Introduction
Metastasis is the process by which tumor cells
migrate from their original site to another host organ by
infiltrating the blood stream (1).
The first step in metastasis is a change in the cells from an
epithelial to a mesenchymal phenotype, termed the
epithelial-mesenchymal transition (EMT) (2,3). During
EMT, cells lose their basal polarity and their contact with one
another, allowing for penetration of the basement membrane
(4), via modulation of protein kinase
B (AKT) activation (5). The
phenotypic transformation includes a transcriptional change in
associated genes, including vimentin, N-cadherin, fibronectin,
Snail, zinc finger E-box-binding homeobox (Zeb), and Twist
(6,7).
These genes are involved in the morphological changes and survival
of transited tumor cells (8).
Transforming growth factor β (TGF-β) is a
multifunctional cytokine that acts as an inhibitor of early-stage
cancer, but promotes EMT in late-stage cancer, as it plays a key
part in the wound healing process, which requires cell migration
(8–11). Upon TGF-β binding to the TGF-β type 2
receptor activin receptor-like kinase 1 (ALK1), the TGF-β type 1
receptor (ALK5) is phosphorylated (10,12), which
then results in the phosphorylation of receptor-regulated mothers
against decapentaplegic homologs (R-SMADs). R-SMADs include SMAD2
and SMAD3, which transduce extracellular ligand signaling from
TGF-β (10,12). The activated R-SMAD combines with the
common-mediator SMAD4, which translocates to the nucleus (13), where regulation of the transcriptional
target genes involved in EMT occurs (14). There is also a TGF-β-associated
non-SMAD signaling pathway that interacts with EMT (15). The pathway is initiated by activation
of phosphoinositide-3 kinase (PI3K)-AKT-mechanistic target of
rapamycin complex 1 (mTORC1) or extracellular signal-regulated
kinase (ERK) signaling, and also leads to cellular changes
(14,16).
In Eastern Asia, the Inula britannica plant
is used to treat bronchitis, disorders of the digestive system, and
inflammation (17,18). A number of biologically active
sesquiterpene lactones have been isolated from this plant, of which
eupatolide is one (19). Several
studies have revealed the biological functions of eupatolide:
Eupatolide exerts anti-inflammatory effects by inhibiting nuclear
factor κ-light-chain-enhancer of activated B cells activation and
nitric oxide and tumor necrosis factor (TNF) production (13,15). In
addition, we hypothesized that eupatolide may exert antitumor
activity, as eupatolide inhibits platelet-derived growth factor
(PDGF)-induced proliferation and migration of vascular smooth
muscle cells (VSMCs) through heme oxygenase 1 (HO-1) induction via
the reactive oxygen species (ROS)/nuclear factor (erythroid-derived
2)-like 2 (NRF2) pathway (20), and
stimulates TNF-related apoptosis-inducing ligand (TRAIL)-induced
apoptosis by downregulation of cellular FLICE-like inhibitory
protein, an inhibitor of the TRAIL signaling pathway (21).
The aim of the present study was to investigate the
effect of eupatolide on proliferation, migration and invasion in
breast cancer cells. Breast cancer cells were treated with
eupatolide to investigate whether eupatolide inhibited migration
and invasion in breast cancer cells. To identify a signaling
pathway by which eupatolide affects migration and invasion, western
blotting and Matrigel assay were used.
Materials and methods
Cell culture, chemicals and
antibodies
Human breast cancer MDA-MB-231 and MCF-7 cells were
obtained from American Type Culture Collection (Manassas, VA, USA)
and maintained in Dulbecco's modified Eagle's medium (HyClone; GE
Healthcare, Chicago, IL, USA) with 10% fetal bovine serum (HyClone;
GE Healthcare). Cells were incubated in a humidified atmosphere at
37°C with 5% CO2 (22).
Human epidermal growth factor (EGF) and human transforming growth
factor-β1 (TGF-β1) were purchased from Sigma-Aldrich (Merck KGaA,
Darmstadt, Germany). The isolation of eupatolide from I.
britannica and its chemical structure have been reported
previously (23). Antibodies against
Snail (cat. no. ab53519), Slug (cat. no. ab27568), SMAD4 (cat. no.
ab40759) and phospho-EGFR (cat. no. ab76195) were purchased from
Abcam (Cambridge, UK). Antibodies against vimentin (cat. no. 3932),
phospho-AKT (Ser473; cat. no. 9271), ERK1/2 (cat. no. 9102),
phospho-ERK1/2 (cat. no. 9106), EGF receptor (EGFR; cat. no. 4267),
phospho-SMAD3 (Ser423/425; cat no. 9520), SMAD3 (cat no. 9523),
phospho-SMAD1/5 (cat. no. 9516), SMAD1 (cat. no. 6944) and SMAD5
(cat. no. 12534) were obtained from Cell Signaling Technology, Inc.
(Danvers, MA, USA). The antibody specific for E-cadherin was
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA).
Proliferation assay
The proliferation assay was performed by direct
counting. MCF-7 and MDA-MB-231 cells were seeded in 12-well plates
at a density of 1×104 cells/well. Once cells were fully
attached to the surface of the plate, they were treated with either
PBS or 10 µM eupatolide for various durations. After 24, 48 or 72 h
of treatment, the proliferating cells were detached using a
trypsin-EDTA solution and counted using a Neubauer haemocytometer
(Miltrnyi Biotech GmbH, Bergisch Gladbach, Germany). The experiment
was performed in triplicate as described previously (24).
Wound healing assay
To assess the effect of eupatolide on the migration
of breast cancer cells, MDA-MB-231 and MCF7 cells were seeded in
96-well plates at a density of 3×104 cells/well. The
confluent cell monolayer was wounded by scraping with a Wound-Maker
provided with the IncuCyte system (Essen Bioscience, Ann Arbor, MI,
USA). Cells were then treated with EGF (100 ng/ml) or TGF-β1 (50
ng/ml) in the presence or absence of 10 µM eupatolide for 24 h and
control cells were treated with PBS. The kinetics of cell migration
were monitored using the IncuCyte system, as described previously
(25).
Invasion assay
MCF-7 and MDA-MB-231 cells (2×104 per
well) suspended in 100 µl serum-free DMEM were seeded into
Transwell inserts (pore size, 8 µm; Costar; Corning Incorporated,
NY, USA) coated with Matrigel. The lower chamber was filled with
500 µl DMEM containing 10% FBS. The cells were then treated with
EGF (100 ng/ml) or TGF-β1 (50 ng/ml) in the presence or absence of
10 µM eupatolide for 24 h and control cells were treated with PBS.
After 24 h, the cells invading the lower chamber were stained with
crystal violet and images of the invaded cells in the bottom
chamber were imaged. To quantify the results, crystal violet stains
were dissolved in 33% acetic acid and then the absorbance at a
wavelength of 570 nm was evaluated using the Epoch microplate
reader (BioTek Instruments Inc., Winooski, VT, USA). Each
experiment was performed three times.
