Introduction
Acute lymphoblastic leukemia (ALL) is one of the
most malignant types of hematological disease, which accounts for
30% of all pediatric cancers (1).
Chemotherapeutic drugs commonly used for ALL treatment include
prednisone, 6-mercaptopurine, dexamethasone, cyclophosphamide,
l-asparaginase, vincristine, daunorubicin or doxorubicin,
methotrexate and cytarabine; with overall cure rates of 80%
(Fig. 1) (2). However, a significant number of patients
develop resistance to these drugs and outcome is poor among
patients who relapse (2). The
pathogenesis of ALL includes changes in gene expression, which are
regulated by diverse transcription factors (3) such as nuclear factor-kappa B (NF-κB),
which has been associated with cell proliferation and survival
(4). Additionally, in solid tumors,
NF-κB exhibits an important function in invasion, angiogenesis,
aggressive tumor growth and chemoresistance (4–6).
Constitutive activation of NF-κB is observed in ~92% of pediatric
ALL patients (7) and thus is one of
the targets for chemosensitization.
Since ~80% of clinical drugs are derived from
natural products, numerous compounds have been identified that
downmodulate NF-κB (8). Curcumin
(diferuloylmethane), a polyphenol derived from the plant Curcuma
longa, has been demonstrated to inhibit NF-κB activation, which
is induced by a wide variety of carcinogens and chemotherapeutic
agents (9). The use of curcumin was
classified by the USA Food and Drug Administration as ‘generally
recognized as safe’ (10).
Furthermore, various clinical trials indicate that curcumin may be
administered at oral doses as high as 8 g/day with no side effects
(11).
The aim of the present study was to investigate the
potential anticancer effect of curcumin on the human REH ALL cell
line, when administered alone and in combination with currently
used therapies. The results indicate that curcumin potentiates the
effect of chemotherapeutic agents against ALL cells by activation
of caspase-3 through downregulation of oxidative stress, NF-κB
activation and various NF-κB-regulated cell survival gene
products.
Materials and methods
Reagents
Curcumin and sodium dodecyl sulfate were obtained
from Sigma-Aldrich (St. Louis, MO, USA). Penicillin, streptomycin,
RPMI-1640 medium, phosphate-buffered saline (PBS), fetal bovine
serum (FBS) and TaqMan Assays for B-cell lymphoma (Bcl)-extra large
(xL), Bcl-2, cyclin D1, survivin, c-Myc and beta-glucuronidase
(GUSB) were obtained from Life Technologies (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). DNA and RNA extraction was
performed using QIAzol Lysis reagent and QIAmp Circulating Nucleic
Acid kit, which were obtained from Qiagen GmbH (Hilden, Germany).
Monoclonal anti-NF-κB (dilution, 1:50; cat. no. A88940),
anti-cluster of differentiation (CD)45 (dilution, 1:10; cat. nos.
IM2652U and IM0782U) and anti-caspase-3 (dilution, 1:50; cat. no.
A88950) antibodies, 7-aminoactinomycin D (7-AAD) viability dye,
PerFix EXPOSE Phospho-Epitopes Exposure kit and IntraPrep™
permeabilization reagent were obtained from Beckman Coulter, Inc.
(Brea, CA, USA). Anti-NF-κB p65 antibody (dilution, 1 µg; cat. no.
sc-8008) was obtained from Santa Cruz Biotechnology, Inc. (Dallas,
TX, USA). Highly sensitive 8-hydroxy-2′-deoxyguanosine (8-OHdG) was
supplied by the Japanese Institute for the Control of Aging
(Fukuroi, Japan). Prednisone, 6-mercaptopurine, dexamethasone,
cyclophosphamide, l-asparaginase, vincristine, daunorubicin,
doxorubicin, methotrexate and cytarabine were provided by PiSA
Farmacéutica (Guadalajara, México).
Cell culture
The REH cell line (St. Jude Children's Research
Hospital, Memphis, TN, USA) was cultured in RMPI-1640 medium
supplemented with 100 U/ml penicillin and 100 g/ml streptomycin
with 10% FBS in a humidified incubator at 37°C with an atmosphere
of 5% CO2. Next, 5×105 REH cells were treated
for 48 h in triplicate with 125 µg/ml prednisone, 250 µg/ml
6-mercaptopurine, 0.4 µg/ml dexamethasone, 50 µg/ml
cyclophosphamide, 5 U l-asparaginase, 25 µg/ml vincristine, 1 µg/ml
daunorubicin, 0.5 µg/ml doxorubicin, 7.5 µg/ml methotrexate and
1.25 µg/ml cytarabine, with or without 20 µM curcumin. Untreated
cells served as the control group.
Viability assay
Cell viability was determined by flow cytometry
using a 7-AAD dye exclusion test. After treatment with the various
drugs, cells were harvested, washed once in PBS and centrifuged at
100 × g at room temperature for 1 min, and incubated with 100 µl
PBS, 20 µl anti-CD45-FITC (dilution, 1:10; cat. no. IM0782U;
Beckman Coulter, Inc.) and 100 µl 7-AAD for 20 min at room
temperature in the dark. Following incubation, the cells were
suspended in PBS and 20,000 events were analyzed using a Gallios
Flow Cytometer (Beckman Coulter, Inc.). Gating was set to exclude
cell debris and autofluorescence.
DNA oxidation
To determine if curcumin prevents oxidative damage
to DNA caused by chemotherapy treatment, oxidative DNA adducts were
measured using the highly sensitive 8-OHdG. DNA was isolated from
the cells using QIAamp DNA Mini kit (Qiagen GmbH) and mixed with 50
µl nuclease-free water. Later, the cells were digested with Mung
Bean Nuclease (6 U; Promega, Madison, WI, USA) at 37°C for 45 min,
followed by treatment with alkaline phosphatase (2 U) for an
additional 45 min. DNA was precipitated with absolute ethanol and
centrifuged at 2,370 × g for 2 min, followed by hydration with 50
µl nuclease-free water. The digested DNA was added to the 8-OHdG
(Highly Sensitive 8-OHdG Check ELISA kit; cat. no. KOG-HS10E; Japan
Institute for the Control of Aging, Fukuroi, Japan.) well strip and
incubated with 50 µl primary monoclonal antibody (dilution, 1 µg/50
µl) specific for 8-OHdG at 4°C overnight. Following incubation, 3
washes were performed with 250 µl washing solution (Japanese
Institute for the Control of Aging) at room temperature, with
agitation of the plate from side to side for 20 seconds, disposing
washing solution each time. The samples were then incubated with 50
µl horseradish peroxidase-conjugated anti-mouse secondary antibody
(dilution, 1 µg/5 µl; cat. no. 405310) for 1 h at room temperature
in the dark. After 3 washes, 50 µl chromatic solution (Japanese
Institute for the Control of Aging) was added and incubated for 15
min at 4°C. The reaction was terminated following addition of 100
µl termination solution (Japanese Institute for the Control of
Aging), and the samples were analyzed at a wavelength of 450 nm in
a plate spectrophotometer.
NF-κB detection
To assess the involvement of curcumin in NF-κB
activation, flow cytometry was performed using
anti-human-phospho-NF-κB p65 antibody (Beckman Coulter, Inc.).
