Introduction
Colorectal cancer (CRC) is the third most commonly
diagnosed cancer in the United States (1). Over the past two decades, the lifetime
risk of developing CRC has decreased in the USA, Europe and Japan
(2). Screenings by fecal occult blood
test or colonoscopy have been demonstrated to reduce the incidence
of CRC and mortality (3). Early
detection and treatment is known as the most effective measure to
improve cancer mortality (4,5). However, despite recent advances in CRC
treatment, once diagnosed, the overall prognosis remains poor
(6,7).
Treatment with several novel combination chemotherapies, including
folinic acid (LV)/5-fluorouracil (5-FU)/oxaliplatin and
LV/5-FU/irinotecan, exhibited high response rates and prolonged
survival time in patients with CRC (8–11).
However, patients with a positive response to chemotherapy remain
as high as 50% of all patients with CRC (7). Additional molecular targets and
biomarkers are required for improvements in chemotherapeutic effect
and patient survival.
Homeobox A (HOXA) cluster genes, located on
chromosome 7p15, serve a fundamental role in embryonic development
(12). HOXA genes are required for
the development of normal organs, including the central nervous
system, axial skeleton, limbs, gut, hematopoietic and urogenital
tract, and internal and external genitalia (13). Therefore, dysregulation of these genes
may potentially lead to the development of malignant tumors.
Numerous studies have reported that HOXA genes are altered in
various types of cancer, including leukemia and skin, prostate,
breast, ovarian, gastric and esophageal cancer, and those
alterations are associated with patient survival or
clinicopathological factors (14–19). In
addition, several HOXA genes are also altered in CRC, and those
alterations appear to be associated with poorer patient prognosis
(20,21).
In the present study, HOXA gene expression was
investigated using reverse transcription-quantitative polymerase
chain reaction (RT-qPCR) or immunohistochemical (IHC) staining in
surgically resected CRC tissues, and the biological significance of
HOXA genes was examined by in vitro experiments to clarify
the function of HOXA genes in CRC.
Materials and methods
Clinical patient samples
A total of 231 patients with CRC, who had undergone
surgical resection at Fukushima Medical University Hospital
(Fukushima, Japan) between January 1991 and December 2007 were
involved in the present study. Specimens from all 231 cases were
used for IHC staining. The mRNA was extracted from tumor and
adjacent non-tumor tissues in 40 of 231 cases for PCR analyses.
Information regarding age, sex, tumor-node-metastasis (TNM) stage
(22,23) and pathological diagnosis, including
lymphatic and venous invasion, were retrospectively collected. The
carcinomas at the time of primary tumor resection were staged
according to the Union for International Cancer Control
classification (22,23). The present study was approved by the
Ethics Committee of the Fukushima Medical University (Fukushima,
Japan; approval no. 1615). Written informed consent was obtained
from all patients.
Cell line culture
The colon cancer cell lines (HCT116, LoVo, RKO,
LS174T, Colo205, Colo201, SW620, LS180, SW837, SW480, HCT15, and
SW48) used in the present study were originally obtained from the
American Type Culture Collection (Manassas, VA, USA). RKO, LS174T
and LS180 cells were cultured in Dulbecco's modified Eagle's medium
supplemented with 10% fetal bovine serum (both from Thermo Fisher
Scientific, Inc., Waltham, MA, USA). SW480, LoVo, HCT15, SW48,
SW620 and HCT116 cells were cultured in RPMI-1640 (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) supplemented with 10% fetal bovine
serum. The monolayer cells were maintained in a 37°C incubator with
5% CO2, observed regularly under a light microscope
(×40) and subcultured when 80–90% confluence was reached.
RT-qPCR
Total RNA was extracted from tumor and non-tumor
tissues from 20 patients and cells using TRIzol reagent (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol
as previously described (24–26). Complementary DNA (cDNA) was
synthesized from 5 µg of the total RNA with a random hexamer using
the SuperScript III First-Strand Synthesis System (Thermo Fisher
Scientific, Inc.) with the GeneAmp PCR system 9700 (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
thermocycling conditions maintained were as follows: Denaturation,
65°C for 5 min; annealing, 25°C for 10 min; elongation, 50°C for 50
min; and termination, 85°C for 5 min and 37°C for 20 min.
Subsequently, the cDNA from CRC tissues were used for the
measurement of expression of 11 HOXA family genes (HOXA1, HOXA2,
HOXA3, HOXA4, HOXA5, HOXA6, HOXA7, HOXA9, HOXA10, HOXA11 and
HOXA13). β-actin was used as an internal control. Primer sequences
are shown in Table I. The PCR product
was loaded onto a 2% agarose gel with 0.5 µg/ml ethidium bromide.
Following electorophoresis, the product were visualized using an
UV-transilluminator (E-Box-1000/20M; Cosmo Bio Co. Ltd., Tokyo,
Japan).
 | Table I.Primer sequences for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I.
