Introduction
Multiple myeloma is characterized by an increased
level of light chain monoclonal immunoglobulin, as well as a
simultaneous decrease in the level of normal immunoglobulin
(1). In multiple myeloma, osteoclasts
are activated resulting in an imbalance of the osteoclast and
osteoblast equilibrium. Osteolytic lesion formation and
osteoporosis is also observed at higher rate in multiple myeloma
(1). Currently, there are a large
number of drugs, as well as stem cell transplantation, in use for
the prevention of multiple myeloma formation (2); however, development of multidrug
resistance and disease relapse has been reported in patients. Thus,
the development of an efficient therapeutic strategy for multiple
myeloma is highly desired.
Natural products, along with their derivatives,
possess the source of a large number of drugs. Studies have
determined that natural products can prevent, decrease, and
possibly defeat several pathologies, including cancer, diabetes,
cardiovascular and neurological disorders (3–6).
Artemisinin is isolated from a herbaceous plant, Artemisia
annua, which is located in China and has been used as an
anti-malarial drug for a number of years (7,8). The
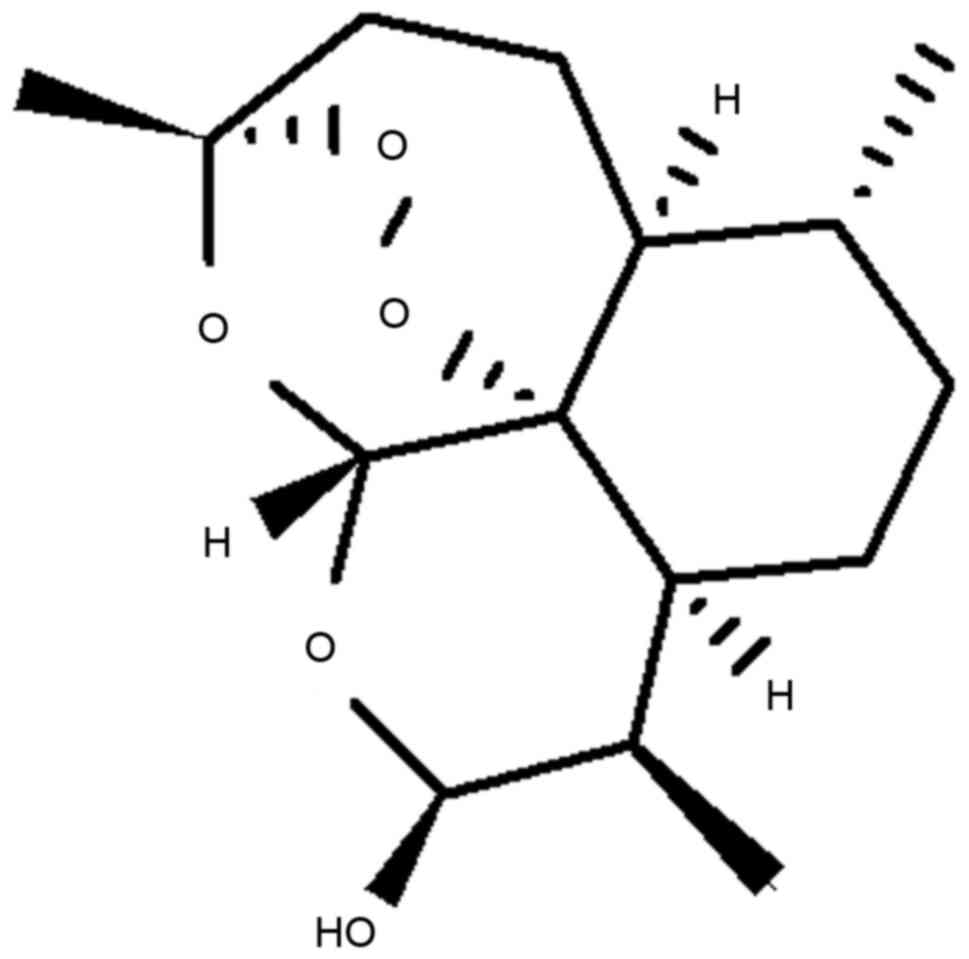
analogs of artemisinin, including dihydroartemisinin (DHA; Fig. 1), artesunate and artemether, also
possess anti-malarial potential and are, therefore, used for the
treatment of malaria (7). Screening
of artemisinin and its derivatives revealed promising potential as
inhibitors of malignant tumor proliferation in vitro
(9–12). They inhibit the proliferation of
breast (12) and ovarian cancer cells
(7) without affecting non-malignant
cells (12,13). Taking into account the anticancer
potential of DHA along with its limited side effects on normal
cells, the present study aimed to investigate the effect of DHA on
multiple myeloma. The results obtained revealed that the DHA
treatment caused the induction of apoptosis in U266 cells through
c-Jun N-terminal kinase (JNK) signaling pathway and c-Jun
activation.
