Introduction
According to the World Health Organization cancer is
a broad term for a large group of diseases that can affect any part
of the body. It is caused by transformation of normal to malignant
cells that behave unusual and grow beyond their usual boundaries.
It is the second leading cause of death globally, and was
responsible for 8.8 million deaths in 2015 (1). Conventional chemo and radiotherapy have
many adverse effects and fail to cure many types of cancer in
humans, thus alternative therapies to treat cancer patients have
become more popular since late last century (2). Among these therapies the use of
oncolytic viruses, which have tropism to malignant cells but not to
normal cells, has been gaining field during the last decades
(3) and even a FDA-approved
virotherapy is already available since 2015 (IMLYGIC™; Amgen, Inc.,
Thousand Oaks, CA, USA). Oncolysis induced by these viruses is
mostly an immunogenic type of cancer cell death that includes
immunogenic apoptosis, necrosis and autophagic cell death (4). As consequence, oncolytic virus induces a
potent post-oncolytic antitumor activity that is considered crucial
for its efficient therapeutic activity (5).
The Newcastle disease virus (NDV) is one of the
various species of viruses that are under clinical evaluation as
vector for oncolytic tumor, gene and immune stimulation therapies
(3). It is member of the
Avulavirus genus in the Paramyxoviridae family (6) and possesses a 15,186 nucleotide negative
single strand RNA genome that encodes six genes including the
nucleocapsid protein (NP), phosphoprotein (P), matrix protein (M),
fusion protein (F), hemagglutinin-neuraminidase (HN), and
RNA-dependent RNA polymerase (L) (7).
Phylogenetic comparison of the full-length HN and F proteins shows
two major divisions represented by class I and class II. Class II
is further divided into at least 8 (I–VIII) genotypes (8) and genotypes I (such as Ulster) and II
(such as La Sota) have the NDV strains most used for virotherapy
(2,3).
Additionally, NDV is classified into three pathotypes depending on
the severity of the disease that it causes in birds: lentogenic
(avirulent), mesogenic (intermediate), or velogenic (virulent)
(7). The cleavage site in the F
protein of the NDV has been shown to be a major determinant of
virulence (9,10). In this regard, pathogenic
classification of NDV strains in birds correlates with their
oncolytic properties in cancer cells, which can be categorized as
either lytic or non-lytic, with velogenic and mesogenic viruses
being lytic and lentogenic viruses in general being non-lytic
(11).
NDV has been demonstrated to mediate its oncolytic
effect by both intrinsic and extrinsic caspase-dependent pathways
of cell death (12). NDV-induced
apoptosis is dependent on upregulation of tumor necrosis factor
(TNF)-related apoptosis-inducing ligand (TRAIL) and caspase
activation, which cause opening of mitochondrial permeability
transition pores, loss of mitochondrial membrane potential, and
subsequent activation of apoptosis process (13). Besides, NDV can also develop
additional immune stimulatory mechanisms. It is known that it
provokes the release of danger signals during virus replication
into the tumor cell cytoplasm (14–16) and
stimulates the immune system to produce cytokines such as
interferons (IFNs) or TNF (17). In
turn, these inflammatory cytokines lead to the activation of
natural killer (NK) cells, monocytes, macrophages and sensitized T
cells, stimulating both innate and adaptive immune response
(17–21).
It is known that in NDV therapy the treatment
outcomes are dependent on the kind of tumor and on the used NDV
strain (2). Besides, to avoid
limiting effects of neutralizing antibodies during virotherapy the
use of different NDV strains is considered. In this study, we
wanted to explore the capability and specificity of a genotype V
recombinant attenuated NDV vaccine, which was made from a velogenic
strain obtained from an outbreak in Puebla (Mexico) in 2005 and
denominated APMV1/Chicken/Mexico/P05/2005 (22), to induce apoptosis over a panel of
tumor cell lines and to stimulate human peripheral blood
mononuclear cells (PBMC) in vitro.
Materials and methods
Cell lines and culture
Tumor cells were kindly provided by Dr Pablo Ortiz
(HL-60 and HCC1954; CIBO, Guadalajara, Mexico), Dr Susana Del Toro
(HeLa; CUCS, Guadalajara, Mexico) and Dr Daniel Cervantes (A549 and
HepG2; UAA, Aguascalientes, Mexico). The human cell lines HeLa
(HPV-18 cervix adenocarcinoma) and HepG2 (hepatoblastoma) (23) were cultured in DMEM medium (Gibco,
Grand Island, NY, USA) supplemented with 1 g/l glucose, 4 mM
L-alanyl-glutamine, 10% heat-inactivated fetal bovine serum (FBS;
Gibco), 50 IU/penicillin and, 50 µg/ml streptomycin (Sigma-Aldrich
Israel Ltd., Rehovot, Israel); the human cell lines HCC1954 (breast
cancer), HL-60 (promyelocytic leukemia) and A549 (lung carcinoma)
were cultured in RPMI-1640 medium (Sigma-Aldrich, St. Louis, MO,
USA) supplemented with 2 g/l glucose, 2 mM L-glutamine, 25 mM
HEPES, 10% heat-inactivated FBS, 50 IU/penicillin and, 50 µg/ml
streptomycin (Sigma-Aldrich Israel Ltd.) at 37°C and 5%
CO2 in a humidified atmosphere. PBMCs were obtained from
healthy donors, after obtaining informed consent, by centrifugation
in Ficoll-Hypaque (Lymphoprep™; Axis-Shield PoC AS, Oslo, Norway)
density gradient at 832 × g for 20 min. Cells from sample/medium
interface were collected and washed three times with phosphate
buffer saline (PBS) pH 7.4 (137 mM NaCl, 2.7 mM KCl, 10 mM
Na2HPO4, 1.8 mM
KH2PO4). Cell suspension was depleted of
erythrocytes by incubation in 75 mM NH4Cl for 5 min,
washed twice in PBS and cultured with supplemented RPMI-1640
medium. The trypan blue dye exclusion assay was performed to
determine cell number and viability of PBMC.
Recombinant NDV
All experiments were carried out with a recombinant
Newcastle disease virus (rNDV-P05), from the Mexican velogenic
strain APMV1/Chicken/Mexico/P05/2005 (22,24)
obtained from a commercial attenuated live vaccine (Investigación
Aplicada, S.A. de C.V., Tehuacan, Mexico). To concentrate and
clarify the virus, one vaccine vial was diluted in 50 ml of PBS and
centrifuged at 3,300 × g at 4°C for 20 min (Hermle Z383K; Hermle
Labortechnik GmbH, Wehingen, Germany). Then the supernatant was
filtered (0.22 µm; Millipore Ireland Ltd., Cork, Ireland) and the
virus pelleted by ultracentrifugation at 150,000 × g at 4°C for 6 h
(Beckman Coulter Optima™ max-xp; Beckman Coulter, Inc.,
Brea, CA, USA) in a 20% sucrose cushion. The virus was suspended in
200 µl of PBS and its concentration and activity were determined by
hemagglutination assay. To determine optimal pH for viral activity,
PBS solutions with different pH (3, 5, 7.4 and 9) were previously
prepared and used to suspend the virus after ultracentrifugation.
To evaluate optimal temperature, the virus was diluted in PBS pH
7.4 and incubated at 22, 37, 42, 60 and 80°C for 2 h.
Hemagglutination assay
Virus concentration and hemagglutinating activity
were determined from 25 µl of ultracentrifuged rNDV-P05 sample by
incubating two-fold serial dilutions of viral samples in 96-well
microtiter plate with 1% human erythrocytes for 45 min at room
temperature. Results were expressed as hemagglutination units (HU),
where one HU represents the highest virus dilution leading to
visible hemagglutination. As positive and negative controls, a
polyvalent influenza vaccine (Flu) and PBS were used,
respectively.