After the treatment the cells were fixed using PerFix Fixative
reagent (Beckman Coulter, Inc.) for 10 min at room temperature and
permeabilized using PerFix Permeabilizing reagent for 5 min at 37°C
in a water bath (PerFix EXPOSE Phospho-Epitopes Exposure kit;
Beckman Coulter, Inc.). A total of 50 µl staining reagent pre-mixed
with 2 µl conjugated anti-NF-κB-AlexaFluor 647 (dilution, 1:50;
cat. no. A88940; Beckman Coulter, Inc.) and 10 µl anti-CD45-FITC
antibodies were added to each tube immediately and incubated at
room temperature for 30 min in the dark. Cells were then washed
with 3 ml 1X wash reagent (PerFix EXPOSE; Beckman Coulter, Inc.) at
room temperature, centrifuged at 300 × g for 5 min. The washing
solution was removed and the cells were suspended in 500 µl final
1X reagent (PerFix EXPOSE; Beckman Coulter, Inc.), and 20,000
events were analyzed using Gallios software version 10 (Beckman
Coulter, Inc.). Gating was set to exclude cell debris and
autofluorescence.
Gene expression
To determine changes in the expression of various
genes that are downregulated by NF-κB, RNA extraction was performed
using QIAzol Lysis reagent. The cells were washed with PBS at room
temperature, centrifuged at 100 × g for 2 min at room temperature
and incubated with 1 ml QIAzol Lysis reagent for 5 min, followed by
the addition of 200 µl chloroform and centrifugation at 12,350 × g
at 4°C for 10 min. The aqueous phase was recovered in 500 µl
isopropanol and incubated at −20°C overnight. Next, samples were
centrifuged at 12,350 × g at 4°C for 10 min and washed twice with
70% ethanol, with centrifugation performed at 9,680 × g at 4°C for
5 min between each wash, and then air dried for 30 min. The samples
were reconstituted in 30 µl diethylpyrocarbonate-treated water
(Invitrogen; Thermo Fisher Scientific, Inc.). Reverse transcription
was performed using 1 µg total RNA and a High-Capacity cDNA Reverse
Transcription kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) in a GeneAmp PCR System 9700 thermal cycler (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The reaction
conditions were as follows: 25°C for 25 min, 37°C for 120 min, 85°C
for 5 min and infinite hold at 4°C. Quantitative polymerase chain
reaction (PCR) was performed using TaqMan Assays for the different
genes and the TaqMan Universal PCR Master Mix (Applied Biosystems;
Thermo Fisher Scientific, Inc.) in a 7900HT Fast Real-Time PCR
System using SDS 2.4 software (Applied Biosystems; Thermo Fisher
Scientific, Inc.). PCR was performed according to the
manufacturer's protocol, and the cycling conditions were as
follows: 50°C for 2 min, 95°C for 10 min, 95°C for 15 sec and 60°C
for 1 min (40 cycles). Data was quantified according to the
relative quantitation 2−∆∆Cq method (12), using the GUSB gene as an
endogenous control and the chemotherapy treatment groups without
curcumin as a calibrator (13).
Caspase-3 detection
To determine whether curcumin potentiates caspase-3
activity, flow cytometry was performed using polyclonal anti-human
cleaved caspase-3 (Asp-175). The cells (5×105) were
mixed with 20 µl anti-CD45-PC5 (dilution, 1:10; cat. no. IM2653U;
Beckman Coulter, Inc.) and incubated for 20 min at room temperature
in the dark. Cells were fixed using 100 µl IntraPrep Fixation
reagent (Beckman Coulter, Inc.) for 15 min at room temperature,
washed in 4 ml PBS and centrifuged at 300 × g for 5 min. Subsequent
to washing, the cells were permeabilized for 5 min at room
temperature using IntraPrep™ Permeabilization reagent (Beckman
Coulter, Inc.). Next, the cells were incubated with 2 µl polyclonal
anti-human cleaved caspase-3 (Asp-175) for 45 min at room
temperature in the dark. The cells were then washed and suspended
in 500 µl PBS and 20,000 events were analyzed using Gallios
software version 10 (Beckman Coulter, Inc.). Gating was set to
exclude cell debris and autofluorescence.
Statistical analysis
Differences in various parameters were compared in
the control and treatment groups using the PASW 18.0 Software
(SPSS, Inc., Chicago, IL, USA). The data were firstly analyzed
using the Kolmogorov-Smirnov test. When the data had a normal
distribution, the groups were compared using the Student's t-test.
When the distribution did not have a normal distribution, the data
were analyzed using Mann-Whitney U test. P<0.05 was considered
to indicate a statistically significant difference.
Results
The antitumor properties of curcumin have been
evaluated in a large number of solid tumors (14), however, less is known regarding
hematological neoplasias.
Curcumin decreases cell viability of
REH cells
To evaluate the effect of curcumin alone on cell
viability, flow cytometry was performed using 7-AAD. To determine
the effect of curcumin on the viability of REH cells, six different
concentrations were investigated (10, 20, 25, 30, 40 and 50 µM).
Curcumin decreased cell viability in a dose-dependent manner in REH
cells, and following treatment with 25, 30, 40 and 50 µM curcumin,
the cell viability was significantly decreased when compared with
the control (P<0.001). A dose of 20 µM curcumin was selected for
further experiments, as the next tested dose of 25 µM curcumin had
statistical differences in cell viability compared with the control
group from a pilot study.