Primer sequences for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) | Size, bp |
|---|
| HOXA1 |
CCCATGGAGGAAGTGAGAAA |
GGACCATGGGAGATGAGAGA | 489 |
| HOXA2 |
AGTCTCGCCTTTAACCTAGCA |
GGCCTCATACTGCTCTCAGG | 571 |
| HOXA3 |
AATGCCAGCAACAACCCTAC |
TGACCAGCGAATGCATAGAG | 535 |
| HOXA4 |
CCCTGGATGAAGAAGATCCA |
GAGGATCGCATCTTGGTGTT | 265 |
| HOXA5 |
GTGAAGAAGCCCTGTTCTCG |
AACGAGATTGAAGGGGGACT | 244 |
| HOXA6 |
CGCGCAAATGAGTTCCTATT |
GACCGAGTTGGACTGTTGGT | 199 |
| HOXA7 |
GGCTTGCCTGCTACTAGTG |
GAAGCTGGAAGCATCTCCAC | 326 |
| HOXA9 |
CCACGCTTGACACTCACACT |
TCGTCTTTTGCTCGGTCTTT | 382 |
| HOXA10 |
GGGGGAAAAAGCCATATCAT |
GGGAGAATTGTGGTGTGCTT | 671 |
| HOXA11 |
GCTTGGAAGCTTCTGGTGAC |
AATTGAGGACAGGCCAACAC | 557 |
| HOXA13 |
CTGGAACGGCCAAATGTACT |
AGAGATTCGTCGTGGCTGAT | 386 |
| β-actin |
GCTCGTCGTCGACAACGGCTC |
CAAACATGATCTGGGTCATCTTCTC | 353 |
PCR amplification using 4 µl of cDNA with the
TaqMan® RT-qPCR kit (Thermo Fisher Scientific, Inc.) for
HOXA family genes was performed with the 7500 Real-time PCR system
(Thermo Fisher Scientific, Inc.) at 94°C for initial denaturation
for 5 min, followed by 25–35 cycles at 94°C for 1 min, 55–60°C for
45 sec and 72°C for 45 sec. Probes for TaqMan gene expression
assays [HOXA9 (assay ID. Hs00266821_m1) and β-actin (assay ID.
Hs99999903_m1)] were purchased from Thermo Fisher Scientific, Inc.,
and β-actin was used as an internal control. The primer sequences
are listed in Table I. Relative HOXA9
gene expression was calculated using the 2−ΔΔCq method
(27), according to the supplier's
protocol (Thermo Fisher Scientific, Inc.).
IHC staining and evaluation
IHC staining was performed on paraffin-embedded
histological sections (thickness, 4 µm) that were fixed in 10%
buffered formalin, using a polymer peroxidase method
(Envision+/HRP; Dako; Agilent Technologies, Inc., Santa Clara, CA,
USA) (24,25). Briefly, following deparaffinization
with xylene and rehydration using a descending alcohol series, the
tissue sections were treated with 0.3% hydrogen peroxide in
methanol for 30 min at room temperature to block endogenous
peroxidase activity. Following rinsing in PBS, the sections were
incubated with anti-HOXA9 antibody (1:1,000; cat. no. NBP2-32356;
Novus Biologicals, LLC, Littleton, CO, USA) at 4°C overnight. An
additional wash in PBS was followed by treatment with ready-to-use
peroxidase-labeled polymer conjugated to goat anti-rabbit
immunoglobulins (cat. no. SM801; ENvision + kit; Dako; Agilent
Technologies, Inc.) as the secondary antibody for 30 min at room
temperature. The staining was visualized with diaminobenzidine,
followed by counterstaining with hematoxylin.
Expression of these proteins was evaluated using
optical microscopy (BX43; Olympus Corporation, Tokyo, Japan) as
positive when the nucleus of the cancerous tissue was stained. The
staining of each specimen was evaluated at ×40 or ×400
magnification by two investigators, Y.W. and K.S. (Fukushima
Medical University School of Medicine, Fukushima, Japan), who were
blinded to the sample name and the clinical outcomes. The rate of
positive-stained cancer cells was evaluated in three randomly
selected areas (size, 200×200 µm) from the tumor specimens. When
the average positive tumor rate was >10%, the tumor was defined
as being positively stained.
Knockdown experiment and cell
counting
The knockdown experiment was performed using a small
interfering RNA (siRNA) method using HOXA9 Stealth RNAi siRNA
(HOXA9-siRNA; cat. no. HSS142497; Thermo Fisher Scientific, Inc.)
and Stealth RNAi siRNA negative control (NC-siRNA; cat. no.
12935-300; Thermo Fisher Scientific, Inc.) using Lipofectamine
RNAiMAX (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. At 1 day prior to transfection, SW48 was
seeded at 2×106 cells/well into the wells of a 10 cm
plate. Transfection with a final concentration of 20 nM siRNA oligo
was performed when the cell confluence reached 30–50%. The cells
were incubated at 37°C 48 h prior to harvest. The number of cells
was subsequently counted after 24 and 48 h siRNA transfection using
a Countess II FL Automated Cell Counter (Thermo Fisher Scientific,
Inc.). All experiments were repeated three times.
Public database
HOXA9 gene aberrations were investigated using The
Cancer Genome Atlas (TCGA) data portal (cBioPortal: http://www.cbioportal.org/).
