Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common tumor in men and the seventh most common tumor in women. It
is the third leading cause of tumor-associated mortality worldwide
(1). HCC has an extremely poor
prognosis, with a 5-year survival rate of 40–50%. Patients with HCC
are usually diagnosed at an advanced tumor stage, which renders
surgical resection inapplicable (2).
The molecular mechanism underlying HCC cell proliferation is not
well understood, and the clarification of this mechanism may
facilitate the prevention and treatment of patients with HCC.
Approximately one-half of HCC patients exhibit
aberrant activation of the Wnt/β-catenin pathway, which indicates
the importance of this signaling cascade in hepatocarcinogenesis
(3–5).
The Wnt/β-catenin signaling pathway is comprehensively involved in
cell proliferation (6). The
Wnt/β-catenin pathway is activated upon the binding of the Wnt
ligands to Frizzled receptors, which is followed by cytosolic
accumulation of β-catenin via interruption of destruction
complex-mediated protein degradation of β-catenin. Stabilized
β-catenin may translocate into the nucleus to induce the
transcription of Wnt/β-catenin pathway target genes, such as cyclin
D1 and cellular v-myc avian myelocytomatosis viral oncogene homolog
(c-MYC) (4,7). The Wnt/β-catenin signaling pathway plays
a vital role in hepatocarcinogenesis. Although several factors,
such as activator protein 1, human zinc finger protein 191,
prospero homeobox protein 1 and T-cell factor (TCF)/lymphocyte
enhancer factor (LEF), have been reported to regulate β-catenin
expression (8–11), the molecular mechanism of the aberrant
expression of β-catenin remains unknown in HCC.
SWItch/sucrose nonfermentable (SWI/SNF) catalytic
subunit SNF2 (SNF2H) is a member of the SWI/SNF chromatin
remodeling family with ATPase activity (12). SNF2H plays a vital role in gene
transcription, DNA repair and DNA replication (12,13).
SWI/SNF complexes are associated with malignant transformation
(14). Previously, several studies
have reported aberrant high expression of SNF2H in breast cancer,
acute leukemia, ovarian cancer and gastric cancer (15–17). These
studies suggest that SNF2H has an important role in cancer
progression. However, whether SNF2H plays a vital role in HCC
progression has not been investigated.
In the present study, it was demonstrated that SNF2H
plays a vital role in HCC cell growth. The SNF2H expression level
was increased in HCC tissues compared with the paratumoral liver
tissues. SNF2H promotes the cell proliferation and colony formation
ability of HCC cells in vitro. SNF2H promotes the growth of
HCC cells by activating the Wnt/β-catenin pathway, which increases
β-catenin protein levels and enhances the nuclear accumulation
β-catenin. These results indicate that SNF2H plays a vital role in
hepatocarcinogenesis.
Materials and methods
Clinical samples
Tissue specimens were collected from 30 patients
with HCC during surgical resection. The collection of the tissue
specimens and associated information from patients was approved by
the Ethics Committee of Renji Hospital, Shanghai Jiaotong
University School of Medicine (Shanghai, China). All patients
enrolled in the present study provided written informed consent
prior to their inclusion.
Plasmid constructs
For lentivirus-mediated expression of SNF2H,
FLAG-tagged full-length SNF2H ORF was cloned in pWPI.1 (plasmid no.
12254; Addgene, Inc., Cambridge, MA, USA). For RNA interference of
SNF2H expression, DNA fragments encoding the hairpin precursors for
shSNF2H#1 (5′-CGTCGAATTAAGGCTGATGTT-3′) and shSNF2H#2
(5′-CGACTGCTGATGTAGTAATTT-3′) were inserted into the pLKO.1-TRC
cloning vector. A scrambled small interfering RNA precursor (Scr)
of similar GC-content to shSNF2H#1 and shSNF2H#2, but without
sequence identity to SNF2H cDNA, was used as the control.
Cell lines, transfection and
lentivirus preparation
The embryo kidney HEK293T and human HCC SMCC-7721,
PLC, BEL-7402 and Huh7 cell lines were acquired from the Cell Bank
of Shanghai Institutes of Biological Sciences, Chinese Academy of
Sciences (Shanghai, China). The cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) with 10% fetal bovine serum (Thermo Fisher
Scientific, Inc.) at 37°C in a 5% CO2 atmosphere. After
culturing cells for 24 h, cells underwent transfection.
The lentivirus plasmid-mediated SNF2H expression
vectors pWPI.1-Flag-SNF2H, pLKO.1-shSNF2H#1 and pLKO.1-shSNF2H#2
were co-transfected with the helper plasmids pMD2.G and psPAX2 into
HEK293T cell to package recombinant lentivirus. In total, 24 h
subsequent to cell transfection, medium was replaced with fresh
DMEM medium to continue cell culture. After 48 h, the lentivirus
supernatants from HEK293T cells were used for the infection of
BEL-7402 and Huh7 cells using Lipofectamine 2000 (Thermo Fisher
Scientific, Inc.).