Combined treatment with
chemotherapeutic agents and curcumin decreases cell viability in
REH cells
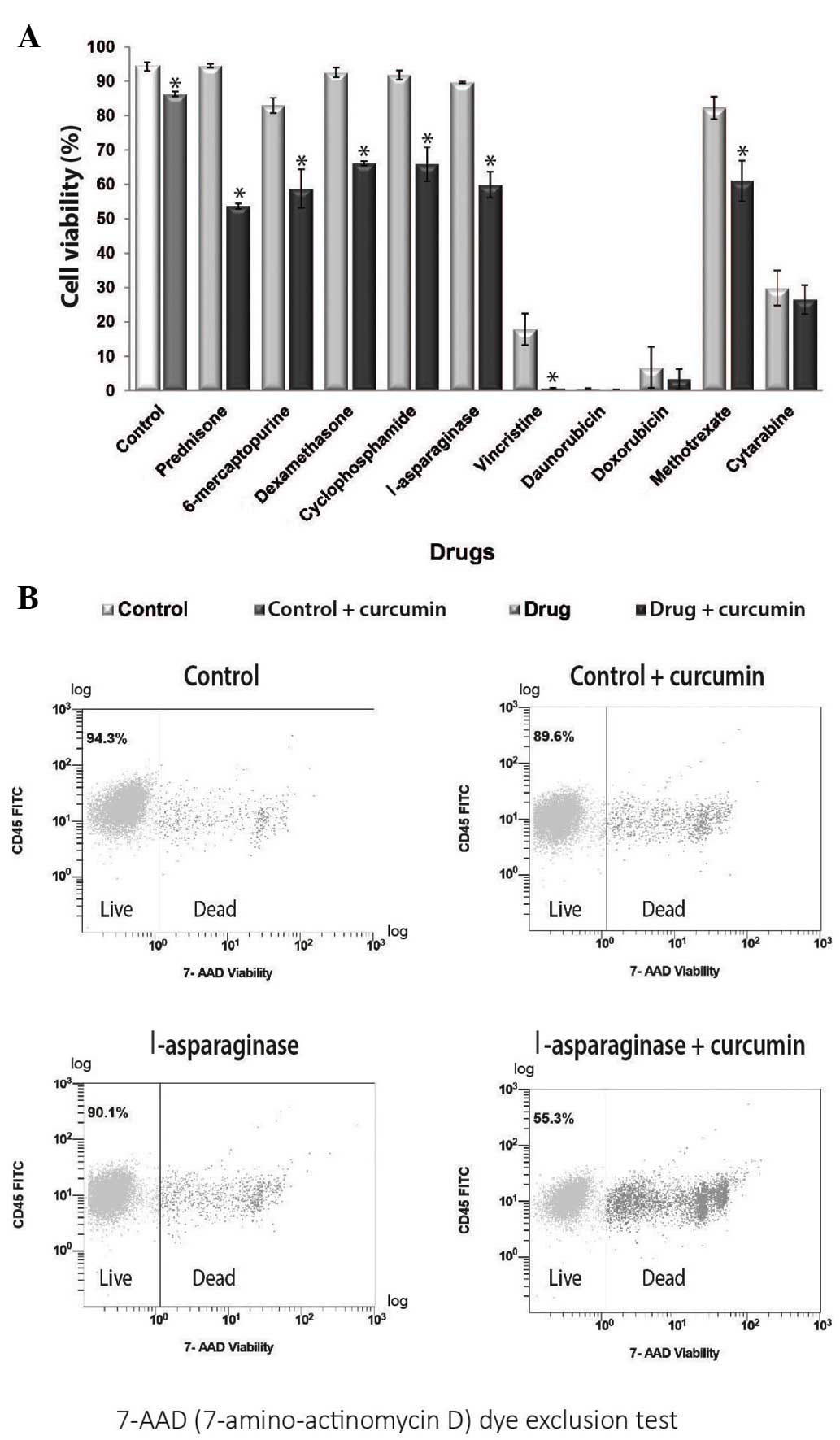
7-AAD flow cytometry was performed to investigate
whether curcumin potentiates the effect of chemotherapeutic agents
and decreases the cell viability of REH cells. The results revealed
that treatment with all chemotherapeutic agents reduced cell
viability when compared with the control. Furthermore, combined
treatment with curcumin resulted in a further decrease in cell
viability for all chemotherapeutic drugs (Fig. 2A). The group treated with curcumin
alone exhibited a cell viability of 86.3%, whereas the group
treated with l-asparaginase alone exhibited a cell viability of
89.7%. Notably, combined treatment with l-asparaginase and curcumin
decreased cell viability to 59.9% (Fig.
2B). The same potentiating effect of curcumin was observed with
prednisone, cyclophosphamide, 6-mercaptopurine, dexamethasone,
vincristine and methotrexate (P<0.05).
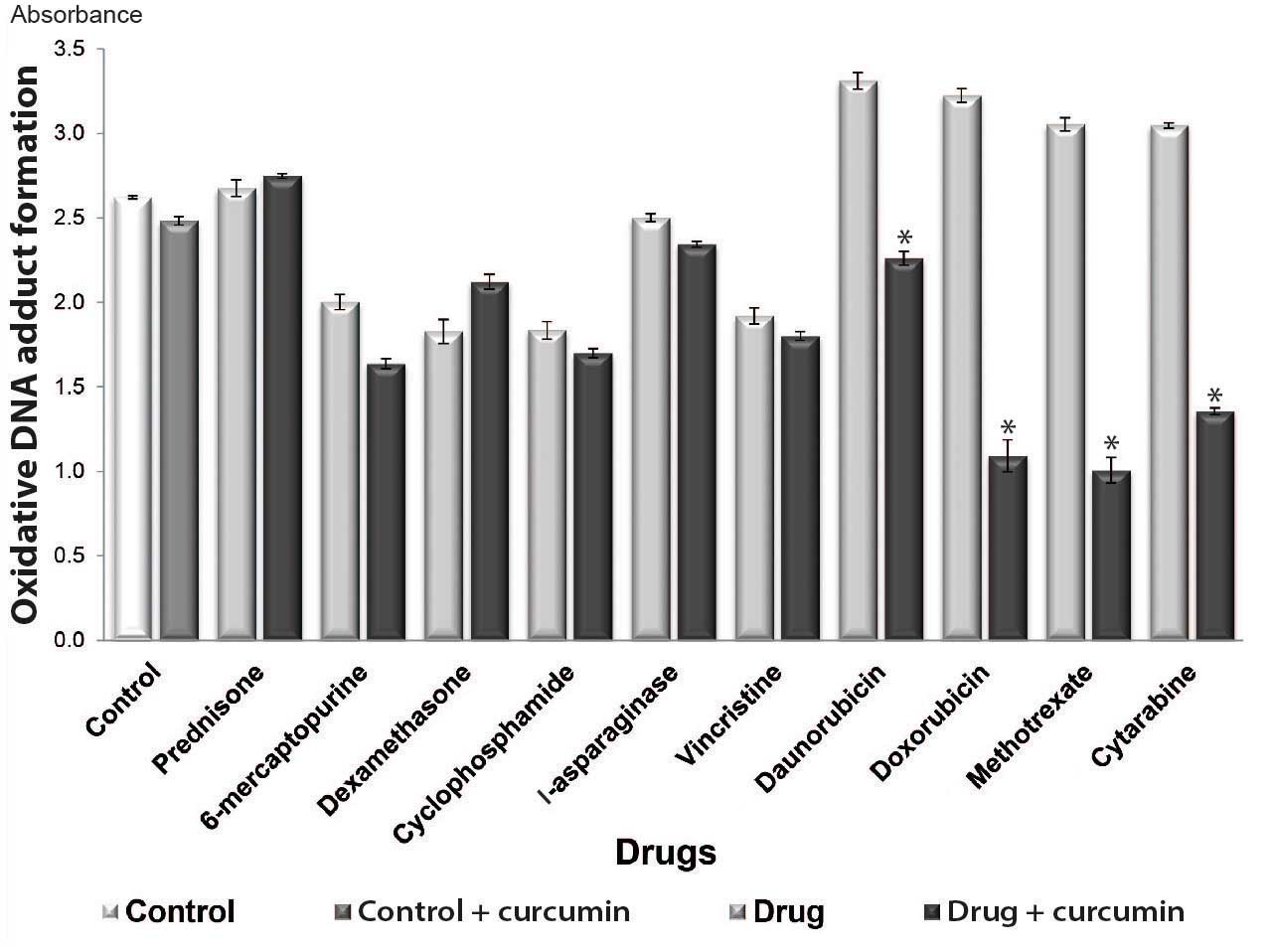
Curcumin prevents DNA oxidation
It has been demonstrated that chemotherapeutic
agents induce DNA damage in normal and abnormal cells (15). To determine whether curcumin acts as
an antioxidant when combined with chemotherapeutic drugs (16), in the present study, oxidative DNA
adduct formation was analyzed in cultures treated with or without
curcumin. The results revealed that oxidative DNA adduct formation
was decreased in all combined treatment groups, with the exception
of the prednisone + curcumin and dexamethasone + curcumin treatment
groups. Significant decreases in DNA adduct formation were observed
in the groups treated with curcumin and daunorubicin, doxorubicin,
methotrexate and cytarabine (P<0.05) (Fig. 3).