Materials and methods
Cell culture
The cell line U266 was purchased from the Shanghai
Institute of Biochemistry and Cellular Biology Chinese Academy of
Sciences (Shanghai, China). The culture medium used for the cell
line was Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The medium contained 10% fetal
bovine serum (HyClone, Logan, UT, USA), penicillin (100 U/ml) and
streptomycin (100 U/ml). Cell culture was carried out in an
incubator with 5% CO2 and 95% air at 37°C.
MTT assay
Effects of DHA on cell proliferation was determined
using an MTT assay. The concentration of the cells was adjusted to
4×106 cells/ml, and 190 µl cell suspension was added
into each well of the 96-well plate. Plates were incubated for a
period of 24 h using a 5% CO2 incubator at 37°C. Various
concentrations of DHA (1, 3, 10, 30 and 100 µmol/l) were added to
the plates and incubation was continued for 12, 24 and 48 h. The
addition of 20 µl MTT solution (5 mg/ml) to each well of the plate
was conducted at 12, 24 and 48 h. Further incubation of the plates
was carried out for 4 h, followed by decantation of supernatant and
the addition of 150 µl dimethyl sulfoxide to every well.
Measurement of the absorbance for each well was performed in
triplicate at a wavelength of 490 nm to determine the cell
viability.
Flow cytometry for analysis of the
cell cycle
The effect of various concentrations (1, 3, 10, 30
and 100 µmol/l) of DHA on cell proportion in various cell cycle
phases were examined using flow cytometry. Exponentially
proliferating cells at a concentration of 3.5×105
cells/ml after 48 h of DHA treatment were collected and
subsequently subjected to PBS washing. Cells were then subjected to
fixing with 70% ethanol at −20°C for 24 h followed by PBS washing
and then stained with propidium iodide in the dark for 5 min at
room temperature. Nuclear DNA was analyzed using a CycleTEST™ Plus
kit (BD Biosciences, San Jose, CA, USA) as per the manufacturer's
protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Following treatment with DHA for a period of 48 h,
cells were treated with TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) to isolate the total RNA according
to the manufacturer's protocol. Samples of 2 µl total RNA were used
for the synthesis of cDNA, and amplification was performed using
the ThermoScript RT-PCR system (Thermo Fisher Scientific, Inc.).
Analysis was performed using a 2% agarose gel and a confirmation
made by nucleotide sequencing. Primer design for RT-PCR was
performed using GenBank sequences and Primer Express®
software for Real-Time PCR (version 3; Applied Biosystems; Thermo
Fisher Scientific, Inc.). The following primers were used for
RT-PCR: GAPDH forward, 5′-TGAACGGGAAGCTCACTGG-3′ and reverse,
5′-TCCACCACCCTGTTGCTGGA-3′; caspase-3 forward,
5′-TTTTTCAGAGGGGATCGTTG-3′ and reverse, 5′-CGGCCTCCACTGGTATTTTA-3′;
and c-Jun forward, 5′-CCCCAAGATCCTGAAACAGA-3′ and reverse,
5′-CCGTTGCTGGACTGGATTAT-3′. Primers were supplied by Hokkaido
System Science Co., Ltd. (Hokkaido, Japan).