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analyses were performed using GraphPad Prism 6 software
(GraphPad Software, Inc., La Jolla, CA, USA). Wilcoxon signed-rank
tests and unpaired Student's t-tests were performed to analyze the
differences among groups. In addition, survival curves were
generated using the Kaplan-Meier method and compared using the log
rank test. A total of 226 patients were used in the Kaplan-Meier
survival analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
HOXA9 expression is upregulated in
CRC
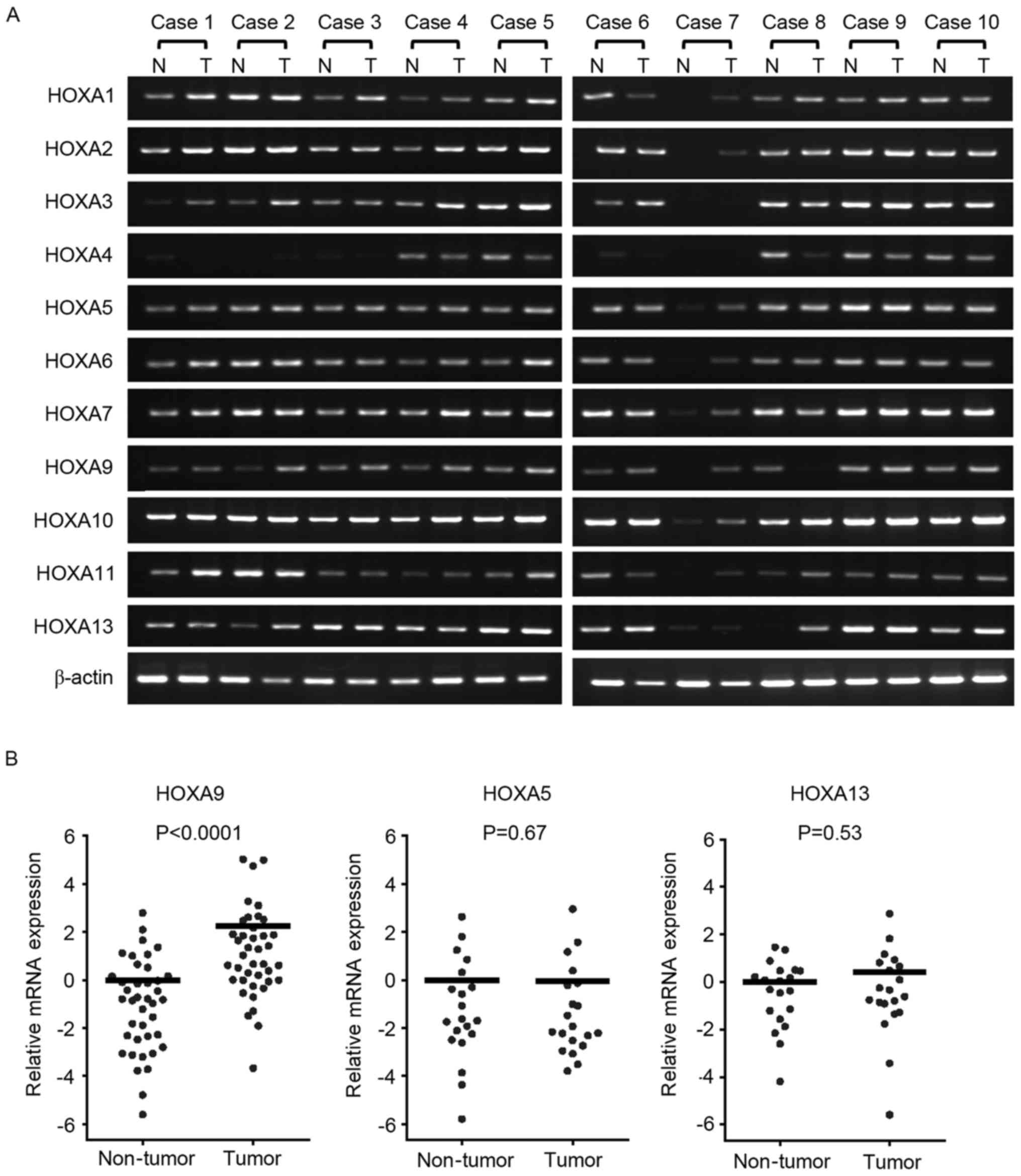
To investigate HOXA gene expression in CRC, the mRNA
expression of each cluster gene was investigated in 10
representative cases by RT-qPCR (Fig.
1A). Although all HOXA genes were detected in tumor and
adjacent non-tumor tissue samples, HOXA9 expression appeared to
have increased in the tumor tissues compared with the non-tumor
tissues as detected by PCR and gel electrophoresis. To investigate
whether there is a difference in expression levels, RT-PCR was
performed, and it was revealed that HOXA9 mRNA expression was
increased in the tumor tissues by 4.71 fold compared with the
non-tumor tissues (P<0.0001; Wilcoxon matched pairs test;
Fig. 1B).