Western blot analysis
Total tissue and cell proteins were obtained using
radioimmunoprecipitation assay buffer lysis buffer with proteinase
inhibitors (protease inhibitor cocktail; Sigma-Aldrich, St. Louis,
MO, USA). Equivalent amounts of proteins (30 µg) were separated by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis and
transferred onto nitrocellulose membranes (Invitrogen; Thermo
Fisher Scientific, Inc.). Subsequent to blocking with 5% skimmed
milk for 30 min, the membranes were incubated with a rabbit
polyclonal anti-SNF2H primary antibody (dilution, 1:500; catalog
no., ab3749; Abcam, Cambridge, MA, USA) at 4°C overnight. A goat
anti-rabbit IgG horseradish peroxidase-conjugated secondary
antibody (dilution, 1:3,000; catalog no., 7074; Cell Signaling
Technology, Inc., Danvers, MA, USA) was used for the subsequent
incubation at room temperature for 1 h. In addition, a mouse
monoclonal β-actin horseradish peroxidase-conjugated antibody
(dilution, 1:5,000; catalog no., ab20272; Abcam) was used as a
loading control. The expression of SNF2H was detected on X-ray film
using the ECL detection system (Pierce; Thermo Fisher Scientific,
Inc.).
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA of the sample was extracted using TRIzol
reagent (Thermo Fisher Scientific, Inc.), and RT was performed
using the PrimeScript RT kit (Takara, Otsu, Japan). For the RT-qPCR
assay, aliquots of cDNA were amplified using SYBR Premix Ex Taq
(Takara). PCR reactions were performed on MXP3000 (Stratagene, La
Jolla, CA, USA). The primer sequences were as follows: SFN2H
forward, CCTTTGAAGATGAAACCAGGGCGC and reverse,
CTGTTAATAGCTCTTCATCCTCCTC; and β-actin forward, TCCCTGGAGAAGAGCTACG
and reverse, GTAGTTTCGTGGATGCCACA.
Cell proliferation
Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) to measure the cell
proliferation. BEL-7402-Mock, BEL-7402-SNF2H, Huh7-scr,
Huh7-shSNF2H#1 and Huh7-shSNF2H#2 cell lines were seeded into
96-well plates at a density of 2×103 cells/well with
DMEM medium containing 10% FBS. At the indicated time points (24,
48, 72 or 96 h), the cell absorbance was detected at 450 nm using
xMark Microplate Spectrophotometer (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The CCK-8 assay procedure was performed