Statistical analysis
Data are expressed as the mean ± standard deviation
(SD) and were analyzed using SPSS software (version 14; SPSS, Inc.,
Chicago, IL, USA). The one-way analysis of variance and Dunnett's
T3 post hoc analysis were used for statistical analysis. P<0.05
was considered to indicate a statistically significant
difference.
Results
DHA decreases in viability of U266
cells
The effect of different concentrations of DHA (1, 3,
10, 30 and 100 µmol/l) for various time durations (12, 24 and 48 h)
on U266 cell proliferation was analyzed using an MTT assay.
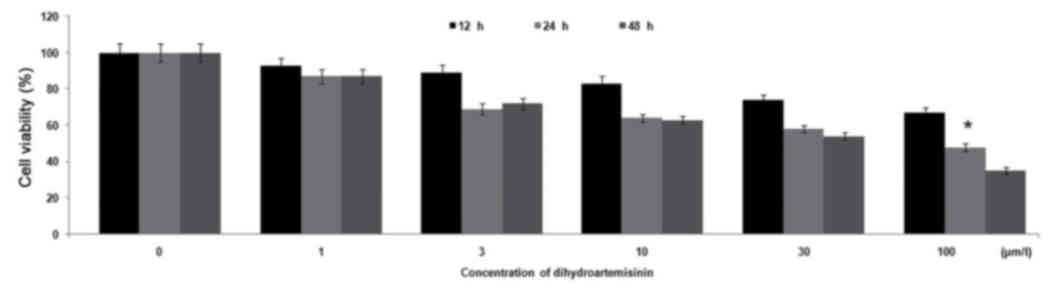
Treatment of U266 cells with DHA caused a significant (P<0.05)
decrease in cell viability compared with control cells. Increasing
the concentration of DHA from 1 to 100 µmol/l reduced cell
viability from 87 to 35%, compared with 100% in the control
cultures (Fig. 2). Treatment of U266
cells with 100 µmol/l DHA for 12, 24 and 48 h caused a reduction in
cell proliferation to 67, 48 and 35%, respectively (Fig. 2).
DHA causes cell cycle arrest in the
sub-G0/G1 phase
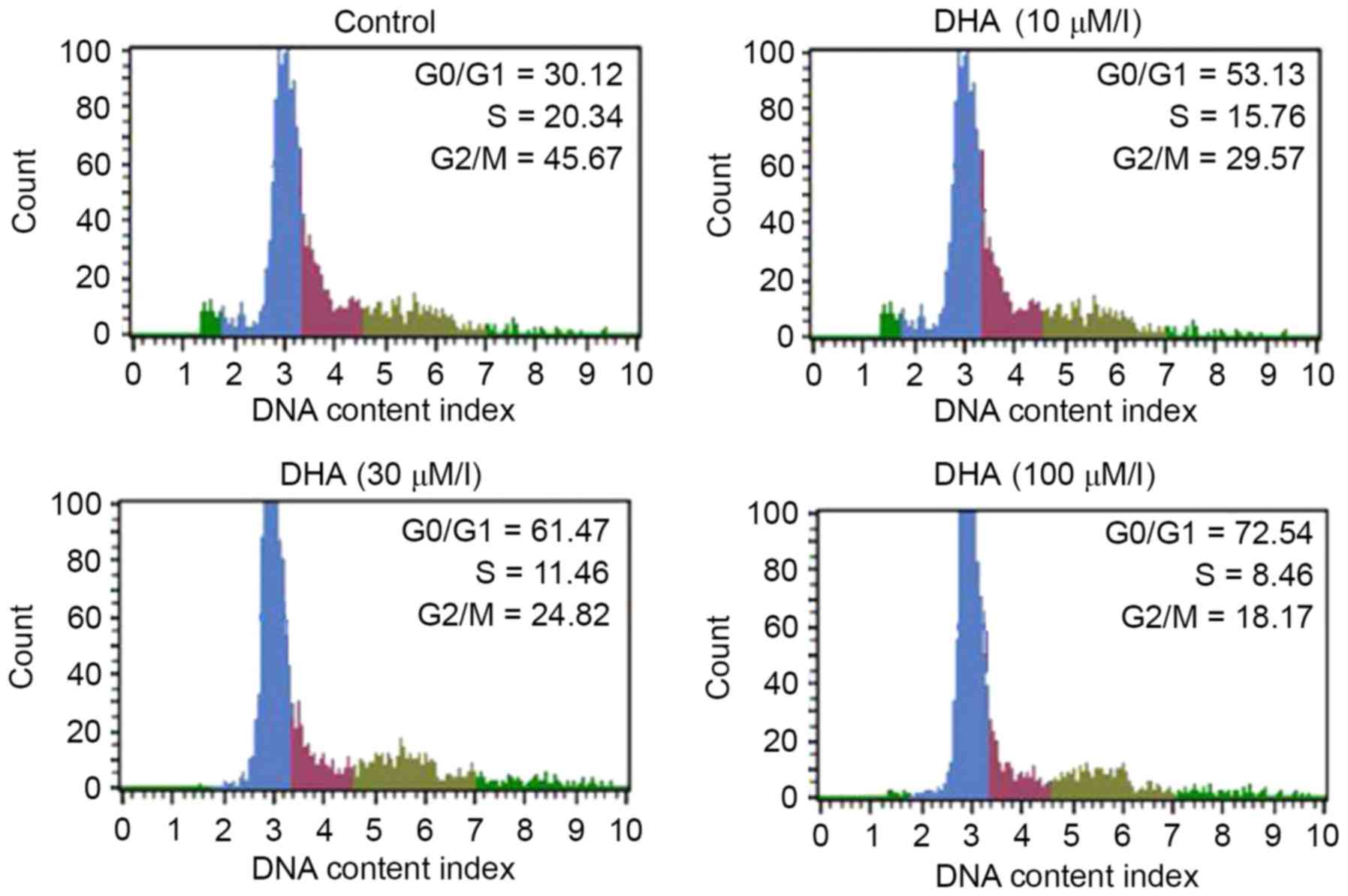
U266 cells were treated with 1, 3, 10, 30 and 100
µmol/l DHA for 48 h and then examined by flow cytometry. A