Upregulated HOXA9 expression is
associated with poorer patient outcomes
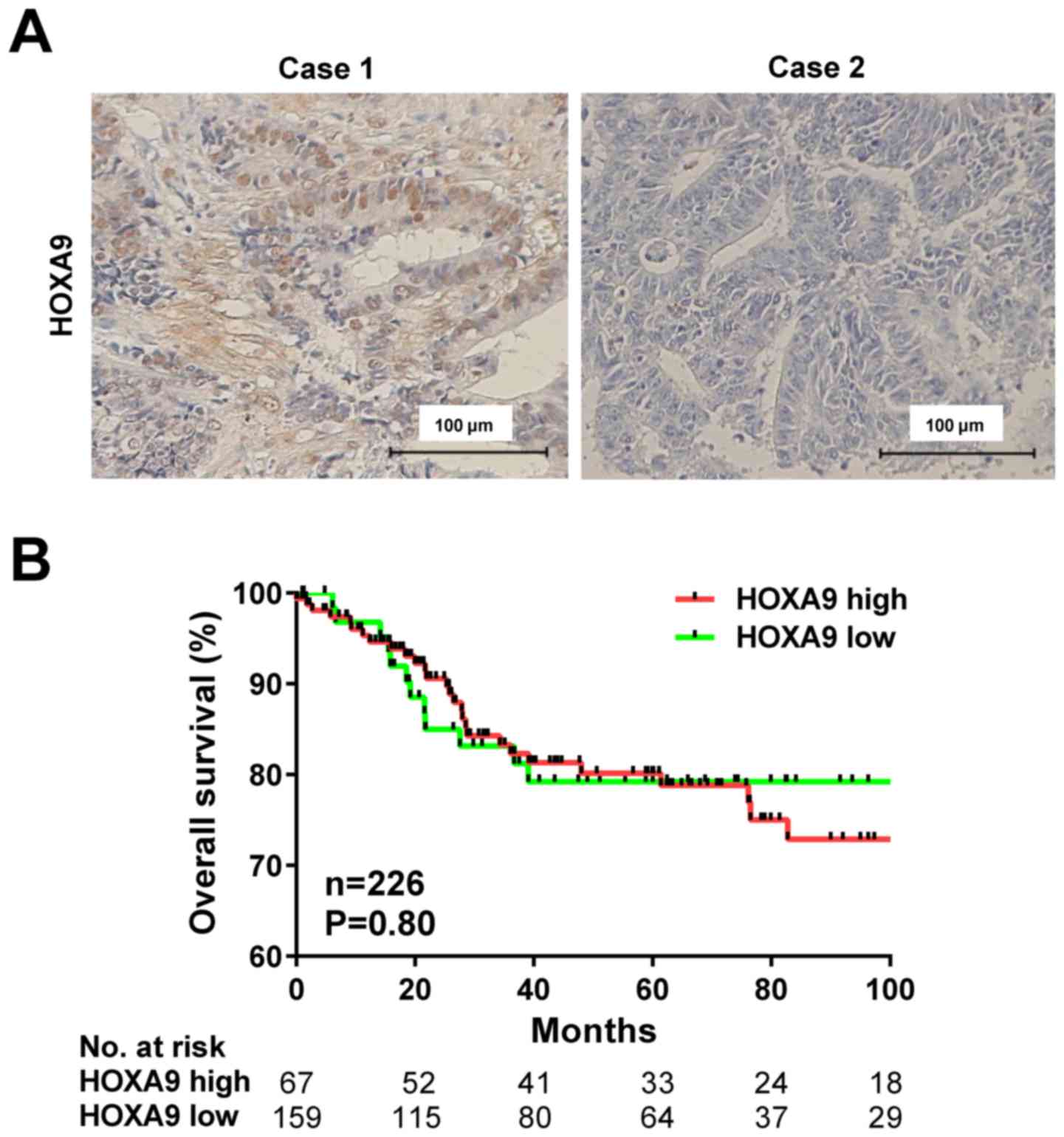
HOXA9 protein expression was evaluated using IHC
staining in 231 patients with CRC (Fig.
2A). HOXA9 expression was detected in the nucleus of the CRC
cells, and positive expression was observed in 161 cases (69.7%),
and negative expression was detected in 70 cases (30.3%).
The association between HOXA9 expression and
clinicopathological factors was analyzed in patients with CRC
(Table II). The rate of positive
expression of HOXA9 was significantly associated with higher TNM
stage (P=0.03) and positive lymph node metastasis (P=0.02).
However, HOXA9 expression was not associated with age, gender,
tumor location, histology, depth of invasion, venous invasion or
liver metastasis. Furthermore, Kaplan-Meier analysis demonstrated
no association between increased HOXA9 levels and overall survival
rate (P=0.80; Fig. 2B).
| Table II.Association between
clinicopathological factors and HOXA9 protein expression as
determined by immunohistochemistry. |
Table II.