RNA isolation and reverse
transcription-polymerase chain reaction (RT-PCR)
MDA-MB-231 cells were seeded and pre-treated with
either PBS or 10 µM eupatolide for 30 min, followed by treatment
with 50 ng/ml TGF-β1 for 1, 2 or 3 h. Total RNA was isolated using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), and 4 µg RNA was reverse transcribed using 0.5
µg of random primers, reverse transcriptase, 1.5 mM of dNTP and 5X
buffer from RevertAid RT Reverse Transcription kit (Fermentas;
Thermo Fisher Scientific, Inc.) at 42°C for 1 h. Once cDNA was
synthesized, ALK1, ALK2, ALK3, ALK4 and ALK5 expression levels were
amplified using the AccuPower RT-PCR PreMix kit (Bioneer
Corporation, Daejeon, Korea) with the following primer sequences:
Slug forward, 5′-GGTCAAGAAGCATTTCAAC-3′ and reverse,
5′-GGTAATGTGTGGGTCCGA-3′; Snail forward, 5′-CAGACCCACTCAGATGTCAA-3′
and reverse, 5′-CATAGTTAGTCACACCTCGT-3′; ALK1 forward,
5′-GTGGCAGCAAGTCACAAAGG-3′ and reverse, 5′-TACCAACTTGCCGGACCATC-3′;
ALK2 forward, 5′-CTCGGCTCTGTCCAGTTTGT-3′ and reverse,
5′-TCACTGGGGTACTCGGAGAG-3′; ALK3 forward,
5′-TCTCCAAAAGCCCAGCTACG-3′ and reverse, 5′-CCTATGACAACAGGGGGCAG-3′;
ALK4 forward, 5′-GTGGTGACGTGGCTGTGAAA-3′ and reverse,
5′-TTTGGAGCAATGTCTATGGT-3′; ALK5 forward, 5′-ATCCCAACAGATGGCAGAG-3′
and reverse, 5′-GGAGAGTTCAGGCAAAGCTG-3′; and GAPDH forward,
5′-GTGTTCCTACCCCCAATGTGT-3′ and reverse,
5′-ATTGTCATACCAGGAAATGAGCTT-3′. GAPDH primers were used as an
internal control. PCR was performed for 30 cycles consisting of 30
sec denaturation at 94°C, 30 sec annealing at 56°C and 30 sec
extension at 72°C. Each PCR was performed in triplicate. The PCR
products were separated by 1% w/v agarose gel electrophoresis,
stained with GelRed (Jomar Life Research, Melbourne, Victoria,
Australia), and then the band intensities were quantitated by
ImageJ software (version 1.51j8; National Institutes of Helath,
Bethesda, MD, USA).
Western blot analysis
MCF-7 and MDA-MB-231 were treated with either PBS or
10 µM eupatolide and lysed using a buffer containing 150 mM NaCl,
1.0% NP-40, 50 mM Tris-HCl at pH 8.0, 1 mM EDTA, protease
inhibitors and phosphatase inhibitors (Roche Applied Science,
Penzberg, Germany). The concentration of protein was determined
using the BCA protein assay kit (Pierce; Thermo Fisher Scientific,
Inc.), the 10 µg of prepared whole-cell lysate samples were
separated according to their size on 10% SDS-PAGE, and proteins
were transferred onto a 0.45-µm pore-size nitrocellulose membrane
for 2 h at room temperature. The membrane was blocked with 3% BSA
in TBS-T for 30 min at 25°C and incubated for overnight at 4°C with
the 1:1,000 dilution of targeted primary antibody (anti-vimentin,
anti-GAPDH, anti-E-cadherin, anti-Snail, anti-Slug, anti-SMAD3,
anti-p-SMAD3 S423/425, anti-SMAD1/5, anti-p-SMAD1/5 S423/425,
anti-SMAD4, anti-p-ERK1/2 T202/Y204, anti-ERK1/2, anti-p-AKT S473,
anti-AKT, anti-SP1, anti-p-SP1 T453 and anti-β-actin antibodies).
The horseradish peroxidase-conjugated goat anti-mouse (cat. no.
A90-131P) or goat anti-rabbit (cat. no. A120-118P) antibodies
(Bethyl Laboratories, Montgomery, TX, USA) at 1:3,000 dilution in
3% BSA in TBS-T were incubated with the membrane at 25°C for 2 h,
and then the membrane were washed 3 times for 5 min with TBS-T
buffer. The membranes were developed by ECL reagent (GE Healthcare)
and bands were detected using the LAS-3000 Image Reader (Fujifilm,
Tokyo, Japan). The protein bands were quantitated by ImageJ
software (version 1.51j8; National Institutes of Health).
Statistical analysis
Values are presented as the mean ± standard
deviation. Multiple comparisons between groups were performed by
one-way analysis of variance (ANOVA). P<0.05 was considered to
indicate a statistically significant difference. All statistical
analyses were performed using GraphPad Prism 5.0 (GraphPad
Software, La Jolla, CA, USA).
Results
Eupatolide suppresses the migration
and invasion of breast cancer cells
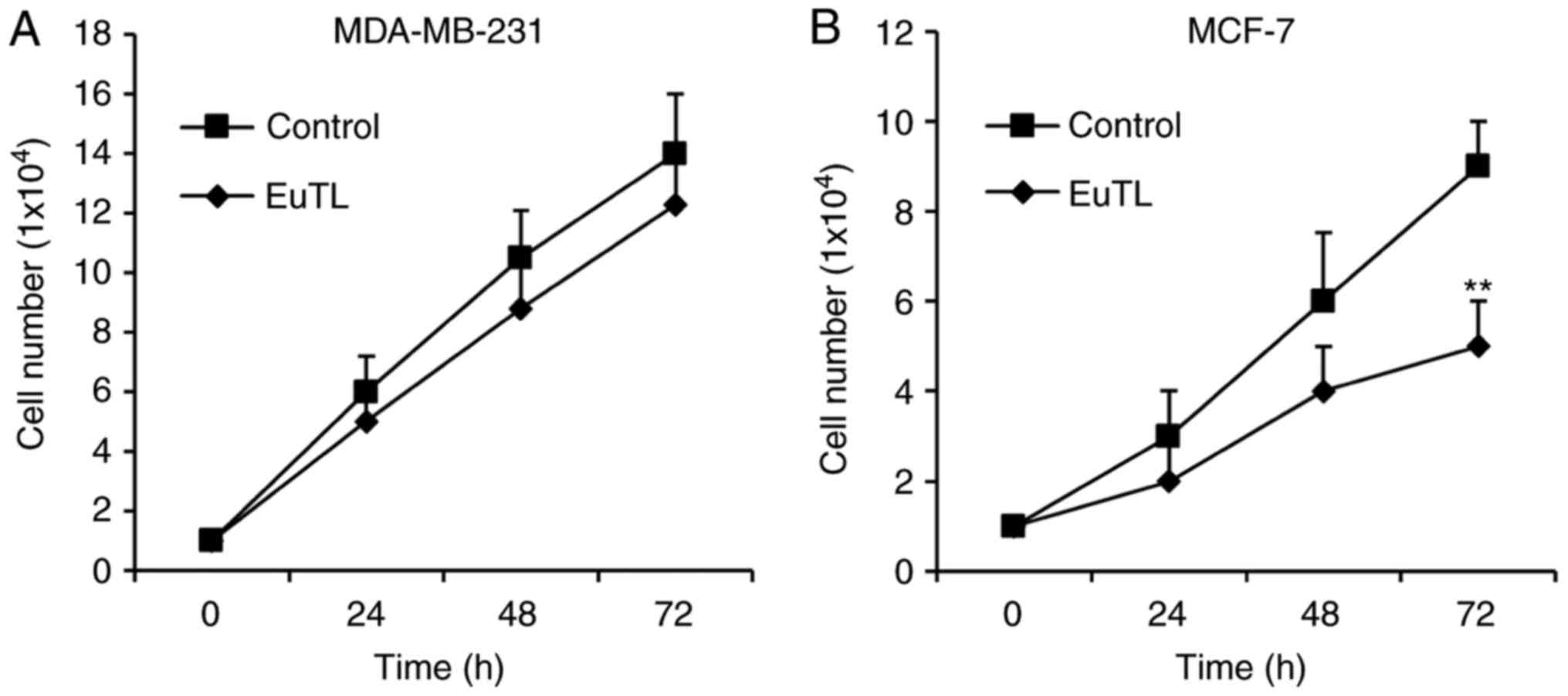
To examine whether eupatolide has an
anti-proliferative effect on breast cancer cells, estrogen
receptor-positive MCF-7 breast cancer cells and estrogen
receptor-negative MDA-MB-231 cells were treated with eupatolide for
3 days, with cell numbers measured each day. To determine the
statistical significance of changes in cell numbers during the 3
days of treatment, ANOVA was performed. In MDA-MB-231 cells, no
significant difference in proliferation rate was observed between
those treated with eupatolide or PBS (Fig. 1A); however, MCF-7 cells treated with
eupatolide exhibited a significantly decreased cell number compared
with the control on the third day of treatment (P<0.01; Fig. 1B).
