Introduction
Docosahexaenoic acid (DHA; 22:6 n-3) is an important
member of the family of omega-3 polyunsaturated fatty acids and
numerous people have added DHA oil to their daily diet (1,2).
Currently, breast cancer (BC) is one of the most common types of
cancer among women worldwide (3).
Dietary unsaturated fatty acids, particularly DHA, are considered
to serve an important role in reducing the risk of developing BC
(4). Certain evidence has
demonstrated that DHA oil is beneficial for inhibiting mammary
gland carcinogenesis (5–10). The mechanisms of DHA oil on the
inhibition of tumor proliferation and the promotion of apoptosis
are complex (5–10). Among the numerous key factors,
oxidative stress serves an important role in the initiation,
promotion, progression and apoptosis of BC by interfering with the
intracellular signal transduction pathways, and inducing DNA damage
(11). The downregulation of
antioxidant enzyme (AOE) expression levels or their activities have
been revealed to be associated with numerous types of cancer,
including breast, prostate, bladder and hepatic cancer, and
multiple myeloma (12–18). Nevertheless, other studies have
reported no significant changes, higher expression or higher
activity levels of AOEs in certain cancer types. For example,
decreased catalase (CAT), with unchanged glutathione-peroxidase
(GPH-PX) and increased levels of superoxide dismutase (SOD) levels
were reported in the A549 lung cancer cell line, and lung cancer
tissues (19). Therefore, maintaining
the appropriate activity levels of AOEs may be essential in
preventing the development of specific cancer types (20). There are specfic key AOEs with
essential roles in protecting cells from oxidative stress,
including SOD, CAT and GPH-PX. The regulation and increases in AOE
activity are associated with cancer (12). To elucidate the anticancer mechanism
of DHA in BC, antioxidant activities, including that of SOD, CAT
and GSH-PX were analyzed using various assays.
The anticancer mechanisms of DHA are complex
(7). Glucose uptake, glycolytic
metabolism, lactate production and total glucose oxidation have
been demonstrated to be significantly decreased in response to DHA
supplementation, thereby decreasing oxidative metabolism, and
enhancing metabolic injury (7).
Furthermore, the metabolic changes in DHA led to intracellular
cyclic (c)AMP and cGMP levels decreasing by 50% in MDA-MB-231, and
BT-474 cancer cell lines, which mediated the phosphorylation of
AMP-activated protein kinase at Thr172, a metabolic stress marker
(7). It effectively provides
rationale for enhancement oncurrent cancer prevention models and
therapies by combining with dietary sources, including DHA oil
(7).
In the present study, the tumor suppressor ratio,
the activities of SOD, GSH-PX and CAT, the expression of
malondialdehyde (MDA), the expression of cAMP/cGMP, and the
expression of Toll-like receptor 4 (TLR-4) and peroxisome
proliferator activated receptor (PPAR)-α factor were detected in
human malignant BC tissues following treatment with DHA oil. The
results aided in explaining the apoptosis mechanism of DHA on human
malignant BC. The current study has clinical significance for the
further development of DHA oil use in BC.
Materials and methods
Samples and reagents
DHA oil (21) (Bohai
algae DHA oil, content ~40%) was provided by the Food Science and
Engineering Laboratory of Jinzhou Medical University (Jinzhou,
China). Human malignant BC tissue samples were provided by the
First Affiliated Hospital of Jinzhou Medical University. The
approval of removed tissues for research purposes was obtained from
the Ethics Committee of The First Affiliated Hospital of Jinzhou
Medical University. Written informed consent was obtained from each
patient.
Hydrocortison (1 gram) was purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Anti-TLR4 antibody
produced in rabbit (100 µg) was purchased from Sigm-Aldrich (Merck
KGaA). Anti-PPARα rabbit antibody (100 µg) was purchased from Abcam
(Cambridge, UK). Horseradish peroxidase (HRP)-conjugated sheep
anti-rabbit IgG H&L (1 mg) was purchased from Abcam. Trypsin,
Hanks, RPMI-1640 medium and Dulbecco's modified Eagle's medium
(DMEM) was purchased from Gibco (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The 10% fetal bovine serum and 10% normal goat
serum was purchased from Hyclone (GE Healthcare Life Sciences,
Logan, UT, USA). Tris-buffered saline (pH 7.6; cat. no. SG EC-925)
was purchased from Shanghai Sango Biotechnology Co. Ltd. (Shanghai,
China). Other reagents were analytical solvents and purchased from
Hongda Co. (Jinzhou, China; https://0ds13046577.atobo.com.cn/).
Human malignant BC tissue culture
Human malignant BC tissues were obtained from the
Breast Surgery Department of the First Affiliated Hospital of
Jinzhou Medical University. The BC tissues were sterilized with 75%
alcohol for 2 min and then washed three times with RPMI-1640. The
BC tissues were sliced into small pieces of 1–2 mm3.
Each BC tissue sample was cultured in RPMI-1640 medium with or
without DHA oil. One of BC tissue samples was used as a control
(control group) and the rest were treated with DHA oil (~40%) with
final DHA concentrations of 100 µg/ml (first group), 150 µg/ml
(second group) and 200 µg/ml (third group). Each group contained
three samples of BC tissues. Each individual BC was placed in a
3.5-cm culture well. All wells were cultured for 24 h with 5%
CO2 at 37°C.
After being cultured with or without DHA for 24 h,
the BC tissues were embedded in paraffin to observe the effect of
DHA oil on the morphology of BC. Five random sections were obtained
from each sample. For research on the signal transduction pathway
of DHA, BC tissues were cultured for 24 h in RPMI-1640 medium with
or without DHA oil, and then the total protein was extracted from
the BC tissues. The total protein content was used to detect the
effect of DHA on BC antioxidant activities. Each sample was
measured three times in parallel.
