Introduction
As a leading cause of mortality, arteriosclerosis
presents a significant risk to human health. It has been
responsible for 16,000,000 heart attacks and 5,800,000 strokes in
2008 in the USA. Although advances have been achieved in medicine,
the prevalence of arteriosclerosis remains (1). Studies have revealed that
arteriosclerosis is characterized by narrowed lumen filled with
thickened smooth muscle cells, which hinders the circulation of
blood flow (1). These vascular smooth
muscle cells (VSMCs) are believed to translocate from the tunica
media to the tunica intima, following rapid and uncontrolled
proliferation (2). Based on the above
knowledge, the suppression of VSMCs is considered to be a feasible
therapeutic strategy against arteriosclerosis.
Further studies have revealed that the activation of
specific molecular signaling pathways, including ERK, Notch, Wnt
and NF-κB, plays an key role in the aberrant proliferation of VSMCs
(3). Natural compounds that are
capable of interrupting the activation of these signaling pathways
have been demonstrated to exert an inhibitory effect on VSMC
proliferation. For instance, ligustrazine was demonstrated to
attenuate VSMC proliferation by suppressing ERK and p38 MAPK
signaling (4). Similarly, the natural
compound reinioside C suppresses the proliferation of smooth muscle
cells by inhibiting the NADPH oxidase-ROS-ERK1/2-NF-κB-AP-1 pathway
(5). Long non-coding RNA (lncRNA), a
type of nuclear RNA molecule that cannot be translated into a
protein product, has previously been confirmed to inhibit the
proliferation of arteriosclerosis-associated VSMCs (6). However, it is unclear whether lncRNA is
an effective therapeutic target for VSMC-directed
anti-arteriosclerosis treatment.
In this study, we observed that oleanolic acid (OA),
a natural compound widely distributed in a wide range of plants
with various verified bioactivities, exerts an inhibitory effect on
the proliferation of VSMCs in rats by increasing lincRNA-p21
expression. Our findings provide evidence that lncRNA mediates the
effect of the natural compound on VSMC biology.
Materials and methods
Primary rat smooth muscle cell
culture
Rat smooth muscle cells were used in the present
study, which was approved by the ethics committee of Qingdao
University (Qingdao, China). The isolation and culturing procedure
has been described previously (7).
PDGF-bb (30 ng/ml, PHG0041, Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) was added to the culture to stimulate the
proliferation of VSMCs.
Chemicals
OA was obtained from Sigma-Aldrich (St. Louis, MO,
USA) and preserved in dimethyl sulfoxide (DMSO). In the study, OA
(20, 40 or 80 µM) was added to the culture of VSMCs, and then the
proliferation rates of VSMCs were determined at the indicated time
points. In the control groups, DMSO was added to the culture.
Cell counting
The number of cells was determined with a Millipore
Scepter™ handheld automated cell counter (Merck Millipore,
Darmstadt, Germany) according to the instructions provided at the
indicated time points. The counting was performed at least three
times. The bars represent the means ± standard deviation.
Bromodeoxyuridine (BrdU) assay
BrdU assay was conducted to detect the proliferation
rates of VSMCs. The protocols are briefly described as follows.
Culture medium was removed and replaced with BrdU labeling solution
at 37°C for 2 h. Then the labeling solution was removed and cells
were washed twice with phosphate-buffered saline (PBS). One
milliliter of 3.7% formaldehyde was used to fix the cells for 15
min at room temperature, then cells were washed twice with PBS.
Next, 1 ml Triton® X-100 permeabilization buffer was
added to each well and the cells were incubated for 20 min at room
temperature. One milliliter of antibody staining buffer was
subsequently added to each well, and anti-BrdU primary antibody was
used to treat cells overnight at room temperature. Fluorescently
labeled secondary antibody was then added and the cells were
incubated for 1 h. Finally, the cells were observed under an
IX71-A12FL/PH microscope (Olympus Corporation, Tokyo, Japan).
BrdU+ cells were counted and recorded for the
calculation of percentages.
lncRNA detection
Quantitative polymerase chain reaction (qPCR) was
performed to detect the lincRNA-p21 expression level. Total RNA was
extracted from the cells with TRIzol solution (Sigma-Aldrich).
Following reverse transcription reaction, qPCR was performed using
TaqMan® 2X Universal PCR master mix (Applied Biosystems;
Thermo Fisher Scientific, Inc.) on a CFX96™ Real-Time PCR detection
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA) supplied
with analytical software. The primers used are as follows: forward:
TCCTTCTTGTGGTTGTGACT; reverse: GGGCTCAAGGGATCGGCCTG.