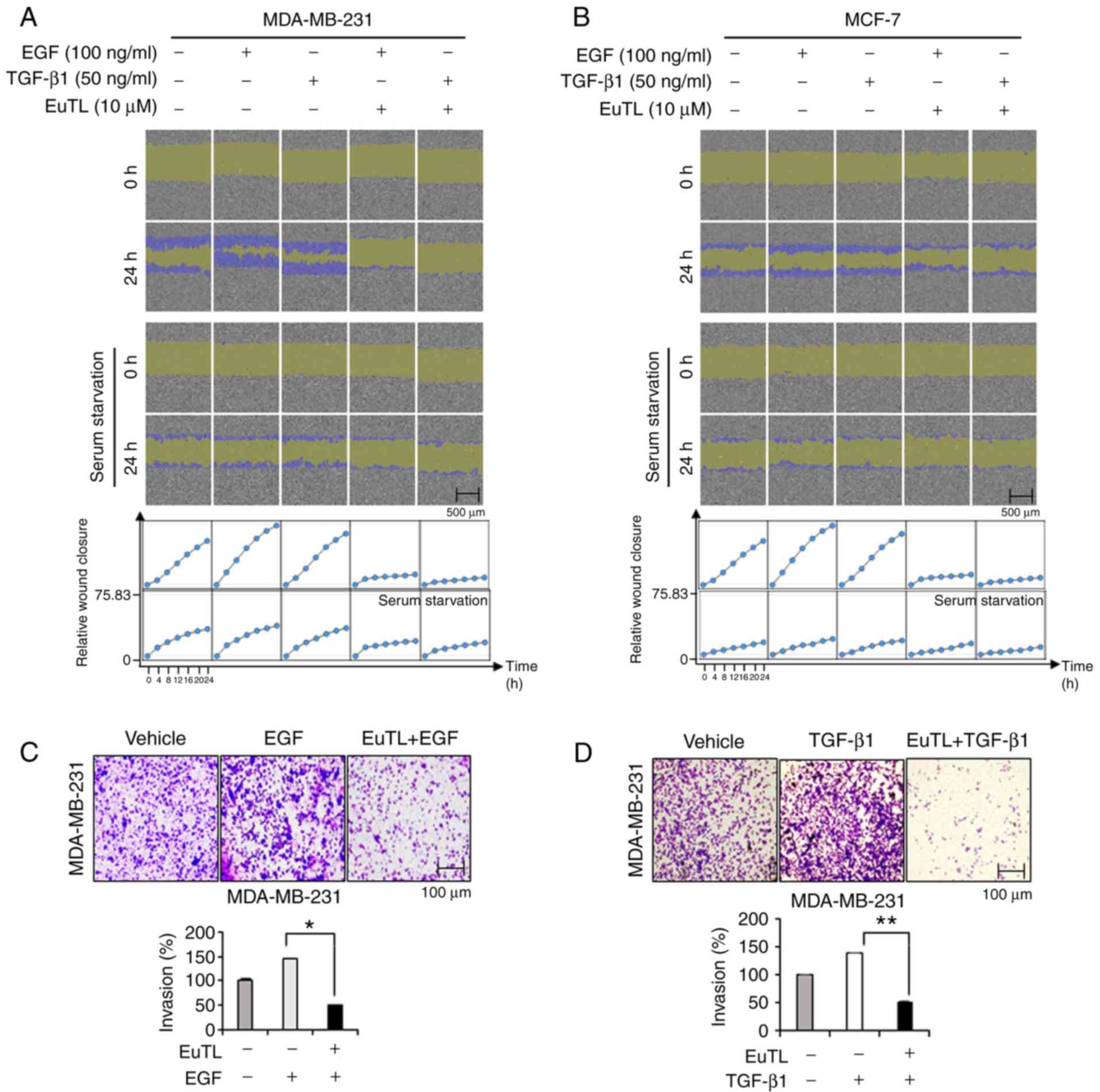
To examine whether eupatolide has an anti-migratory
effect on breast cancer cells stimulated with EGF and TGF-β1
(critical growth factors that can induce EMT and metastasis)
(26), a confluent monolayer of cells
was scratched and then treated with eupatolide for 24 h. As the
migration of MCF-7 cells could be affected by the
anti-proliferative effect of eupatolide, cells were treated with
eupatolide for 24 h only to exclude this influence, and analyzed
using the IncuCyte live-cell imaging analyzer. EGF and TGF-β1
treatment greatly increased the migration of cells compared that in
the untreated control group, and treatment with eupatolide clearly
inhibited the migration induced by EGF and TGF-β1 (Fig. 2A and B). MCF-7 cells migrated more
slowly than MDA-MB-231 cells (Fig.
2B); the slow migration of MCF-7 cells was possibly due to
their non-metastatic nature (27,28).