according to the manufacturer's instructions.
Colony formation assay
BEL-7402-Mock, BEL-7402-SNF2H, Huh7-scr,
Huh7-shSNF2H#1 and Huh7-shSNF2H#2 cell lines were separately seeded
into 6-well plates at a density of 3×102 cells/well
using DMEM medium containing 10% FBS, and the cells were cultured
at 37°C in a 5% CO2 atmosphere. After 10–14 days of
culturing, the cells were washed with PBS and fixed using 10%
formaldehyde for 30 min. Finally, the cells were stained with
Giemsa stain for 60 min.
Statistical Analysis
Pearson's χ2 test or Fisher's exact test
were used to compare SNF2H expression levels between tissue samples
and the Student's t-test was used for quantitative variables.
P<0.05 was considered to indicate a statistically significant
difference.
Results
SNF2H is highly expressed in HCC
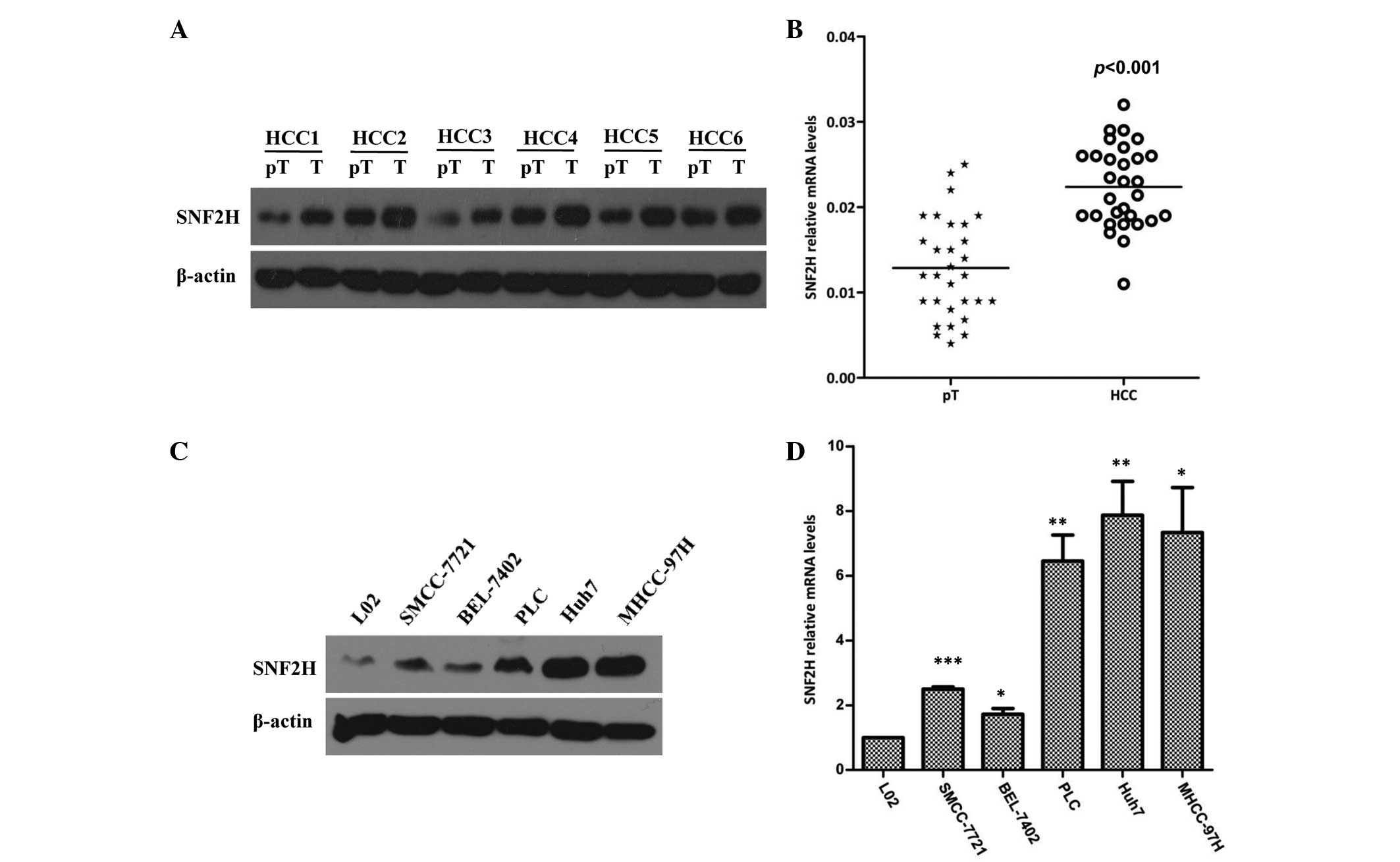
The expression of SNF2H was first assessed in HCC
and paired paratumoral liver tissues. The SNF2H protein level was
significantly increased by 1.7-fold in HCC tissues compared with
the matched paratumoral liver tissues (P=0.018; Fig. 1A). To confirm these results, the
expression of SNF2H in 30 HCC specimens and the paired paratumoral
liver tissues was evaluated. It was found that the expression of
SNF2H in HCC specimens was increased 1.9-fold compared with the
paired paratumoral liver tissues (P<0.001; Fig. 1B). Next, the expression of SNF2H in
the normal liver cell and HCC SMCC-7721, BEL-7402, PLC, Huh7 and
MHCC-97H cell lines was examined. Western blotting showed that the
HCC Huh7 and MHCC97H cell lines had increased SNF2H expression
compared with the normal liver L02 cell line (Fig. 1C). It was also found that the mRNA
level of SNF2H in the HCC SMCC-7721, BEL-7402, PLC, Huh7 and
MHCC-97H cell lines was increased 2.5-fold, 1.7-fold, 6.34-fold,
7.72-fold and 7.09-fold, respectively, compared with the mRNA level
in the L02 cell line (P=0.0018, P=0.0160, P=0.0025, P=0.0027 and
P=0.0102, respectively; Fig. 1D).
Overall, these results indicate that the high expression of SNF2H
may improve the development of HCC.
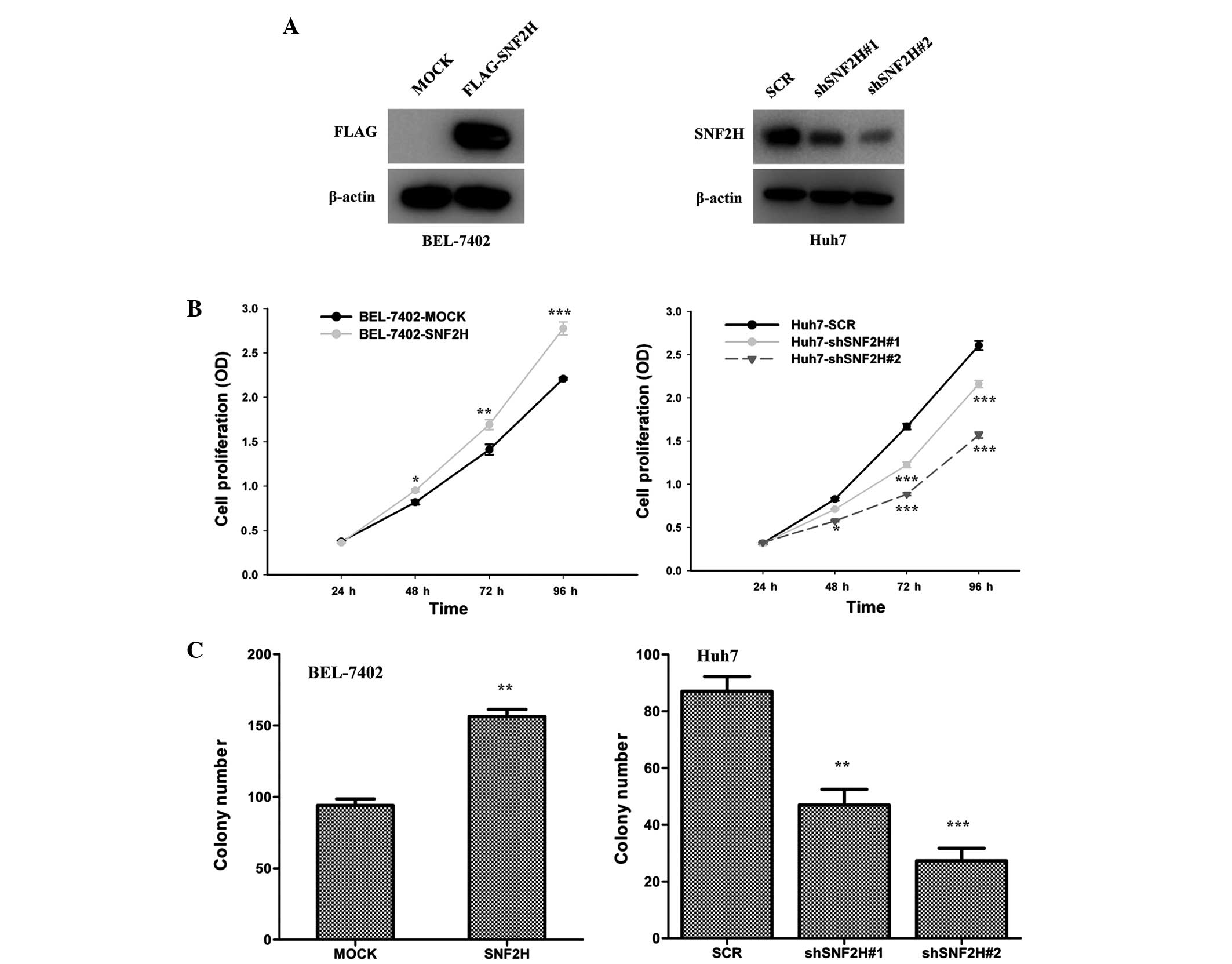
Overexpression of SNF2H improves HCC
cell growth
BEL-7402 and Huh7 cells were used as model HCC cell
lines for the following investigations. Lentivirus-mediated
