Introduction
Colorectal cancer (CRC) is a leading cause of cancer
mortality worldwide, with over 1.2 million new cases and more than
600,000 mortalities every year (1).
In China, the incidence has increased rapidly since the 1980s
(2,3).
Distant metastasis following surgery is the main cause of treatment
failure. There are few effective strategies to treat CRC once
first-line approaches have failed. Therefore, improved
understanding of the role and mechanism of signaling pathways
including Notch and Wnt in colorectal carcinogenesis is critical
for the development of novel therapeutics.
A hallmark of tumors is the alteration of signaling
pathways that control cellular differentiation during developmental
processes, including the Wnt, Notch and Hedgehog pathways (4). Notably, there is evidence that crosstalk
exists among these pathways at the molecular level, and the key
nodes of intersection may provide opportunities for effective
targeted therapies. Multiple intracellular signaling pathways
including Wnt/β-catenin signaling, epidermal growth factor
receptor/Ras signaling and Notch signaling play major roles and
have demonstrated crosstalk in intestinal development and
tumorigenesis (5). Notch and
Wnt/β-catenin play key roles not only in maintaining the growth and
proliferation of CRC but also in cellular drug resistance and
cancer recurrence by regulating colon cancer stem cells (6–8).
Therefore, understanding the effects of Wnt and Notch signaling
pathway inhibitors on tumor progression is essential.
In the present study, we investigated the effects of
the inhibitors of Wnt and Notch signaling pathway synergistically
on the proliferation, migration and cell cycle of HCT-116 cells,
and the effects on tumor growth in a transplantation tumor model.
We further studied the expression of molecules in these two
signaling pathways treated by inhibitors rDDK-1 and LY374973.
Materials and methods
Cell culture
Human HCT-116 cell lines were cultured in RPMI-1640
(Invitrogen Life Technologies, San Diego, CA, USA) supplemented
with 10% fetal bovine serum at 37°C and 5% of
CO2/air.
Proliferation assay, Transwell
migration assay and cell cycle assay
3-(4, 5-Dimethylthiazol-2-yl)-2,
5-diphenyltetrazolium bromide (MTT) assay was used to estimate the
cell viability. HCT-116 cells were seeded at an initial density of
5,000 cells per well in a flat-bottomed 96-well cell culture plate
and allowed to grow for 48 h in a humidified 5% CO2, 95%
air atmosphere in an incubator maintained at 37°C. Twenty
microliters of MTT (5 mg/ml) solution (Sigma Chemical Co., St.
Louis, MO, USA) were added to each well and then incubated for 4 h
at 37°C. After the media were removed, 200 µl dimethyl sulfoxide
was added to each well to dissolve the formazan formed. After 30
min incubation at room temperature, the plates were scanned with a
microplate reader that was set at 490 nm for measuring the
absorbance.
The migration of cells was assayed in Transwell cell
culture chambers with 6.5-mm diameter polycarbonate membrane
filters having an 8 µm pore size. Briefly, 4×104 cells
in 100 µl serum-free medium were added to the upper chamber of the
device, and the lower chamber was filled with 600 µl culture medium
with 20% fetal bovine serum. After 10 h of incubation at 37°C, the
cells of the lower chamber were analyzed.
HCT-116 cells at a density of 2×106
cells/well were placed in 12-well plates and incubated with
inhibitors of the Wnt and Notch signaling pathways for 48 h before
the cells were harvested by centrifugation. The cells were then
trypsinized, washed with phosphate-buffered saline (PBS) and
treated with 50 µg/ml cold propidium iodide solution for 30 min in
the dark. Flow cytometric analysis was performed on a FACSCalibur
instrument (Becton-Dickinson, San Jose, CA, USA). The percentage of
cells in the G0/G1, S and G2/M phases was determined by flow
cytometry.
Immunofluorescence and microscopy
Cells were cultivated on cover glass slides in
24-well culture plates. After washing with PBS four times, cells
were fixed in 4% paraformaldehyde for 30 min. All of the following
washing and incubation steps were carried out with PBS/Tween-20
(PBST) on a rocker: cells were washed for 5 min four times and
permeabilized with 0.05% saponin for 5 min. After another three
washing steps, cells were fixed with pre-chilled ice-cold 100%
acetone for 2 min on ice and immediately washed three times.
Blocking with 4% bovine serum albumin in PBST for 30 min was
followed by overnight incubation at 4°C with the primary antibody.
The next day cells were washed three times and incubated with the
secondary antibody for 1 h. For double-staining, the procedure was
repeated with the next antibodies. Cells were imaged with a
fluorescence microscope (Axiovert 40C; Carl Zeiss Microscopy, Inc.,
Thornwood, NY, USA).
Xenografts in BALB/c mice
A total of 24, 6–8-week-old, female BALB/c mice
(Center of Experimental Animal, Kunming Medical University,
Kunming, China) were subcutaneously injected with 1×106
HCT-116 cells/animal. All animals were housed under a controlled
room humidity (50±10%), and maintained under a 12-h light/dark
cycle with free access to water and food. Upon tumor formation, the
animals were divided into four groups: i) control group (n=6); ii)
rDDK-1 group (5 mg/kg, administered once weekly, all three times,
n=6); iii) LY374973 group (4 mg/kg, administered once weekly, all
three times, n=6); and iv) rDDK-1+LY374973 group (n=6). The study
was approved by the ethics committee of Guangdong General Hospital
(Guangzhou, China; approval number 2015268A).