significant (P<0.05) enhancement was observed in the U266 cell
population in sub-G0/G1 phase with an
increase in DHA concentration from 1 to 100 µmol/l. Treatment with
1, 3, 10, 30 and 100 µmol/l DHA increased the
sub-G0/G1 phase cell population to 33.13,
38.25, 54.91, 74.47 and 88.54%, respectively (Fig. 3).
DHA causes an increase in c-Jun
expression in U266 cells
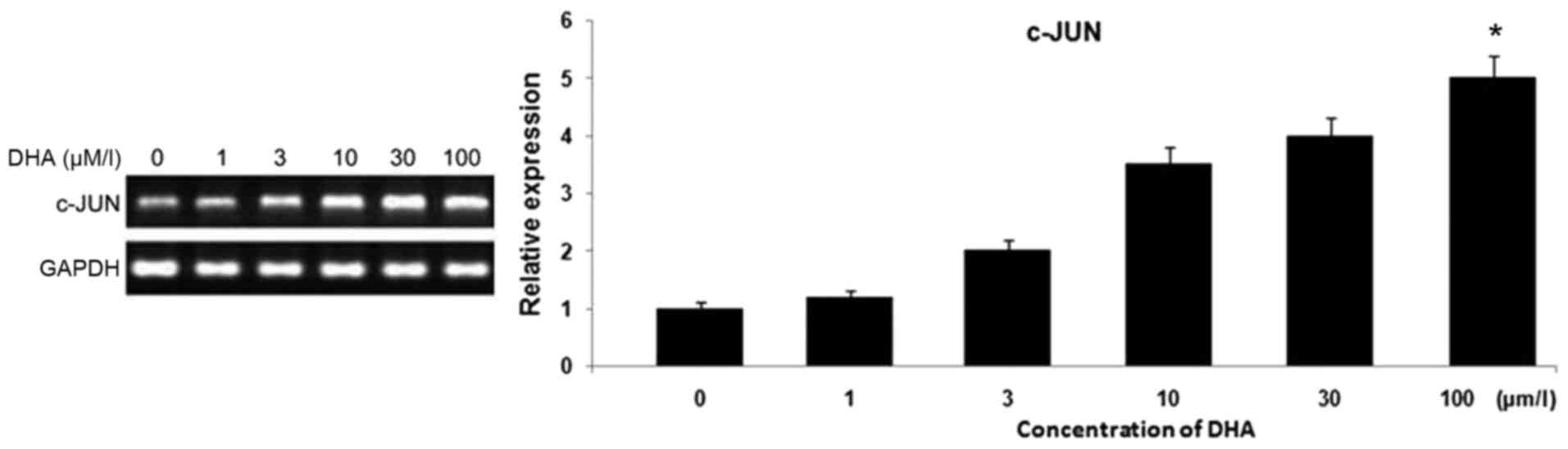
The expression of c-Jun in DHA treated or untreated
U266 cells was examined using RT-PCR analysis. Treatment of the
cells for 48 h with DHA caused a significant increase in c-Jun
expression (Fig. 5). DHA treatment at
1, 3, 10, 30 and 100 µmol/l caused enhancements in the level of
c-Jun to 0.174±0.001, 0.254±0.002, 0.387±0.001, 0.502±0.003 and
0.679±0.005 respectively, compared with 0.982±0.001 in the control
cells.
DHA causes activation of the JNK
signaling pathway in multiple myeloma cells
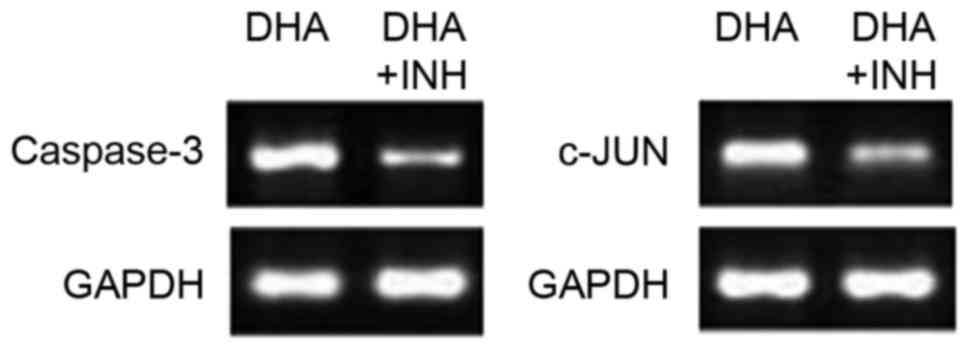
In order to confirm whether the DHA-mediated
increase in caspase-3 and c-Jun expression in U266 cells is
associated with JNK signaling, cells were treated with SP600125
(JNK signaling pathway inhibitor; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Treatment with SP600125 (2 mg/ml) following
incubation with DHA caused a significant decrease in caspase-3 and
c-Jun expression in U266 cells compared with cultures incubated
with DHA alone (Fig. 6). These
findings confirmed that DHA treatment increases the level of
caspase-3 and c-Jun in the cells through activation of the JNK
signaling pathway.
Discussion
DHA and its various derivatives have the potential
of being malignant tumor proliferation inhibitors in vitro
(3–6).
They inhibit the proliferation of breast (6) and ovarian cancer cells (7) without affecting non-malignant cells
(6,7).
Apoptosis serves an important function in the maintenance of
homeostasis in the body by removing unwanted cells (6). Any disturbance in the process of
apoptosis results in the onset of various diseases, including
cancer and autoimmune disease (7). In
the bone marrow, increased levels of malignant plasma cells result
in the development of multiple myeloma, which prevents cancer cell
apoptosis and causes an alteration in the rate of cell
proliferation. The present study was performed to determine the
influence of DHA on a multiple myeloma human cell line. The results
revealed that DHA caused a significant reduction in the viability
of multiple myeloma cells compared with the control cells.