overexpression of SNF2H in the BEL-7402 cells and knockdown of
SNF2H expression of SNF2H in the Huh7 cells were used to evaluated
the function of SNF2H in HCC cell growth. Overexpression of SNF2H
in BEL-7402 cells and knockdown of the expression of SNF2H in the
Huh7 cells were performed (Fig. 2A).
Overexpression of SNF2H in BEL-7402 cells improved the cell growth
over 96 h (24 h, P=0.1070; 48 h, P=0.0015; 72 h, P<0.001; 96 h,
P<0.001), but knockdown of SNF2H expression in Huh7 cells
inhibited cell growth (shSNF2H#1 vs. SCR: 24 h, P=0.97; 48 h,
P=0.0054; 72 h, P<0.001; and 96 h, P<0.001) (shSNF2H#2 vs.
SCR: 24 h, P=0.345; 48 h, P=0.001; 72 h, P<0.001; and 96 h,
P<0.001) (Fig. 2B). Overexpression
of SNF2H also increased the number of BEL-7402 cell colonies by
1.6-fold (P<0.001) (Fig. 2C).
However, knockdown of SNF2H expression reduced the number of Huh7
cell colonies by 50 and 70%, respectively (shSNF2H#1 vs. SCR,
P=0.006; shSNF2H#2 vs. SCR, P<0.001) (Fig. 2C). These results indicate that SNF2H
improves HCC cell growth.
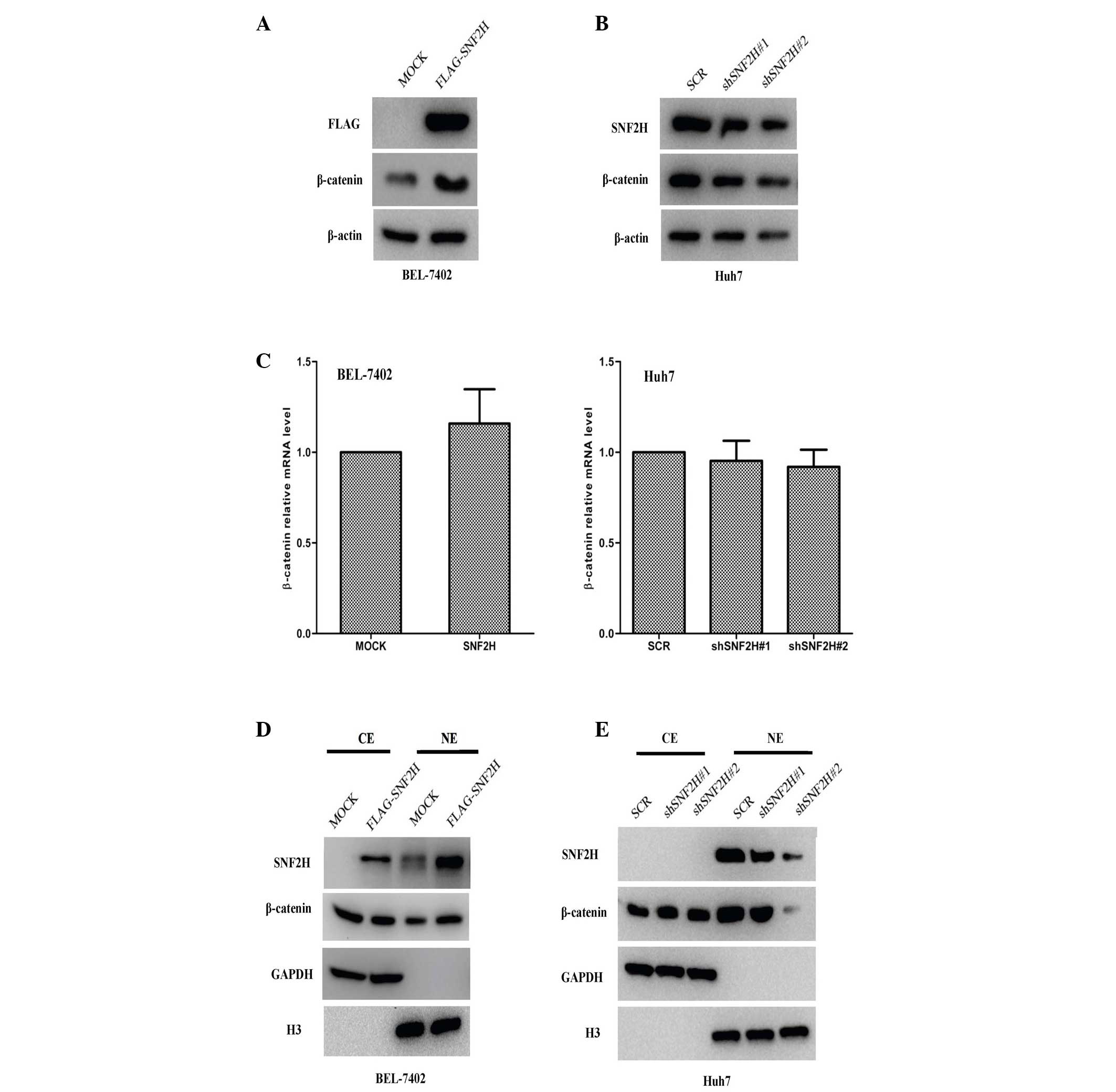
SNF2H promotes the protein levels of
β-catenin and enhances its nuclear accumulation
The Wnt/β-catenin pathway is always aberrantly
activated during the development of HCC, and the β-catenin
expression level is often increased in HCC (3–5,18,19).
Therefore, the present study investigated whether SNF2H enhances
β-catenin expression to promote tumor growth. It was found that the
β-catenin protein level in BEL-7402 cells was upregulated 2.6-fold
by the exogenous expression of SNF2H (P=0.0012; Fig. 3A), but the β-catenin protein level was
downregulated by 30% (shSNF2H#1 vs. SCR, P=0.008) and 70%
(shSNF2H#2 vs. SCR, P=0.001) in Huh7 cells by SNF2H knockdown
(Fig. 3B). The increase in β-catenin
protein levels may result in induced β-catenin transcription or
improved β-catenin protein stability. The present study
investigated whether SNF2H used this mechanism to upregulate
β-catenin protein levels. Subsequently, it was observed that
neither SNF2H overexpression in BEL-7402 cells nor SNF2H knockdown
in Huh7 cells resulted in different β-catenin mRNA levels compared
with the control BEL-7402-MOCK (increase of 1.1-fold; P=0.158) and
Huh7-Scr cells (shSNF2H#1 vs. SCR, decrease of 7%, P=0.62;