Quantitative polymerase chain reaction
(qPCR)
qPCR was used to detect the mRNA expression of
molecules in the Wnt and Notch signaling pathways. The PCR
reactions were performed in a total volume of 20 µl, including 10
µl 2X Power SYBR®-Green PCR master mix (Applied
Biosystems, Warrington, UK), 2 µl cDNA (5 ng/µl) and 1 µl primer
mix (10 µM each). The PCR amplification and detection were carried
out using the LightCycler 480 II (Roche Applied Science, Basel,
Switzerland) as follows: an initial denaturation at 95°C for 10
min; 40 cycles of 95°C for 15 sec and 60°C for 1 min. The relative
gene expression was calculated using the comparative CT method
(9). The gene expression of the
target gene normalized to an endogenous reference (GAPDH) and
relative to the calibrator was given by the formula
2−ΔΔCq (10).ΔCq was
calculated by subtracting the average GAPDH Cq from the average Cq
of the gene of interest. The ratio defines the level of relative
expression of the target gene to that of GAPDH.
Western blot analysis
Cells were washed with ice-cold PBS and lysed in
RIPA buffer [50 mmol/l Tris (pH 7.5), 150 mmol/l NaCl, 1% NP-40,
0.5% sodium deoxycholate, 0.1% sodium dodecyl sulphate (SDS)]
containing phenylmethylsulfonyl fluoride (PMSF; 1 mmol/l) and
protease inhibitors (2 g/ml; Protease inhibitor cocktail set III,
Calbiochem, Billerica, MA, USA) on ice for 30 min. The lysates were
clarified by centrifugation at 13,000 × g for 30 min at 4°C.
The total protein concentration was estimated using a Protein Assay
kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Proteins were
separated by sodium dodecyl sulphate-polyacrylamide gel,
transferred to polyvinylidene difluoride membranes (EMD Millipore,
Billerica, MA, USA), blocked and probed with antibodies against
β-catenin (1:1,000; sc-65480; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA), c-myc (1:1,000; sc-40; Santa Cruz Biotechnology,
Inc.), Jagged (1:1,000; sc-390177; Santa Cruz Biotechnology, Inc.),
Notch1 (1:1,000; sc-373891; Santa Cruz Biotechnology, Inc.), Notch2
(1:1,000; sc-5545; Santa Cruz Biotechnology, Inc.), DLL4 (1:1,000;
ab7280; Abcam, Cambridge, MA, USA) and Pra-1 (1:1,000; ab76413;
Abcam). Upon washing, blots were incubated with horseradish
peroxidase-conjugated secondary antibodies and visualized by super
enhanced chemiluminescence detection reagent (Applygen, Beijing,
China).
Statistical analysis
Statistical analyses were conducted using Student's
t-tests with the statistical software SPSS 15.0 (SPSS, Inc.,
Chicago, IL, USA). A corresponding two-sided P-value <0.05 was
considered to indicate a statistically significant difference.
Results
Effect of inhibition of Wnt and Notch
signaling pathway on proliferation, migration and cell cycle of
HCT-116 cells
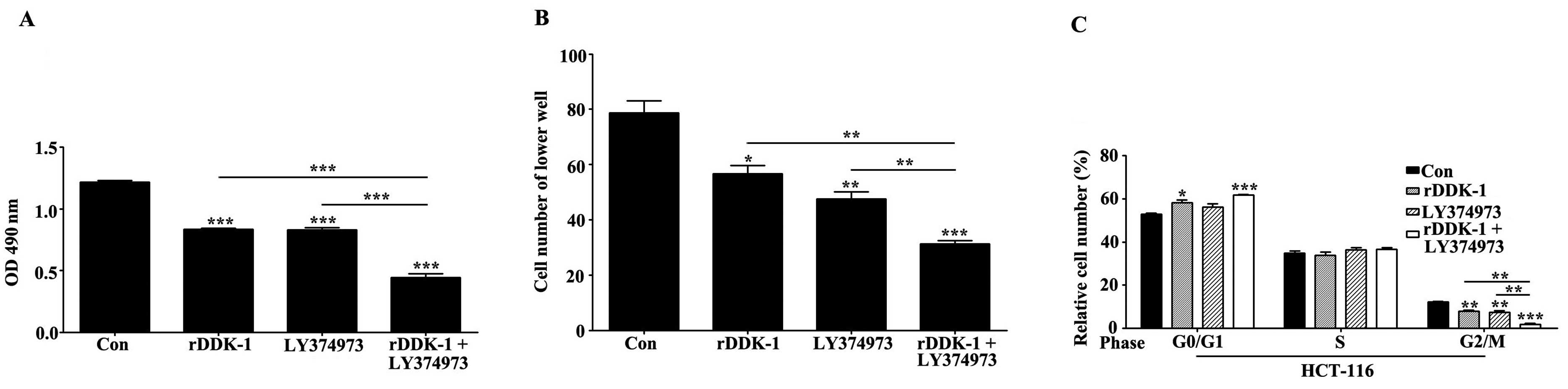
To investigate the effects of inhibition of the Wnt
and Notch signaling pathway on the proliferation, migration and
cell cycle of HCT-116 cells, we used the specific inhibitor rDDK-1
for the Wnt signaling pathway and LY374973 for the Notch signaling
pathway. We observed that rDDK-1 and LY374973 reduced the
proliferation of HCT-116 cells when used separately, and
synergistically inhibited the proliferation (Fig. 1A). We also revealed that rDDK-1 and
LY374973 reduced the migration ability and percentage of G2/M phase
cells. rDDK-1 and LY374973 could further synergistically inhibit
migration and the G2/M percentage (Fig.