Viral protein separation and
staining
A volume of 5 µl from a 10,000 HU/ml sample of
concentrated virus was resolved by 10% sodium dodecyl
sulfate-polyacrilamide gel (SDS-PAGE) using a Mini Protean III
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and then
stained with Coomassie blue R-250 staining for visual
interpretation as previously illustrated (25).
Apoptosis detection
The capacity of virus to induce apoptosis on tested
cells was determined following the instructions of a commercial
enzyme-linked immunosorbent assay (ELISA) kit (Cell Death Detection
ELISAPLUS; Roche Diagnostics GmbH, Mannheim, Germany)
for the detection of histone-associated-DNA-fragments (nucleosomes)
in cytoplasm as an indicator of late apoptosis. In brief, tumor
cell lines and PBMC were seeded in a 96-well plate at
104 cells/well in 200 µl for 24 and 48 h in presence (10
and 50 HU) or absence (PBS vehicle, control) of rNDV-P05. After
incubation, cells were lysed and centrifuged at 200 × g for 10 min
(Eppendorf 5415C; Eppendorf, New York, NY, USA), and supernatants
(cytoplasmic fraction) were evaluated for the detection of
nucleosomes by ELISA. Apoptosis levels were calculated as the
enrichment factor (EF) of nucleosomes released into the cytoplasm
as follows: EF = OD405 nm of treated cells/OD405
nm non-treated cells (control).
PBMC stimulation assay and cytokine
quantification
For human PBMC stimulation, 106 cells
were seeded in 24-well plates with 1 ml of supplemented RPMI-1640
medium. After 4–6 h acclimatization, cells were co-cultured with
rNDV-P05 (10 and 20 HU) or with an equivalent volume of PBS
(vehicle control) for 24 h. As positive control, 5 µg/ml of
lipopolysaccharide (LPS; Sigma-Aldrich) was added to cells. After
stimulation, supernatants from all groups were kept at −80°C until
utilization. Measurement of IFN-α, IFN-γ, TNF-α and TRAIL was
determined from supernatants by commercial ELISA kits [human IFN-α
platinum (eBioscience, Vienna, Austria); human IFN-γ (Thermo Fisher
Scientific, Inc., Waltham, MA, USA); human TNF-α (Life
Technologies, Merelbeke, Belgium); human TRAIL (CD253; Abcam,
Cambridge, UK)]. Absorbance was read using an iMark™
microplate reader (Bio-Rad Laboratories, Tokyo, Japan) and
concentration for each cytokine was determined carrying out a curve
with the standards supplied by the kit.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Human PBMC were stimulated as previously described,
but co-cultured with rNDV-P05 or positive/negative controls only 4
h. Total RNA was isolated from all groups using a commercial kit
Allprep RNA/protein (Qiagen GmbH, Hilden, Germany) following the
company's instructions. Purified RNA was quantified with a NanoDrop
2000 Spectrophotometer (Thermo Fisher Scientific, Inc.) with the
A260/280 ratio and only samples with ratio >1.8 were employed.
RNA (50 ng) was retrotranscripted to complementary DNA (cDNA) using
the kit SuperScript™ II Reverse Transcriptase
(Invitrogen, Carlsbad, CA, USA), and then 100 ng of cDNA were used
as template for qPCR according to SYBR-Green technology kit
(Bio-Rad Laboratories, Inc.). All qPCR experiments were carried out
in a 7500 Fast Real Time PCR system Applied Biosystems V2.0.
Specific primers for TNF-α (ID: 25952110c1), IFN-α
(ID: 11067751a1), IFN-γ (ID: 56786137c1), TRAIL (ID:
300193031c1), and β-actin (ID: 4501885a1) genes (Table I) were chosen from Primer Bank
(https://pga.mgh.harvard.edu/primerbank/) and used at a
final concentration of 120 (TNF-α and IFN-α) or 200 nM (IFN-γ,
TRAIL, and β-actin). The cycling conditions were: initial
denaturation at 95°C for 3 min, then denaturation at 95°C for 10
sec, annealing at 60°C (TNF-α), 59°C (IFN-α), 60°C (IFN-γ), 60°C
(TRAIL) and, 60°C (β-actin) for 30 sec, for 40 cycles. Target gene
expression was reported as the fold change in the normalized signal
relative to β-actin gene through the ΔΔCq method (26).
 | Table I.Oligonucleotides for gene expression
quantification. |
Table I.
Oligonucleotides for gene expression
quantification.
| Gene | GenBank
accession | Oligonucleotides
(5′>3′) |
|---|
| TNF-α | NM_000594 | F:
CCTCTCTCTAATCAGCCCTCTG |
|
|
| R:
GAGGACCTGGGAGTAGATGAG |
| IFN-α | NM_000605 | F:
GCTTGGGATGAGACCCTCCTA |
|
|
| R:
CCCACCCCCTGTATCACAC |
| IFN-γ | NM_000619 | F:
TCGGTAACTGACTTGAATGTCCA |
|
|
| R:
TCGCTTCCCTGTTTTAGCTGC |
| TRAIL | NM_001190942 | F:
TGCGTGCTGATCGTGATCTTC |
|
|
| R:
GCTCGTTGGTAAAGTACACGTA |
| β-actin | NM_001101 | F:
CATGTACGTTGCTATCCAGGC |
|
|
| R:
CTCCTTAATGTCACGCACGAT |
Statistical analysis
Data represent the mean ± SEM from at least three
independent experiments. All analyses were done with the GraphPad
Prism 5.0 software (GraphPad Software, San Diego, CA, USA).
Statistical analysis was performed by a two-sided Student's t-test,
except for comparing NDV treated against control in antitumor
assays, in which one-way ANOVA with Dunnett's post test was
employed. A P-value <0.05 was considered significant.
Results
rNDV-P05 characterization
rNDV-P05 proteins from ultracentrifuged samples were
resolved and identified through SDS-PAGE (Fig. 1A). Five major bands corresponded to
the molecular weights of the structural proteins of NDV, including
HN (75 kDa as a monomer and 150 kDa as dimer), F (65 kDa), NP
(50–55 kDa), and M (40 kDa) proteins. rNDV-P05 activity was
determined by hemagglutination assay (Fig. 1B) and 1 HU of rNDV-P05 sample was
titered at 1:128. Furthermore, in order to find out the best
conditions for rNDV handling, different pH and temperatures were
applied to virus after ultracentrifugation and then
hemagglutination assay was developed. Detailed results are
summarized in Table II.
Hemagglutination titers of rNDV changed according to pH, with the
lowest value observed at pH 3 (1:32; P<0.001), and one titer
decreased at pH 5 and pH 9 (1:64; P<0.001) compared to pH 7.4
(1:128). On the other hand, temperatures higher than 42°C totally
abolished hemagglutinating activity of the virus.