shSNF2H#2 vs. SCR, increase of 8%, P=0.574) (Fig. 3C). In addition, there were no
differences in the cytoplasmic level of β-catenin. However, the
nuclear level of β-catenin was markedly upregulated 2-fold in the
SNF2H overexpression BEL-7402 cells compared with the control group
(P=0.003) (Fig. 3D). Knockdown of
SNF2H did not change the cytoplasmic level of β-catenin, but the
nuclear level of β-catenin was markedly downregulated by 20 and
70%, respectively, compared with the control group in the Huh7
cells (shSNF2H#1 vs. SCR, P=0.023; shSNF2H#2 vs. SCR, P=0.002)
(Fig. 3E). These results indicate
that SNF2H may promote β-catenin protein levels and enhance the
nuclear accumulation of β-catenin in HCC cells.
SNF2H activates the Wnt/β-catenin
signaling pathway
SNF2H increases the protein level of β-catenin and
enhance its nuclear translocation in HCC cells. These findings
indicate that SNF2H may drive the Wnt/β-catenin signaling pathway
in HCC cells. To test this hypothesis, it was investigated whether
overexpression of SNF2H in BEL-7402 cells could regulate the
transcription of the Wnt/β-catenin target genes, such as cyclin D1
and c-MYC. SNF2H overexpression in BEL-7402 cells significantly
upregulated c-MYC and cyclin D1 mRNA by 2.5-fold (P=0.0056 vs.
MOCK) and 2.2-fold (P<0.001 vs. MOCK), respectively, and protein
levels (Fig. 4A and B). However,
knockdown of the expression of SNF2H decreased the mRNA expression
of cyclin D1 by 60% (shSNF2H#1 vs. SCR, P<0.001; shSNF2H#2 vs.
SCR, P=0.0053) and c-MYC by 50% (shSNF2H#1 vs. SCR, P=0.0037;
shSNF2H#2 vs. SCR, P<0.001) and also decreased protein levels in
Huh7 cells compared with the control group (Fig. 4A and B). It was investigated whether
SNF2H could activate the LEF luciferase reporter that is always
used as indicator of the level of Wnt/β-catenin signaling pathway
activity. Indeed, exogenous expression of SNF2H in HEK-293T cells
could enhance the LEF luciferase reporter activity by 3.0-fold
compared with the control group (P<0.001) (Fig. 4C). Next, it was investigated whether
SNF2H could also activate the LEF luciferase reporter activity in
the HCC cells. Overexpression of SNF2H in BEL-7402 cells enhanced
LEF luciferase reporter activity by 2.0-fold compared with the
control group (P=0.0031) (Fig. 4D).