Immunoblot assays
Total protein was extracted from cells, followed by
1% polyacrylamide gel electrophoresis-based separation. After 0.45
µm membrane transferring and 5% milk blocking, samples were
incubated with the following primary antibodies: anti-p-IKKβ, 1:500
(BD Biosciences, San Jose, CA, USA); anti-total IKKβ antibody,
1:1,000 (BD Biosciences); anti-IκBα antibody, 1:1,000 (Cell
Signaling Technology, Inc., Danvers, MA, USA; anti-IκBβ antibody,
1:1,000 (Cell Signaling Technology); anti-p65 antibody, 1:1,000
(Cell Signaling Technology), and subsequently appropriate secondary
antibodies. Following the treatment of substrates, the bands were
visualized using a Tanon-4200 imaging system (Tiangen, China).
Statistical analysis
Statistical analysis in our study was performed
using a two-tailed Student's t-test. P<0.05 and P<0.01 were
considered to indicate a significant and a very significant
statistical difference between two groups, respectively.
Results
Oleanolic acid suppresses
proliferation of VSMCs in a dose- and time-dependent manner
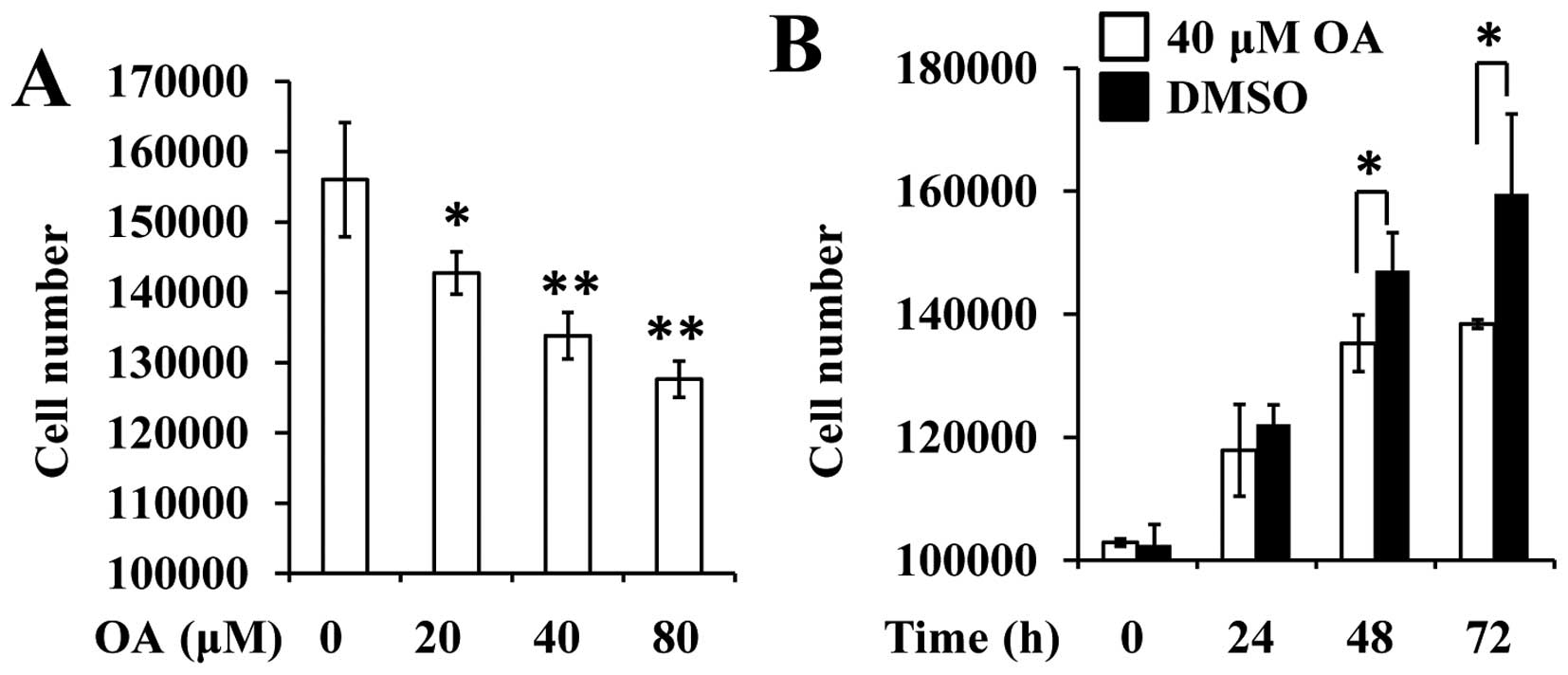
First, we employed cell counting and BrdU assay to
detect the effect of OA on rat VSMCs. The results revealed that the
number of VSMCs was significantly reduced 48 h after treatment with
the various concentrations of OA (Fig.