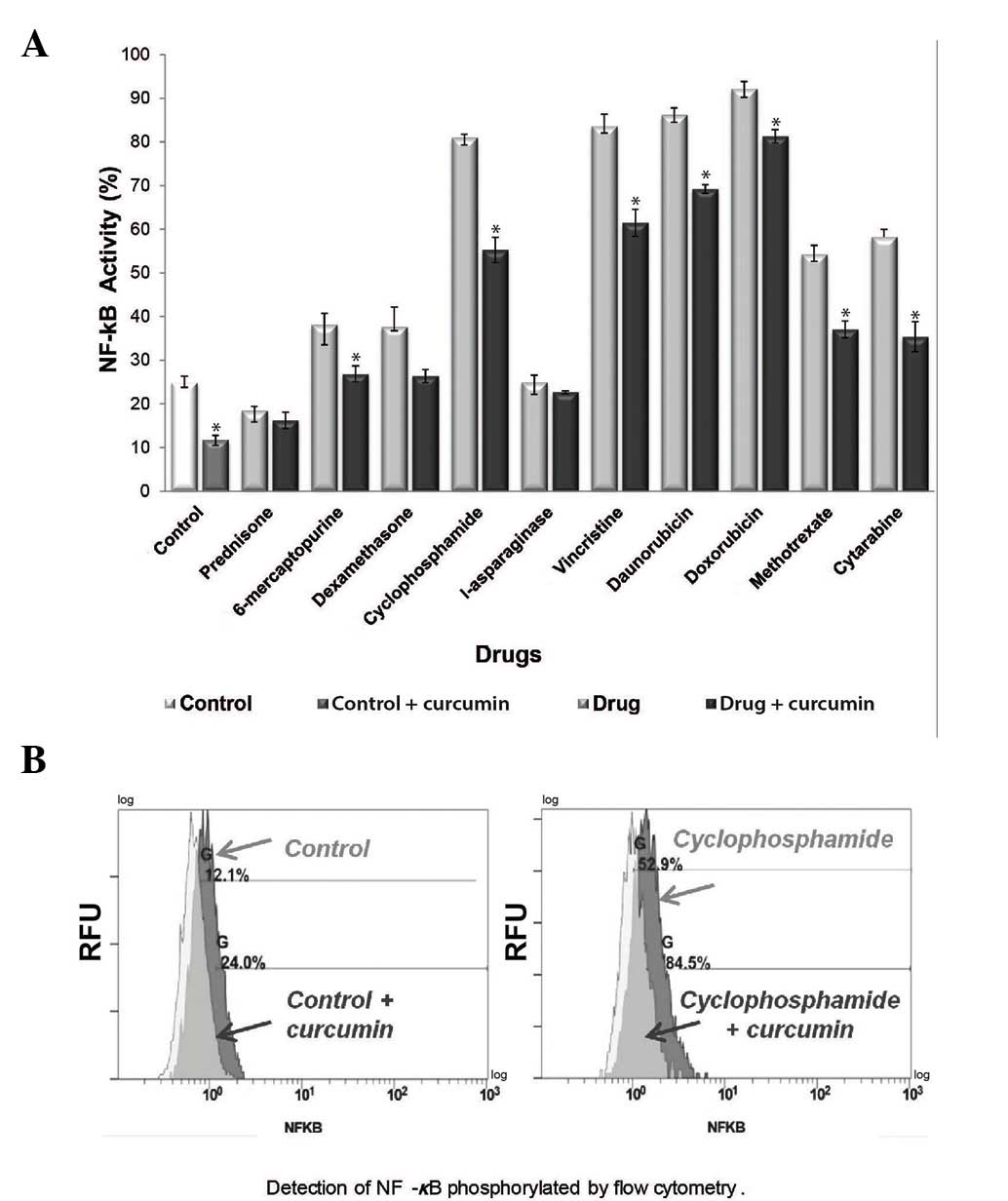
Curcumin decreases NF-κB activation in
cells treated with chemotherapeutic agents
It has been reported that constitutive NF-κB
activation occurs in ALL (7,17), and a previous study has suggested that
chemotherapy alone may increase this activation (18). To determine whether curcumin decreases
the levels of active NF-κB in the REH cell line, the levels of
NF-κB phosphorylated at Ser536 were evaluated by flow cytometry.
The results demonstrated that treatment with 8/10 of the
therapeutic agents led to increased NF-κB activity when compared
with the control group (P<0.05), whereas all of the combined
treatment groups (chemotherapeutic agent + curcumin) exhibited
decreased NF-κB activation (Fig. 4A).
The control and curcumin alone treatment groups exhibited an NF-κB
activation rate of 24.8 and 11.7%, respectively. The
cyclophosphamide treatment group exhibited an NF-κB activation rate
of 81%, while combined treatment with cyclophosphamide and curcumin
decreased the NF-κB activation rate to 55.3% (P<0.05) (Fig. 4B). The most significant NF-κB
inactivation was observed following combined treatment with
curcumin and 6-mercaptopurine, cyclophosphamide, vincristine,
daunorubicin, doxorubicin, methotrexate and cytarabine
(P<0.05).
Curcumin affects the expression of
NF-κB target genes
Since apoptosis and proliferation-related genes
Bcl-2, Bcl-xl, survivin, cyclin D1 and c-Myc, have all been
demonstrated to be regulated by NF-κB (19), in the present study, the expression of
these genes was analyzed. Gene expression was analyzed in the
chemotherapy-treated groups with and without curcumin. The gene
expression fold change was calculated using the group with
chemotherapy without curcumin as calibrator and the chemotherapy
with curcumin group as a target group. The gene expression of the
anti-apoptosis gene Bcl-xL was decreased by 2.5 fold in the
vincristine group; however, this difference was not significant
(P=0.513) (Fig. 5A). Furthermore,
Bcl-2 gene expression was decreased by 6.5 fold in the vincristine
+ curcumin group (P<0.05) (Fig.
5B). The expression of the survivin gene was decreased by 9.7
fold in the vincristine + curcumin group (P<0.05), when compared
with the vincristine group (Fig. 5C).
The expression of the proliferative gene c-Myc increased in the
methotrexate + curcumin group and decreased by 3.8 fold in the
doxorubicin + curcumin group when compared with their calibrator
(same chemotherapy without curcumin); however, these changes were
not significant (P=0.827 and P=0.275, respectively) (Fig. 5D). Notably, the expression of the
cyclin D1 gene was increased in 9/10 of the co-treated groups (with
the exception of the doxorubicin group) compared with their
calibrator; however, no significant differences were identified
(Fig. 5E). These findings indicate
that curcumin did not result in a downregulation pattern in the
c-Myc and cyclin D1 groups.