Association between
clinicopathological factors and HOXA9 protein expression as
determined by immunohistochemistry.
|
|
| HOXA9 IHC, n |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Total | Positive | Negative | P-value |
|---|
| Total | 231 | 161 | 70 |
|
| Age |
|
|
| 1.00 |
| ≥60
years | 174 | 121 | 53 |
|
| <60
years | 57 | 40 | 17 |
|
| Gender |
|
|
| 0.77 |
|
Male | 136 | 96 | 40 |
|
|
Female | 95 | 65 | 30 |
|
| Stage |
|
|
| 0.03 |
| 0 |
5 |
4 | 1 |
|
| I | 37 | 25 | 12 |
|
| II | 85 | 52 | 33 |
|
|
III | 66 | 52 | 14 |
|
| IV | 38 | 28 | 10 |
|
| Tumor location |
|
|
| 0.65 |
|
Colon | 165 | 113 | 52 |
|
|
Rectum | 66 | 48 | 18 |
|
| Histology |
|
|
| 0.82 |
|
tub | 203 | 142 | 61 |
|
|
por | 28 | 19 | 9 |
|
| Depth |
|
|
| 0.28 |
| T1 | 28 | 19 | 9 |
|
| T2 | 27 | 22 | 5 |
|
| T3 | 159 | 106 | 53 |
|
| T4 | 17 | 14 | 3 |
|
| Lymphatic
invasion |
|
|
| 1.00 |
|
Absent | 48 | 34 | 14 |
|
|
Present | 183 | 127 | 56 |
|
| Venous
invasion |
|
|
| 0.60 |
|
Absent | 50 | 33 | 17 |
|
|
Present | 181 | 128 | 53 |
|
| Lymph node
metastasis |
|
|
| 0.02 |
|
Negative | 135 | 86 | 49 |
|
|
Positive | 96 | 75 | 21 |
|
| Liver
metastasis |
|
|
| 1.00 |
|
Negative | 200 | 139 | 61 |
|
|
Positive | 31 | 22 | 9 |
|
Knockdown of HOXA9 has no effect on
growth in colon cancer cells
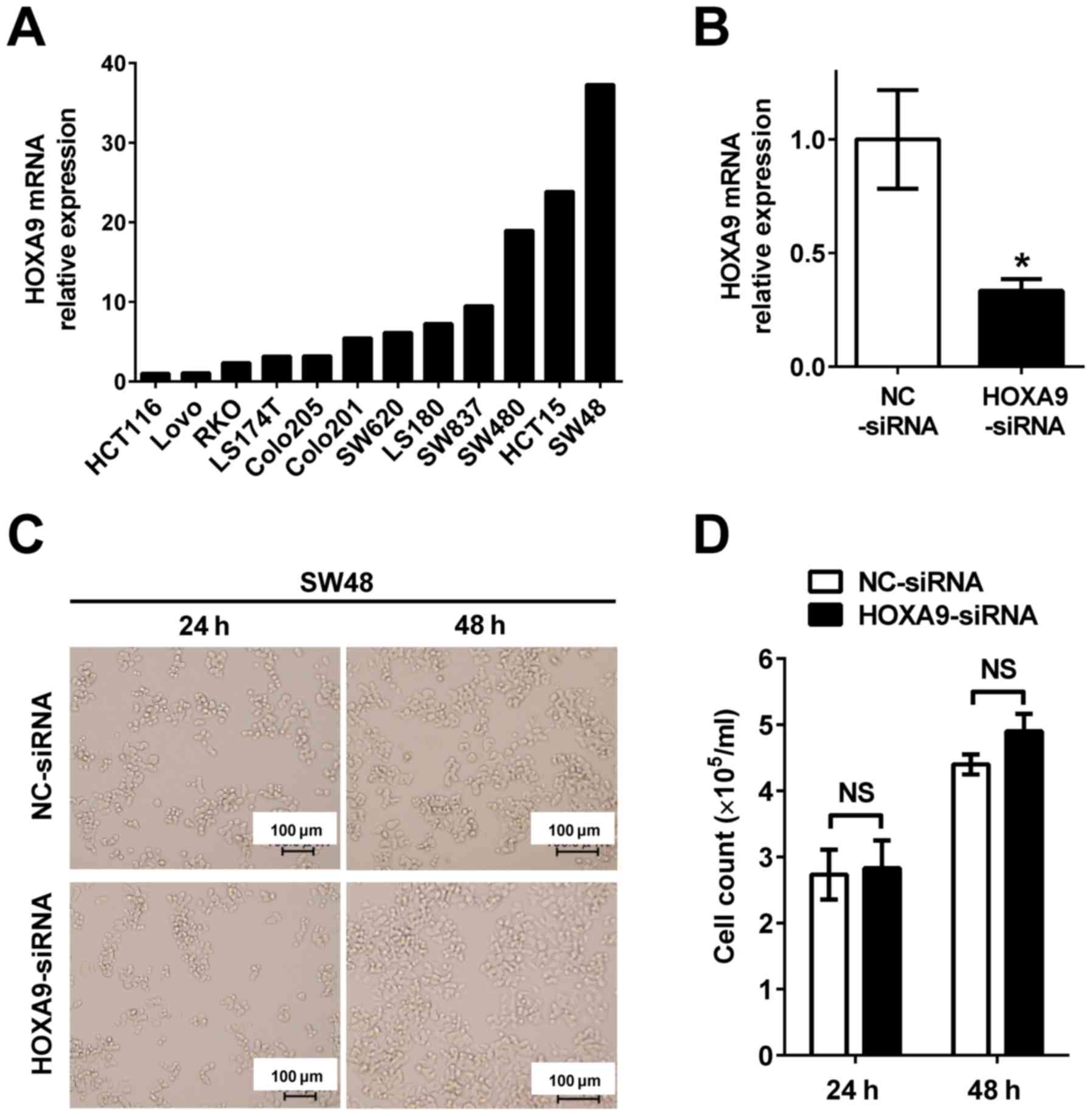
To evaluate the role of HOXA9 in colon cancer
progression, gene knockdown of HOXA9 was performed to investigate
cell proliferation. HOXA9 expression in 12 colon cancer cell lines
(HCT116, LoVo, RKO, LS174T, Colo205, Colo201, SW620, LS180, SW837,
SW480, HCT15 and SW48) was evaluated to select the appropriate cell
line for subsequent experiments (Fig.