1A). Similarly, 40 µM OA was observed to decrease the number of
cells at the indicated time points (24, 48 and 72 h, Fig. 1B).
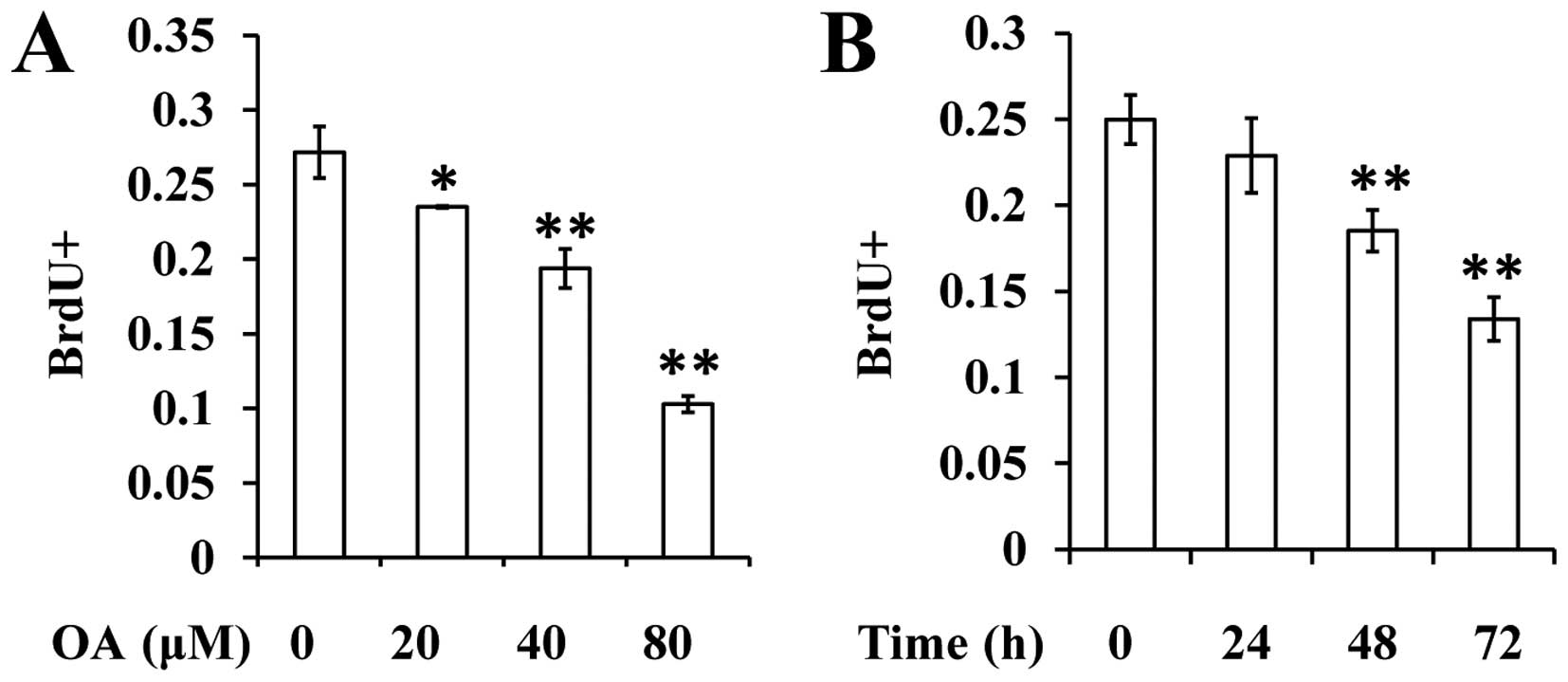
BrdU assay was conducted to determine the effect of
OA on the proliferation of VSMCs. The results revealed that OA is
able to decrease the percentage of BrdU+ cells in a
dose- and time-dependent manner (Fig. 2A
and B).