1B and C).
Effect of inhibitors rDDK-1 and
LY374973 on Wnt and Notch pathway molecules in HCT-116 cells
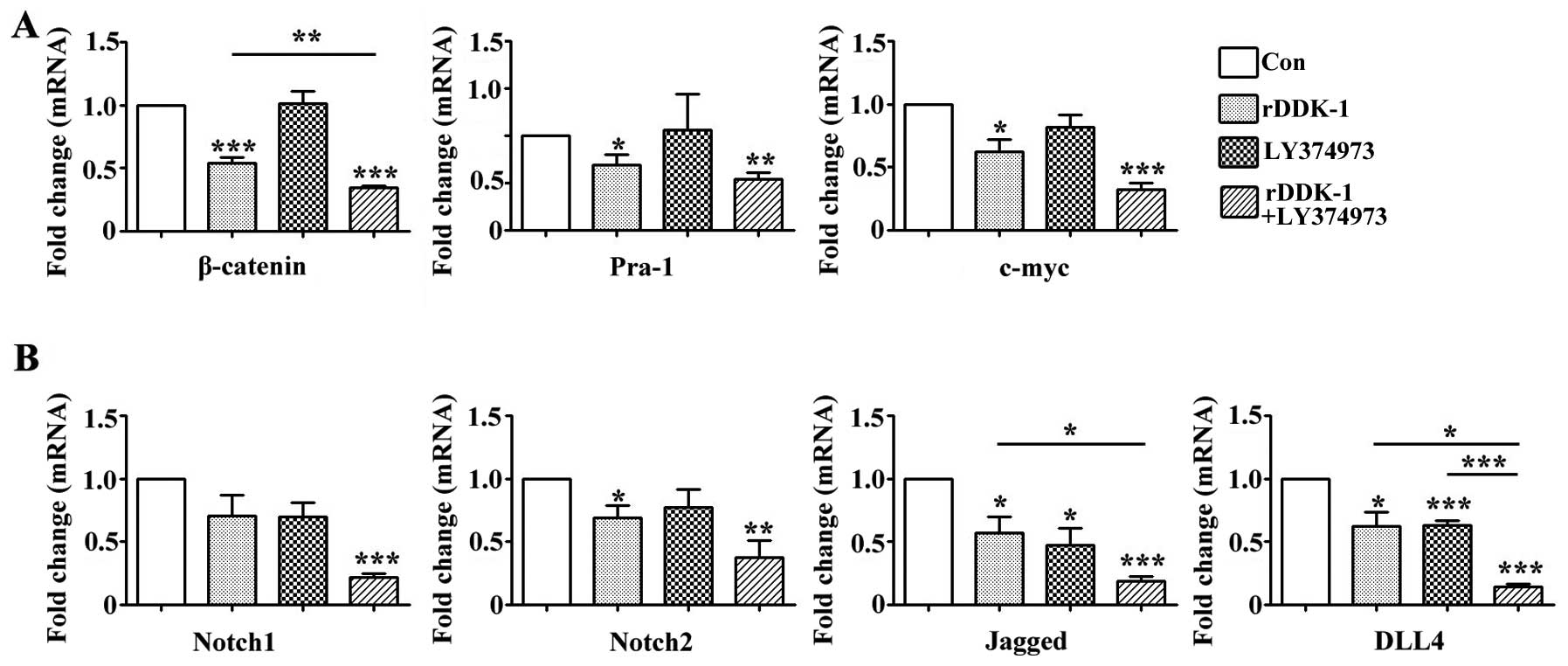
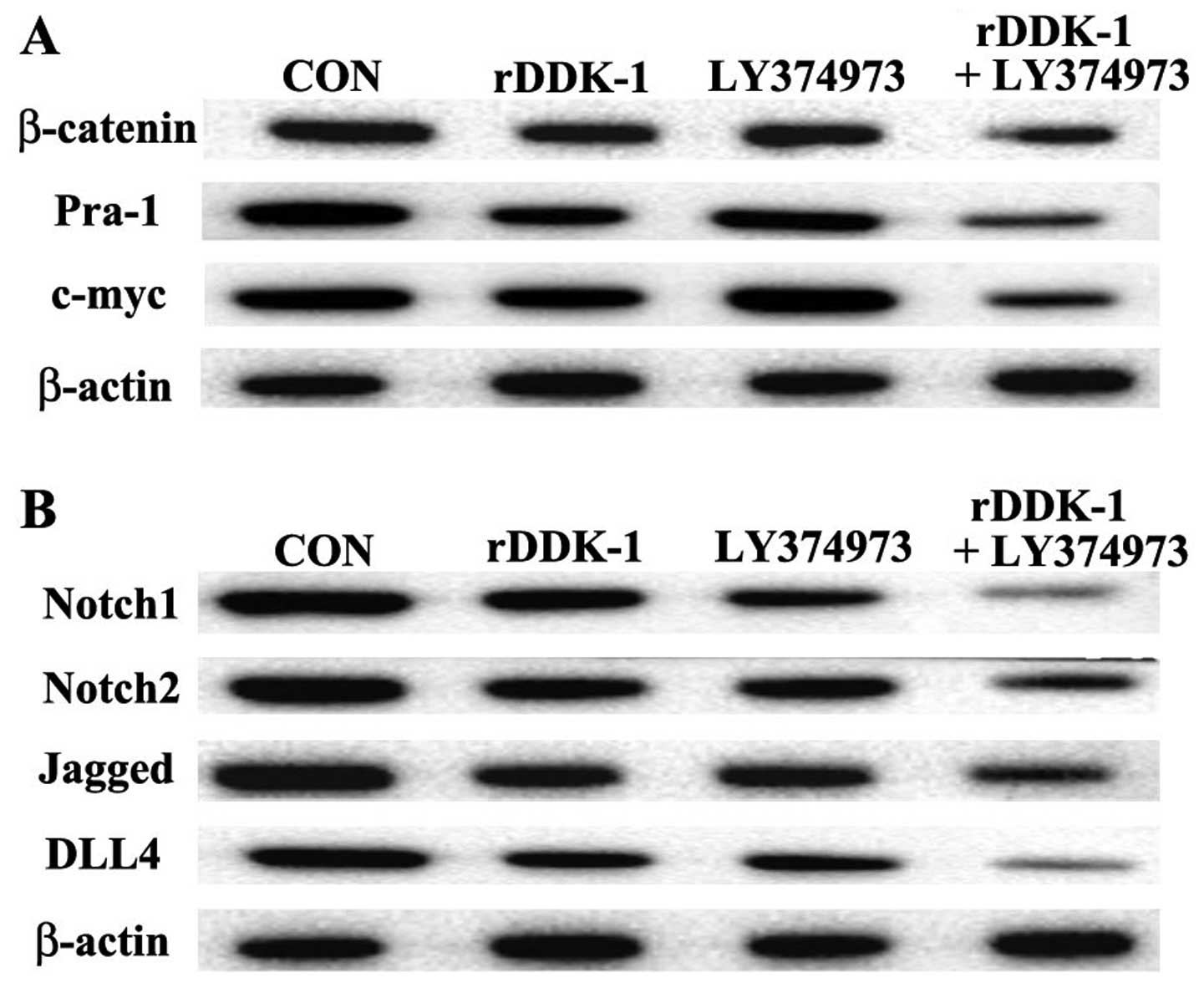
Our results revealed that rDDK-1 reduced the mRNA
and protein expression of Wnt signaling pathway molecules
β-catenin, Pra-1 and c-myc, and also inhibited the expression of
Notch signaling pathway genes Notch2, Jagged and DLL4 (Figs. 2 and 3).
LY374973 reduced the mRNA and protein expression level of Jagged
and DLL4, but had no effect on Wnt signaling pathway molecules
Pra-1 and c-myc (Figs. 2 and 3). With regard to the mRNA expression level,
LY374973 further increased the rDDK-1 inhibition of the mRNA
expression of β-catenin, Jagged and DLL4 but not Pra-1, c-myc,
Notch1 and Notch2 compared with rDDK-1 alone (Fig. 2). Inconsistently, with regard to the
protein expression level, LY374973 together with rDDK-1 reduced the
protein expression level of all detected Wnt and Notch pathway
genes (Fig. 3).