Whether eupatolide also affects the migration of
cells under serum-starvation conditions was assessed. MDA-MB-231
and MCF-7 cells arrested at G0 phase were not
significantly migrated compared to non-arrested cells. These
findings indicated that eupatolide more efficiently inhibited the
migration of growth factor-stimulated cells than unstimulated
cells. The result was quantified by measuring cell density in the
wound area at 2-h intervals for 24 h (Fig. 2A and B). To confirm this effect on
migration further, an invasion assay was performed using Transwell
plates. Because MCF-7 cells are non-metastatic, only MDA-MB-231
cells were used for the invasion assay. MDA-MB-231 cells were
loaded into Transwell chambers and treated with EGF and TGF-β1 as
well as eupatolide or control. EGF and TGF-β1 treatment increased
the invasion of MDA-MB-231 cells, but co-treatment with eupatolide
markedly reduced EGF- and TGF-β1-enhanced invasion (Fig. 2C and D). Eupatolide exhibited an
anti-proliferative effect in non-metastatic MCF-7 cells and an
anti-migration ability in metastatic MDA-MB-231 cells. However, the
anti-migratory effect of eupatolide is unlikely to be caused by the
anti-proliferative effect in MCF-7 cells, as the anti-proliferative
effect only reached significance on the third day following the
treatment with eupatolide.
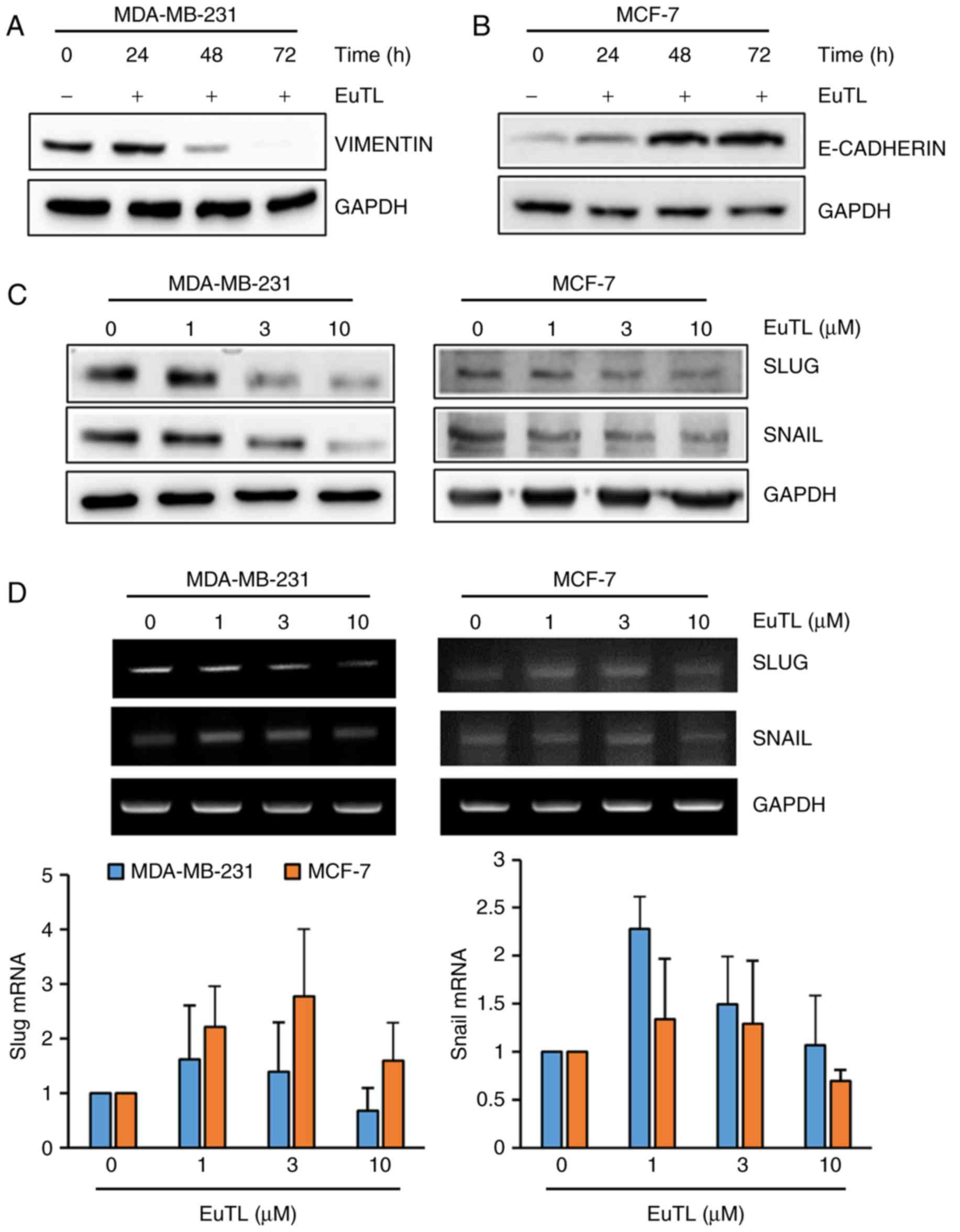
Eupatolide suppresses EMT by blocking
the expression of Snail and Slug
Since eupatolide inhibited the migration of breast
cancer cells, the expression of genes associated with EMT was
assessed. Since vimentin maintains the cytoskeletal architecture
and tissue integrity, it is involved in migration of cells.
Therefore, MDA-MB-231 cells were treated with eupatolide for 24, 48
or 72 h at a concentration of 10 µM to examine the effect on
vimentin expression. Eupatolide treatment markedly inhibited
vimentin expression in a time-dependent manner (Fig. 3A). The adherens junction protein
E-cadherin contributes to the maintenance of the epithelial barrier
function by homotypic interactions (29). Since MCF-7 cells express much higher
levels of E-cadherin than MDA-MB-231 cells (30), MCF-7 cells were treated with
eupatolide. Treatment with eupatolide increased E-cadherin
expression in a time-dependent manner (Fig. 3B), indicating that eupatolide-mediated
E-cadherin expression contributed to the suppression of cell
migration. Subsequently, whether eupatolide could inhibit the
expression level of transcription factors known to regulate
EMT-associated gene expression was assessed. MDA-MB-231 and MCF-7
cells were treated with eupatolide and the expression levels of
Snail and Slug were measured by western blotting. The expression
levels of the two proteins were decreased in both types of cell in
a dose-dependent manner (Fig. 3C). As
Snail and Slug are unstable proteins (7), the mRNA levels of Snail and Slug were
also examined using RT-qPCR in duplicate. No significant difference
was observed between the eupatolide-treated and control groups;
thus, treatment with eupatolide did not affect Snail and Slug mRNA
expression in these cells (Fig. 3D),
indicating that eupatolide contributes to the destabilization of
Snail and Slug proteins.