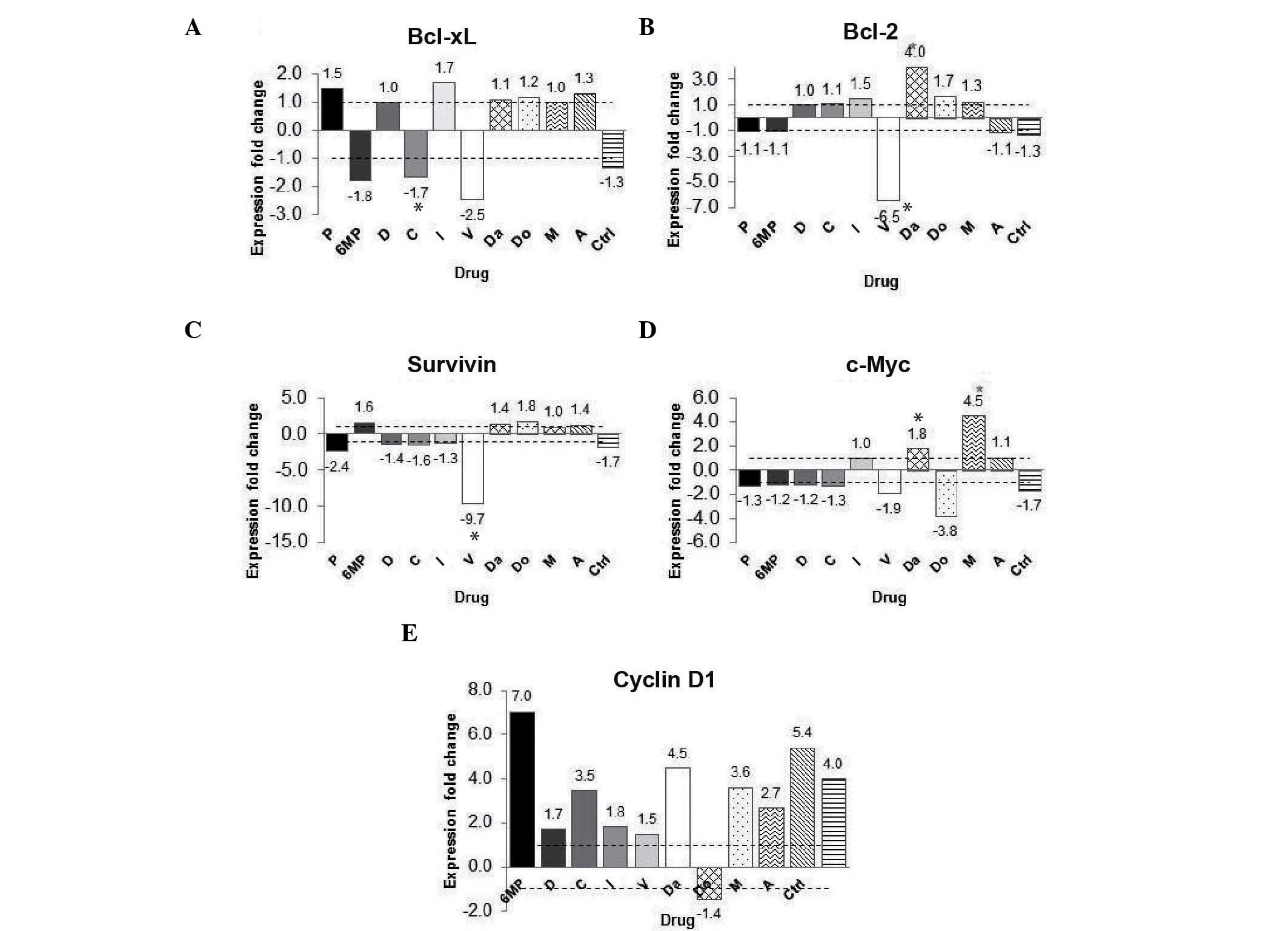 | Figure 5.Effect of curcumin on NF-κB-regulated
gene expression. (A) Changes in fold expression of the (A)
Bcl-extra large, (B) Bcl-2, (C) survivin, (D)
c-Myc and (E) cyclin D1 genes. The data are presented
as the fold-change in gene expression between the groups treated
with chemotherapy alone and the groups treated with chemotherapy
and curcumin.*P<0.05 vs. control. P, prednisone; 6MP,
6-mercaptopurine; D, dexamethasone; C, cyclophosphamide; L,
l-asparaginase; V, vincristine; Da, daunorubicin; Do, doxorubicin;
M, methotrexate; A, cytarabine; Ctrl, control; Bcl, B-cell
lymphoma; xL, extra large; NF-κB, nuclear factor-kappa B. |
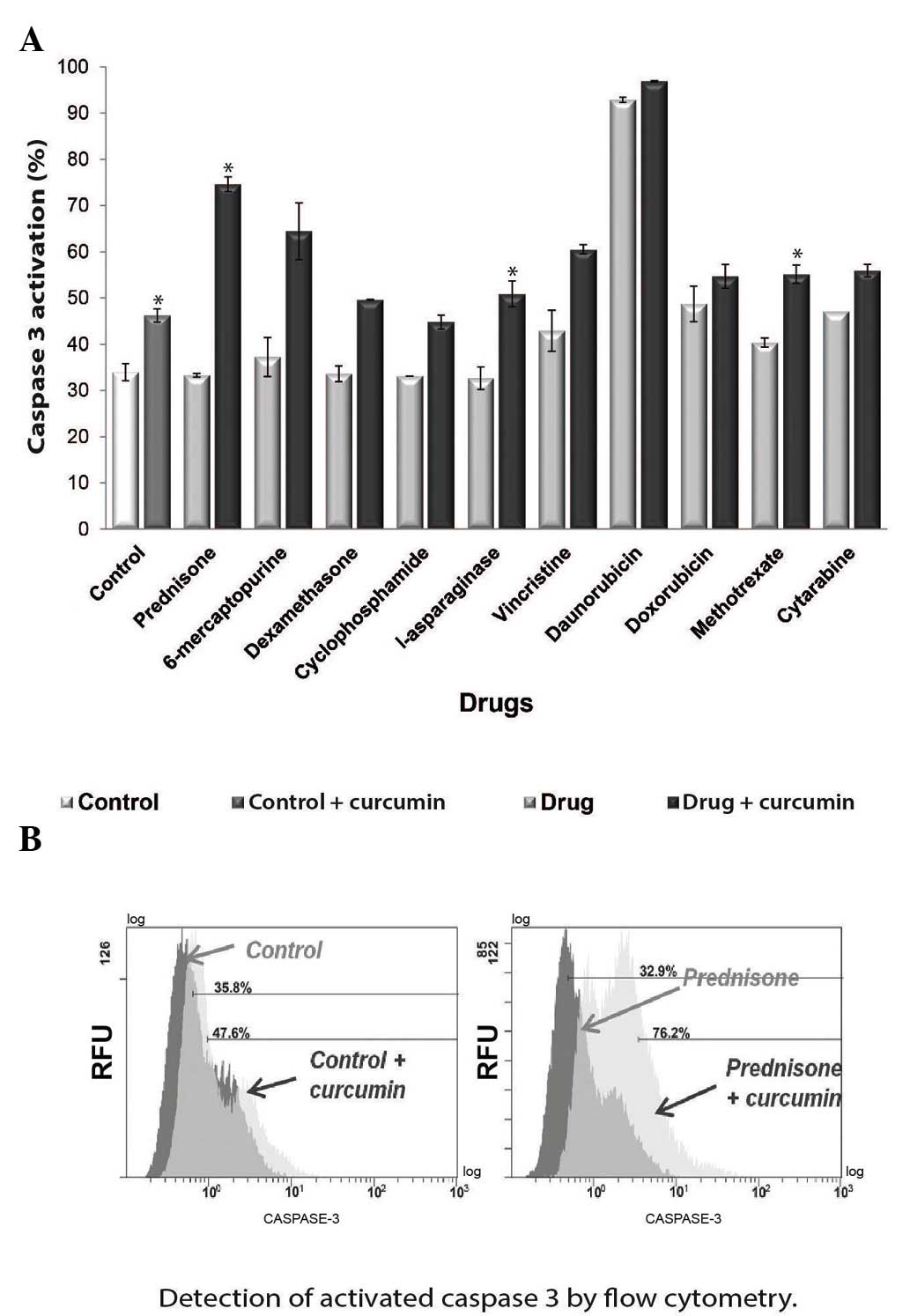
Curcumin activates caspase-3
As NF-κB has been demonstrated to exhibit an
anti-apoptosis effect (20), in the
present study, the effect of curcumin on apoptosis was
investigated. The activity of cleaved caspase-3, the effector
protein of the receptor-mediated and chemical-induced apoptosis
pathways (21), was evaluated. To
determine if curcumin induces caspase-3 activation, flow cytometry
was performed to analyze the percentage of active caspase-3 in the
chemotherapy-treated cultures with or without curcumin. The results
demonstrated that curcumin increased caspase-3 activity following
treatment with all the chemotherapeutic agents tested (Fig. 6A). The most significant increases in
caspase-3 activity were identified in the prednisone + curcumin
(Fig. 6B), l-asparaginase + curcumin
and methotrexate + curcumin groups. However, treatment with
curcumin alone also significantly increased caspase-3 activity
(P<0.05).