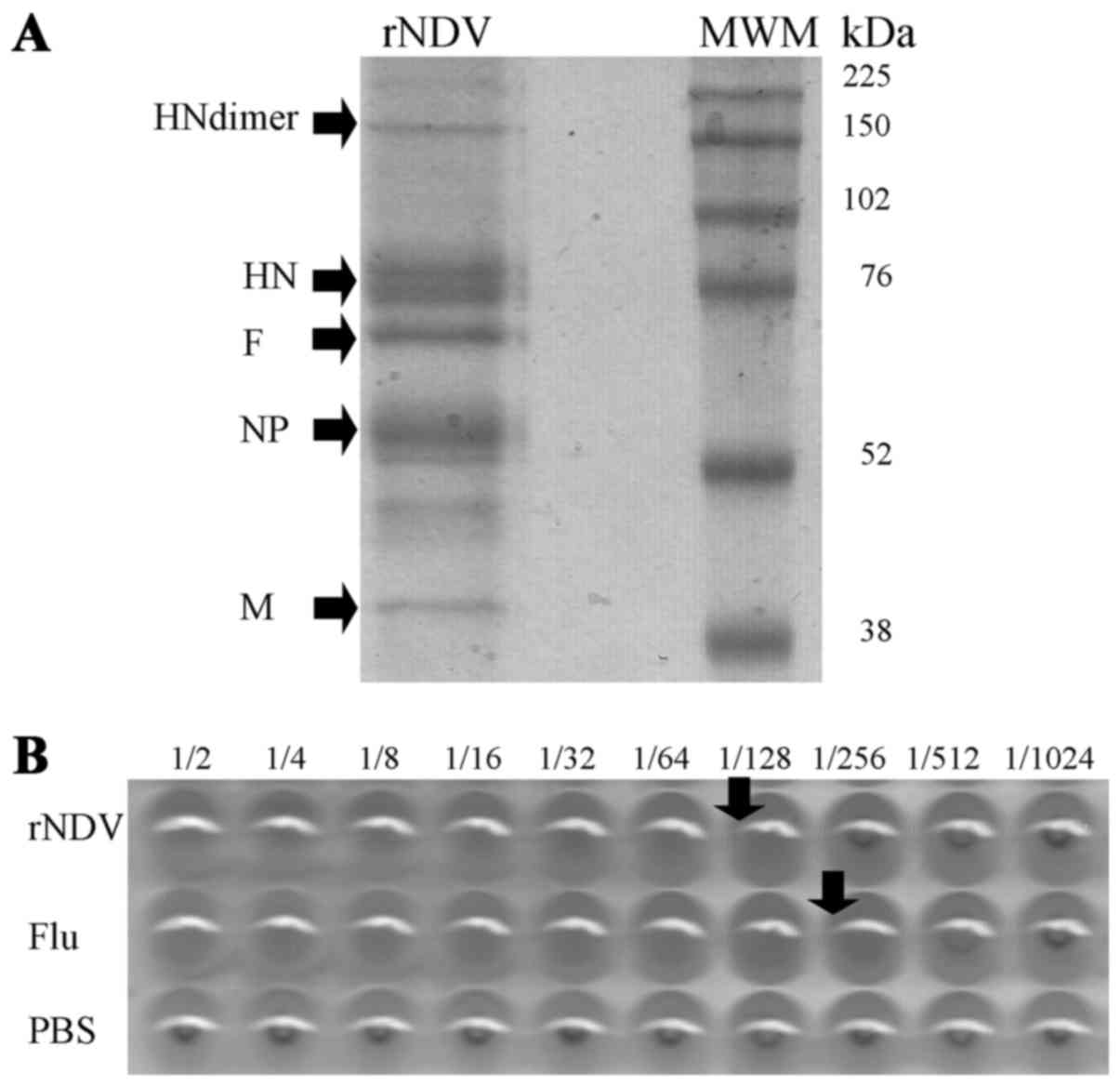 | Figure 1.Characterization of proteins and
hemagglutinating activity of rNDV. (A) Sodium dodecyl
sulfate-polyacrilamide gel from concentrated virus after
ultracentrifugation. Arrows indicate molecular weight for the major
viral protein: HN as monomer and dimer, F, NP, M. (B)
Representative hemagglutination assay for determination of activity
and concentration from rNDV-P05 stocks. PBS and Flu vaccine were
used as negative and positive controls, respectively. HN,
hemagglutinin-neuraminidase; F, fusion protein; NP, nucleocapsid
protein; M, matrix protein; MWM, molecular weight marker; kDa, kilo
Daltons; rNDV-P05, recombinant attenuated NDV strain P05; PBS,
phosphate buffer saline; Flu, polyvalent influenza vaccine. |
| Table II.Physicochemical characterization of
rNDV-P05 activity by hemagglutination assay. |
Table II.
Physicochemical characterization of
rNDV-P05 activity by hemagglutination assay.
|
| pH | Temperature
(°C) |
|---|
|
|
|
|
|---|
| Activity | 3 | 5 | 7.4 | 9 | 22 | 37 | 42 | 60 | 80 |
|---|
| Inverse of HU | 32a | 64a | 128 | 64a | 128 | 128 | 128 | 0 | 0 |
rNDV-P05 induces apoptosis in HeLa,
HCC1954, HL-60 and HepG2 cells, but not in A549 cells and PBMC
Antitumor activity of rNDV-P05 was determined as the
capacity of the virus to induce apoptosis on different tumor cell
lines. Four of the five tumor cell lines showed sensitivity to the
rNDV, mostly in a dose and time dependent way (Fig. 2). For HeLa cells, EF values increased
when cells were incubated with 10 or 50 HU of virus at both times
of incubation, in relation to control cells (P<0.001 to 10 HU at
48 h, P<0.001 to 50 HU at 24 and 48 h; Fig. 2A). The pro-apoptotic effect of 50 HU
of virus was significantly greater than that of 10 HU at 48 h
(P<0.01). Incubation time was also relevant in antitumor
activity, as 48 h showed greater effect than 24 h, both with 10
(P<0.001) or 50 HU (P<0.05) of the virus. For HCC1954 cells,
EF significantly augmented at 24 h (P<0.05 to 10 HU, P<0.001
to 50 HU) and 48 h (P<0.001) when incubated with both doses of
rNDV-P05 (Fig. 2B). This
pro-apoptotic effect was significantly greater when cells were
incubated with the highest dose of virus and during the longest
time (P<0.001). For HL-60 cells, as shown in Fig. 2C, EF values augmented when cells were
treated with 10 or 50 HU of virus for 24 and 48 h (P<0.01 and
P<0.05 to 10 HU at 24 and 48 h, P<0.001 and P<0.01 to 50
HU at 24 and 48 h) and the antiviral effect of 50 HU of the virus
was significantly greater than that of 10 HU when cells were
exposed during 24 h (P<0.05). Surprisingly, it was observed a
significant decrease in EF values as incubation time was prolonged
using both 10 HU (P<0.05) and 50 HU (P<0.01) of rNDV. In the
case of HepG2 cells, incubations with rNDV-P05 induced a
significant increment in values of EF as compared to control
conditions (P<0.01 and P<0.001 to 10 HU at 24 and 48 h,
P<0.001 to 50 HU at 24 and 48 h; Fig.
2D). The antitumor effect of rNDV was greater when HepG2 cells
were incubated with 50 HU of the virus independently of the
incubation time (P<0.05), but exposures of 48 h induced a
significant increase in pro-apoptotic effect in relation to that of
24 h only when 10 HU of the virus were used (P<0.001). Finally,
A549 cells and PBMC were not susceptible to apoptosis induction at
any time or with any used concentration of rNDV (Fig. 2E and F). All together, the results
showed that the pro-apoptotic effect of rNDV-P05 and its magnitude
is specific to particular tumor cell lines because there is not an
evident effect on A549 cell line. Besides, no pro-apoptotic effect
was observed on PBMC from healthy donors, that is important to
study subsequently the immunostimulatory activity of the virus on
PBMC.