3A). As HOXA9 was most significantly expressed in SW48 cells
compared with all other cells, they were chosen for subsequent
knockdown experiments. HOXA9 was subsequently knocked down by
transfection with siRNA oligonucleotide in the SW48 cells
(HOXA9-siRNA) (Fig. 3B). No
morphological changes were observed in the HOXA9 knocked down cells
(Fig. 3C). Furthermore, cell
proliferation was not decreased between HOXA9 knocked down cells
and the negative control (Fig.
3D).
Discussion
In the present study, RT-qPCR analysis indicated
that HOXA9 mRNA was upregulated in CRC tumor tissues compared with
non-tumor tissues. Consistent with the results of the present
study, expression of HOXA9 has been reported to be significantly
increased in esophageal squamous cell cancer (17) and lung cancer (28) compared with normal non-tumor tissues.
However, HOXA9 expression was decreased in hepatocellular
carcinoma, as well as ovary, lung and bladder cancer tissues
(29–32). Such suppression was reported to be due
to hypermethylation of the HOXA9 promoter CpG island (29–32).
Another known regulator of HOXA9 mRNA expression is
microRNA (miR) (33). It has been
reported that HOXA9 is directly suppressed by upregulated miR-196b
expression in non-small cell lung cancer cells (33). Although one study has reported that
miR-196b is downregulated in cervical cancer (34), another study has suggested that HOXA9
is also downregulated in cervical cancer (35). These results indicate that the
expression and aberration mechanisms of HOXA9 are
tumor-specific.
The authors of the present study investigated
whether HOXA9 gene aberrations attributed to high HOXA9 expression
using the publicly available cBioPortal. HOXA9 gene amplification
resulting in high HOXA9 expression was observed in 10/287 (3.5%)
cases of stomach adenocarcinoma (36), 4/109 (3.7%) cases of pancreatic cancer
(37) and 12/184 (6.5%) cases of
esophageal cancer (TCGA provisional). However, HOXA9 gene
amplification resulting in high HOXA9 expression was only detected
in 1/212 (0.5%) cases in CRC (38).
These findings indicated that gene amplification may not be
involved in the increase of HOXA9 mRNA transcript levels in CRC.
The underlying mechanism for the increase of HOXA9 mRNA in CRC
tumors is yet to be elucidated. Notably, a similar upregulation of
mRNA in human tumors has also been reported with other members of
the HOX gene family (12,13,39).
However, the main mechanism involved in this process also remains
unknown (12,13,39).
The present study investigated HOXA9 protein
expression in CRC tumor tissues and found that ~70% of patients
with CRC exhibited positive staining for HOXA9. Furthermore, the
increase in HOXA9 expression was significantly associated with
positive lymph node metastasis in the present cohort. Consistent
with the present results, a number of studies have demonstrated the
involvement of HOX genes in lymph node metastasis (13,20,39). HOXC6
expression was upregulated in lymph node metastasis-positive oral
squamous cell carcinoma compared with lymph node
metastasis-negative oral squamous cell carcinoma (40). By contrast, HOXB7 expression was
downregulated in breast cancer with lymph node metastasis compared
with lymph node-negative breast cancer (41). Notably, no association between HOXA9
expression and patient survival was observed in the present cohort.
This may be due to differences in tumor stage, as the population
comprised primarily stage II and III patients.
In conclusion, to the best of our knowledge, this is
the first study to report that HOXA9 expression is upregulated in
CRC and is associated with lymph node metastasis. Additional
studies are required to reveal the role of HOXA9 and its viability
as a candidate therapeutic target or biomarker.
Acknowledgements
The present study was supported by the Japan Society
for the Promotion of Science KAKENHI (grant no. 15k10143).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katanoda K, Hori M, Matsuda T, Shibata A,
Nishino Y, Hattori M, Soda M, Ioka A, Sobue T and Nishimoto H: An
updated report on the trends in cancer incidence and mortality in
Japan, 1958–2013. Jpn J Clin Oncol. 45:390–401. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mayer RJ, Venook AP and Schilsky RL:
Progress against GI cancer during the American Society of Clinical
Oncology's first 50 years. J Clin Oncol. 32:1521–1530. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brenner H, Kretschmann J, Stock C and
Hoffmeister M: Expected long-term impact of screening endoscopy on
colorectal cancer incidence: A modelling study. Oncotarget.