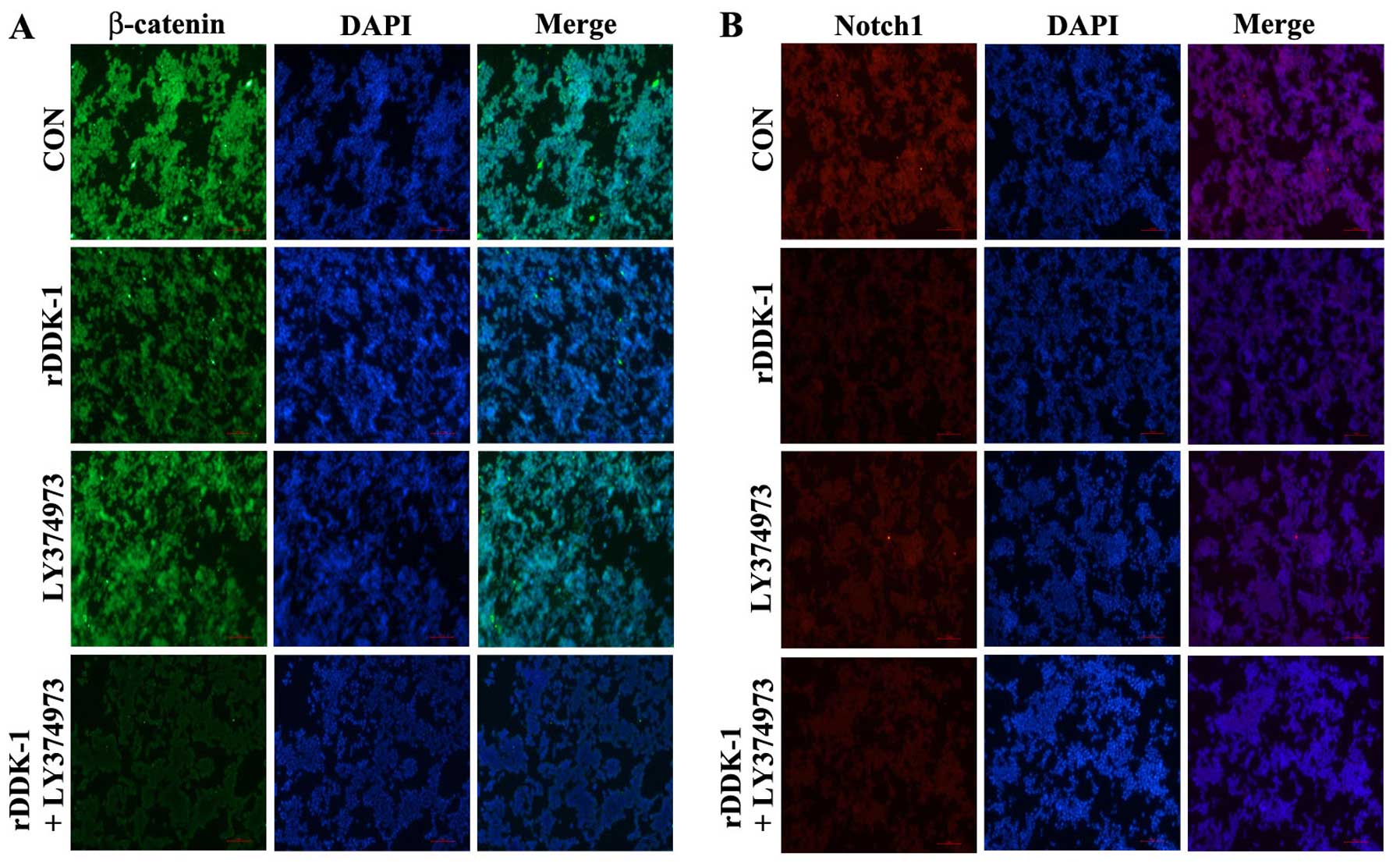
In the immunofluorescence assay, rDDK-1 was able to
inhibit the expression of β-catenin and Notch1, and LY374973
increased this effect on β-catenin. However, LY374973 could only
slightly inhibit the expression of Notch1 but not β-catenin
(Fig. 4).
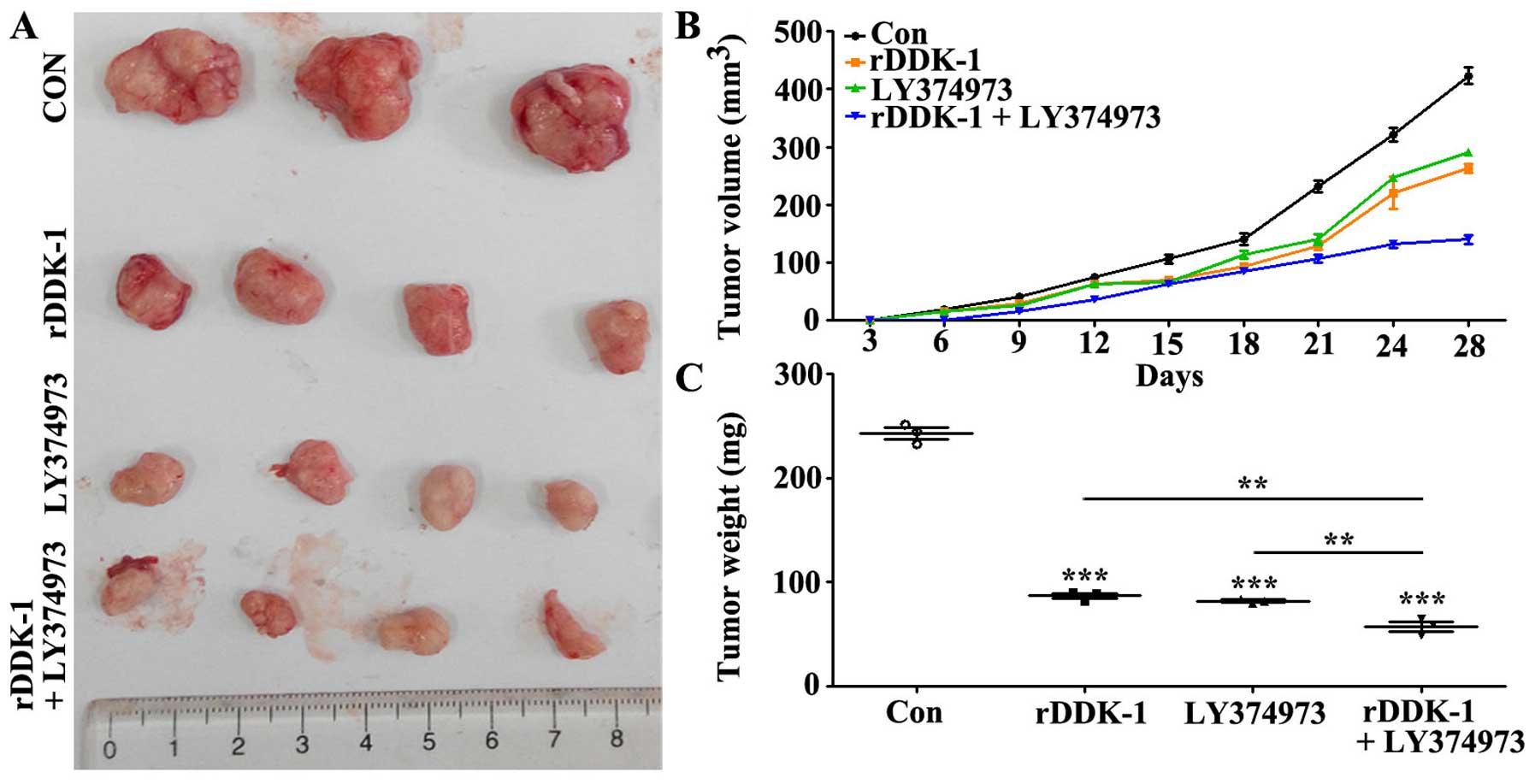
Effect of inhibition of Wnt and Notch
signaling pathway on tumor growth in transplantation tumor
model
We further investigated the effects of inhibition of
Wnt and Notch signaling pathway in a transplantation tumor model.