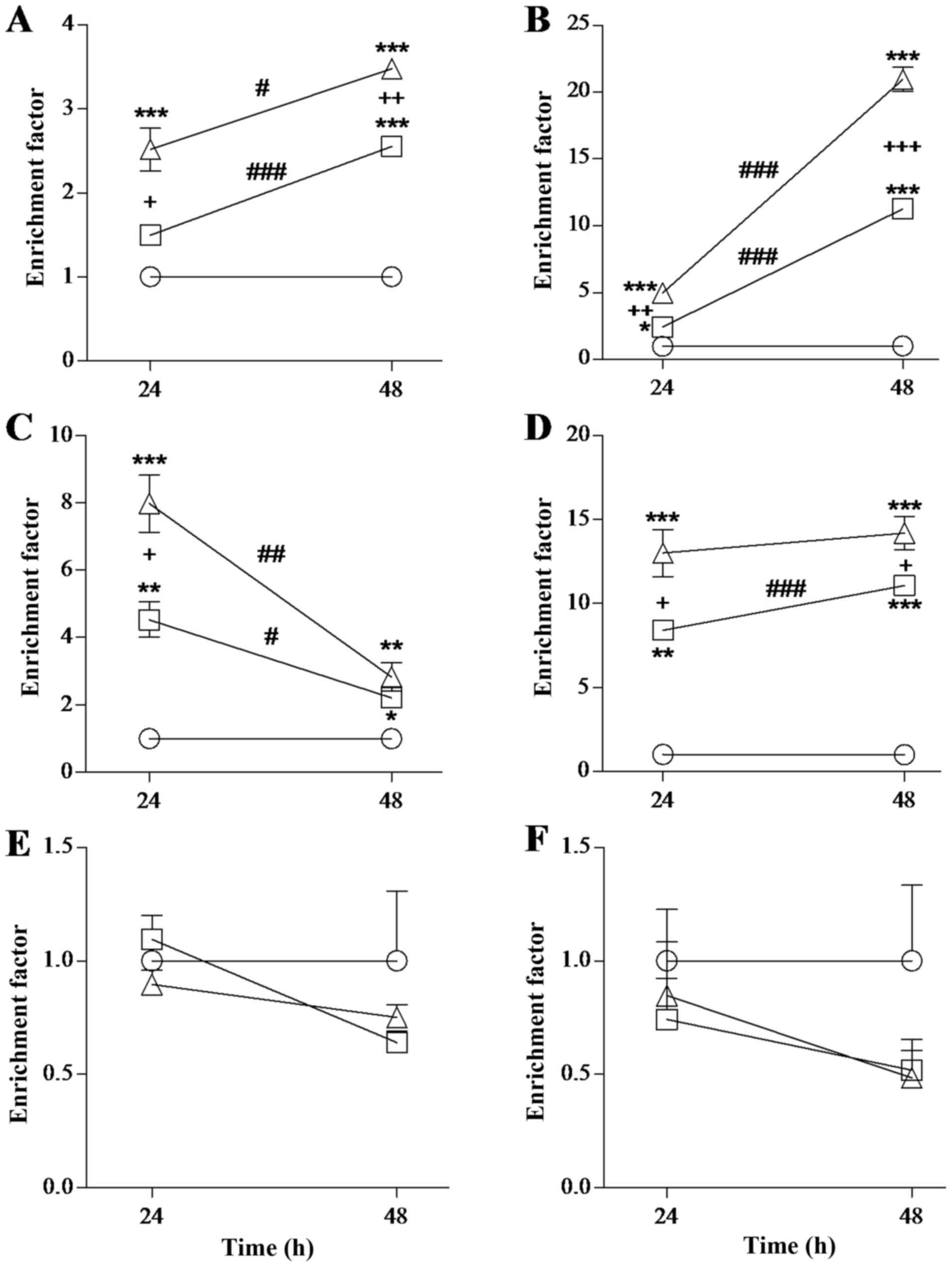 | Figure 2.Evaluation of rNDV antitumor activity
over different cell lines. (A) HeLa cells, (B) HCC1954 cells, (C)
HL-60 cells, (D) HEpG2 cells, (E) A549 cells and (F) PBMC were
exposed to different doses of rNDV-P05 [10 (◻) and 50 (∆) HU] at 24
and 48 h. The EF was evaluated. An equal volume of PBS without
virus was added to cells for control assay (○). Results are
presented as the mean ± standard error of three independent
experiments done for each cell line. *P<0.05, **P<0.01,
***P<0.001 compared to control; +P<0.05,
++P<0.01, +++P<0.001, 10 compared to 50
HU; #P<0.05, ##P<0.01,
###P<0.001, 24 compared to 48 h. All conditions
tested on A549 and PBMC were not statistically significant.
rNDV-P05, recombinant attenuated NDV strain P05; HeLa, HPV-18
cervix adenocarcinoma; HepG2, hepatoblastoma; HCC1954, breast
cancer; HL-60, promyelocytic leukemia; A549, lung carcinoma; PBMC,
peripheral blood mononuclear cells; EF, enrichment factor; PBS,
phosphate buffer saline; HU, hemagglutination units. |
rNDV-P05 induces cytokine release by
PBMC
To investigate whether rNDV-P05 can stimulate immune
cells, PBMCs from five healthy donors were incubated with 10 and 20
HU of the virus and several cytokines in supernatants were
quantified. Twenty-four h of stimulation with rNDV-P05 induced the
secretion of high amounts of TNF-α, IFN-α, TRAIL and IFN-γ from
PBMC, as shown in Fig. 3. For TNF-α,
both NDV doses significantly increased the secretion of this
cytokine by PBMC compared to control cells (P<0.001; Fig. 3A), and it was equivalent to half of
the concentration released in response to LPS. Besides, particular
values of this cytokine for each donor were very similar. As shown
in Fig. 3B, low amounts of TRAIL were
secreted by control or LPS-stimulated cells, while incubation with
10 or 20 HU of rNDV-P05 significantly increased the release of
TRAIL by PBMC, compared to control conditions (P<0.01).
Interestingly, cells from donors four and five recorded values of
TRAIL in response to virus that were similar to that of control
cells. In relation to IFN-α (Fig.
3C), control and LPS-stimulated cells from all donors did not
secrete detectable amount of the cytokine, however both NDV doses
considerably induced the release of IFN-α (P<0.05 to 10 HU,
P<0.01 to 50 HU). It is noteworthy that again the lowest
response to NDV treatment was observed in donors four and five.
Finally, both doses of virus and LPS significantly increased the
secretion of IFN-γ as compared to control cells (P<0.05 to LPS,
P<0.01 to 50 HU, P<0.001 to 10 HU; Fig. 3D). In this case, cells from donor four
released the lowest concentration of IFN-γ in response to rNDV
stimulation.
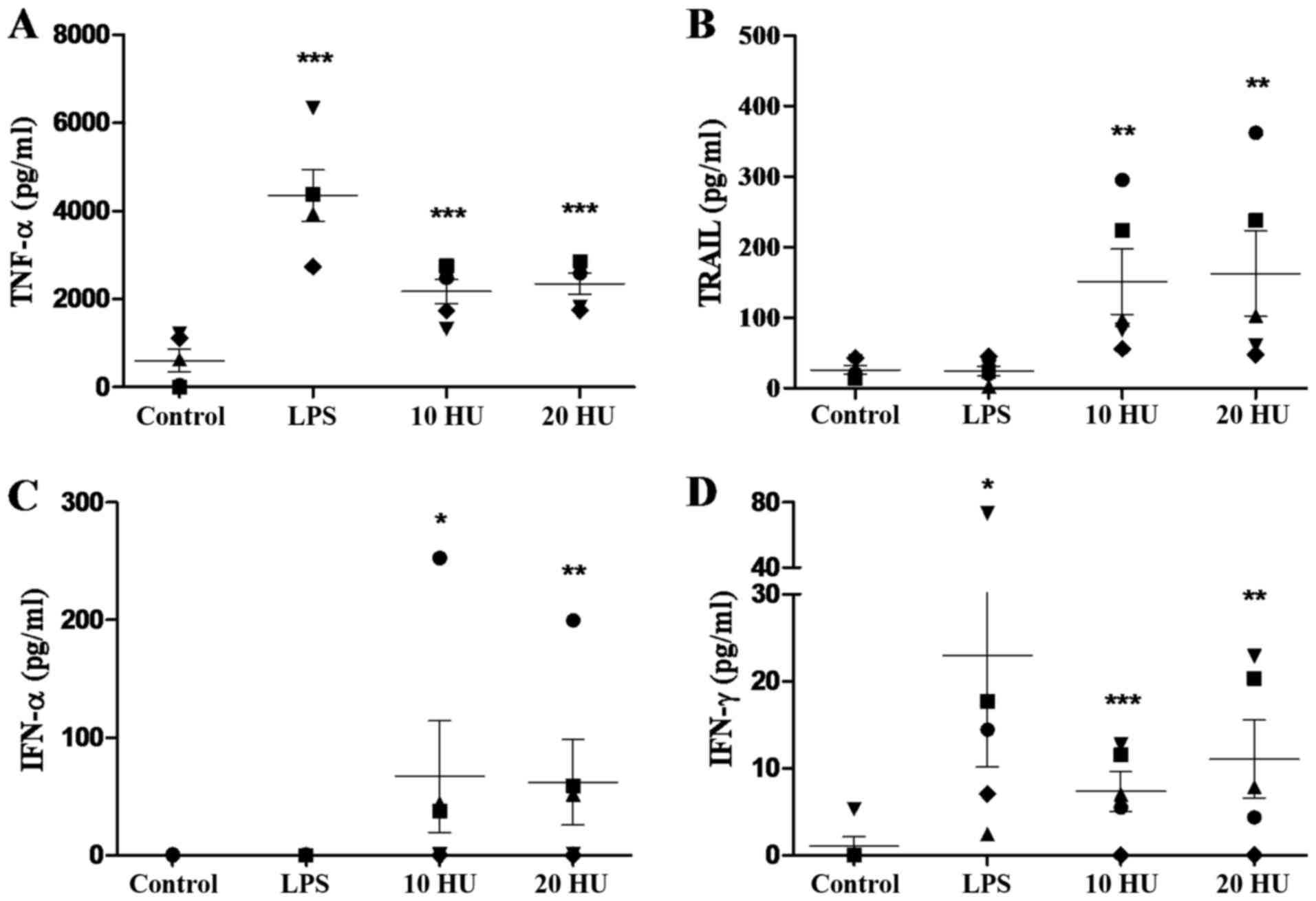 | Figure 3.Immunostimulatory activity of
rNDV-P05 over PBMC evaluated by ELISA. (A) TNF-α, (B) TRAIL, (C)
IFN-α, and (D) IFN-γ release by PBMC after 24 h of co-culture with
rNDV-P05 (10 and 20 HU/ml) or LPS (5 µg/ml). Control cells were
incubated with PBS. Results are presented as the mean ± standard
error. Cytokines were determined by duplicate in PBMC isolated from
five healthy donors: donor 1 (●); 2 (■); 3 (▲); 4 (♦); and 5 (▼).