LincRNA-p21 expression is restored in
VSMCs by OA
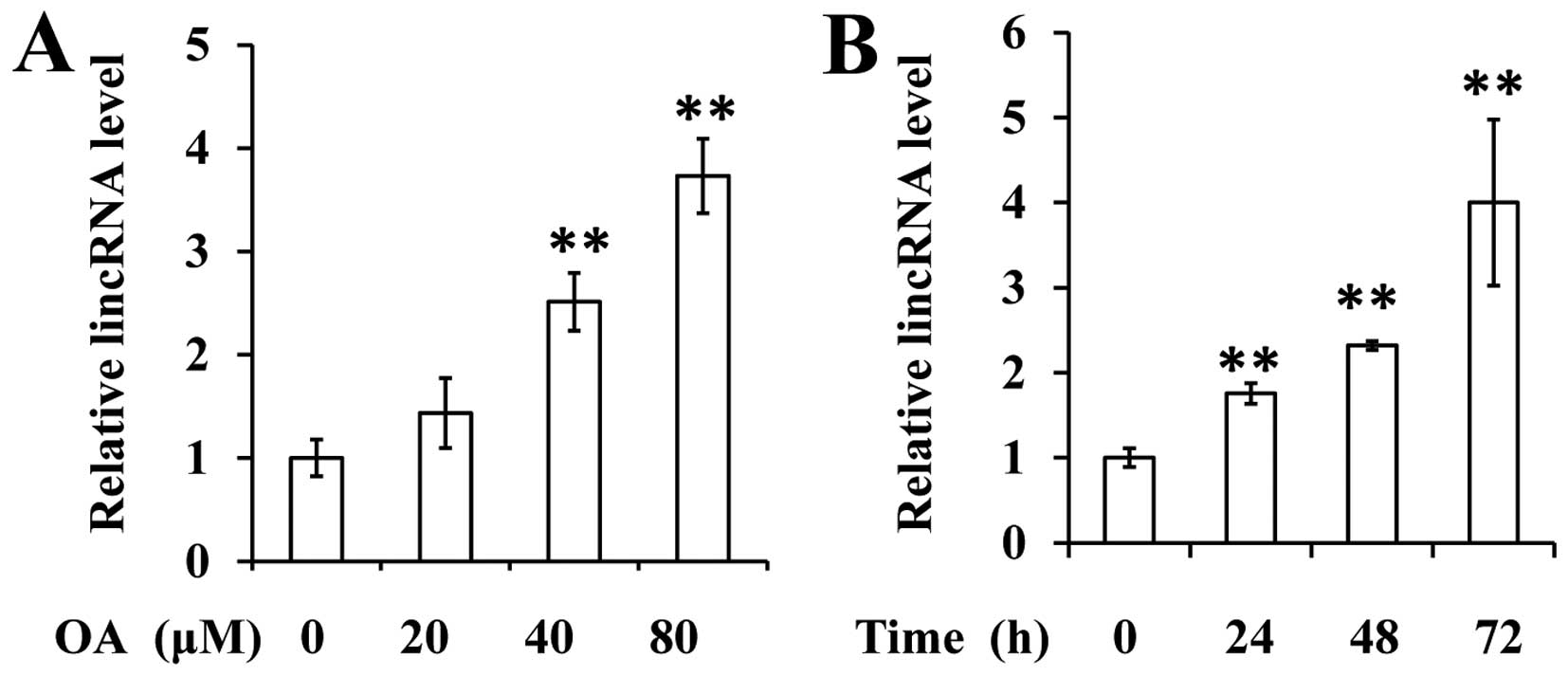
Given the fact that lincRNA-p21 downregulation
contributes to the uncontrolled proliferation of VSMCs, we aimed to
elucidate whether OA affected the expression of lincRNA-p21. The
data revealed that OA significantly increases the expression of
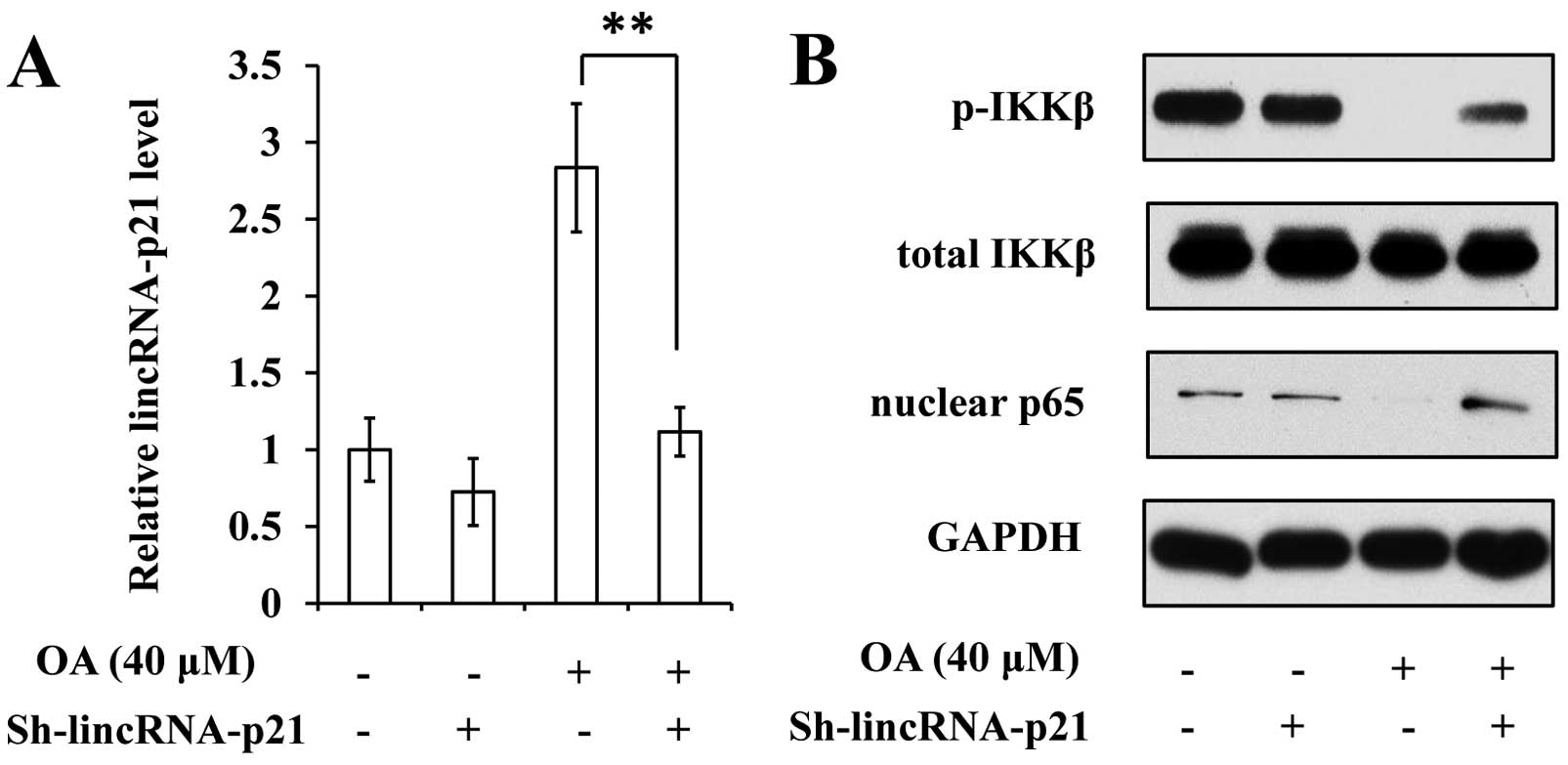
lincRNA-p21 in VSMCs in a dose- and time-dependent manner (Fig. 3A and B).
NF-κB signaling is suppressed in VSMCs
by OA
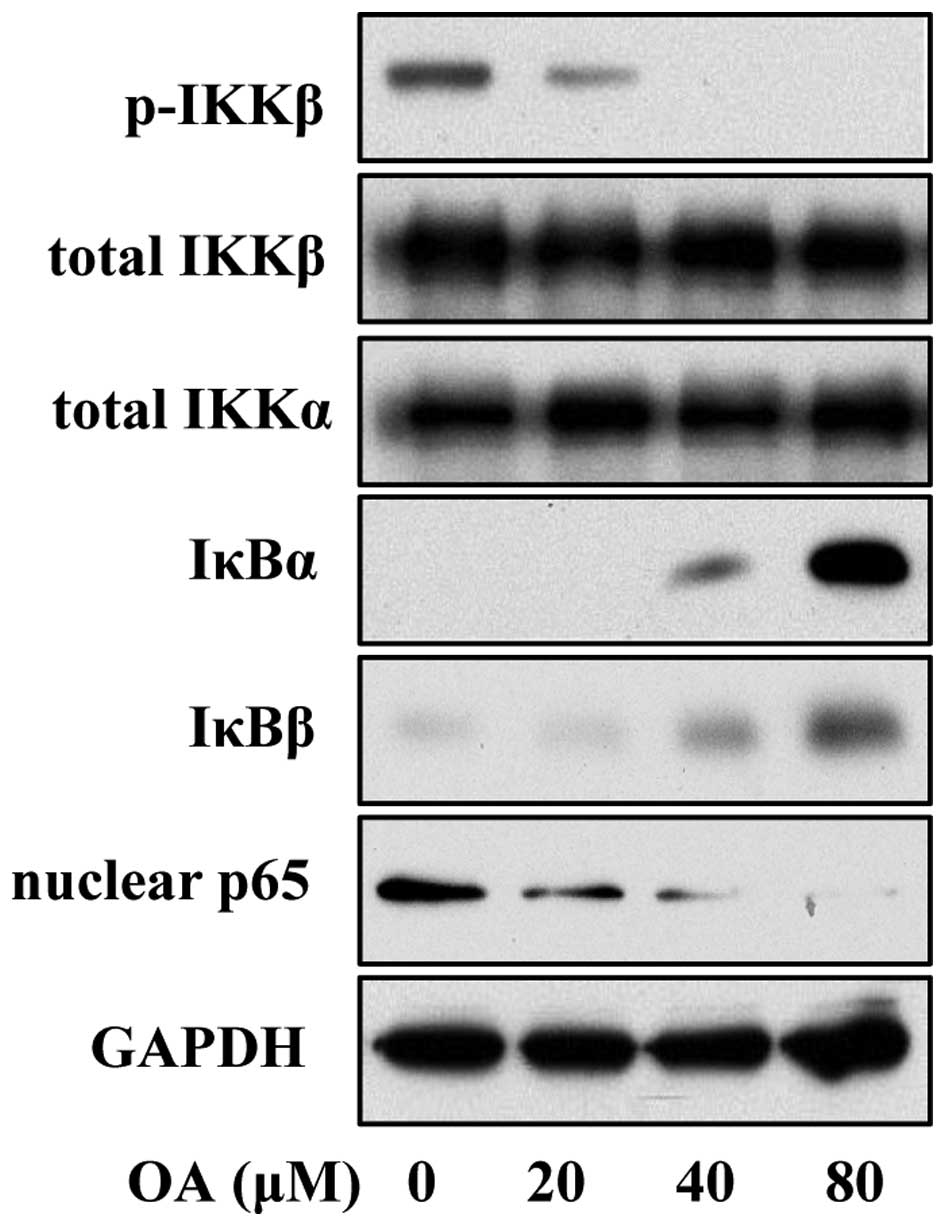
A previous study reported that the lincRNA-p21 level
is inversely correlated with the activation of NF-κB in rheumatoid
arthritis, implying a regulatory effect of lincRNA-p21 on NF-κB
signaling (8). Thus, we employed
immunoblot analysis to determine the activation of the NF-κB
pathway in OA-treated VSMCs. The data revealed that OA treatment
induced lower expression of phosphorylated IKK, increased the level
of IκBβ and reduced the abundance of nuclear p65, suggesting that
OA suppressed the activation of the NF-κB pathway (Fig. 4).
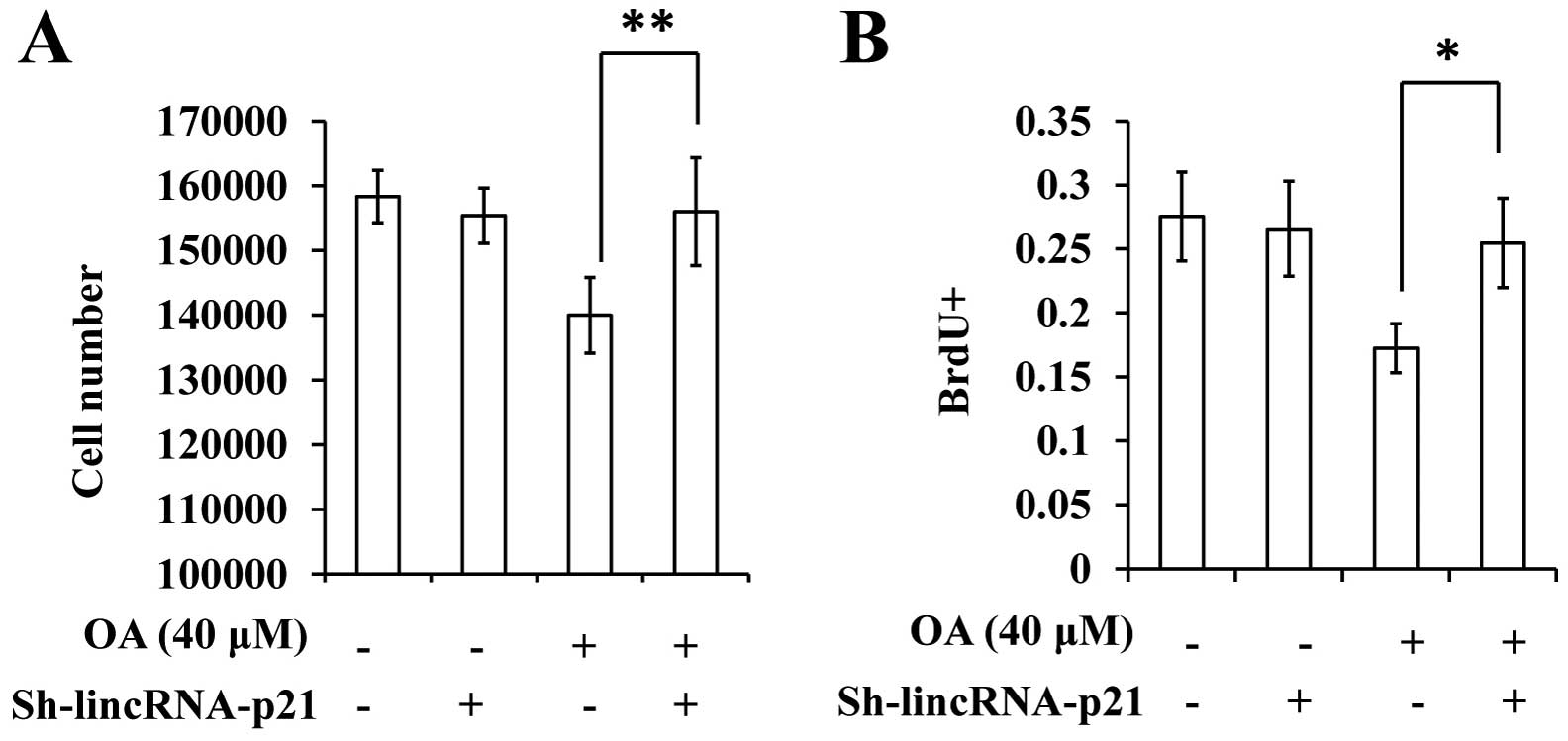
Suppression of lincRNA-p21 rescues the