Increasing the concentration of DHA from 1 to 100 µmol/l reduced
cell viability from 87 to 35%, compared with 100% in the control
cultures. Flow cytometric analysis revealed a significant increase
in the population of U266 cells in the
sub-G0/G1 phase when the concentration of DHA
was increased from 1 to 100 µmol/l. Treatment with 1, 3, 10, 30 and
100 µmol/l DHA increased the sub-G0/G1 phase
cell population to 3.13, 8.25, 24.91, 31.47 and 38.54%,
respectively.
Transduction of signals and the expression of
proteins are mediated by the mitogen-activated protein kinase
(MAPK) (14). Various members of the
MAPK family, including the extracellular signal regulated kinase,
JNK/stress-activated protein kinase (SAPK) and p38, are in dynamic
equilibrium with one another and serve a vital function in
maintaining cell survival and apoptosis (15,16). JNK
is a serine/threonine protein kinase and, due to its interaction
with c-Jun as well as its ability to phosphorylate, it is called
the c-Jun N-terminal kinase (15).
The center of transcription factor-activated protein-1 is c-Jun,
which on combining with Fos causes activation of transforming
growth factor [activating transcription factor (ATF)-2, liver
regeneration factor-1/ATF-3 and Jun dimerization protein-1]. The
activation of c-Jun and ATF-2 is followed by the activation of a
transcription factor, which in turn complexes with Fas via
caspase-8 activation (17). Studies
have indicated that JNK causes cell apoptosis through B-cell
lymphoma (Bcl)-2 and Bcl-extra-large activation (18). The mitochondrial pathway of cell
apoptosis involves cytochrome c release and caspase-9
activation (19). It is reported that
in multiple myeloma cells, anti-Fas monoclonal antibodies induce
apoptosis through the expression of JNK/SAPK and transcription
factor c-Jun (20,21). Activation of c-Jun leads to the
apoptosis signal transduction pathway resulting in cell apoptosis
(22). Results from the present study
revealed that DHA treatment resulted in the elevation of the
expression levels of c-Jun (JNK pathway member) and caspase-3 in
multiple myeloma cells. The addition of SP600125, an inhibitor of
JNK, to the cell culture medium resulted in the reduction of c-Jun
and caspase-3 expression. These findings suggest that DHA induces
apoptosis in multiple myeloma cells by activating the JNK signaling
pathway through the activation of c-Jun.
Thus, the present study demonstrated that DHA caused
an inhibition of proliferation for multiple myeloma cells through
JNK signaling pathway activation. Therefore, DHA can be used for
the treatment of multiple myeloma.
Glossary
Abbreviations
Abbreviations:
|
DHA
|
dihydroartemisinin
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
|
JNK
|
c-Jun N-terminal kinase
|
References
|
1
|
Wei ZL and Wang XH: The development of
NF-κB in the multiple myeloma. Med Res. 36:98–101. 2007.