Histological observation
For hematoxylin-eosin (H.E.) staining, sections were
rinsed in double distilled water and were incubated with
hematoxylin solution stain for 5 min at 25°C. After 5 min, sections
were washed with running tap water, sequentially followed by
differentiation for 30 sec in 1% acid-alcohol (hydrochloric acid
and ethanol) at 25°C and then washed for 1 min again with running
tap water. Subsequently, all sections were stained with eosin for
30 sec and then dehydrated with different concentrations alcohol
(70, 80, 90 and 100%) for 2 min each. Sections were covered with
xylene-based mounting medium, after two changes of xylene for 5 min
each time at 25°C. The effect of DHA on the H.E. BC sections was
observed using an inverted microscope (OLYMPUS, IX73+DP73, 12V 100W
halogen lamp). Analysis of the tissue areas occupied was performed
using Image Pro 5.0 Plus software (Media Cybernetics, Inc.,
Rockville, MD, USA). Suppressor ratio (%)=(DHA treatment group
entity tissue area-the control group entity tissue area)/control
group entity tissue area ×100%.
Effect of DHA on the antioxidant
activities of BC tissues
Detection of the activities of
antioxidant enzymes
The BC tissues were centrifuged at 1,500 × g for 15
min at 4°C to remove the debris following homogenizing with normal
saline solution (0.9% sodium chloride). The supernatant was
transferred into new tubes for the evaluation of the SOD, CAT and
GSH-PX activities. The measurements were performed following the
manufacturer's protocol of an assay kit (SOD: cat. no. A001-1; CAT:
cat. no. A007-2; GSH-PX: cat. no. A005; Nanjing KeyGen Biotech Co.,
Ltd., Nanjing, China).
SOD activities were determined (at 550 nm) using the
Xanthine oxidase method and expressed as nU/mg protein (22). CAT was measured by the reaction of CAT
scavenging H2O2, and ammonium molybdate was
added to generate a pale yellow complex (maximum absorption peak at
405 nm). CAT activities were expressed as mU/mg protein (23). GSH-PX was reacted with
dithiobis-nitrobenzoic acid to produce a yellow compound,
5-dithio-bis2-nitrobenzoic acid dithiobis-nitrobenzoic acid anion
(maximum absorption peak at 420 nm). The concentrations of GSH-PX
were expressed as nU/mg protein (24).
Detection of MDA content
Fatty-acid peroxidation MDA content in the
homogenate (0.1 ml) was measured using the thiobarbituric acid
(TBA) method according to the manufacturer's protocol (cat. no.
A003-1; Nanjing KeyGen Biotech Co., Ltd.). MDA and TBA condensate
to produce a red product (maximum absorption peak at 532 nm). Thus,
the MDA content was calculated by measuring the 532 nm absorbance
and expressed as nmol/mg protein (23,24).
cAMP and cGMP assays
The cAMP and cGMP content of BC tissues were
determined following the manufacturer's protocol (cAMP: cat. no.
80204; cGMP: cat. no. 80104; Neweast Biotech Company, Wuhan, China)
by ELISA. The cAMP and cGMP extracts were diluted (1:10) with
sample diluent. Optical densities were measured at 450 nm (25,26).
Immunohistochemistry
The expression of TLR-4 factor was measured by
immunohistochemistry. Briefly, 4-µm thick sections were
deparaffinized and the endogenous peroxidase activity was blocked
for 10 min with 3% hydrogen peroxide at 25°C. Subsequently,
sections were treated for 30 min with 10% normal goat serum in
Tris-buffered saline (pH=7.6) at 37°C. Then, sections were
incubated with monoclonal anti-TLR4 rabbit antibody (1:200; cat.
no. PRS3141-100UG) overnight at 4°C. After washing three times with
PBS, they were incubated with the HRP-sheep anti-rabbit IgG H&L
secondary antibody (1:500; cat. no. ab6721) at room temperature for
1 h, followed by incubation with the color reagent
3,3′-diaminobenzidine for 3 min at 25°C (26). Analysis of the average gray density
value (GDV) of TLR-4 was performed using Image Pro 5.0 Plus
software.
Western blotting
All BC tissues were stored within liquid nitrogen
prior to protein extraction. Briefly, tissues were homogenized with
0.5 ml ice-cold lysis buffer (pH 7.5, 20 mM Tris-HCl, 1 mM EDTA, 1
mM DTT, 5 mM MgCl2, 20 µg/ml aprotinin, 2 mM sodium
orthovanadate and 1 mM PMSF). The homogenates were centrifuged at
10,000 × g at 4°C for 20 min and the supernatant was removed. The
protein concentration was determined using the bicinchoninic acid
method with bovine serum albumin as the standard. Samples (30 µg/ml
protein per lane) were boiled for 5 min, separated using 15%
SDS-PAGE and transferred onto a nitrocellulose membrane (EMD
Millipore, Billerica, MA, USA). Subsequently, membranes were
blocked with 5% bovine serum albumin at room temperature for 1.5 h
and incubated with the anti-PPARα rabbit antibody (1:1,000; cat.
no. ab8934) at 4°C for 12 h. The membrane was then washed three
times with TBS Tween 20 buffer and incubated with the secondary
HRP-conjugated sheep anti-rabbit IgG H&L antibody (1:200; cat.
no. ab6721) at room temperature for 1 h. The GDV of specific bands
was measured with Quantity One version 4.62 software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) (27).