*P<0.05, **P<0.01, ***P<0.001 compared to control.
rNDV-P05, recombinant attenuated NDV strain P05; PBMC, peripheral
blood mononuclear cells; ELISA, enzyme-linked immunosorbent assay;
TNF-α, tumor necrosis factor-α; TRAIL, tumor necrosis factor
related apoptosis inducing ligand; IFN, interferon; LPS,
lipopolysaccharide; PBS, phosphate buffer saline; HU,
hemagglutination units. |
Early cytokine response at mRNA level
induced by rNDV-P05 in PBMC
As previously described, stimulation of PBMC with
rNDV was evaluated in a late stage (24 h) through ELISA for
cytokine concentration in culture supernatants. Additionally, an
early response was determined by real time qPCR at 4 h after
stimulation (Fig. 4). PBMC treated
with 20 HU of virus increased the expression of TNF-α 3.06-fold
(P<0.01; Fig. 4A), TRAIL 8.64-fold
(P<0.01; Fig. 4B), IFN-α
10.80-fold (P<0.01; Fig. 4C), and
IFN-γ 11.41-fold (P<0.05; Fig.
4D), compared to unstimulated PBMC. At this time, LPS-treated
PBMC only increased significantly the expression of TRAIL compared
to control group (P<0.05; Fig.
4B).
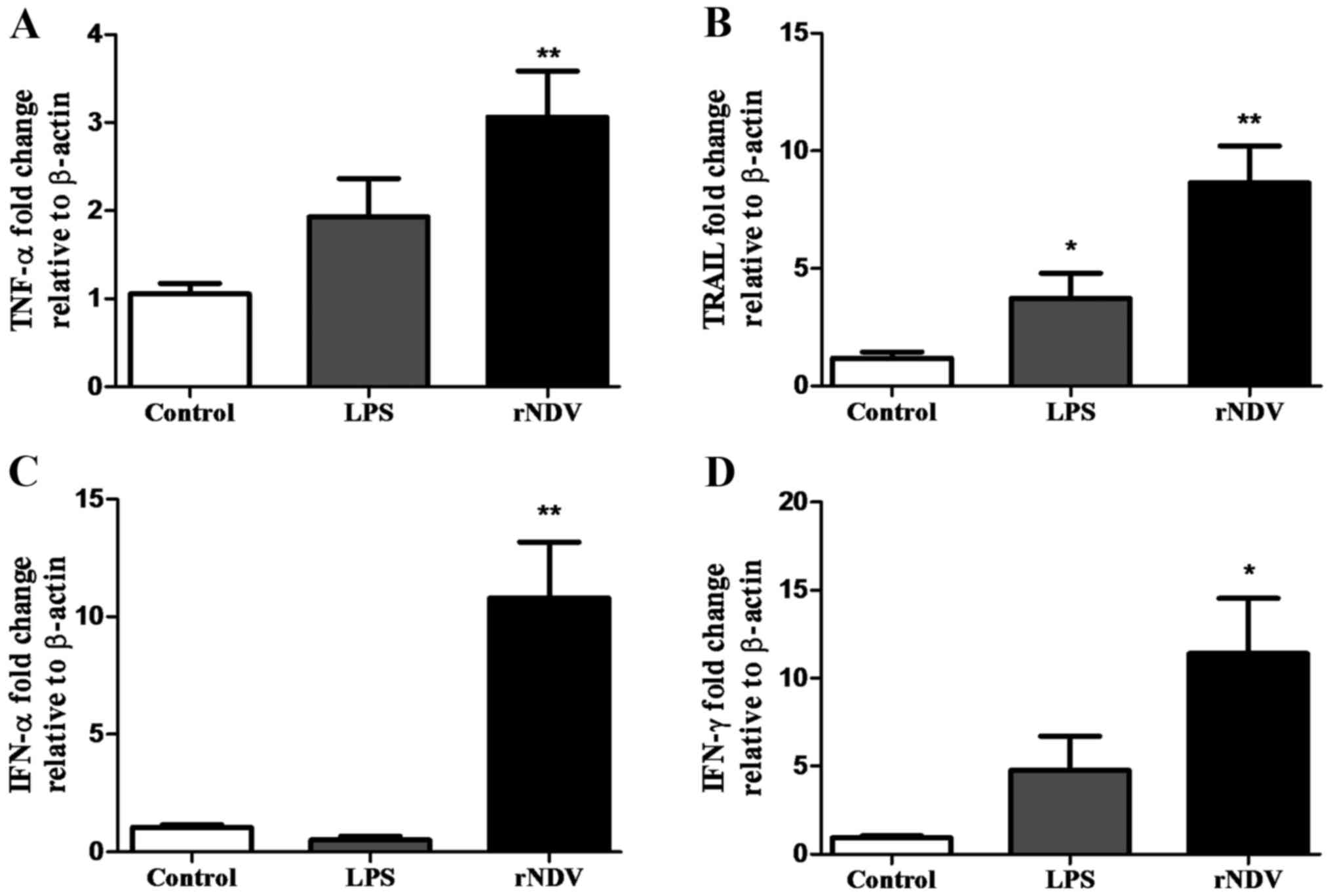 | Figure 4.Cytokine expression assessment in
PBMC stimulated with rNDV-P05 by quantitative PCR. Changes in the
expression of (A) TNF-α, (B) TRAIL, (C) IFN-α,
and (D) IFN-γ genes were evaluated at 4 h after co-culture
with rNDV-P05 20 HU/ml or LPS (5 µg/ml). Control cells were
incubated with PBS. β-actin gene was used as housekeeping
gene. Results are presented as the mean ± standard error. Total RNA
from three healthy donors was used and quantified in triplicate.