7:48168–48179. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brenner H, Chang-Claude J, Seiler CM and
Hoffmeister M: Long-term risk of colorectal cancer after negative
colonoscopy. J Clin Oncol. 29:3761–3767. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Venook AP, Weiser MR and Tepper JE:
Colorectal cancer: All hands on deck. Am Soc Clin Oncol Educ Book.
1–89. 2014.
|
|
7
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
de Gramont A, Figer A, Seymour M, Homerin
M, Hmissi A, Cassidy J, Boni C, Cortes-Funes H, Cervantes A, Freyer
G, et al: Leucovorin and fluorouracil with or without oxaliplatin
as first-line treatment in advanced colorectal cancer. J Clin
Oncol. 18:2938–2947. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Douillard JY, Cunningham D, Roth AD,
Navarro M, James RD, Karasek P, Jandik P, Iveson T, Carmichael J,
Alakl M, et al: Irinotecan combined with fluorouracil compared with
fluorouracil alone as first-line treatment for metastatic
colorectal cancer: A multicentre randomised trial. Lancet.
355:1041–1047. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Goldberg RM, Sargent DJ, Morton RF, Fuchs
CS, Ramanathan RK, Williamson SK, Findlay BP, Pitot HC and Alberts
SR: A randomized controlled trial of fluorouracil plus leucovorin,
irinotecan, and oxaliplatin combinations in patients with
previously untreated metastatic colorectal cancer. J Clin Oncol.
22:23–30. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tournigand C, André T, Achille E, Lledo G,
Flesh M, Mery-Mignard D, Quinaux E, Couteau C, Buyse M, Ganem G, et
al: FOLFIRI followed by FOLFOX6 or the reverse sequence in advanced
colorectal cancer: A randomized GERCOR study. J Clin Oncol.
22:229–237. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pezzani L, Milani D, Manzoni F, Baccarin
M, Silipigni R, Guerneri S and Esposito S: HOXA genes cluster:
Clinical implications of the smallest deletion. Ital J Pediatr.
41:312015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Quinonez SC and Innis JW: Human HOX gene
disorders. Mol Genet Metab. 111:4–15. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Novak P, Jensen T, Oshiro MM, Wozniak RJ,
Nouzova M, Watts GS, Klimecki WT, Kim C and Futscher BW: Epigenetic
inactivation of the HOXA gene cluster in breast cancer. Cancer Res.
66:10664–10670. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Strathdee G, Holyoake TL, Sim A, Parker A,
Oscier DG, Melo JV, Meyer S, Eden T, Dickinson AM, Mountford JC, et
al: Inactivation of HOXA genes by hypermethylation in myeloid and
lymphoid malignancy is frequent and associated with poor prognosis.
Clin Cancer Res. 13:5048–5055. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bach C, Buhl S, Mueller D, García-Cuéllar
MP, Maethner E and Slany RK: Leukemogenic transformation by HOXA
cluster genes. Blood. 115:2910–2918. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen KN, Gu ZD, Ke Y, Li JY, Shi XT and Xu
GW: Expression of 11 HOX genes is deregulated in esophageal
squamous cell carcinoma. Clin Cancer Res. 11:1044–1049.
2005.PubMed/NCBI
|
|
18
|
Widschwendter M, Apostolidou S, Jones AA,
Fourkala EO, Arora R, Pearce CL, Frasco MA, Ayhan A, Zikan M,
Cibula D, et al: HOXA methylation in normal endometrium from
premenopausal women is associated with the presence of ovarian
cancer: A proof of principle study. Int J Cancer. 125:2214–2218.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Orlovsky K, Kalinkovich A, Rozovskaia T,
Shezen E, Itkin T, Alder H, Ozer HG, Carramusa L, Avigdor A,
Volinia S, et al: Down-regulation of homeobox genes MEIS1 and HOXA
in MLL-rearranged acute leukemia impairs engraftment and reduces
proliferation. Proc Natl Acad Sci USA. 108:pp. 7956–7961. 2011;
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kanai M, Hamada J, Takada M, Asano T,
Murakawa K, Takahashi Y, Murai T, Tada M, Miyamoto M, Kondo S and
Moriuchi T: Aberrant expressions of HOX genes in colorectal and
hepatocellular carcinomas. Oncol Rep. 23:843–851. 2010.PubMed/NCBI
|
|
21
|
Bhatlekar S, Addya S, Salunek M, Orr CR,
Surrey S, McKenzie S, Fields JZ and Boman BM: Identification of a
developmental gene expression signature, including HOX genes, for
the normal human colonic crypt stem cell niche: Overexpression of
the signature parallels stem cell overpopulation during colon
tumorigenesis. Stem Cells Dev. 23:167–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Union for International Cancer Control
(UICC), . TNM Classification of Malignant Tumors. Gospodarowicz MK,
Wittekind Ch and Sobin LH: 7th. Wiley-Blackwell; Oxford: 2009
|
|
23
|
Sobin LH and Compton CC: TNM seventh