effect of OA on the proliferation of VSMCs
Given the finding that OA treatment led to increased
expression of lincRNA-p21, we subsequently aimed to investigate the
significance of this event for the inhibitory effect of OA on VSMC
proliferation. We infected cells with a lentiviral vector
expressing shRNA targeting lincRNA-p21 (sh-lincRNA-p21; Fig. 5A). Following this treatment, NF-κB
signaling in VSMCs was observed to resist the effect of OA
(Fig. 5B). The suppression of
lincRNA-p21 appeared to eliminate the inhibitory effect of OA on
the proliferation of VSMCs (Fig. 6A and
B).
Discussion
OA has been well documented to exert various
bioactivities. Its effect on VSMCs was studied by Feng et al
(9). The ERK/Nrf2/HO-1 signaling
pathway was confirmed to mediate the protective effect of OA on
H2O2-induced oxidative damage in VSMCs.
However, the effect of OA on the proliferation of VSMCs had not
been studied previously. In this study, we provided evidence that
OA reduces the proliferation rates of rat VSMCs, which play a
significant role in the progression of arteriosclerosis.
Although the aberrant activation of certain
molecular signaling pathways has been established to be crucial for
arteriosclerosis, a previous study added lncRNA to the major
regulators for the onset and progression of this vascular disease
(6). lincRNA-p21, which is
underexpressed in arteriosclerosis patients, suppresses the
proliferation of VSMCs by promoting the transcription of numerous
p53 targets. Restoring p53 expression has been demonstrated to
ameliorate the severity of arteriosclerosis. However, it is unclear
whether lincRNA-p21 is an effective therapeutic target for
arteriosclerosis. Our study provided evidence that lincRNA-p21
mediates the effect of OA on the proliferation of VSMCs. VSMC
proliferation is assumed to contribute to the progression of
arteriosclerosis, implying that lincRNA-p21 may be a promising
target for anti-arteriosclerosis therapy.
The significance of the restoration of lincRNA-p21
by OA is not limited to the treatment of cardiovascular diseases.
Cancer, which is the second highest cause of human mortality, is
resistant to chemotherapy due to the low specificity and numerous
side effects of current antitumor drugs. A number of natural
compounds, including OA, have been demonstrated to suppress tumor
growth without significant side effects (10–14). OA is
therefore considered to be a promising compound in the clinical
application of cancer treatment. The association between
lincRNA-p21 and cancer has also been studied previously (15). LincRNA-p21 suppresses the malignant
phenotypes of cancer through multiple pathways (16–18). More
notably, lincRNA-p21 may be used to determine the clinical
prognosis of colon cancers (19).
However, it has not been determined whether lincRNA-p21 is
associated with the antitumor activity of OA. Further studies
should be conducted in order to address this issue.
Taken together, we observed that OA suppresses the
proliferation of VSMCs by increasing the expression of lincRNA-p21.
Our findings indicated that targeting lincRNA-p21 may be an
effective strategy for anti-arteriosclerosis treatment, and that OA
is a candidate compound for arteriosclerosis therapy.
References
|
1
|
Sun AS and Renaud M: Enhancement of
5′-nucleotidase activity of human leukemic cells after
fractionation: implications for cancer and aging. Mutat Res.