*P<0.05, **P<0.01 compared to control. PBMC, peripheral blood
mononuclear cells; rNDV-P05, recombinant attenuated NDV strain P05;
TNF-α, tumor necrosis factor-α; TRAIL, tumor necrosis factor
related apoptosis inducing ligand; IFN, interferon; LPS,
lipopolysaccharide; HU, hemagglutination units; PBS, phosphate
buffer saline. |
Discussion
Recently, increasing attention has focused on
alternative therapies with lesser side effects than current
treatments for cancer (chemo and radiotherapy). An approach for
this concern is the use of oncolytic viruses, especially NDV, which
had demonstrated the capability to kill malignant cells as a result
of activation of apoptotic pathways, both in vitro and in
vivo (2,27). However, it has been reported that
treatment response is dependent on the used NDV strain
(2). In this sense, our goal was to determine whether a
recombinant attenuated NDV from a genotype V velogenic strain
(22,24) possesses hemagglutinating, antitumor
and immunostimulatory activity.
Firstly, we demonstrated that rNDV-P05 has
hemagglutinating activity, which was partially lost under acid or
basic conditions, and totally at higher temperatures than 42°C.
These data gave us an idea for correct and optimum handling of the
virus. Our results are similar to those described to LaSota and R2B
strains (28), with the exception
that rNDV-P05 remains with the same hemagglutination titter even at
42°C while LaSota and R2B strains reduced one titer (28). That is important since
hemagglutinating activity has been demonstrated to play a crucial
role in antitumor and immunostimulatory activities (15).
Regarding to antineoplastic activity of NDV, it is
already clear that oncolytic strains are mostly velogenic and that
lentogenic strains are more immunostimulatory than
antiproliferative (2,27). Although rNDV-P05 expresses HN and F
proteins from a velogenic strain, it replicates as a lentogenic
strain due to a replacement in the cleavage site in F protein from
polybasic to monobasic amino acids (24). Keeping this in mind, our next aim
characterizing rNDV-P05 was to explore whether the virus had the
capability or not to induce apoptosis over a panel of tumor cell
lines from a variety of tissues. In this study, we have
demonstrated that rNDV-P05 was able to induce apoptosis over four
from five tested cell lines, which had previously shown
susceptibility to NDV (29–32). In solid tumors, this is important for
virotherapy, given that the cell death induced by the virus in the
tumor microenvironment can evoke not only local, but also a
systemic tumor-specific immune surveillance against primary and
metastatic malignant cells. This effect can be mediated through the
release of tumor-associated antigens, damage-associated molecular
patterns, pathogen-associated molecular patterns and inflammatory
cytokines that can help to develop a local and systemic Th1
response (33–35).
Furthermore, it is worthy of mention that due to the
fact that neutralizing antibodies (mostly anti-HN and anti-F) may
limit repeated delivery of the virus (36) and therefore decrease its efficacy,
approaches such as the use of different viral strains or engineered
viruses expressing different surface glycoproteins can be
considered (27). It has been
demonstrated that neutralizing antibodies are strain specific
(37). In this sense, the rNDV
proposed here could be considered for virotherapy, not only by the
fact that belongs to a different genotype of most used NDV strains
for virotherapy (22), but also
because it has shown to possess promising antineoplastic action
against numerous cancer cell lines and to be safe when co-cultured
with normal cells (like PBMC) from healthy donors.
Additionally to the antitumor activity of rNDV-P05,
we found that the virus has the capability to upregulate the
expression of a group of cytokines involved in tumor clearance,
such as TNF-α, TRAIL, IFN-α and IFN-γ. TNF-α is a
membrane-integrated protein and a soluble cytokine with
inflammatory and antitumor activity (38–41).
Interestingly, in this study the attenuated rNDV-P05 triggered a
significant increase of TNF-α in culture supernatants and an early
upregulation at the mRNA level. Previous studies in vitro
using the lytic NDV strain 73-T showed that human PBMC upregulate
the expression of TNF-α in supernatant after 24 h of stimulation
(18,42). Other reports have demonstrated the
capability of NDV to stimulate the production of this cytokine,
although it was in murine macrophages (19,43). In
patients with advanced solid tumors, the administration of a single
or multiple doses of the lentogenic NDV PV701 induced the rejection
of the tumors with an increased amount in serum of pro-inflammatory
cytokines, such as TNF-α (44). Thus,
TNF-α overexpression might be involved in the antineoplastic
activity of NDV-stimulated PBMC.
As a member of the apoptosis-inducing TNF family,
TRAIL has shown its ability to kill a broad range of malignant
cells in vitro and in vivo (45,46).
Particularly for NDV, it has been demonstrated the overexpression
of TRAIL in membrane of human PBMC stimulated with NDV Ulster,
especially in monocytes and T cell populations (15). In another study, NDV Ulster-activated
human monocytes enhanced their cytotoxic activity over different
cell lines, which was TRAIL-dependent and similar to that for
IFN-α-activated monocytes (47). More
recently, the NDV strain 7793 was used to active murine NK cells.
NDV 7793-activated NK cells overexpressed TRAIL in membrane and in
supernatants respect to untreated cells, and the effect was similar
to that of IFN-γ-activated NK cells (48). According to our study, human PBMC
stimulated with the rNDV increased significantly the levels of
TRAIL secreted into the culture supernatants, as well as the mRNA
expression. TRAIL expression in LPS-stimulated cells was only
significant when it was measured by qPCR; this result could be
explained by the fact that the mRNA levels might reflect the total
of TRAIL molecules (membrane-attached and soluble) and TRAIL in
culture supernatants might reflect only the secreted forms of
TRAIL. Due to the fact that we detected a prompt upregulation of
TRAIL mRNA in response to rNDV incubation, we can propose that as
an early event the increase in TRAIL expression is mostly dependent
on direct viral stimulation, as previously described (49,50).
However, it is known that TRAIL can be upregulated also by IFN-α/β
(51,52) and IFN-γ (52,53).
Interestingly, we detected high concentrations of IFN-α/γ in
response to rNDV stimulation; thus, we cannot exclude that part of
soluble TRAIL measured in PBMC cultures comes from populations of
monocytes and/or T-cells stimulated with type I IFNs, as it has
been already reported (20,47).
Kinetic expression of IFN-α by PBMC in response to
NDV has been previously evaluated (54). In accordance with that study, we found
that stimulation of human PBMC with rNDV, but not with LPS, induced
an upregulation of IFN-α gene compared to control. At
protein level, we also found an augmentation of IFN-α in the
culture supernatants from rNDV-stimulated cells compared to
control. An increment in IFN-α secretion in response to the lytic
NDV 73-T and Ulster strains has been previously reported (15,18).
However, in other study using NDV Ulster strain the production of
IFN-α was not detected neither in populations of monocytes nor
plasmacytoid dendritic cells (DCs) obtained from human PBMC; only a
recombinant version of the virus that expressed
granulocyte-macrophage colony-stimulating factor was able to
increase significantly IFN-α levels (20). In this regard, rNDV-P05 was able to
induce IFN-α secretion at the level reported to NDV Ulster
(20) and without the requirement to
express a foreign protein to trigger it. Increased levels of this
cytokine have also been detected in serum from patients treated
with NDV (44). The fast production
of IFN-α is the first-line defense mechanism after viral contact
with innate immune system, as it possesses antiproliferative
properties (55–57), remarking the importance of its
production during virotherapy.