Discussion
In the present study, the effect of curcumin as a
phytochemical with chemopreventive and antitumor properties was
investigated, as curcumin has been previously used in herbal
medicine and as a dietary compound with non-toxic effects (22). The aim of the present study was to
evaluate the effect of curcumin in the human ALL REH cell line in
combination with a variety of therapeutic agents used to treat ALL.
The results revealed that NF-κB activation was decreased in all
chemotherapeutic agent + curcumin groups, and a subsequent increase
in apoptosis was also observed.
Curcumin was demonstrated to decrease the
chemotherapeutic activation of NF-κB. Previous studies have
revealed that chemotherapeutic agents increase NF-κB activation
(18), while curcumin is able to
reduce it (23). In the present
study, with the exception of prednisone, treatment with all the
chemotherapeutic agents tested resulted in increased NF-κB
activation when compared with the control groups. However,
treatment with 20 µM curcumin downregulated the constitutively
active NF-κB in the REH cell line, both alone and in combination
with all the therapeutic agents tested. It is postulated that this
downregulation occurs via activation of the inhibitor of kappa B α
(24).
The results of the present study indicate that
curcumin functions as a sensitizer in tumor cells and potentiates
the antitumor effect of the tested chemotherapeutic agents, as
shown by the decreased cell viability observed. Previous evidence
that curcumin may potentiate the antitumor effect of
chemotherapeutic agents in ALL, was initially observed in
ALL-derived Jurkat, REH and RS4;11 cell lines exposed to
l-asparaginase and curcumin via inhibition of protein kinase B
(AKT) and AKT-regulated gene products (25). In the present study, a decrease in
cell viability and an increase in apoptosis was observed following
treatment with cyclophosphamide, which was potentiated by curcumin.
By contrast, curcumin has been demonstrated to inhibit
cyclophosphamide-induced tumor regression in a breast cancer murine
model (26). The results of the
present study are in agreement with previous studies that have
demonstrated a decrease in cell viability following curcumin
treatment and the synergistic effect of curcumin following
co-treatment with l-asparaginase in leukemia Jurkat, REH and RS4;11
cell lines (25), and vincristine in
multiple myeloma cells (27). In the
present study, the observed decrease in cell viability following
co-treatment with curcumin and vincristine, daunorubicin or
doxorubicin was not statistically significant, which may be due to
cytotoxicity. Notably, the chemotherapeutic agents in which the
decrease in cell viability was most evident in the presence of
curcumin exhibited the highest levels of cell viability following
treatment with the chemotherapeutic agents alone. Thus, the
decrease in cell survival may be attributed to curcumin.
It was also reported that the downregulation of
NF-κB led to apoptosis of ALL cells, as indicated by the increased
expression of the apoptosis effector protein caspase-3 (27). Previously, the antitumoral effect of
curcumin was associated with caspase-3 activation (28), and in the present study, all
co-treatment groups exhibited an increase in caspase-3 activity. In
contrast to a previous study (29),
in the present study, treatment with vincristine alone activated
caspase-3, and the reported increase in caspase-3 activity
following combined treatment with l-asparaginase and curcumin was
confirmed (30). Notably, in the
present study, in the 6-mercaptopurine + curcumin group, the marked
increase in caspase-3 activity was not significant, despite the
significant decrease in cell viability observed. Therefore, it can
be hypothesized that cell death may be activated via an alternative
pathway. In the present study, curcumin treatment did not lead to
the downregulation of anti-apoptosis and proliferative genes, as
previously described (31),
indicating that an alternative pathway may be activated, instead of
that involving NF-κB. A previous study revealed that, in AML
daunorubicin-resistant cell lines, apoptosis increased following
combined treatment with daunorubicin and curcumin (32), and in the present study, an increase
in caspase-3 activity was observed in the daunorubicin and curcumin
co-treated cell cultures, although this increase was not
statistically significant.
Oxidative stress caused by therapeutic agents used
for the treatment of ALL cause damage in non-cancerous tissues
(33) leading to the formation of
oxidative DNA adducts (34). Curcumin
may act as a scavenger of the free radicals caused by the therapy,
subsequently reducing these molecules (35). Notably, a previous study revealed that
in the NG108-15 (glioblastoma/neuroblastoma hybrid) cell line,
curcumin protected the cells from oxidative damage when
administered in combination with hydrogen peroxide, but not
following pre-treatment (36). In the
present study, cell cultures were treated simultaneously with the
chemotherapeutic agents and curcumin, which may explain the free
radical scavenging properties of curcumin. Notably, not all of the
chemotherapeutic agents tested in the present study exhibited
increased levels of 8-OHdG when compared with the control group.