However, knockdown of SNF2H expression in Huh7 cells decreased LEF
luciferase reporter activity by 50% (shSNF2H#1 vs. SCR, P=0.0166)
and 60% (shSNF2H#2 vs. SCR, P=0.0018) compared with the control
group (Fig. 4E). These results
indicate that SNF2H activates the Wnt/β-catenin signaling pathway
to promote HCC cell growth.
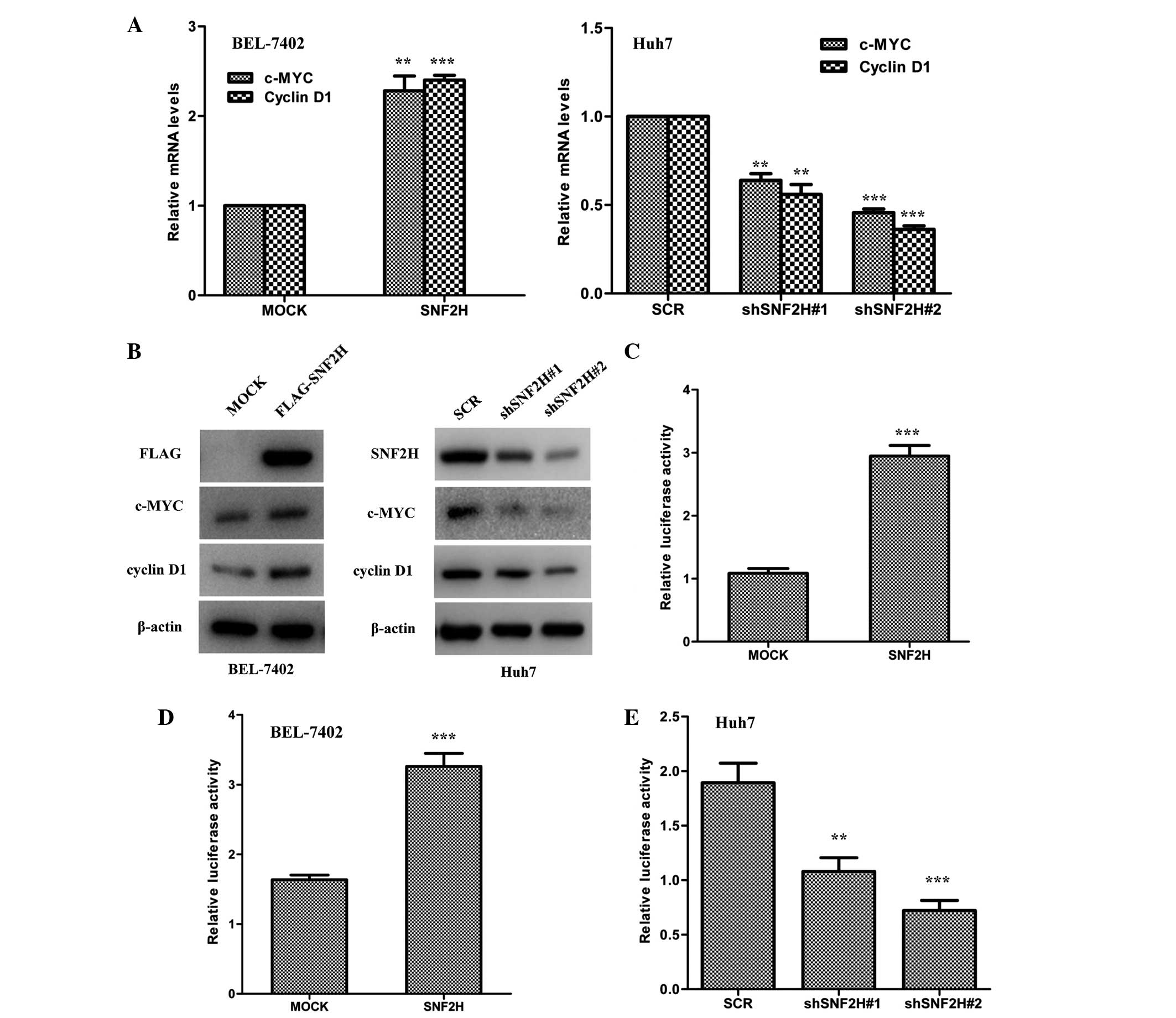 | Figure 4.SNF2H activates the Wnt/β-catenin
pathway. (A) Left, SNF2H overexpression in BEL-7402 cells
upregulated c-MYC and cyclin D1 mRNA levels. Right, SNF2H knockdown
in Huh7 cells downregulated c-MYC and cyclin D1 mRNA levels. The
mean ± SD from three independent experiments is presented as a
relative ratio to the control, which was assigned a value of 1.0.