According with early reports, the role that IFN-γ
plays in tumor rejection during NDV therapy is not clear. For
instance, stimulation of human PBMC with NDV 73-T for 24 h did not
enhance the production of this cytokine, at least not at detectable
levels (18). A different approach
using MCF-7 mammary cancer cells infected with NDV Ulster as a
tumor vaccine was not able to induce the secretion of IFN-γ in
human T-cells after five days of co-culture. Only a recombinant
version of the virus expressing interleukin (IL)-2 enhanced the
concentration of IFN-γ in supernatant, when compared to control
groups (21). Recently, employing a
genotype VIII velogenic strain (AF2240) over human PBMC, it was
demonstrated the capacity of this strain to stimulate IFN-γ
production (17). Here, we found that
PBMC (except from donor 4) respond to rNDV-P05 and LPS with an
upregulation in IFN-γ secretion. An increase at mRNA level was also
observed. Our results highlight that although rNDV-P05 is
recombinantly attenuated, it triggers a similar effect when
compared with a velogenic strain (17), without needing the expression of
foreign cytokines, such as IL-2 (21). The upregulation in IFN-γ induced by
rNDV on human PBMC could be beneficial in vivo, due to IFN-γ
is involved in activating anticancer immunity (T cells, NK cells
and DCs), in inhibiting the activity of immune-suppressive cells
(regulatory T cells and myeloid-derived suppressor cells), as well
as in the conversion of tumor-associated macrophages (58,59).
In conclusion, taking together our and previous
reports, there is a heterologous response of tumor and immune cells
after NDV stimulation and it is noteworthy that the genotype of the
strain applied for virotherapy seems to be, at least in part, the
reason for this effect. This study shows for the first time that a
genotype V NDV possesses antitumor and immunostimulatory
activities. Due to rNDV-P05 performed a promising antitumor
activity and was able to stimulate the expression of four key
antitumor cytokines, the present study opens the path for its use
in an alternate application of different viral strains during
virotherapy.
Acknowledgements
This study was supported by the National Council of
Science and Technology of Mexico (grant no. 207977). Oscar Antonio
Ortega-Rivera has a doctoral fellowship from the National Council
of Science and Technology of Mexico. The authors wish to thank Dr
Carlos Olvera Sandoval and Ms Pamela Gallegos Alcalá for excellent
technical assistance and Dr Daniel Cervantes García for his support
with PCR analysis.
References
|
1
|
World Health Organization (WHO), . 2017,
https://www.who.int/cancer/en/April
10–2017
|
|
2
|
Schirrmacher V: Fifty years of clinical
application of Newcastle disease virus: Time to celebrate!
Biomedicines. 4:E162016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Russell SJ, Peng KW and Bell JC: Oncolytic
virotherapy. Nat Biotechnol. 30:658–670. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kroemer G, Galluzzi L, Kepp O and Zitvogel
L: Immunogenic cell death in cancer therapy. Annu Rev Immunol.
31:51–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guo ZS, Liu Z and Bartlett DL: Oncolytic
immunotherapy: Dying the right way is a key to eliciting potent
antitumor immunity. Front Oncol. 4:742014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mayo MA: Virus taxonomy-Houston 2002. Arch
Virol. 147:1071–1076. 2002.PubMed/NCBI
|
|
7
|
Alexander DJ: Newcastle disease and other
avian paramyxoviruses. Rev Sci Tech. 19:443–462. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Czeglédi A, Ujvári D, Somogyi E, Wehmann
E, Werner O and Lomniczi B: Third genome size category of avian
paramyxovirus serotype 1 (Newcastle disease virus) and evolutionary
implications. Virus Res. 120:36–48. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Morrison T, McQuain C, Sergel T, McGinnes
L and Reitter J: The role of the amino terminus of F1 of the
Newcastle disease virus fusion protein in cleavage and fusion.
Virology. 193:997–1000. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Panda A, Huang Z, Elankumaran S, Rockemann
DD and Samal SK: Role of fusion protein cleavage site in the
virulence of Newcastle disease virus. Microb Pathog. 36:1–10. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ahlert T and Schirrmacher V: Isolation of
a human melanoma adapted Newcastle disease virus mutant with highly
selective replication patterns. Cancer Res. 50:5962–5968.
1990.PubMed/NCBI
|
|
12
|
Elankumaran S, Rockemann D and Samal SK:
Newcastle disease virus exerts oncolysis by both intrinsic and
extrinsic caspase-dependent pathways of cell death. J Virol.
80:7522–7534. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kumar R, Tiwari AK, Chaturvedi U, Kumar
GR, Sahoo AP, Rajmani RS, Saxena L, Saxena S, Tiwari S and Kumar S:
Velogenic Newcastle disease virus as an oncolytic virotherapeutics:
In vitro characterization. Appl Biochem Biotechnol. 167:2005–2022.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gallucci S and Matzinger P: Danger
signals: SOS to the immune system. Curr Opin Immunol. 13:114–119.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeng J, Fournier P and Schirrmacher V:
Induction of interferon-alpha and tumor necrosis factor-related
apoptosis-inducing ligand in human blood mononuclear cells by
hemagglutinin-neuraminidase but not F protein of Newcastle disease
virus. Virology. 297:19–30. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fournier P, Zeng J and Schirrmacher V: Two
ways to induce innate immune responses in human PBMCs: Paracrine
stimulation of IFN-α responses by viral protein or dsRNA. Int J
Oncol. 23:673–680. 2003.PubMed/NCBI
|
|
17
|
Lam HY, Yusoff K, Yeap SK, Subramani T,
Abd-Aziz S, Omar AR and Alitheen NB: Immunomodulatory effects of
Newcastle disease virus AF2240 strain on human peripheral blood
mononuclear cells. Int J Med Sci. 11:1240–1247. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zorn U, Dallmann I, Grosse J, Kirchner H,
Poliwoda H and Atzpodien J: Induction of cytokines and cytotoxicity
against tumor cells by Νewcastle disease virus. Cancer Biother.
9:225–235. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Schirrmacher V, Bai L, Umansky V, Yu L,
Xing Y and Qian Z: Newcastle disease virus activates macrophages
for anti-tumor activity. Int J Oncol. 16:363–373. 2000.PubMed/NCBI
|
|
20
|
Janke M, Peeters B, de Leeuw O, Moorman R,
Arnold A, Fournier P and Schirrmacher V: Recombinant Newcastle
disease virus (NDV) with inserted gene coding for GM-CSF as a new
vector for cancer immunogene therapy. Gene Ther. 14:1639–1649.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Janke M, Peeters B, Zhao H, de Leeuw O,
Moorman R, Arnold A, Ziouta Y, Fournier P and Schirrmacher V:
Activation of human T cells by a tumor vaccine infected with
recombinant Newcastle disease virus producing IL-2. Int J Oncol.
33:823–832. 2008.PubMed/NCBI
|
|
22
|
Absalón AE, Mariano-Matías A,
Vásquez-Márquez A, Morales-Garzón A, Cortés-Espinosa DV,
Ortega-García R and Lucio-Decanini E: Complete genome sequence of a
velogenic Newcastle disease virus isolated in Mexico. Virus Genes.
45:304–310. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|
|
24
|
Garzón-Morales JA, Lucio-Decanini E,
Cortes-Espinosa DV and Absalón-Constantino AE: Newcastle disease
virus and the use thereof as a vaccine US Patent 2013/0315956 A1.
Filing date: November 18, 2011. Publication date. November
28–2013
|
|
25
|
Gallagher SR: Unit 7.3. SDS-polyacrylamide
gel electrophoresis (SDS-PAGE). Curr Protoc Essent Lab Tech.