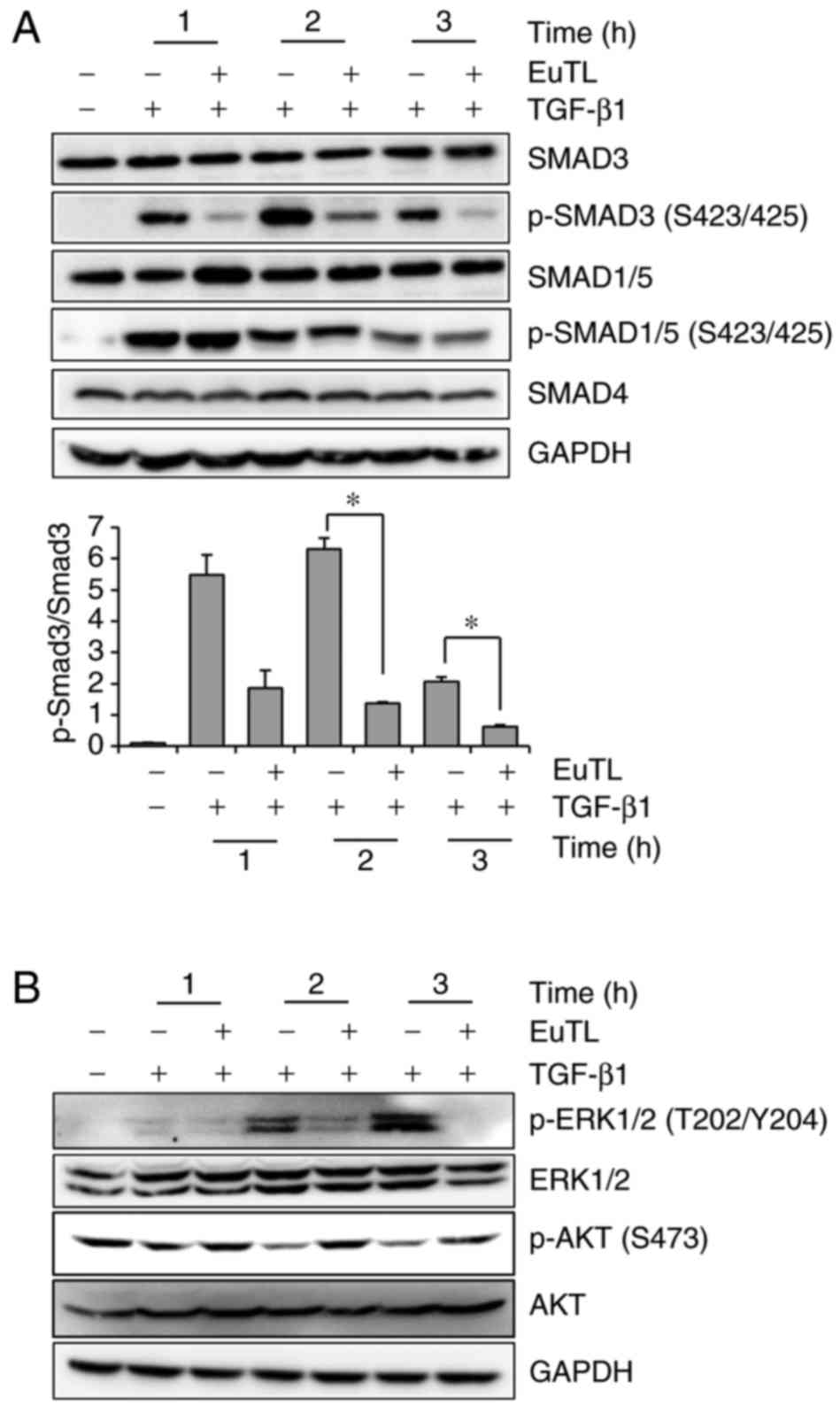
Eupatolide blocks TGF-β1-induced SMAD3
activation
To investigate which signaling pathway was targeted
by eupatolide, the SMAD signaling pathway was examined, as
TGF-β1-mediated SMAD3 activation increases Snail expression
(31). MDA-MB-231 cells were treated
with TGF-β1 in the presence of eupatolide and then examined by
western blotting to determine the level of phospho-SMAD3 and
phospho-SMAD1/5. The levels of phospho-SMAD3, which were increased
by TGF-β1 treatment, were decreased by eupatolide treatment in a
time-dependent manner (Fig. 4A).
However, SMAD1/5 phosphorylation was not affected by eupatolide
treatment. Certain non-SMAD-dependent signaling pathways are also
induced by TGF-β in EMT. TGF-β induces the phosphorylation of
tyrosine residues on both type-I (ALK5) and type-II (ALK1) TGF-β
receptors and/or on SHC adaptor protein 1 (32). The phosphorylated tyrosine residues
recruit growth factor receptor-bound protein 2/son of sevenless
homolog (GRB2/SOS) to activate ERK through RAS proto-oncogene,
GTPase (RAS), Raf-1 proto-oncogene, serine/threonine kinase (RAF)
and their downstream mitogen-activated protein kinase (MAPK)
cascades. ERK then regulates target gene transcription through
downstream transcription factors, in conjunction with SMADs, to
control EMT (33). To investigate
whether eupatolide affects these signaling pathways, levels of
phospho-ERK were examined. Pre-treatment with eupatolide 30 min
prior to TGF-β1 treatment inhibited ERK phosphorylation at 1 h
after treatment (Fig. 4B), indicating
that eupatolide also inhibited the TGF-β1-mediated non-SMAD
pathway. The PI3K/AKT pathway is another non-SMAD pathway that
contributes to TGF-β-induced EMT (15). Thus, whether eupatolide affected the
AKT signaling pathway was investigated. Contrary to what was
expected, TGF-β1 inhibited the basal level of AKT phosphorylation
at the S473 residue, which was recovered by incubation with
eupatolide (Fig. 4B).