The results revealed that rDDK-1 and LY374973 decreased the tumor
volume and weight when used separately, and could further
synergistically inhibit the tumor volume and weight (Fig. 5B and C).
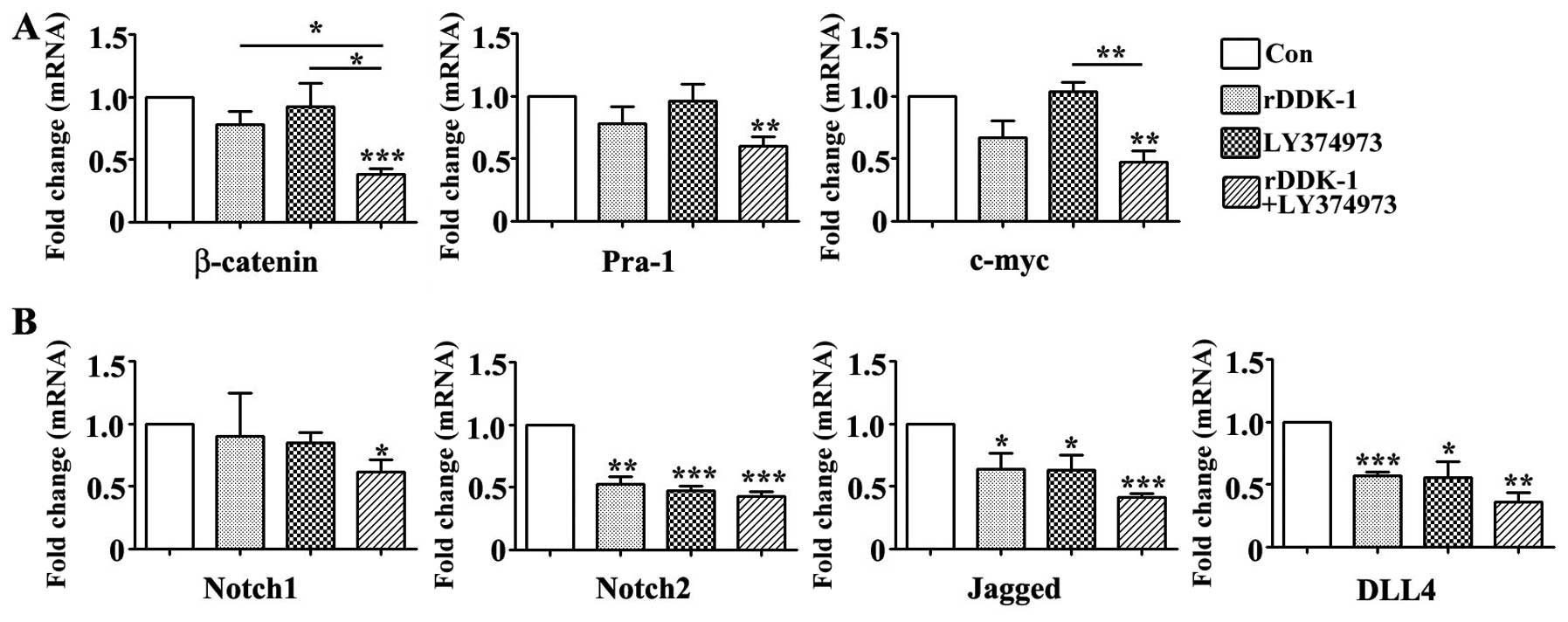
Effect of inhibitors rDDK-1 and
LY374973 on Wnt and Notch pathway molecules in transplantation
tumor model
Our results indicated that LY374973 could inhibit
the mRNA and protein expression of Notch2, Jagged and DLL4, but had
no effect on Wnt signaling pathway genes in tumor tissues (Figs. 6 and 7).