Statistical analysis
Each experiment was performed in parallel three
times. All data are expressed as the mean ± standard deviation.
Statistical analysis was performed using one-way analysis of
variance and was analyzed further by Tukey's honest significant
difference test (28). Different
lowercase letters in the same column represent significant
differences at P≤0.05 and different capital letters represent
significant differences at P≤0.01. All data analyses were conducted
using SPSS 19.0 software (IBM Corp., Armonk, NY, USA).
Results
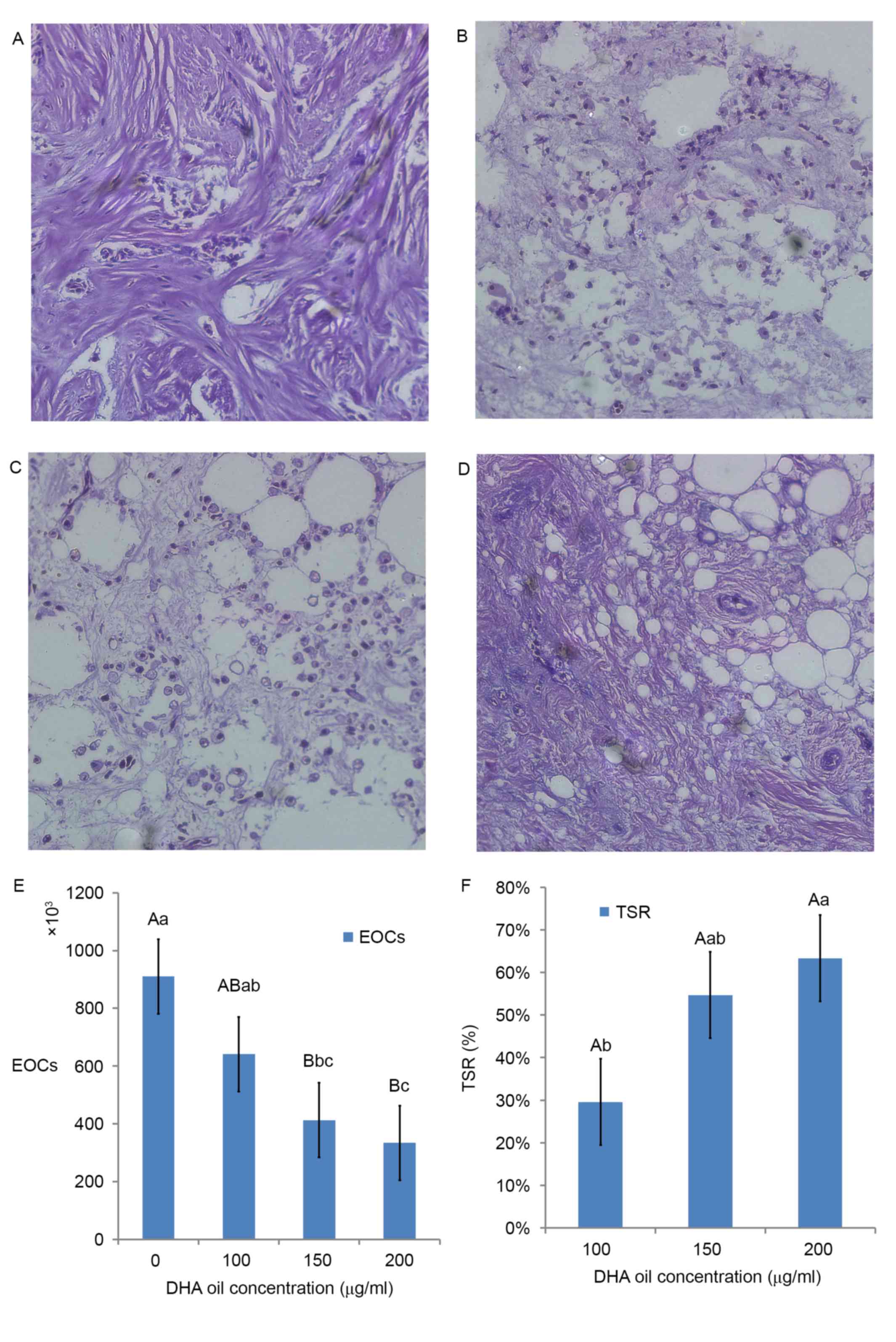
Effects of DHA on BC morphology and
the suppressor ratio of DHA in BC
From the results of paraffin sections (H.E.
staining), as DHA oil concentration increased, the morphology of BC
cells became more irregular. For example, apoptosis of BC cells was
more evident compared with the control group (Fig. 1A). The surrounding of tissues appeared
to be shedding, phagocytized and the cell connections had
disappeared (Fig. 1B). There were
more vesicles observed within cells (Fig.
1C). The overall volume of BC cells had shrunk, whereas the
cytoplasmic density of BC cells had increased. Chromatin
agglutination occurred, and the nuclear membrane and nucleolus were
broken or absent (Fig. 1D).
In Fig. 1E, the area
sizes of human malignant breast tissues in the 150 µg/ml DHA group
were significantly reduced compared with the control (0 µg/ml DHA
group). The suppressor ratio of DHA in the 200 µg/ml DHA group was
significantly higher compared with the ratio of the 100 µg/ml DHA
group (Fig. 1F).
Effect of DHA on the activities of
AOEs
The activities of t-SOD, CAT and GSH-PX in the BC
tissues increased in a DHA dose-dependent manner. Compared with the
control group (500.33±23.20 nU/mg protein), the activities of SOD
was significantly enhanced by 22.4% in the 100 µg/ml DHA group
(612.37±14.98 nU/mg protein; P<0.001) and reached the maximum at
79.24% in the 200 µg/ml DHA group (896.78±38.87 nU/mg protein;
P<0.001; Fig. 2A).