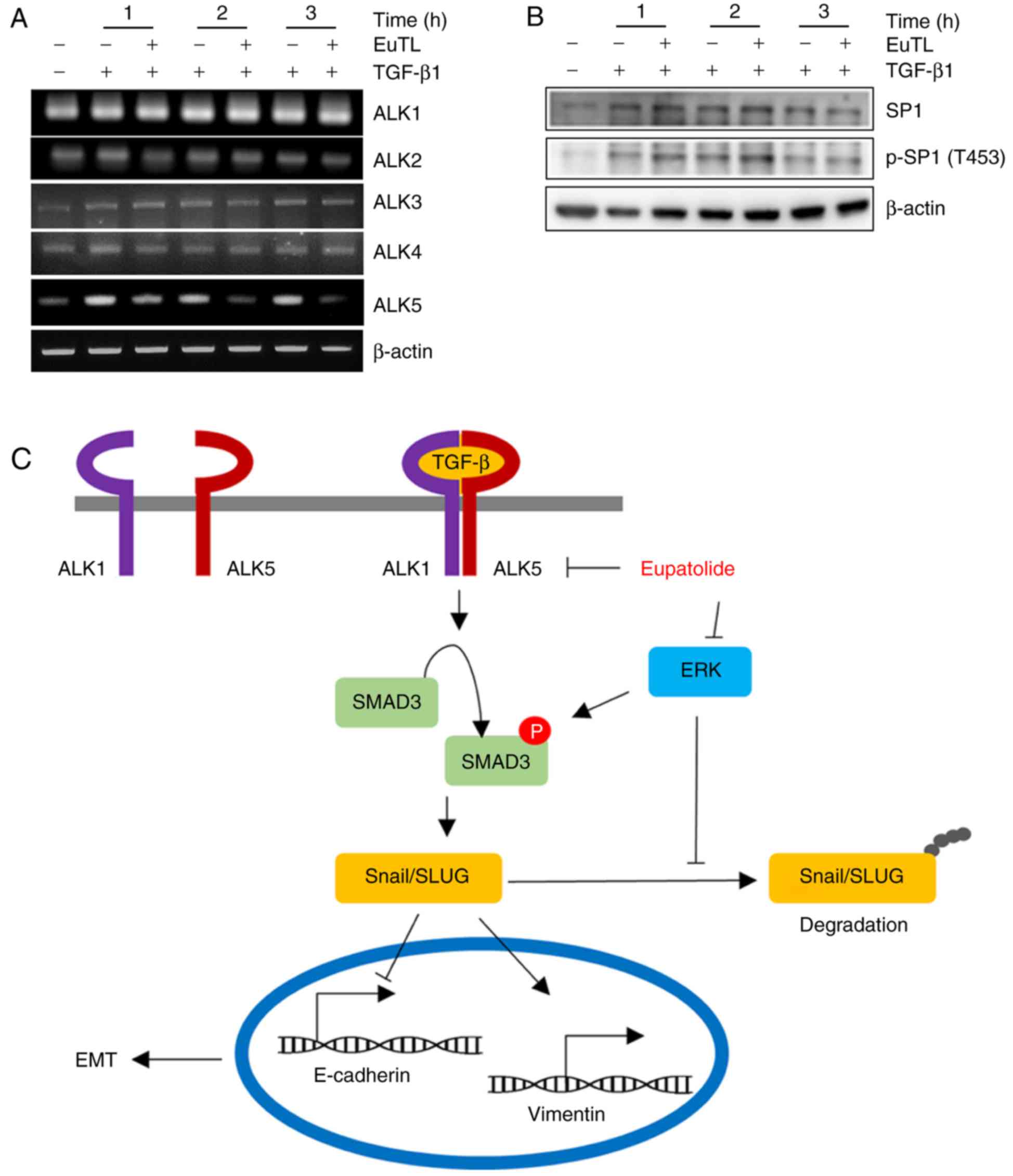
Eupatolide represses TGF-β1-induced
ALK5 expression
Since SMAD3 activation is mediated by the
heterodimerization of ALK5 and ALK1, the transcript levels of ALK1,
ALK2, ALK3, ALK4, and ALK5 were examined following eupatolide
treatment. TGF-β1 treatment increased the level of ALK5
transcription, which was decreased upon co-treatment with
eupatolide in a time-dependent manner. However, co-treatment with
eupatolide and TGF-β1 did not affect the levels of ALK1, ALK2, ALK3
and ALK4 transcription (Fig. 5A). As
TGF-β is a potent enhancer of cell migration and invasion, and ALK5
is the critical receptor component of the TGF-β receptor
heterodimer, the inhibition of ALK5 transcription by eupatolide is
potentially a critical mechanism for the suppression of the TGF-β1
signaling pathway (34). The Sp1
transcription factor (SP1) is known to increase ALK5 expression. To
assess whether SP1 activation was blocked by eupatolide treatment,
cells were treated with TGF-β1 and eupatolide, and the levels of
SP1 and of phospho-SP1 were examined. TGF-β1-activated SP1
phosphorylation was not affected by eupatolide treatment (Fig. 5B). We therefore hypothesize that
eupatolide inhibits TGF-β1 signaling pathway in two ways: By
inhibiting ALK5 transcription and TGF-β1-activated ERK expression,
leading to Snail and Slug degradation (Fig. 5C). Thus, eupatolide could be used as
an inhibitor of the TGF-β1 signaling pathway, leading to the
suppression of EMT.
Discussion
The majority of carcinomas are of epithelial origin
and capable of metastasizing to other organs (35). For this process, epithelial tumor
cells acquire mesenchymal characteristics and lose epithelial
characteristics in the process of EMT. This is required for cells
to translocate to other organs, mediated via an increase in the
migration and invasion of tumor cells. As cancer cells lose
epithelial characteristics during EMT, it is worthwhile examining
whether eupatolide affects the epithelial phenotype of cancer
cells. Since eupatolide treatment inhibits the migration and
invasion of breast cancer cells by increasing E-cadherin expression
and decreasing expression of vimentin, we hypothesize that
treatment of eupatolide inhibits the metastasis of cancer cells by
enhancement of the epithelial phenotype.
Eupatolide is a sesquiterpene lactone that can be
isolated from traditional herbal extracts, and which exhibits
various biological functions, including anti-inflammatory,
anti-proliferative and anti-migratory activities (19,20,36). Since
eupatolide inhibits the PDGF-mediated proliferation and migration
of vascular smooth muscle cells through HO-1 induction via the
ROS-NRF2 pathway (20), it is
conceivable that eupatolide may inhibit the migration of breast
cancer cells regardless of their estrogen receptor expression
status. In a previous study, it was shown that eupatolide
sensitizes breast cancer cells to TRAIL-induced apoptosis (19); it is therefore possible that the
eupatolide-mediated inhibition of migration in cancer cells could
be caused by the suppression of cell proliferation. The present
study indicates that eupatolide inhibited the proliferation of
MCF-7 cells on day 3 after treatment, although this did not occur
in MDA-MB-231 cells. As MCF-7 cells are non-metastatic, whereas
MDA-MB-231 cells are, it is likely that the inhibition of migration
in metastatic cells is not caused by the inhibition of
proliferation. To prove this hypothesis, various non-metastatic and
metastatic breast cancer cells require examination.
The TGF-β ligand superfamily is classified into
three subgroups, TGF-βs, activins and bone morphogenetic proteins
(33,37), and there are five type-II receptors
and seven different type-I receptors, termed ALK1-ALK7 (38). Binding of each TGF-β ligand induces
type II-type I receptor heterodimerization and the combination of
type II-type I receptor is dependent on a binding ligand type.
After the receptor heterodimerization, constitutively active type
II receptor phosphorylates and activates the type I receptor. The
specificity and versatility of TGF-β signaling depend on which
SMADs are activated (38). Receptor
complexes containing ALK1, ALK2, ALK3 and ALK6 phosphorylate SMAD1,
SMAD5 and SMAD8 (39), whereas
receptor complexes containing ALK4, ALK5, and ALK7 phosphorylate
SMAD2 and SMAD3 (40). In the present
study, eupatolide suppressed SMAD3 phosphorylation but did not
affect SMAD1/5 phosphorylation. Thus, ALK4, ALK5 and ALK7 may be
upstream kinases, rather than ALK1, ALK2, ALK3 and ALK6.
Transcriptional analysis revealed that eupatolide specifically
suppressed ALK5 transcription, even though a transcription factor
responsible for downregulation of ALK5 transcription was not
identified.