(B) Left, overexpression of SNF2H increased c-MYC and cyclin D1
protein levels in BEL-7402 cells. Right, knockdown of SNF2H
decreased c-MYC and cyclin D1 protein levels in Huh7 cells. (C)
SNF2H activated the LEF reporter. HEK293T cells were co-transfected
with the SNF2H and the LEF reporter expression constructs. (D)
Overexpression of SNF2H upregulated LEF reporter activity in
BEL-7402 cells. (E) Knockdown of SNF2H expression downregulated LEF
reporter activity in Huh7 cells. The mean ± SD of normalized
luciferase activity from three independent experiments is
presented. Significant differences were determined using Student's
t-test. **P<0.01; ***P<0.001. SNF2H, SWItch/sucrose
nonfermentable catalytic subunit SNF2; c-MYC, cellular v-myc avian
myelocytomatosis viral oncogene homolog; mRNA, messenger RNA; LEF,
lymphocyte enhancer factor; SD, standard deviation. |
Discussion
HCC is the fifth most common cancer and the third
most common cause of cancer-associated mortality worldwide. The
incidence and mortality of HCC has been increasing rapidly
worldwide in previous decades. China alone accounts for >50% of
cases of HCC worldwide (20,21). Due to the poor prognosis of HCC
patients, the molecular mechanism of HCC development urgently
requires clarification. Although numerous genes associated with the
development of HCC have been confirmed (22–26), the
mechanism of HCC development has not been well elucidated. In the
present study, the expression of SNF2H in HCC specimens and the
paired paratumoral liver tissues was assessed. It was found that
the expression of SNF2H in HCC specimens was increased compared
with the paired paratumoral liver tissues. The present findings
suggest that SNF2H may be a novel intervention target for the
treatment of HCC.
The extremely poor prognosis of HCC is mainly due to
tumor growth and vascular invasion (27–30). The
molecular mechanism of HCC cell growth is not well known.
Therefore, a thorough knowledge of the molecular mechanisms of HCC
cell growth is extremely important for the therapeutics for HCC
patients. The present results showed that SNF2H plays a vital role
in promoting HCC cell growth and colony formation ability,
indicating that SNF2H is a vital factor for the development of
HCC.
The Wnt/β-catenin signaling pathway is aberrantly
activated during the development of HCC (3–5). When
binding with Wnt ligands, the destruction complex, which includes
adenomatous polyposis coli protein, casein kinase 1, Axin and
glycogen synthase kinase 3, may not induce the degradation of
β-catenin. Further, stabilized β-catenin may induce the expression
of Wnt/β-catenin target genes by interacting with TCF/LEF
transcription factors. Previous data show that certain Wnt target
genes, such as cyclin D1 and c-MYC, regulate tumor cell growth
(6,31). However, the association between SNF2H
and the Wnt/β-catenin pathway in HCC remains unknown. In the
present study, SNF2H was shown to activate the Wnt/β-catenin
pathway in HCC cells by upregulating β-catenin protein levels and
enhancing β-catenin nuclear translocation. The mechanism for
SNF2H-mediated upregulation of the expression of β-catenin requires
elucidation. Neither SNF2H-overexpression BEL-7402 cells nor
SNF2H-knockdown Huh7 cells regulated β-catenin mRNA levels. Thus,
it is possible that SNF2H may interfere with the degradation
machinery of β-catenin. The mechanism of SNF2H enhancement of the
nuclear translocation of β-catenin is a vital focus for additional
investigation, which may indicate a novel target for the treatment
of HCC patients.
SNF2H plays an important role in cell growth and
viability (32,33). Stopka et al (13) reported that knockout of SNF2H reduced
cell proliferation by the inhibition of mitosis. The present
results showed that SNF2H also promotes cell proliferation in HCC.
The role of SNF2H in carcinogenesis is well established. Mohamed
et al (12) reported that
SNF2H showed higher expression in prostatic neoplasia compared with
benign prostatic hyperphasia. The results of this study suggest
that SNF2H is important in prostate carcinogenesis (12). Previously, another study observed
increased expression of SNF2H in gastric cancer samples compared
with normal mucosa (16). However, no
study has reported an association between SNF2H and HCC. The
present study demonstrated that SNF2H expression was markedly
upregulated in HCC tissues compared with the paired paratumoral
liver tissues. The present results indicate that SNF2H may be a
promising therapeutic target strategy for HCC patients.
Acknowledgements
The present study was sponsored by the National
Natural Science Foundation of China (grant nos. 81322025 and
81371875), and the Shanghai Committee of Science and Technology,
China (grant no. 14140901000) and the Foundation for Innovative
Research Groups of the National Natural Science Foundation of China
(grant no. 81421001).
References
|
1
|
Ferlay J, Shin H, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Forner A, Hessheimer AJ, Isabel Real M and
Bruix J: Treatment of hepatocellular carcinoma. Crit Rev Oncol
Hematol. 60:89–98. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boyault S, Rickman DS, deReyniès A,
Balabaud C, Rebouissou S, Jeannot E, Hérault A, Saric J, Belghiti
J, Franco D, et al: Transcriptome classification of HCC is related
to gene alterations and to new therapeutic targets. Hepatology.
45:42–52. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takigawa Y and Brown AM: Wnt signaling in
liver cancer. Curr Drug Targets. 9:1013–1024. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Polakis P: The many ways of Wnt in cancer.
Curr Opin Genet Dev. 17:45–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Valenta T, Hausmann G and Basler K: The
many faces and functions of β-catenin. EMBO J. 31:2714–2736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guigon CJ, Kim DW, Zhu X, Zhao L and Cheng
SY: Tumor suppressor action of liganded thyroid hormone receptor
beta by direct repression of beta-catenin gene expression.
Endocrinology. 151:5528–5536. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Q, Dashwood WM, Zhong X, Al-Fageeh and
Dashwood RH: Cloning of the rat beta-catenin gene (Ctnnb1) promoter
and its functional analysis compared with the Catnb and CTNNB1
promoters. Genomics. 83:231–242. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu G, Jiang S, Wang C, et al: Zinc finger
transcription factor 191, directly binding to β-catenin promoter,
promotes cell proliferation of hepatocellular carcinoma.