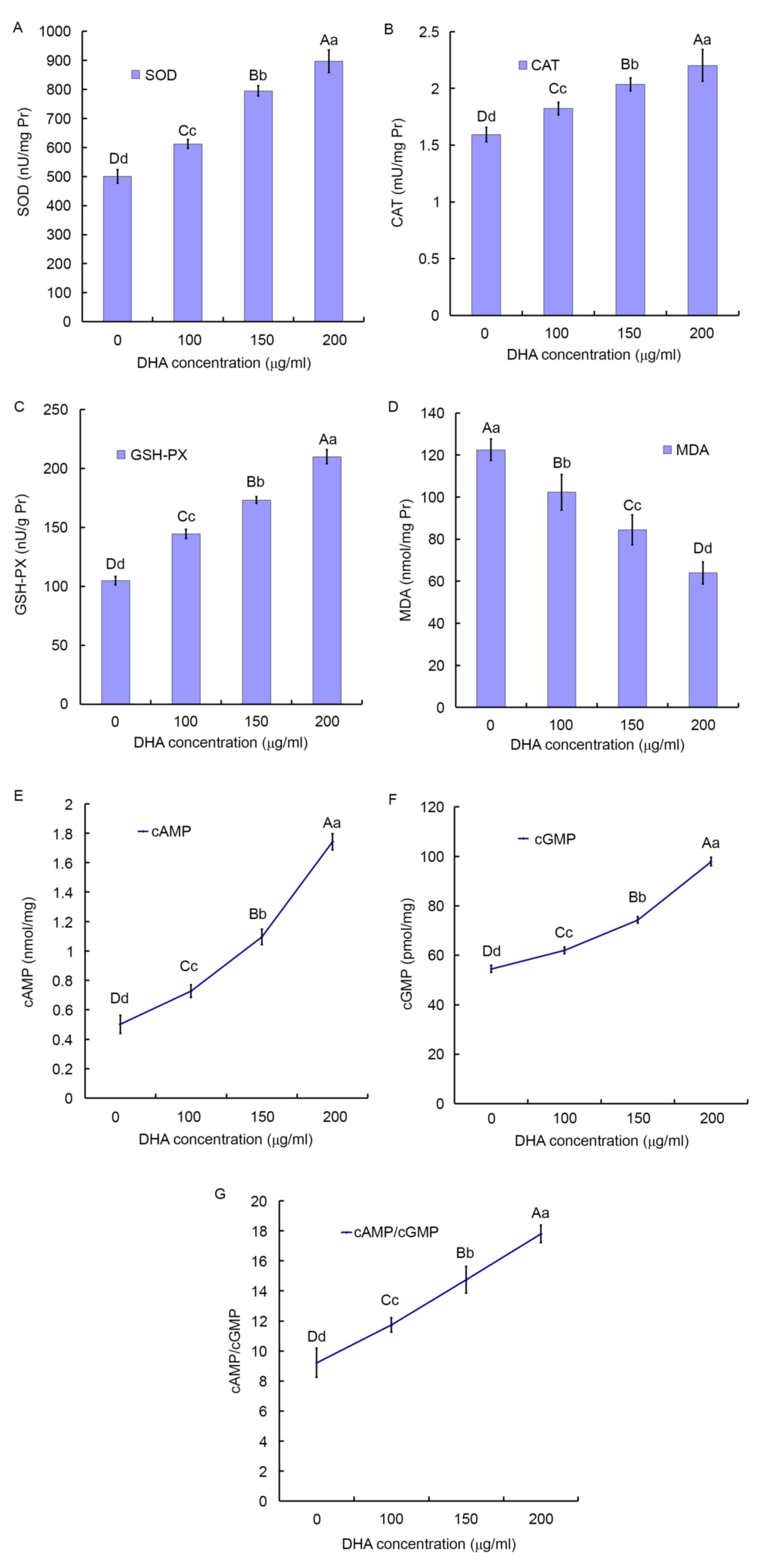 | Figure 2.Effect of DHA on the activity of
antioxidant enzymes and secondary messengers. (A) SOD, (B) GSH-PX
and (C) CAT activities, and (D) MDA concentration in the BCTs
incubated for 24 h with increasing concentrations of DHA. The
results are expressed as the mean ± standard deviation of four
independent experiments, two samples per experiment performed in
triplicate. The homogeneity of variance comparisons were analyzed
using SPSS 19.0 software. Statistical analysis was analyzed by
Tukey's honest significant difference test following one-way
analysis of variance. Different lowercase letters in the same
column represent significant differences at P≤0.05 and different
capital letters represent significant differences at P≤0.01, but
not significantly different (P>0.05) for the same letter
(Table I). DHA serves as a death
receptor via cancer-specific cAMP/cGMP upregulation. Effects of DHA
on (E) cAMP and (F) cGMP levels in the BCTs following 24 h. (G)
Effects of DHA on the ratio of cAMP/cGMP levels in the BCTs
following 24 h. Different lowercase and uppercase letters indicate
the significance of multiple comparisons tests of the results.
Different lowercase letters in the same column represent
significance differences at P≤0.05 and different capital letters
represent significant differences at P≤0.01 (Table I). SOD, super oxide dismutase; DHA,
docosahexaenoic acid; CAT, catalase; GSH-PX,
glutathione-peroxidase; MDA, malondialdehyde; cAMP, cyclic AMP;
cGMP, cyclic GMP; BCTs, breast cancer tissues. |
Compared with the control group (1.59±0.06 mU/mg
protein), the activities of CAT was enhanced by 14.44% the 100
µg/ml DHA group (1.82±0.05 mU/mg protein; P<0.001) and reached
the maximum at 38.30% in the 200 µg/ml DHA group (2.20±0.14 mU/mg
protein; P<0.001; Fig. 2B).