TGF-β antisense oligonucleotides have been used
widely in preclinical studies for cancer therapy (41). Treatment with AP-12009, an antisense
oligonucleotide against TGF-β2, resulted in a significant increase
in survival time (42) another
antisense oligonucleotide targeted against TGF-β1, AP-11014, is
being developed for treatment of human non-small cell lung,
colorectal and prostate cancer (43).
In addition to these relatively large TGF-β inhibitors, developing
small molecular TGF-β inhibitors that are selectively targeted to
inhibit ALK5 is of interest, as they could inhibit all TGF-β
isoforms (44), unlike, for instance,
antisense oligonucleotides that only target a specific TGF-β
isoform. In addition to the SMAD signal transduction pathway, TGF-β
also activates other intracellular signaling pathways, including
the MAPK, PI3K/AKT, and Rho-like GTPase pathways (45). TGF-β phosphorylates its receptors on
tyrosine residues, leading to the recruitment of GRB2/SOS to
activate ERK through RAS proto-oncogene/Raf-1 proto-oncogene
(RAS/RAF) cascades (46,47). Activated ERK then stimulates
TGF-β-target gene transcription in conjunction with SMADs, also
stabilizing Snail and Slug to control EMT (48). In the present study, eupatolide
suppressed ERK activation, which could contribute to the
destabilization of Snail and Slug. Therefore, eupatolide is a
promising candidate small anti-TGF-β inhibitor, as it is able to
inhibit TGF-β-mediated Snail and Slug stabilization via ERK and
ALK5 transcription.
The PI3K/AKT pathway is another non-SMAD pathway
that contributes to TGF-β-induced EMT (33). In numerous other TGF-β-induced
responses, the activation of PI3K or AKT antagonizes TGF-β-induced
apoptosis and growth inhibition via a physical interaction between
AKT and SMAD3 (37,38), which is independent of AKT
phosphorylation. The interaction between AKT and SMAD3 prevents
ALK5-mediated phosphorylation and the nuclear localization of
SMAD3, which leads to the inhibition of SMAD3-mediated
transcription (31,32). It means that PI3K/AKT can negatively
regulate TGF-β-induced SMADs signaling pathways (49). The present study found that TGF-β
inhibited the basal level of AKT phosphorylation rather than
activating AKT, whereas treatment with eupatolide increased AKT
phosphorylation. Thus, the eupatolide-mediated increase in AKT
phosphorylation may enhance the degree of interaction between AKT
and SMAD3, which contributes to the inhibition of TGF-β signal
transduction. On the other hand, AKT activation is known to inhibit
glycogen synthase kinase-3β (GSK-3β), leading to the stabilization
of Snail and Slug. However, treatment with eupatolide destabilized
Snail and Slug, with a corresponding activation of AKT. This
discrepancy can be explained as follows: AKT binds directly to
unphosphorylated SMAD3 regardless of AKT phosphorylation, leading
to the prevention of SMAD3 activation. Thus, regardless of the
eupatolide-mediated increase in AKT phosphorylation, if
phosphorylated AKT and SMAD3 interaction is enhanced, AKT
phosphorylation can contribute to the inhibition of the TGF-β
signaling pathway. Conversely, Snail destabilization with AKT
activation upon eupatolide treatment may be explained if GSK-3β
inactivation upon eupatolide treatment can be affected more by ERK
inhibition than AKT activation.
As traditional herbal extracts have been used to
treat cancer in East Asia, it is necessary to provide scientific
evidence that these traditional anticancer therapies are effective.
The present study reveals a partial molecular mechanism by which
eupatolide inhibits the migration and invasion of breast tumor
cells. However, one limitation of this study is that effect of
eupatolide on the inhibition of cell migration was only assessed
in vitro, owing to a lack of available eupatolide for the
conduction of an in vivo experiment. If similar results are
observed in an animal model, the development of eupatolide-derived
chemicals to prevent cancer metastasis may be worthwhile.
Acknowledgements
The present study was supported by grants from the
National Research Foundation of Korea grant funded by the Korean
government (no. NRF-2016R1A2B2011683) and the Science Research
Center of the National Research Foundation funded by the Korean
government (no. 2016R1A5A1011974).
References
|
1
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: New insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchytnal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Singh A and Settleman J: EMT, cancer stem
cells and drug resistance: An emerging axis of evil in the war on
cancer. Oncogene. 29:4741–4751. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tam WL and Weinberg RA: The epigenetics of
epithelial-mesenchymal plasticity in cancer. Nat Med. 19:1438–1449.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin YC, Lee YC, Li LH, Cheng CJ and Yang
RB: Tumor suppressor SCUBE2 inhibits breast-cancer cell migration
and invasion through the reversal of epithelial-mesenchymal
transition. J Cell Sci. 127:85–100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu Y and Zhou BP: Snail: More than EMT.
Cell Adh Migr. 4:199–203. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Marcucci F, Stassi G and De Maria R:
Epithelial-mesenchymal transition: A new target in anticancer drug
discovery. Nat Rev Drug Discov. 15:311–325. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Heldin CH, Vanlandewijck M and Moustakas
A: Regulation of EMT by TGFβ in cancer. FEBS Lett. 586:1959–1970.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wakefield LM and Roberts AB: TGF-beta
signaling: Positive and negative effects on tumorigenesis. Curr
Opin Genet Dev. 12:22–29. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kubiczkova L, Sedlarikova L, Hajek R and
Sevcikova S: TGF-β - an excellent servant but a bad master. J
Transl Med. 10:1832012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cunha SI and Pietras K: ALK1 as an
emerging target for antiangiogenic therapy of cancer. Blood.
117:6999–7006. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Whitman M and Raftery L: TGFbeta signaling
at the summit. Development. 132:4205–4210. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Curado F, Spuul P, Egaña I, Rottiers P,
Daubon T, Veillat V, Duhamel P, Leclercq A, Gontier E and Génot E:
ALK5 and ALK1 play antagonistic roles in transforming growth factor
β-induced podosome formation in aortic endothelial cells. Mol Cell
Biol. 34:4389–4403. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang YE: Non-Smad pathways in TGF-beta
signaling. Cell Res. 19:128–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schniewind B, Groth S, Müerköster S
Sebens, Sipos B, Schäfer H, Kalthoff H, Fändrich F and Ungefroren
H: Dissecting the role of TGF-beta type I receptor/ALK5 in
pancreatic ductal adenocarcinoma: Smad activation is crucial for
both the tumor suppressive and prometastatic function. Oncogene.