edition: What's new, what's changed: Communication from the
International Union Against Cancer and the American Joint Committee
on Cancer. Cancer. 116:5336–5339. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Okano M, Kumamoto K, Saito M, Onozawa H,
Saito K, Abe N, Ohtake T and Takenoshita S: Upregulated Annexin A1
promotes cellular invasion in triple-negative breast cancer. Oncol
Rep. 33:1064–1070. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tachibana K, Saito M, Imai JI, Ito E,
Yanagisawa Y, Honma R, Saito K, Ando J, Momma T, Ohki S, et al:
Clinicopathological examination of dipeptidase 1 expression in
colorectal cancer. Biomed Rep. 6:423–428. 2017.PubMed/NCBI
|
|
26
|
Saito M, Shiraishi K, Matsumoto K,
Schetter AJ, Ogata-Kawata H, Tsuchiya N, Kunitoh H, Nokihara H,
Watanabe S, Tsuta K, et al: A three-microRNA signature predicts
responses to platinum-based doublet chemotherapy in patients with
lung adenocarcinoma. Clin Cancer Res. 20:4784–4793. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Calvo R, West J, Franklin W, Erickson P,
Bemis L, Li E, Helfrich B, Bunn P, Roche J, Brambilla E, et al:
Altered HOX and WNT7A expression in human lung cancer. Proc Natl
Acad Sci USA. 97:pp. 12776–12781. 2000; View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shin SH, Kim BH, Jang JJ, Suh KS and Kang
GH: Identification of novel methylation markers in hepatocellular
carcinoma using a methylation array. J Korean Med Sci.
25:1152–1159. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Beachy SH, Onozawa M, Silverman D, Chung
YJ, Rivera MM and Aplan PD: Isolated Hoxa9 overexpression
predisposes to the development of lymphoid but not myeloid
leukemia. Exp Hematol. 41:518–529.e5. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Thorsteinsdottir U, Mamo A, Kroon E,
Jerome L, Bijl J, Lawrence HJ, Humphries K and Sauvageau G:
Overexpression of the myeloid leukemia-associated Hoxa9 gene in
bone marrow cells induces stem cell expansion. Blood. 99:121–129.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Aine M, Sjödahl G, Eriksson P, Veerla S,
Lindgren D, Ringnér M and Höglund M: Integrative epigenomic
analysis of differential DNA methylation in urothelial carcinoma.
Genome Med. 7:232015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu SL, Lee DC, Sohn HA, Lee SY, Jeon HS,
Lee JH, Park CG, Lee HY, Yeom YI, Son JW, et al: Homeobox A9
directly targeted by miR-196b regulates aggressiveness through
nuclear Factor-kappa B activity in non-small cell lung cancer
cells. Mol Carcinog. 55:1915–1926. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
How C, Hui AB, Alajez NM, Shi W, Boutros
PC, Clarke BA, Yan R, Pintilie M, Fyles A, Hedley DW, et al:
MicroRNA-196b regulates the homeobox B7-vascular endothelial growth
factor axis in cervical cancer. PLoS One. 8:e678462013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Alvarado-Ruiz L, Martinez-Silva MG,
Torres-Reyes LA, Pina-Sanchez P, Ortiz-Lazareno P, Bravo-Cuellar A,
Aguilar-Lemarroy A and Jave-Suarez LF: HOXA9 is underexpressed in
cervical cancer cells and its restoration decreases proliferation,
migration and expression of epithelial-to-mesenchymal transition
genes. Asian Pac J Cancer Prev. 17:1037–1047. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cancer Genome Atlas Research Network, .
Comprehensive molecular characterization of gastric adenocarcinoma.
Nature. 513:202–209. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Witkiewicz AK, McMillan EA, Balaji U, Baek
G, Lin WC, Mansour J, Mollaee M, Wagner KU, Koduru P, Yopp A, et
al: Whole-exome sequencing of pancreatic cancer defines genetic
diversity and therapeutic targets. Nat Commun. 6:67442015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cancer Genome Atlas Network, .
Comprehensive molecular characterization of human colon and rectal
cancer. Nature. 487:330–337. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bhatlekar S, Fields JZ and Boman BM: HOX
genes and their role in the development of human cancers. J Mol Med
(Berl). 92:811–823. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hassan NM, Hamada J, Murai T, Seino A,
Takahashi Y, Tada M, Zhang X, Kashiwazaki H, Yamazaki Y, Inoue N
and Moriuchi T: Aberrant expression of HOX genes in oral dysplasia
and squamous cell carcinoma tissues. Oncol Res. 16:217–224. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Makiyama K, Hamada J, Takada M, Murakawa
K, Takahashi Y, Tada M, Tamoto E, Shindo G, Matsunaga A, Teramoto
K, et al: Aberrant expression of HOX genes in human invasive breast
carcinoma. Oncol Rep. 13:673–679. 2005.PubMed/NCBI
|