Compared with the control group (104.91±3.51 mU/mg
protein), the activity of GSH-PX was enhanced by 37.75% in the 100
µg/ml DHA group (144.51±3.95 mU/mg protein; P<0.001) and reached
the maximum at 100.17% in the 200 µg/ml DHA group (210.00±6.01
mU/mg protein; P<0.001; Fig.
2C).
Effect of DHA on the concentration of
MDA in the BC tissues
In Fig. 2D, the MDA
concentration of BC tissues was significantly reduced compared with
the control. Compared with the control group (122.49±5.15 nmol/mg
protein), the mean MDA concentration in the 100 µg/ml DHA group
(102.31±8.49 nmol/mg protein) was significantly decreased by 16.48%
(P<0.001) and in the 200 µg/ml DHA group was decreased by 52.22%
(63.96±5.24 nmol/mg protein; P<0.001).
DHA serves as a death receptor via the
upregulation of cancer-specific cAMP/cGMP
Intracellular cAMP and cGMP are the secondary
messengers involved in the generation of BC. The levels of cAMP and
cGMP of the BC tissues with DHA treatment were significantly higher
compared with the control groups (Fig. 2E
and F).
Compared with the control group (0.50±0.062
nmol/mg), the mean concentration of cAMP in the 100 µg/ml DHA group
(0.73±0.043 nmol/mg; P<0.001) significantly increased by 44.78%
and reached the maximum at 246.77% in the 200 µg/ml DHA group
(1.74±0.056 nmol/mg; P<0.001). Compared with the 100 and 150
µg/ml DHA group (0.73±0.043 and 1.10±0.053 nmol/mg, respectively),
the mean concentration of cAMP in the 200 µg/ml DHA group was
significantly increased by 139.52, and 59.13%, respectively
(Fig. 2E).
Compared with the control group (54.50±1.42 m
nmol/mg), the mean concentration of cGMP in the 100 µg/ml DHA group
(62.01±1.41 nmol/mg; P<0.001) significantly increased by 13.79%
and reached the maximum at 79.64% in the 200 µg/ml DHA group
(97.89±1.78 nmol/mg; P<0.001). Compared with the 100 and 150
µg/ml DHA group (62.01±1.41 and 74.32±1.27 nmol/mg, respectively),
the mean concentration of cGMP in the 200 µg/ml DHA group
significantly increased by 57.87, and 31.72% respectively (Fig. 2F).
Compared with the control group, the 200 µg/ml DHA
group revealed significantly higher concentrations of cAMP/cGMP
with a 2-fold increase (Fig. 2G).
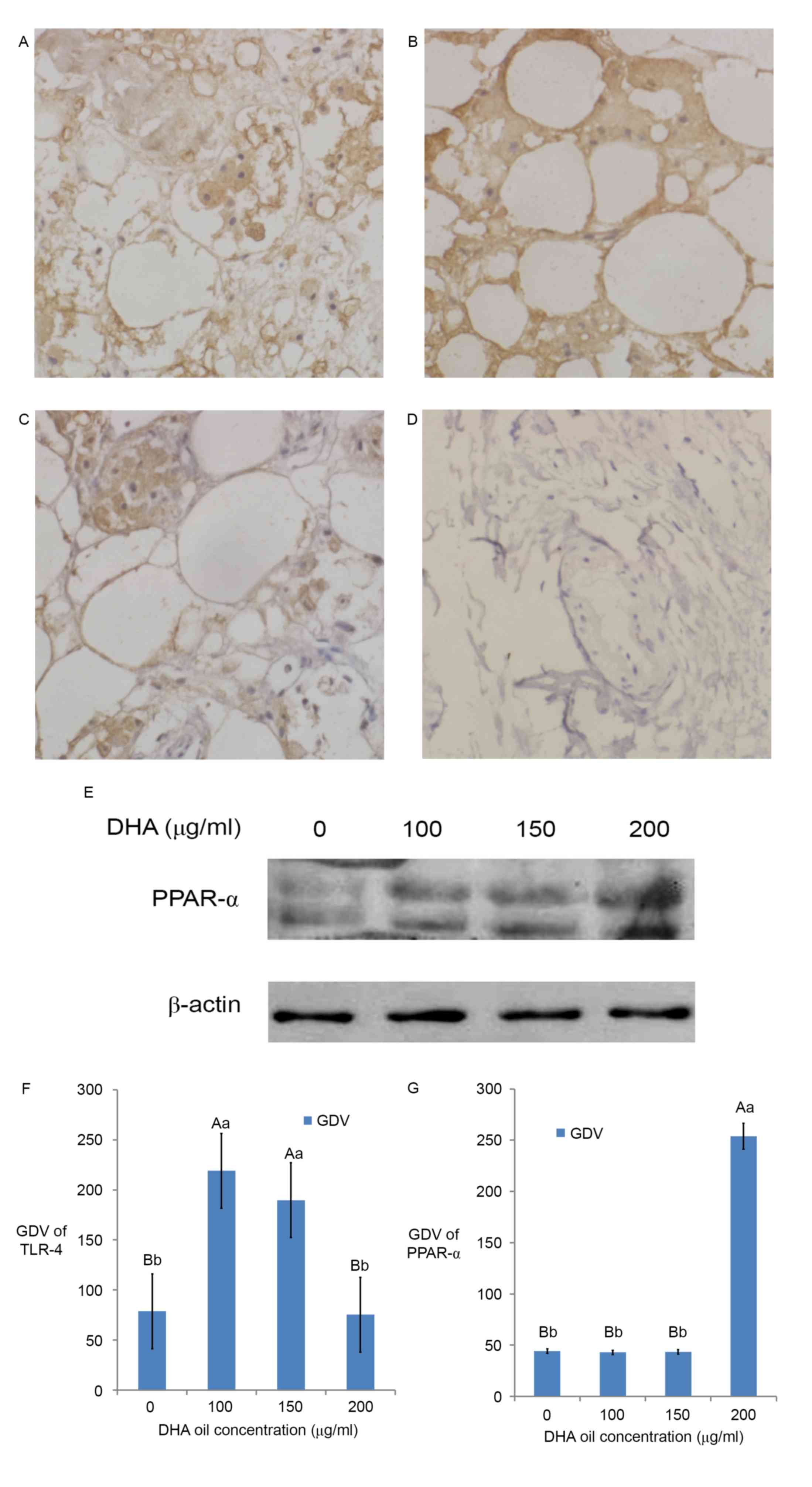
Effect of DHA on the expression and
localization of TLR-4 factor by immunohistochemistry
The immunohistochemical staining of the control
group (Fig. 3) was brown in the cell
membrane and cytoplasm. The GDV of TLR-4 showed that TLR-4 mainly
existed in the cell membrane and cytoplasm (Fig. 3A-D and F). The results revealed that
there was TLR-4 expression in the immunohistochemistry sections of
breast cancer tissues (Fig. 3A and
F), thus TLR-4 may participate in the generation of BC and
localized primarily in the membrane and cytoplasm of BC cells. When
DHA was added at 100 µg/ml (Fig. 3B),
TLR-4 expression reached the maximum (Fig. 3F); however, the expression of TLR-4 in
the 200 µg/ml (Fig. 3D) DHA group was
similar to that of the control group (P>0.05, Fig. 3F). The expression of TLR-4 in the 150
µg/ml (Fig. 3C) DHA group was similar
to the 100 µg/ml DHA group (Fig. 3B;
P>0.05, Fig. 3F).
Effect of DHA on the expression of
PPAR-α by western blotting
The western blotting result (Fig. 3E) and the GDV (Fig. 3G) of the control group bands revealed
that PPAR-α protein expressed in breast cancer tissue. The results
revealed that PPAR-α participated in the generation of BC in
