APC/C activity during mitosis in response to
microtubule disruption. Oncogene. 27:4456–4466. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bi X, Lin Q, Foo TW, Joshi S, You T, Shen
HM, Ong CN, Cheah PY, Eu KW and Hew CL: Proteomic analysis of
colorectal cancer reveals alterations in metabolic pathways:
Mechanism of tumorigenesis. Mol Cell Proteomics. 5:1119–1130. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gatenby RA and Gillies RJ: Why do cancers
have high aerobic glycolysis? Nat Rev Cancer. 4:891–899. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Valentini G, Maggi M and Pey AL: Protein
stability, folding and misfolding in human PGK1 deficiency.
Biomolecules. 3:1030–1052. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ning QY, Wu JZ, Zang N, Liang J, Hu YL and
Mo ZN: Key pathways involved in prostate cancer based on gene set
enrichment analysis and meta analysis. Genet Mol Res. 10:3856–3887.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Vander Heiden MG: Targeting cancer
metabolism: A therapeutic window opens. Nat Rev Drug Discov.
10:671–684. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tennant DA, Durán RV and Gottlieb E:
Targeting metabolic transformation for cancer therapy. Nat Rev
Cancer. 10:267–277. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lindskog S: Structure and mechanism of
carbonic anhydrase. Pharmacol Ther. 74:1–20. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dasgupta S, Tripathi PK, Qin H,
Bhattacharya-Chatterjee M, Valentino J and Chatterjee SK:
Identification of molecular targets for immunotherapy of patients
with head and neck squamous cell carcinoma. Oral Oncol. 42:306–316.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Parkkila S, Rajaniemi H, Parkkila AK,
Kivela J, Waheed A, Pastorekova S, Pastorek J and Sly WS: Carbonic
anhydrase inhibitor suppresses invasion of renal cancer cells in
vitro. Proc Natl Acad Sci USA. 97:2220–2224. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lascorz J, Hemminki K and Försti A:
Systematic enrichment analysis of gene expression profiling studies
identifies consensus pathways implicated in colorectal cancer
development. J Carcinog. 10:72011. View Article : Google Scholar : PubMed/NCBI
|