With regard to the protein expression level, rDDK-1 together with
LY374973 could further synergistically inhibit the expression level
of all detected Wnt and Notch pathway genes compared with rDDK-1 or
LY374973 alone (Fig. 7).
Discussion
Understanding the role and mechanism of signaling
pathways including Notch and Wnt in colorectal carcinogenesis is
likely to be critical for the development of novel therapeutics.
Previous studies revealed that Wnt/β-catenin and Notch signaling
play major roles and had crosstalk in intestinal development and
tumorigenesis (5). In addition,
increased Notch/Wnt signaling also promoted the early onset of
adenoma formation in APC-Cidn1 mice (11).
Previous studies have revealed that Wnt and Notch
signaling pathways worked together with transcription factors
including Slug, Snail and Twist to suppress E-cadherin, and finally
to increase tumor progression and migration (12). Gopalakrishnan et al reported
that β-catenin interacted significantly with Notch intracellular
domain in adenoma and adenocarcinoma compared with normal tissues.
This interaction activated CyclinD1 and Hes1 and finally promoted
cell proliferation (13).
Membrane-bound Notch is physically associated with unphosphorylated
β-catenin in stem and colon cancer cells and negatively regulates
post-translational accumulation of β-catenin protein by altering
the endocytic adaptor protein Numb and lysosomal activity (14). Bordonaro et al revealed that
the Notch ligand Delta-like 1 augmented the activity of the Wnt
signaling pathway and transcriptionally upregulated the connective
tissue growth factor gene and promoted cell growth in colon cancer
(15). Zhu et al divided CRC
into three transcriptional subtypes, and identified driver networks
or pathways for each group. Genomic alterations in the Wnt
signaling pathway were common among all three subtypes, however.
Unique combinations of pathway alterations including Wnt, VEGF and
Notch drove distinct molecular and clinical phenotypes in different
CRC subtypes (16). Prasetyanti et
al observed that Notch signals coordinated self-renewal and
lineage determination not only in normal cells, but also at the
adenoma and carcinoma stage in humans and mice (17). Notably, the Wnt pathway exhibited a
heterogeneous activity pattern that determined stemness in all
stages of disease, whereas it was previously predicted to be
constitutively active in adenomas and carcinomas.
Notch and Wnt/β-catenin signaling also intersect in
stem and progenitor cells and regulate each other
transcriptionally. Notch and Wnt signaling function together to
regulate colonic progenitor cell division and differentiation.
Reedijk et al reported that Notch signaling was required for
adenoma formation in response to elevated Wnt pathway signaling
that occurred in an APCMin mouse model of human adenomatous
polyposis coli (18). DLL4 and JAG1
were the Notch ligand genes. Katoh et al observed that JAG1
was widely expressed in a number of cancers including colon cancer,
head and neck cancer and gastric cancer. JAG1 was a Wnt-dependent
Notch signaling activator and was the key molecule maintaining the
homeostasis of stem and progenitor cells (19). Wnt and Notch pathways regulate the
self-renewal of normal stem cells (20).
Deregulation of Notch and Wnt signaling pathways
plays a significant role in normal and cancer stem cells (CSCs)
(21). These signaling pathways that
are involved in proliferation and maintenance of CSCs lead to the
development of CRC (22). A number of
promising targets including Wnt/β-catenin and Notch signaling have
been identified as useful targets to prevent or therapeutically
inhibit CRC development (1,23). Singh et al revealed that green
tea catechin epigallocatechin-3-gallate blocks carcinogenesis by
affecting a wide array of signal transduction pathways, including
Notch and Wnt (24).
Okuhashi et al reported that the Notch
inhibitors γ-secretase inhibitors together with the Wnt inhibitor
quercetin (Qu) could suppress the growth of DND-41 T-cell acute
lymphoblastic leukemia cells synergistically, and that Qu treatment
reduced the levels of Notch1 protein and its active fragment in
DND-41 cells (24). However, there
were no reports about the synergistic anti-tumor effects of Notch
and Wnt inhibitors together in other types of tumors, including
CRC. Our study demonstrated that rDDK-1 (an inhibitor of the Wnt
signaling pathway) and LY374973 (an inhibitor of the Notch
signaling pathway) synergistically inhibited the proliferation,
migration and G2/M percentage of HCT-116 cell lines, and could
further synergistically inhibit the tumor volume and weight in a
transplantation tumor model. Our results may pave to way for using
inhibitors of the Wnt and Notch signaling pathways together to
treat CRC.
Further study should be conducted to explore the
mechanism of Wnt and Notch signaling pathway crosstalk in CRC and
to validate the effect of inhibition of the Wnt and Notch signaling
pathways in other animals.
References
|
1
|
Curtin JC: Novel drug discovery
opportunities for colorectal cancer. Expert Opin Drug Discov.