Fig. 3E and G. The expression of
PPAR-α in BC tissues increased with DHA and reached the maximum in
the 200 µg/ml DHA group (Fig. 3E;
P<0.01; Fig. 3G). The expression
of PPAR-α in the control group, 100 µg/ml DHA group and 150 µg/ml
DHA group are similar (Fig. 3E;
P>0.05).
Discussion
The results of the H.E. staining in the present
study demonstrated that DHA oil induced the apoptosis of BC cells.
The suppressor ratio of DHA treatment revealed that DHA
significantly inhibited the growth and induced the apoptosis of BC
cells.
Oxidative stress is the most important cause of cell
damage, leading to the occurrence and development of cardiovascular
disease. AOEs protect cells from free radicals and oxidative
stress, contributing to the prevention of cancer. Numerous studies
have reported that the upregulated expression or higher activity of
AOEs may be used as effective strategies for cancer prevention and
therapy. For example, MRN-100, as an adjuvant therapy was
demonstrated to be effective in the treatment of esophageal/gastric
carcinoma, exerting an antioxidant effect in the stomach and blood
tissues by increasing the levels of GSH-PX, SOD, CAT, GSH-PX, and
the total antioxidant capacity (29).
Ganoderma lucidum significantly enhanced the levels of SOD,
CAT and GPH-PX in the plasma, liver, and mammary tissues, thus
being an effective chemopreventive agent against BC (30). Tangeretin increased the levels of
AOEs, including SOD, CAT, GST and GSH-PX significantly, indicating
to be effective, and efficient for the treatment of BC (31).
SOD is a class of AOEs that catalyzes the
dismutation of superoxide radicals into H2O2
and O2. Subsequently, CAT and GSH-PX catalyze
H2O2 decomposition to H2O and
O2 (20).
H2O2 is not only a reactive oxygen species,
but also a major signaling molecule (32). Multiple studies have indicated that
mitochondrial H2O2 is a direct and effective
apoptosis inducer (32,33). The present study demonstrated that DHA
was able to simultaneously upregulate the expression levels and
activities of SOD, CAT, and GSH-PX in BC tissues. Therefore, DHA
may inhibit the proliferation of BC cells and the associated
oxidative stress mechanism.
Oxidative stress produces fatty-acid peroxidation
whose metabolites possess high toxicities and mutagenic properties
(34). The main fatty-acid
peroxidation is MDA (34). The
results in the current study revealed that DHA significantly
decreased the MDA concentration of BC tissues.
The cyclic nucleotides cAMP and cGMP have been
recognized as important signaling molecules within cells. Under
normal physiological conditions, cyclic nucleotides regulate a
myriad of biological processes. In addition, altered cyclic
nucleotide signaling has been observed in a number of
pathophysiological conditions, including cancer. Several studies
have demonstrated that activation of cyclic nucleotide signaling
leads to the inhibition of proliferation and activation of
apoptosis in numerous types of cancer cells, such as bladder
(35,36), breast (37–41), colon
(42–44), hepatoma (45), leukemia (46,47), lung
(36,48), lymphoma (49,50),
ovarian (36,51), pituitary (52), prostate (36) and skin cancer (53). In the present study, the results
demonstrated that DHA produced significant increase in the ratio of
cAMP/cGMP levels (P<0.001), suggesting that DHA inhibits the
proliferation and induces apoptosis of BC cells by increasing the
ratio of cAMP/cGMP.