219:295–302. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chaabane C, Coen M and Bochaton-Piallat
ML: Smooth muscle cell phenotypic switch: implications for foam
cell formation. Curr Opin Lipidol. 25:374–379. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rudijanto A: The role of vascular smooth
muscle cells on the pathogenesis of atherosclerosis. Acta Med
Indones. 39:86–93. 2007.PubMed/NCBI
|
|
4
|
Yu L, Huang X, Huang K, Gui C, Huang Q and
Wei B: Ligustrazine attenuates the platelet-derived growth
factor-BB-induced proliferation and migration of vascular smooth
muscle cells by interrupting extracellular signal-regulated kinase
and P38 mitogen-activated protein kinase pathways. Mol Med Rep.
12:705–711. 2015.PubMed/NCBI
|
|
5
|
Hong D, Bai YP, Shi RZ, Tan GS, Hu CP and
Zhang GG: Inhibitory effect of reinioside C on vascular smooth
muscle cells proliferation induced by angiotensin II via inhibiting
NADPH oxidase-ROS-ENK1/2-NF-kappaB-AP-1 pathway. Pharmazie.
69:698–703. 2014.PubMed/NCBI
|
|
6
|
Wu G, Cai J, Han Y, Chen J, Huang ZP, Chen
C, Cai Y, Huang H, Yang Y, Liu Y, et al: LincRNA-p21 regulates
neointima formation, vascular smooth muscle cell proliferation,
apoptosis, and atherosclerosis by enhancing p53 activity.
Circulation. 130:1452–1465. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qin X, Qiu C and Zhao L: Maslinic acid
protects vascular smooth muscle cells from oxidative stress through
Akt/Nrf2/HO-1 pathway. Mol Cell Biochem. 390:61–67. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Spurlock CF III, Tossberg JT, Matlock BK,
Olsen NJ and Aune TM: Methotrexate inhibits NF-κB activity via long
intergenic (noncoding) RNA-p21 induction. Arthritis Rheumatol.
66:2947–2957. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Feng J, Zhang P, Chen X and He G: PI3K and
ERK/Nrf2 pathways are involved in oleanolic acid-induced heme
oxygenase-1 expression in rat vascular smooth muscle cells. J Cell
Biochem. 112:1524–1531. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu J, Zheng L, Ma L, Wang B, Zhao Y, Wu
N, Liu G and Lin X: Oleanolic acid inhibits proliferation and
invasiveness of Kras-transformed cells via autophagy. J Nutr
Biochem. 25:1154–1160. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu J, Wu N, Ma LN, Zhong JT, Liu G, Zheng
LH and Lin XK: p38 MAPK signaling mediates mitochondrial apoptosis
in cancer cells induced by oleanolic acid. Asian Pac J Cancer Prev.
15:4519–4525. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu J, Zheng L, Zhong J, Wu N, Liu G and
Lin X: Oleanolic acid induces protective autophagy in cancer cells
through the JNK and mTOR pathways. Oncol Rep. 32:567–572.
2014.PubMed/NCBI
|
|
13
|
Liu J, Zheng L, Wu N, Ma L, Zhong J, Liu G
and Lin X: Oleanolic acid induces metabolic adaptation in cancer
cells by activating the AMP-activated protein kinase pathway. J
Agric Food Chem. 62:5528–5537. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu J, Wu N, Ma L, Liu M, Liu G, Zhang Y
and Lin X: Oleanolic acid suppresses aerobic glycolysis in cancer
cells by switching pyruvate kinase type M isoforms. PLoS One.
9:e916062014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gutschner T and Diederichs S: The
hallmarks of cancer: A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang F, Zhang H, Mei Y and Wu M:
Reciprocal regulation of HIF-1α and lincRNA-p21 modulates the
Warburg effect. Mol Cell. 53:88–100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang G, Li Z, Zhao Q, Zhu Y, Zhao C, Li X,
Ma Z, Li X and Zhang Y: LincRNA-p21 enhances the sensitivity of
radiotherapy for human colorectal cancer by targeting the
Wnt/β-catenin signaling pathway. Oncol Rep. 31:1839–1845.
2014.PubMed/NCBI
|
|
18
|
Dimitrova N, Zamudio JR, Jong RM, Soukup
D, Resnick R, Sarma K, Ward AJ, Raj A, Lee JT, Sharp PA and Jacks
T: LincRNA-p21 activates p21 in cis to promote Polycomb target gene
expression and to enforce the G1/S checkpoint. Mol Cell.
54:777–790. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhai H, Fesler A, Schee K, Fodstad O,
Flatmark K and Ju J: Clinical significance of long intergenic
noncoding RNA-p21 in colorectal cancer. Clin Colorectal Cancer.
12:261–266. 2013. View Article : Google Scholar : PubMed/NCBI
|