However, all co-treated groups exhibited a reduction in 8-OHdG
molecules compared with the control group, indicating that curcumin
controlled the free radicals produced by the therapeutic agents,
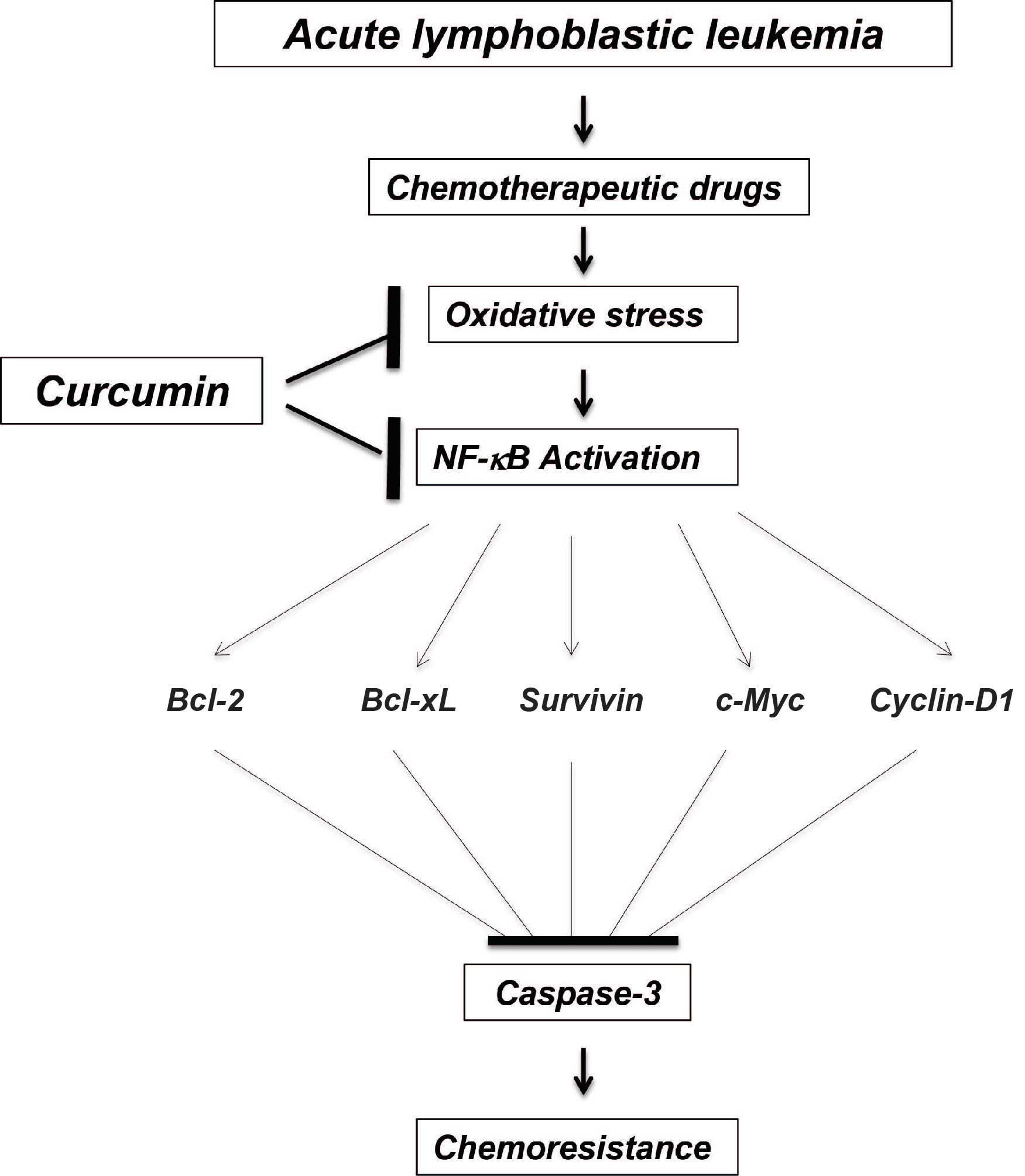
resulting in less DNA damage. A proposed model that demonstrates
the possible mechanism of action of combined treatment with
curcumin and chemotherapeutic agents for ALL is shown in Fig. 7.
Clinical studies investigating the efficacy of
curcumin for the treatment of pancreatic (37) and colorectal cancer (38) have yielded positive results. However,
to the best of our knowledge, no studies have investigated
pediatric hematological neoplasias to date. These promising
clinical trials in solid tumors, considered together with the
decrease in multi-drug resistance gene expression over curcumin in
primary ALL cell cultures (39) and
potentiation through curcumin of l-asparaginase (25) correspond to the only studies in
vitro, highlighting the importance of the present study.
Additional in vitro and mice models to assess the effect of
combined treatment with chemotherapeutic agents and curcumin are
required. Possible interactions between curcumin at various
concentrations and chemotherapeutic agents cannot be excluded and
thus, more studies that investigate the possible interactions
between curcumin and chemotherapeutic agents are also required.
In conclusion, the present study revealed that
curcumin inhibits survival, increases apoptosis and decreases DNA
oxidation of REH leukemia cells in an NF-κB-dependent manner, both
alone and in combination with all the therapeutic agents tested
(prednisone, 6-mercaptopurine, dexamethasone, cyclophosphamide,
l-asparaginase, vincristine, daunorubicin, doxorubicin,
methotrexate and cytarabine). The application of this compound in
the treatment of pediatric lymphoblastic leukemia may improve the
outcome of patients. Overall, the present results indicate that
curcumin may improve the efficacy of chemotherapeutic agents
against ALL.
Acknowledgements
The authors would like to thank Dr Bharat B.
Aggarwal (Cytokine Research Section Department of Molecular
Oncology, The University of Texas, M. D. Anderson Cancer Center,
Houston, TX, USA) for assistance with the present study, the St.
Jude Children's Research Hospital (Memphis, TN, USA) for providing
the REH cell line and all the staff members of the Cytogenetics
Unit (Pediatric Hematology and Oncology Service, Pediatric
Division, Civil Hospital from Guadalajara, Guadalajara, Jalisco,
México). The present study was partially supported by the State
Council of Science and Technology of Jalisco (Consejo Estatal de
Ciencia y Tecnología de Jalisco; Guadalajara, México; grant no.
5-2010-1-1083).
References
|
1
|
Pui CH: Childhood Leukemias. 3rd.
Cambridge University Press; Cambridge: 2012, View Article : Google Scholar
|
|
2
|
Pui CH and Jeha S: New therapeutic
strategies for the treatment of acute lymphoblastic leukaemia. Nat
Rev Drug Discov. 6:149–165. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Libermann TA and Zerbini LF: Targeting
transcription factors for cancer gene therapy. Curr Gene Ther.
6:17–33. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brown M, Cohen J, Arun P, Chen Z and Van
Waes C: NF-kappaB in carcinoma therapy and prevention. Expert Opin
Ther Targets. 12:1109–1122. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Montagut C, Tusquets I, Ferrer B,
Corominas JM, Bellosillo B, Campas C, Suarez M, Fabregat X, Campo
E, Gascon P, et al: Activation of nuclear factor-kappaB is linked
to resistance to neoadjuvant chemotherapy in breast cancer
patients. Endocr Relat Cancer. 13:607–616. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li F and Sethi G: Targeting transcription
factor NF-kappaB to overcome chemoresistance and radioresistance in
cancer therapy. Biochim Biophys Acta. 1805:167–180. 2010.PubMed/NCBI
|
|
7
|
Kordes U, Krappmann D, Heissmeyer V,
Ludwig WD and Scheidereit C: Transcription factor NF-kappaB is
constitutively activated in acute lymphoblastic leukemia cells.
Leukemia. 14:399–402. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sethi G, Sung B and Aggarwal BB: Nuclear
factor-kappaB activation: From bench to bedside. Exp Biol Med
(Maywood). 233:21–31. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ravindran J, Prasad S and Aggarwal BB:
Curcumin and cancer cells: How many ways can curry kill tumor cells
selectively? AAPS J. 11:495–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lao CD, Ruffin MT IV, Normolle D, Heath
DD, Murray SI, Bailey JM, Boggs ME, Crowell J, Rock CL and Brenner
DE: Dose escalation of a curcuminoid formulation. BMC Complement
Altern Med. 6:102006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gupta SC, Kismali G and Aggarwal BB:
Curcumin, a component of turmeric: From farm to pharmacy.