Increasing evidence suggests an association between
chronic inflammation and cancer development (54). Recent evidence suggests that
inflammation and oxidative stress play pivotal roles in the
development of clinical conditions like cancers (55) and metabolic syndrome (56). Nonetheless, the underlying molecular
signaling pathways associating inflammation, oxidative stress and
BC cell death are not well defined.
TLR-4 is an important member of the Toll-like
receptor family that are exogenous or endogenous ligands, and
activate the nuclear factor (NF)-κB signal transduction pathway and
the transcription of the early inflammatory cytokine genes. Ahmed
et al (57) demonstrated that
lipopolysaccharide, the TLR-4 agonist, enhances 4T1 tumor growth
and migration, by increasing the rate of angiogenesis, vascular
invasiveness, and tumor invasion. This effect is more evident in
TLR-4−/− mice, suggesting that TLR-4 on host immune
cells may serve an essential role in inhibiting BC genesis and
tumor metastasis (58). TLR4 exerts
both a defensive role at the host level and a negative role at the
cancer cell level in this murine metastatic breast tumor model
(58). Other data suggested that the
TLR-4 agonist may induce pro- or anti-tumorigenic effects (57–60). The
results in the present study revealed that TLR-4 was highly
expressed in the BC tissue of the 100 µg/ml DHA treatment group. In
the BC tissue, TLR-4 was localized in the cell membrane and
cytoplasm, similar to that observed by Yang et al (61). The expression of TLR-4 in the
cytoplasm may be due to the presence of BC with lymph node
metastasis. Thus, this suggests that TLR-4 participates in the
generation of BC and DHA upregulated TLR-4 to induce the apoptosis
of BC. The expression of TLR-4 was significantly reduced when DHA
was at 200 µg/ml, this may be due to the fact that different
concentrations of DHA use different signaling pathways in breast
cancer. The low concentrations DHA (100 µg/ml) stimulated the
expression of TLR-4 to induce the apoptosis of BC. However, high
concentrations DHA (200 µg/ml) decreased the expression of TLR-4
signaling molecules to promote the BC tissue apoptosis
thoroughly.
PPAR-α, a nuclear transcription receptor, belongs to
the PPAR family, and regulates the expression of numerous genes and
proteins (62). Activation of PPAR-α
has been reported to serve an important role in glucose
homeostasis, fatty acid oxidation, lipid metabolism and the
inflammatory process (63). The
activation of PPAR-α was demonstrated to block the transcription of
NF-κB and activator protein-1 signaling pathways (62–64).
Previously, PPAR-α-specific agonists have been reported to inhibit
the proliferation of various cancer cells in cultured cell lines
and in engraft nude mouse models (65–70).
Yessoufou et al (71) revealed
that TLR-4 mutant mice exhibited significantly higher PPAR-α
expression levels following a methionine/choline-deficient diet,
while levels in wild types did not change. This suggests that high
expression levels of TLR-4 increase the expression of PPAR-α.
However, little is known regarding the molecular and cellular
mechanisms of PPAR-α-mediated growth inhibition of cancer cells. In
the present study, it was demonstrated that DHA promoted the
expression of PPAR-α and we hypothesized that DHA regulates the
glycolipid metabolic pathways in BC tissues, thus leading to the
apoptosis of BC cells.
In conclusion, DHA induced the activities of SOD,
CAT and GSH-PX, and decreased the concentration of MDA in the BC
tissues. Furthermore, DHA significantly increased the ratio of
cAMP/cGMP levels, and promoted the expression of TLR-4 and PPAR-α,
in order to induce the apoptosis of BC cells (Fig. 4). DHA may be used as a dietary
treatment or for prevention of BC in the future.
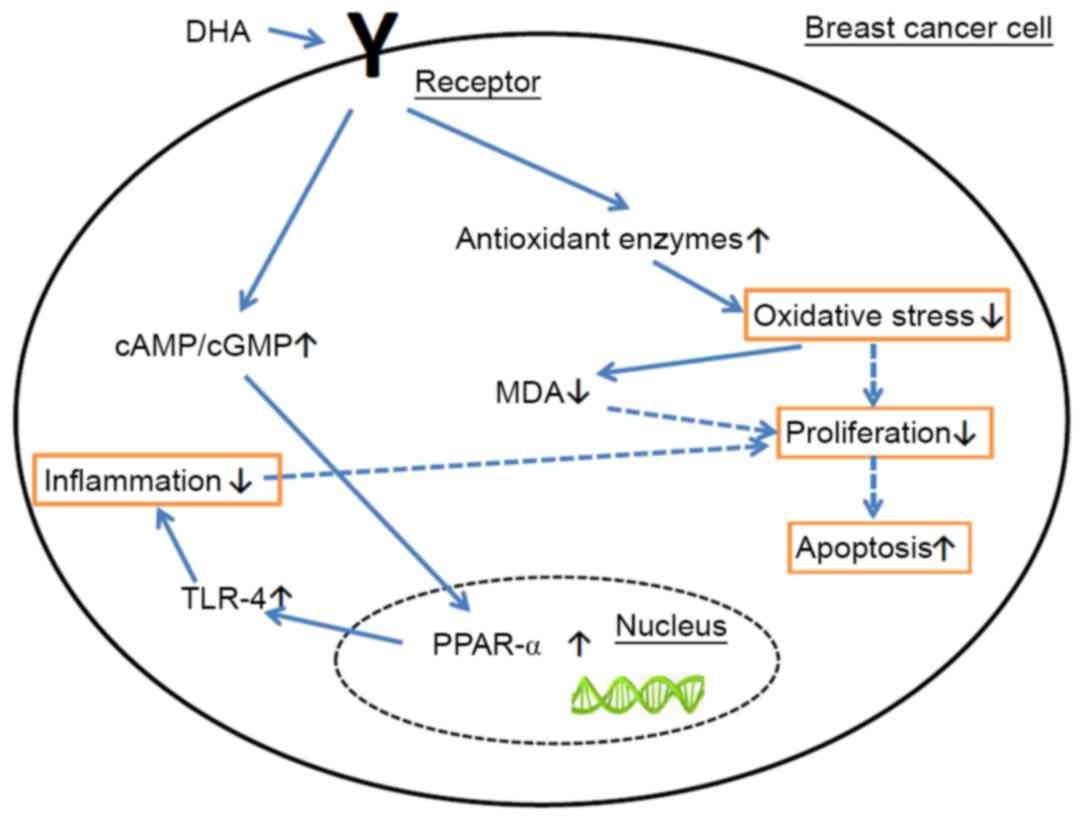 | Figure 4.Apoptosis mechanism of DHA. DHA
induced the levels of SOD, CAT, and GSH-PX and decreased the