|
|
2
|
Spisek R, Charalambous A, Mazumder A,
Vesole DH, Jagannath S and Dhodapkar MV: Bortezomib enhances
dendritic cell (DC)-mediated induction of immunity to human myeloma
via exposure of cell surface heat shock protein 90 on dying tumor
cells: Therapeutic implications. Blood. 109:4839–4845. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Carocho M and Ferreira IC: A review on
antioxidants, prooxidants and related controversy: Natural and
synthetic compounds, screening and analysis methodologies and
future perspectives. Food Chem Toxicol. 51:15–25. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mecocci P and Polidori MC: Antioxidant
clinical trials in mild cognitive impairment and Alzheimer's
disease. Biochim Biophys Acta. 1822:631–638. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hawkes CA, Ng V and McLaurin JA: Small
molecule inhibitors of Aβ-aggregation and neurotoxicity. Drug Dev
Res. 70:111–124. 2009. View Article : Google Scholar
|
|
6
|
Joynera PM and Cichewicz RH: Bringing
natural products into the fold-exploring the therapeutic lead
potential of secondary metabolites for the treatment of
protein-misfolding related neurodegenerative diseases. Nat Prod
Rep. 28:26–47. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Meshnick SR: Artemisinin: Mechanisms of
action, resistance and toxicity. Int J Parasitol. 32:1655–1660.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
O'Neill PM: Medicinal chemistry: A worthy
adversary for malaria. Nature. 430:838–839. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Efferth T, Dunstan H, Sauerbrey A, Miyachi
H and Chitambar CR: The anti-malarial artesunate is also active
against cancer. Int J Oncol. 18:767–773. 2001.PubMed/NCBI
|
|
10
|
Huang XJ, Ma ZQ, Zhang WP, Lu YB and Wei
EQ: Dihydroartemisinin exerts cytotoxic effects and inhibits
hypoxia inducible factor-1alpha activation in C6 glioma cells. J
Pharm Pharmacol. 59:849–856. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nam W, Tak J, Ryu JK, Jung M, Yook JI, Kim
HJ and Cha IH: Effects of artemisinin and its derivatives on growth
inhibition and apoptosis of oral cancer cells. Head Neck.
29:335–340. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Singh NP and Lai H: Selective toxicity of
dihydroartemisinin and holotransferrin toward human breast cancer
cells. Life Sci. 70:49–56. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen T, Li M, Zhang R and Wang H:
Dihydroartemisinin induces apoptosis and sensitizes human ovarian
cancer cells to carboplatin therapy. J Cell Mol Med. 13:1358–1370.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liang XM and Yang KD: Caspase, JNK/SAPK,
P38 MAPK and apoptosis. Foreign Med Sci (Section Hygiene). 35:5–10.
2008.(In Chinese).
|
|
15
|
Du L, Wang FY, Zhang L and Liu T: Advance
in the research of JNK dependent apoptosis. China Trop Med.
18:841–844. 2008.(In Chinese).
|
|
16
|
Xiao Y, Yang FQ, Li SP, Gao JL, Hu G, Lao
SC, Conceição EL, Fung KP, Wangl YT and Lee SM: Furanodiene induces
G2/M cell cycle arrest and apoptosis through MAPK signaling and
mitochondria-caspase pathway in human hepatocellular carcinoma
cells. Cancer Biol Ther. 6:1044–1050. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Papa S, Zazzeroni F, Pham CG, Bubici C and
Franzoso G: Linking JNK signaling to NF-kappaB: A key to survival.
J Cell Sci. 117:5197–5208. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Murakami Y, Aizu-Yokota E, Sonoda Y, Ohta
S and Kasahara T: Suppression of endoplasmic reticulum stress
induced caspase activation and cell death by the over expression of
Bcl-xl or Bcl-2. J Biochem. 141:401–410. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peng J, Mao XO, Stevenson FF, Hsu M and
Andersen JK: The herbicide paraquat induces dopaminergic nigral
apoptosis through sustained activation of the JNK pathway. J Biol
Chem. 279:32626–32632. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Largo C, Alvarez S, Saez B, Blesa D,
Martin-Subero JI, González-García I, Brieva JA, Dopazo J, Siebert
R, Calasanz MJ and Cigudosa JC: Identification of overexpressed
genes in frequently gained/amplified chromosome regions in multiple
myeloma. Hematologica. 91:184–191. 2004.
|
|
21
|
Lin HH, Chen JH, Kuo WH and Wang CJ:
Chemopreventive properties of Hibiscus sabdariffa L. on human
gastric carcinoma cells through apoptosis induction and JNK/p38
MAPK signaling activation. Chem Biol Interac. 165:59–75. 2007.
View Article : Google Scholar
|
|
22
|
Junttila MR, Li SP and Westermarck J:
Phosphatase-mediated crosstalk between MAPK signaling pathways in
the regulation of cell survival. FASEB J. 22:954–965. 2008.
View Article : Google Scholar : PubMed/NCBI
|