2012:1–28. 2011.
|
|
26
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative CT method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zamarin D and Palese P: Oncolytic
Newcastle disease virus for cancer therapy: Old challenges and new
directions. Future Microbiol. 7:347–367. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rani S, Gogoi P and Kumar S: Spectrum of
Newcastle disease virus stability in gradients of temperature and
pH. Biologicals. 42:351–354. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Alabsi AM, Bakar SA, Ali R, Omar AR, Bejo
MH, Ideris A and Ali AM: Effects of Newcastle disease virus strains
AF2240 and V4-UPM on cytolysis and apoptosis of leukemia cell
lines. Int J Mol Sci. 12:8645–8660. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lv Z, Zhang TY, Yin JC, Wang H, Sun T,
Chen LQ, Bai FL, Wu W, Ren GP and Li DS: Enhancement of anti-tumor
activity of Newcastle disease virus by the synergistic effect of
cytosine deaminase. Asian Pac J Cancer Prev. 14:7489–7496. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jiang K, Li Y, Zhu Q, Xu J, Wang Y, Deng
W, Liu Q, Zhang G and Meng S: Pharmacological modulation of
autophagy enhances Newcastle disease virus-mediated oncolysis in
drug-resistant lung cancer cells. BMC Cancer. 14:5512014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rajmani RS, Gupta SK, Singh PK, Gandham
RK, Sahoo AP, Chaturvedi U and Tiwari AK: HN protein of Newcastle
disease virus sensitizes HeLa cells to TNF-α-induced apoptosis by
downregulating NF-κB expression. Arch Virol. 161:2395–2405. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ni J, Galani IE, Cerwenka A, Schirrmacher
V and Fournier P: Antitumor vaccination by Newcastle disease virus
hemagglutinin-neuraminidase plasmid DNA application: Changes in
tumor microenvironment and activation of innate anti-tumor
immunity. Vaccine. 29:1185–1193. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Donnelly OG, Errington-mais F, Steele L,
Hadac E, Scott K, Peach H, Phillips RM, Bond J, Harrington K, Vile
R, et al: Measles virus causes immunogenic cell death in human
melanoma. Gene Ther. 20:7–15. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zamarin D, Holmgaard RB, Subudhi SK, Park
JS, Mansour M, Palese P, Merghoub T, Wolchok JD and Allison JP:
Localized oncolytic virotherapy overcomes systemic tumor resistance
to immune checkpoint blockade immunotherapy. Sci Transl Med.
6:226–232. 2014. View Article : Google Scholar
|
|
36
|
Seal BS, King DJ and Sellers HS: The avian
response to Newcastle disease virus. Dev Comp Immunol. 24:257–268.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Miller PJ, King DJ, Afonso CL and Suarez
DL: Antigenic differences among Newcastle disease virus strains of
different genotypes used in vaccine formulation affect viral
shedding after a virulent challenge. Vaccine. 25:7238–7246. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Balkwill FR, Lee A, Aldam G, Moodie E,
Thomas JA, Tavernier J and Fiers W: Human tumor xenografts treated
with recombinant human tumor necrosis factor alone or in
combination with interferons. Cancer Res. 46:3990–3993.
1986.PubMed/NCBI
|
|
39
|
Talmadge JE, Phillips H, Schneider M, Rowe
T, Pennington R, Bowersox O and Lenz B: Immunomodulatory properties
of recombinant murine and human tumor necrosis factor. Cancer Res.
48:544–550. 1988.PubMed/NCBI
|
|
40
|
van der Veen AH, de Wilt JH, Eggermont AM,
van Tiel ST, Seynhaeve ALB and ten Hagen TL: TNF-alpha augments
intratumoural concentrations of doxorubicin in TNF-alpha-based
isolated limb perfusion in rat sarcoma models and enhances
anti-tumour effects. Br J Cancer. 82:973–980. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Seynhaeve AL, Hoving S, Schipper D,
Vermeulen CE, De Wiel-Ambagtsheer GA, VanTiel ST, Eggermont AM and
Ten Hagen TL: Tumor necrosis factor-alpha mediates homogeneous
distribution of liposomes in murine melanoma that contributes to a
better tumor response. Cancer Res. 67:9455–9462. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lorence RM, Rood PA and Kelley KW:
Newcastle disease virus as an antineoplastic agent: Induction of
tumor necrosis factor-alpha and augmentation of its cytotoxicity. J
Natl Cancer Inst. 80:1305–1312. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ahmed I, Ahmad U, Keong YY, Manna NA and
Othman F: Induction of nitric oxide and TNF-α in Newcastle disease
virus (NDV) AF2240 infected RAW 264.7 macrophages and their
cytotoxic activity on MDA-MB-231 breast cancer cell line. J Cancer
Sci Ther. 6:478–482. 2014. View Article : Google Scholar
|
|
44
|
Pecora AL, Rizvi N, Cohen GI, Meropol NJ,
Sterman D, Marshall JL, Goldberg S, Gross P, O'Neil JD, Groene WS,
et al: Phase I trial of intravenous administration of PV701, an
oncolytic virus, in patients with advanced solid cancers. J Clin
Oncol. 20:2251–2266. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ashkenazi A, Pai RC, Fong S, Leung S,
Lawrence DA, Marsters SA, Blackie C, Chang L, McMurtrey AE, Hebert
A, et al: Safety and antitumor activity of recombinant soluble Apo2
ligand. J Clin Invest. 104:155–162. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Walczak H, Miller RE, Ariail K, Gliniak B,
Griffith TS, Kubin M, Chin W, Jones J, Woodward A, Le T, et al:
Tumoricidal activity of tumor necrosis factor-related
apoptosis-inducing ligand in vivo. Nat Med. 5:157–163. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Washburn B, Weigand MA, Grosse-Wilde A,
Janke M, Stahl H, Rieser E, Sprick MR, Schirrmacher V and Walczak
H: TNF-related apoptosis-inducing ligand mediates tumoricidal
activity of human monocytes stimulated by Newcastle disease virus.
J Immunol. 170:1814–1821. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Song DZ, Liang Y, Xiao Q, Yin J, Gong JL,
Lai ZP, Zhang ZF, Gao LX and Fan XH: TRAIL is involved in the
tumoricidal activity of mouse natural killer cells stimulated by
Newcastle disease virus in vitro. Anat Rec (Hoboken).
296:1552–1560. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Vidalain PO, Azocar O, Lamouille B, Astier
A, Rabourdin-Combe C and Servet-Delprat C: Measles virus induces
functional TRAIL production by human dendritic cells. J Virol.
74:556–559. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chaperot L, Blum A, Manches O, Lui G,
Angel J, Molens JP and Plumas J: Virus or TLR agonists induce
TRAIL-mediated cytotoxic activity of plasmacytoid dendritic cells.
J Immunol. 176:248–255. 2005. View Article : Google Scholar
|
|
51
|
Ehrlich S, Infante-Duarte C, Seeger B and
Zipp F: Regulation of soluble and surface-bound TRAIL in human T
cells, B cells and monocytes. Cytokine. 24:244–253. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Griffith TS, Wiley SR, Kubin MZ, Sedger
LM, Maliszewski CR and Fanger NA: Monocyte-mediated tumoricidal
activity via the tumor necrosis factor-related cytokine, TRAIL. J
Exp Med. 189:1343–1354. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fanger NA, Maliszewski CR, Schooley K and
Griffith TS: Human dendritic cells mediate cellular apoptosis via
tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). J
Exp Med. 190:1155–1164. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Löseke S, Grage-Griebenow E, Wagner A,
Gehlhar K and Bufe A: Differential expression of IFN-alpha subtypes
in human PBMC: Evaluation of novel real-time PCR assays. J Immunol
Methods. 276:207–222. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Goodbourn S, Didcock L and Randall RE:
Interferons: Cell signalling, immune modulation, antiviral response
and virus countermeasure. J Gen Virol. 81:2341–2364. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Onoguchi K, Yoneyama M, Takemura A, Akira
S, Taniguchi T, Namiki H and Fujita T: Viral infections activate
types I and III interferon genes through a common mechanism. J Biol
Chem. 282:7576–7581. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ivashkiv LB and Donlin LT: Regulation of
type I interferon responses. Nat Rev Immunol. 14:36–49. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Schroder K, Hertzog PJ, Ravasi T and Hume
DA: Interferon-gamma: An overview of signals, mechanisms and
functions. J Leukoc Biol. 75:163–189. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Parker BS, Rautela J and Hertzog PJ:
Antitumour actions of interferons: Implications for cancer therapy.
Nat Rev Cancer. 16:131–144. 2016. View Article : Google Scholar : PubMed/NCBI
|