Biofactors. 39:2–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Beillard E, Pallisgaard N, van der Velden
VH, Bi W, Dee R, van der Schoot E, Delabesse E, Macintyre E,
Gottardi E, Saglio G, et al: Evaluation of candidate control genes
for diagnosis and residual disease detection in leukemic patients
using 'real-time' quantitative reverse-transcriptase polymerase
chain reaction (RQ-PCR) - a Europe against cancer program.
Leukemia. 17:2474–2486. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Anand P, Sundaram C, Jhurani S,
Kunnumakkara AB and Aggarwal BB: Curcumin and cancer: An ‘old-age’
disease with an ‘age-old’ solution. Cancer Lett. 267:133–164. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen Y, Jungsuwadee P, Vore M, Butterfield
DA and St Clair DK: Collateral damage in cancer chemotherapy:
Oxidative stress in nontargeted tissues. Mol Interv. 7:147–156.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Blakemore LM, Boes C, Cordell R and Manson
MM: Curcumin-induced mitotic arrest is characterized by spindle
abnormalities, defects in chromosomal congression and DNA damage.
Carcinogenesis. 34:351–360. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xue TY, Xu W, An Q, Wu Y, Xu CP and Zhang
XY: Expression of nuclear transcription factor kappaB in childhood
acute lymphoblastic leukemia and its significance. Zhongguo Shi Yan
Xue Ye Xue Za Zhi. 15:767–771. 2007.(In Chinese). PubMed/NCBI
|
|
18
|
Bottero V, Busuttil V, Loubat A, Magné N,
Fischel JL, Milano G and Peyron JF: Activation of nuclear factor
kappaB through the IKK complex by the topoisomerase poisons SN38
and doxorubicin: A brake to apoptosis in HeLa human carcinoma
cells. Cancer Res. 61:7785–7791. 2001.PubMed/NCBI
|
|
19
|
Aggarwal BB and Gehlot P: Inflammation and
cancer: How friendly is the relationship for cancer patients? Curr
Opin Pharmacol. 9:351–369. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karin M: NF-kappaB as a critical link
between inflammation and cancer. Cold Spring Harb Perspect Biol.
1:a0001412009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Anto RJ, Mukhopadhyay A, Denning K and
Aggarwal BB: Curcumin (diferuloylmethane) induces apoptosis through
activation of caspase-8, BID cleavage and cytochrome c release: Its
suppression by ectopic expression of Bcl-2 and Bcl-xl.
Carcinogenesis. 23:143–150. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shishodia S, Amin HM, Lai R and Aggarwal
BB: Curcumin (diferuloylmethane) inhibits constitutive NF-kappaB
activation, induces G1/S arrest, suppresses proliferation, and
induces apoptosis in mantle cell lymphoma. Biochem Pharmacol.
70:700–713. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kasinski AL, Du Y, Thomas SL, Zhao J, Sun
SY, Khuri FR, Wang CY, Shoji M, Sun A, Snyder JP, et al: Inhibition
of IkappaB kinase-nuclear factor-kappaB signaling pathway by
3,5-bis(2-flurobenzylidene)piperidin-4-one (EF24), a novel
monoketone analog of curcumin. Mol Pharmacol. 74:654–661. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang H, Geng QR, Wang L and Lu Y: Curcumin
potentiates antitumor activity of L-asparaginase via inhibition of
the AKT signaling pathway in acute lymphoblastic leukemia. Leuk
Lymphoma. 57:1376–1382. 2012. View Article : Google Scholar
|
|
26
|
Somasundaram S, Edmund NA, Moore DT, Small
GW, Shi YY and Orlowski RZ: Dietary curcumin inhibits
chemotherapy-induced apoptosis in models of human breast cancer.
Cancer Res. 62:3868–3875. 2002.PubMed/NCBI
|
|
27
|
Bharti AC, Donato N, Singh S and Aggarwal
BB: Curcumin (diferuloylmethane) down-regulates the constitutive
activation of nuclear factor-kappa B and IkappaBalpha kinase in
human multiple myeloma cells, leading to suppression of
proliferation and induction of apoptosis. Blood. 101:1053–1062.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chan WH, Wu HY and Chang WH: Dosage
effects of curcumin on cell death types in a human osteoblast cell
line. Food Chem Toxicol. 44:1362–1371. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Al-Katib A, Arnold AA, Aboukameel A, Sosin
A, Smith P, Mohamed AN, Beck FW and Mohammad RM: I-kappa-kinase-2
(IKK-2) inhibition potentiates vincristine cytotoxicity in
non-Hodgkin's lymphoma. Mol Cancer. 9:2282010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang H, Geng QR, Wang L and Lu Y: Curcumin
potentiates antitumor activity of L-asparaginase via inhibition of
the AKT signaling pathway in acute lymphoblastic leukemia. Leuk
Lymphoma. 53:1376–1382. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kunnumakkara AB, Anand P and Aggarwal BB:
Curcumin inhibits proliferation, invasion, angiogenesis and
metastasis of different cancers through interaction with multiple
cell signaling proteins. Cancer Lett. 269:199–225. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rao J, Xu DR, Zheng FM, Long ZJ, Huang SS,
Wu X, Zhou WH, Huang RW and Liu Q: Curcumin reduces expression of
Bcl-2, leading to apoptosis in daunorubicin-insensitive CD34+ acute
myeloid leukemia cell lines and primary sorted CD34+ acute myeloid
leukemia cells. J Transl Med. 9:712011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen Y, Jungsuwadee P, Vore M, Butterfield
DA and St Clair DK: Collateral damage in cancer chemotherapy:
Oxidative stress in nontargeted tissues. Mol Interv. 7:147–156.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Valavanidis A, Vlachogianni T and Fiotakis
C: 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of
oxidative stress and carcinogenesis. J Environ Sci Health C Environ
Carcinog Ecotoxicol Rev. 27:120–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thangapazham RL, Sharma A and Maheshwari
RK: Multiple molecular targets in cancer chemoprevention by
curcumin. AAPS J. 8:E443–E449. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mahakunakorn P, Tohda M, Murakami Y,
Matsumoto K, Watanabe H and Vajaragupta O: Cytoprotective and
cytotoxic effects of curcumin: Dual action on H2O2-induced
oxidative cell damage in NG108-15 cells. Biol Pharm Bull.
26:725–728. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dhillion N, Aggarwal BB, Newman RA, Wolff
RA, Kunnumakkara AB, Abbruzzese JL, Ng CS, Badmaev V and Kurzrock
R: Phase II trial of curcumin in patients with advanced pancreatic
cancer. Clin Cancer Res. 14:4491–4499. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
He ZY, Shi CH, Wen H, Li FL, Wang BL and
Wang J: Upregulation of p53 expression in patients with colorectal
cancer by administration of curcumin. Cancer Invest. 29:208–213.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Anuchapreeda S, Thanarattanakorn P,
Sittipreechacharn S, Tima S, Chanarat P and Limtrakul P: Inhibitory
effect of curcumin on MDR1 gene expression in patient leukemic
cells. Arch Pharm Res. 29:866–873. 2006. View Article : Google Scholar : PubMed/NCBI
|