26:4850–4862. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Khan AL, Hussain J, Hamayun M, Gilani SA,
Ahmad S, Rehman G, Kim YH, Kang SM and Lee IJ: Secondary
metabolites from Inula britannica L. and their biological
activities. Molecules. 15:1562–1577. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee YH, Lee NK and Paik HD: Antimicrobial
characterization of inula britannica against helicobacter pylori on
gastric condition. J Microbiol Biotechnol. 26:1011–1017. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee J, Hwangbo C, Lee JJ, Seo J and Lee
JH: The sesquiterpene lactone eupatolide sensitizes breast cancer
cells to TRAIL through down-regulation of c-FLIP expression. Oncol
Rep. 23:229–237. 2010.PubMed/NCBI
|
|
20
|
Kim N, Hwangbo C, Lee S and Lee JH:
Eupatolide inhibits PDGF-induced proliferation and migration of
aortic smooth muscle cells through ROS-dependent heme oxygenase-1
induction. Phytother Res. 27:1700–1707. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hugo H, Ackland ML, Blick T, Lawrence MG,
Clements JA, Williams ED and Thompson EW: Epithelial-mesenchymal
and mesenchymal-epithelial transitions in carcinoma progression. J
Cell Physiol. 213:374–383. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee S, Jeong AL, Park JS, Han S, Jang CY,
Kim KI, Kim Y, Park JH, Lim JS, Lee MS and Yang Y: IK-guided PP2A
suppresses Aurora B activity in the interphase of tumor cells. Cell
Mol Life Sci. 73:3375–3386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jin HZ, Lee D, Lee JH, Lee K, Hong YS,
Choung DH, Kim YH and Lee JJ: New sesquiterpene dimers from Inula
britannica inhibit NF-kappaB activation and NO and TNF-alpha
production in LPS-stimulated RAW264.7 cells. Planta Med. 72:40–45.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee S, Han S, Jeong AL, Park JS and Yang
Y: Depletion of IK causes mitotic arrest through aberrant
regulation of mitotic kinases and phosphatases. FEBS Lett.
588:2844–2850. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sinha C, Ren A, Arora K, Moon CS,
Yarlagadda S, Zhang W, Cheepala SB, Schuetz JD and Naren AP:
Multi-drug resistance protein 4 (MRP4)-mediated regulation of
fibroblast cell migration reflects a dichotomous role of
intracellular cyclic nucleotides. J Biol Chem. 288:3786–3794. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li L, Qi L, Liang Z, Song W, Liu Y, Wang
Y, Sun B, Zhang B and Cao W: Transforming growth factor-β1 induces
EMT by the transactivation of epidermal growth factor signaling
through HA/CD44 in lung and breast cancer cells. Int J Mol Med.
36:113–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guttilla IK, Phoenix KN, Hong X, Tirnauer
JS, Claffey KP and White BA: Prolonged mammosphere culture of MCF-7
cells induces an EMT and repression of the estrogen receptor by
microRNAs. Breast Cancer Res Treat. 132:75–85. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vergara D, Valente CM, Tinelli A,
Siciliano C, Lorusso V, Acierno R, Giovinazzo G, Santino A,
Storelli C and Maffia M: Resveratrol inhibits the epidermal growth
factor-induced epithelial mesenchymal transition in MCF-7 cells.
Cancer Lett. 310:1–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
van Roy F and Berx G: The cell-cell
adhesion molecule E-cadherin. Cell Mol Life Sci. 65:3756–3788.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chao YL, Shepard CR and Wells A: Breast
carcinoma cells re-express E-cadherin during mesenchymal to
epithelial reverting transition. Mol Cancer. 9:1792010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kocic J, Bugarski D and Santibanez JF:
SMAD3 is essential for transforming growth factor-β1-induced
urokinase type plasminogen activator expression and migration in
transformed keratinocytes. Eur J Cancer. 48:1550–1557. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pannu J, Nakerakanti S, Smith E, ten Dijke
P and Trojanowska M: Transforming growth factor-beta receptor type
I-dependent fibrogenic gene program is mediated via activation of
Smad1 and ERK1/2 pathways. J Biol Chem. 282:10405–10413. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wrighton KH, Lin X and Feng XH:
Phospho-control of TGF-beta superfamily signaling. Cell Res.
19:8–20. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cantelli G, Crosas-Molist E, Georgouli M
and Sanz-Moreno V: TGFBeta-induced transcription in cancer. Semin
Cancer Biol. 42:60–69. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Seyfried TN and Huysentruyt LC: On the
origin of cancer metastasis. Crit Rev Oncog. 18:43–73. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee J, Tae N, Lee JJ, Kim T and Lee JH:
Eupatolide inhibits lipopolysaccharide-induced COX-2 and iNOS
expression in RAW264.7 cells by inducing proteasomal degradation of
TRAF6. Eur J Pharmacol. 636:173–180. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schmierer B and Hill CS: TGFbeta-SMAD
signal transduction: Molecular specificity and functional
flexibility. Nat Rev Mol Cell Biol. 8:970–982. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Feng XH and Derynck R: Specificity and
versatility in tgf-beta signaling through Smads. Annu Rev Cell Dev
Biol. 21:659–693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Daly AC, Randall RA and Hill CS:
Transforming growth factor beta-induced Smad1/5 phosphorylation in
epithelial cells is mediated by novel receptor complexes and is
essential for anchorage-independent growth. Mol Cell Biol.
28:6889–6902. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Valcourt U, Kowanetz M, Niimi H, Heldin CH
and Moustakas A: TGF-beta and the Smad signaling pathway support
transcriptomic reprogramming during epithelial-mesenchymal cell
transition. Mol Biol Cell. 16:1987–2002. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Connolly EC, Freimuth J and Akhurst RJ:
Complexities of TGF-β targeted cancer therapy. Int J Biol Sci.
8:964–978. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schlingensiepen KH, Schlingensiepen R,
Steinbrecher A, Hau P, Bogdahn U, Fischer-Blass B and Jachimczak P:
Targeted tumor therapy with the TGF-beta 2 antisense compound AP
12009. Cytokine Growth Factor Rev. 17:129–139. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Akhurst RJ: Large- and small-molecule
inhibitors of transforming growth factor-beta signaling. Curr Opin
Investig Drugs. 7:513–521. 2006.PubMed/NCBI
|
|
44
|
Ling LE and Lee WC: Tgf-beta type I
receptor (Alk5) kinase inhibitors in oncology. Curr Pharm
Biotechnol. 12:2190–2202. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Smith AL, Robin TP and Ford HL: Molecular
pathways: Targeting the TGF-β pathway for cancer therapy. Clin
Cancer Res. 18:4514–4521. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pelicci G, Lanfrancone L, Grignani F,
McGlade J, Cavallo F, Forni G, Nicoletti I, Grignani F, Pawson T
and Pelicci PG: A novel transforming protein (SHC) with an SH2
domain is implicated in mitogenic signal transduction. Cell.
70:93–104. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee MK, Pardoux C, Hall MC, Lee PS,
Warburton D, Qing J, Smith SM and Derynck R: TGF-beta activates Erk
MAP kinase signalling through direct phosphorylation of ShcA. EMBO
J. 26:3957–3967. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Derynck R and Zhang YE: Smad-dependent and
Smad-independent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Guo X and Wang XF: Signaling cross-talk
between TGF-beta/BMP and other pathways. Cell Res. 19:71–88. 2009.
View Article : Google Scholar : PubMed/NCBI
|