Hepatology. 55:1830–1839. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu Y, Ye X, Zhang JB, et al: PROX1
promotes hepatocellular carcinoma proliferation and sorafenib
resistance by enhancing β-catenin expression and nuclear
translocation. Oncogene. 34:5524–5535. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mohamed MA, Greif PA, Diamond J, Sharaf O,
Maxwell P, Montironi R, Young RA and Hamilton PW: Epigenetic
events, remodelling enzymes and their relationship to chromatin
organization in prostatic intraepithelial neoplasia and prostatic
adenocarcinoma. BJU Int. 99:908–915. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Stopka T and Skoultchi AI: The ISWI ATPase
Snf2h is required for early mouse development. Proc Natl Acad Sci
USA. 100:14097–14102. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Reisman D, Glaros S and Thompson EA: The
SWI/SNF complex and cancer. Oncogene. 28:1653–1668. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jin Q, Mao X, Li B, Guan S, Yao F and Jin
F: Overexpression of SMARCA5 correlates with cell proliferation and
migration in breast cancer. Tumour Biol. 36:1895–1902. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Stopka T, Zakova D, Fuchs O, Kubrova O,
Blafkova J, Jelinek J, Necas E and Zivny J: Chromatin remodeling
gene SMARCA5 is dysregulated in primitive hematopoietic cells of
acute leukemia. Leukemia. 14:1247–1252. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gigek CO, Lisboa LC, Leal MF, Silva PN,
Lima EM, Khayat AS, Assumpção PP, Burbano RR and Smith Mde A:
SMARCA5 methylation and expression in gastric cancer. Cancer
Invest. 29:162–166. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cui J, Zhou X, Liu Y and Tang Z: Mutation
and overexpression of the beta-catenin gene may play an important
role in primary hepatocellular carcinoma among Chinese people. J
Cancer Res Clin Oncol. 127:577–581. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lai TY, Su CC, Kuo WW, Yeh YL, Kuo WH,
Tsai FJ, Tsai CH, Weng YJ, Huang CY and Chen LM: β-catenin plays a
key role in metastasis of human hepatocellular carcinoma. Oncol
Rep. 26:415–422. 2011.PubMed/NCBI
|
|
20
|
Farazi PA and DePinho RA: Hepatocellular
carcinoma pathogenesis: From genes to environment. Nat Rev Cancer.
6:674–687. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen L, Chan TH, Yuan YF, et al: CHD1L
promotes hepatocellular carcinoma progression and metastasis in
mice and is associated with these processes in human patients. J
Clin Invest. 120:1178–1191. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Y, Zhang JB, Qin Y, et al: PROX1
promotes hepatocellular carcinoma metastasis by way of
up-regulating hypoxia-inducible factor 1α expression and protein
stability. Hepatology. 58:692–705. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chuma M, Sakamoto M, Yasuda J, Fujii G,
Nakanishi K, Tsuchiya A, Ohta T, Asaka M and Hirohashi S:
Overexpression of cortactin is involved in motility and metastasis
of hepatocellular carcinoma. J Hepatol. 41:629–636. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lau SH, Sham JS, Xie D, Tzang CH, Tang D,
Ma N, Hu L, Wang Y, Wen JM, Xiao G, Zhang WM, et al: Clusterin
plays an important role in hepatocellular carcinoma metastasis.
Oncogene. 25:1242–1250. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun CK, Ng KT, Sun BS, Ho JW, Lee TK, Ng
I, Poon RT, Lo CM, Liu CL, Man K and Fan ST: The significance of
proline-rich tyrosine kinase2 (Pyk2) on hepatocellular carcinoma
progression and recurrence. Br J Cancer. 97:50–57. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tung-Ping Poon R, Fan ST and Wong J: Risk
factors, prevention, and management of postoperative recurrence
after resection of hepatocellular carcinoma. Ann Surg. 232:10–24.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang W, Lu Y, Xu Y, Xu L, Zheng W, Wu Y,
Li L and Shen P: Estrogen represses hepatocellular carcinoma (HCC)
growth via inhibiting alternative activation of tumor-associated
macrophages (TAMs). J Biol Chem. 287:40140–40149. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang WL, Wei L, Huang WQ, Li R, Shen WY,
Liu JY, Xu JM, Li B and Qin Y: Vigilin is overexpressed in
hepatocellular carcinoma and is required for HCC cell proliferation
and tumor growth. Oncol Rep. 31:2328–2334. 2014.PubMed/NCBI
|
|
30
|
Jiang YF, Yang ZH and Hu JQ: Recurrence or
metastasis of HCC: Predictors, early detection and experimental
antiangiogenic therapy. World J Gastroenterol. 6:61–65. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schmidt-Ott KM and Barasch J:
WNT/beta-catenin signaling in nephron progenitors and their
epithelial progeny. Kidney Int. 74:1004–1008. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Corona DF, Längst G, Clapier CR, Bonte EJ,
Ferrari S, Tamkun JW and Becker PB: ISWI is an ATP-dependent
nucleosome remodeling factor. Mol Cell. 3:239–245. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
VargaWeisz PD and Becker PB:
Chromatin-remodeling factors: Machines that regulate? Curr Opin
Cell Biol. 10:346–353. 1998. View Article : Google Scholar : PubMed/NCBI
|