8:1153–1164. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lei T, Chen WQ, Zhang SW, Lei TH, Ying Q,
He ZY and Wang XH: Prevalence trend of colorectal cancer in 10
cities and counties in China from 1988 to 2002. Zhonghua Zhong Liu
Za Zhi. 31:428–433. 2009.(In Chinese). PubMed/NCBI
|
|
3
|
Li HL, Gao YT, Zheng Y, Zhang W, Gao LF,
Xu B and Xiang YB: Incidence trends of colorectal cancer in urban
Shanghai, 1973-2005. Zhonghua Yu Fang Yi Xue Za Zhi. 43:875–879.
2009.(In Chinese). PubMed/NCBI
|
|
4
|
Bertrand FE, Angus CW, Partis WJ and
Sigounas G: Developmental pathways in colon cancer: crosstalk
between WNT, BMP, Hedgehog and Notch. Cell Cycle. 11:4344–4351.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu WK, Wang XJ, Cheng AS, Luo MX, Ng SS,
To KF, Chan FK, Cho CH, Sung JJ and Yu J: Dysregulation and
crosstalk of cellular signaling pathways in colon carcinogenesis.
Crit Rev Oncol Hematol. 86:251–277. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Saif MW and Chu E: Biology of colorectal
cancer. Cancer J. 16:196–201. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mishra L, Banker T, Murray J, Byers S,
Thenappan A, He AR, Shetty K, Johnson L and Reddy EP: Liver stem
cells and hepatocellular carcinoma. Hepatology. 49:318–329. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ischenko I, Seeliger H, Schaffer M, Jauch
KW and Bruns CJ: Cancer stem cells: how can we target them? Curr
Med Chem. 15:3171–3184. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang G, Brennan C, Rook M, Wolfe JL, Leo
C, Chin L, Pan H, Liu WH, Price B and Makrigiorgos GM: Balanced-PCR
amplification allows unbiased identification of genomic copy
changes in minute cell and tissue samples. Nucleic Acids Res.
32:e762004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pope JL, Ahmad R, Bhat AA, Washington MK,
Singh AB and Dhawan P: Claudin-1 overexpression in intestinal
epithelial cells enhances susceptibility to adenamatous polyposis
coli-mediated colon tumorigenesis. Mol Cancer. 13:1672014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Umar S: Enteric pathogens and cellular
transformation: bridging the gaps. Oncotarget. 5:6573–6575. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gopalakrishnan N, Saravanakumar M,
Madankumar P, Thiyagu M and Devaraj H: Colocalization of β-catenin
with Notch intracellular domain in colon cancer: a possible role of
Notch1 signaling in activation of CyclinD1-mediated cell
proliferation. Mol Cell Biochem. 396:281–293. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kwon C, Cheng P, King IN, Andersen P,
Shenje L, Nigam V and Srivastava D: Notch post-translationally
regulates β-catenin protein in stem and progenitor cells. Nat Cell
Biol. 13:1244–1251. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bordonaro M, Tewari S, Atamna W and
Lazarova DL: The Notch ligand Delta-like 1 integrates inputs from
TGFbeta/Activin and Wnt pathways. Exp Cell Res. 317:1368–1381.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu J, Wang J, Shi Z, Franklin JL, Deane
NG, Coffey RJ, Beauchamp RD and Zhang B: Deciphering genomic
alterations in colorectal cancer through transcriptional
subtype-based network analysis. PLoS One. 8:e792822013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Prasetyanti PR, Zimberlin CD, Bots M,
Vermeulen L, Fde S Melo and Medema JP: Regulation of stem cell
self-renewal and differentiation by Wnt and Notch are conserved
throughout the adenoma-carcinoma sequence in the colon. Mol Cancer.
12:1262013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Reedijk M, Odorcic S, Zhang H, Chetty R,
Tennert C, Dickson BC, Lockwood G, Gallinger S and Egan SE:
Activation of Notch signaling in human colon adenocarcinoma. Int J
Oncol. 33:1223–1229. 2008.PubMed/NCBI
|
|
19
|
Katoh M and Katoh M: Notch ligand, JAG1,
is evolutionarily conserved target of canonical WNT signaling
pathway in progenitor cells. Int J Mol Med. 17:681–685.
2006.PubMed/NCBI
|
|
20
|
Korkaya H and Wicha MS: Selective
targeting of cancer stem cells: a new concept in cancer
therapeutics. BioDrugs. 21:299–310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Karamboulas C and Ailles L: Developmental
signaling pathways in cancer stem cells of solid tumors. Biochim
Biophys Acta. 1830:2481–2495. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roy S and Majumdar AP: Signaling in colon
cancer stem cells. J Mol Signal. 7:112012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pandurangan AK and Esa NM: Dietary
non-nutritive factors in targeting of regulatory molecules in
colorectal cancer: an update. Asian Pac J Cancer Prev.
14:5543–5552. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Singh BN, Shankar S and Srivastava RK:
Green tea catechin, epigallocatechin-3-gallate (EGCG): mechanisms,
perspectives and clinical applications. Biochem Pharmacol.
82:1807–1821. 2011. View Article : Google Scholar : PubMed/NCBI
|