concentration of MDA in the BC tissues. DHA increased the ratio of
cAMP/cGMP levels and promoted the expression of TLR-4 and PPAR-α,
in order to induce the apoptosis of BC cells. DHA, docosahexaenoic
acid; PPAR, peroxisome proliferator activated receptor; TLR-4,
Toll-like receptor 4; CAT, catalase; MDA, malondialdehyde; cAMP,
cyclic AMP; cGMP, cyclic GMP. |
Acknowledgements
The present study was supported by The National
Natural Science Fund of China (grant no. 81401189), the Chancellor
Hong Boze Fund of Jinzhou Medical College (grant no.
XZJJ20130101-03) and The Natural Science Foundation of Liaoning
Province (grant no. 20170540385).
References
|
1
|
Meadus WJ, Turner TD, Dugan ME, Aalhus JL,
Duff P, Rolland D, Uttaro B and Gibson LL: Fortification of pork
loins with docosahexaenoic acid (DHA) and its effect on flavour. J
Anim Sci Biotechnol. 4:462013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Opperman M, Marais de W and Spinnler
Benade AJ: Analysis of omega-3 fatty acid content of South African
fish oil supplements. Cardiovasc J Afr. 22:324–329. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DeSantis C, Ma J, Bryan L and Jemal A:
Breast cancer statistics, 2013. CA Cancer J Clin. 64:52–62. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Murff HJ, Shu XO, Li H, Yang G, Wu X, Cai
H, Wen W, Gao YT and Zheng W: Dietary polyunsaturated fatty acids
and breast cancer risk in Chinese women: A prospective cohort
study. Int J Cancer. 128:1434–1441. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Blanckaert V, Kerviel V, Lépinay A,
Joubert-Durigneux V, Hondermarck H and Chénais B: Docosahexaenoic
acid inhibits the invasion of MDA-MB-231 breast cancer cells
through upregulation of cytokeratin-1. Int J Oncol. 46:2649–2955.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu J and Ma DW: The role of n-3
polyunsaturated fatty acids in the prevention and treatment of
breast cancer. Nutrients. 6:5184–5223. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mouradian M, Kikawa KD, Dranka BP, Komas
SM, Kalyanaraman B and Pardini RS: Docosahexaenoic acid attenuates
breast cancer cell metabolism and the Warburg phenotype by
targeting bioenergetic function. Mol Carcinog. 54:810–820. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xue M, Wang Q, Zhao J, Dong L, Ge Y, Hou
L, Liu Y and Zheng Z: Docosahexaenoic acid inhibited the
Wnt/β-catenin pathway and suppressed breast cancer cells in vitro
and in vivo. J Nutr Biochem. 25:104–110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rahman MM, Veigas JM, Williams PJ and
Fernandes G: DHA is a more potent inhibitor of breast cancer
metastasis to bone and related osteolysis than EPA. Breast Cancer
Res Treat. 141:341–352. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lijing G, Xingyuan Q, Zhuping S, Jialing
G, Bing L, et al: Research progress in the tumor suppression
mechanisms of marine polyunsaturated fatty acids DHA. Sci Technol
Food Industry. 22:385–391. 2013.
|
|
11
|
Klaunig JE and Kamendulis LM: The role of
oxidative stress in carcinogenesis. Annu Rev Pharmacol Toxicol.
44:239–267. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Khan MA, Tania M, Zhang DZ and Chen HC:
Antioxidant enzymes and cancer. Chin J Cancer Res. 22:87–92. 2010.
View Article : Google Scholar
|
|
13
|
Arsova-Sarafinovska Z, Eken A, Matevska N,
Erdem O, Sayal A, Savaser A, Banev S, Petrovski D, Dzikova S,
Georgiev V, et al: Increased oxidative/nitrosative stress and
decreased antioxidant enzyme activities in prostate cancer. Clin
Biochem. 42:1228–1235. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
De Craemer D, Pauwels M, Hautekeete M and
Roels F: Alterations of hepatocellular peroxisomes in patients with
cancer. Catalase cytochemistry and morphometry. Cancer.
71:3851–3858. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Elchuri S, Oberley TD, Qi W, Eisenstein
RS, Jackson Roberts L, Van Remmen H, Epstein CJ and Huang TT:
CuZnSOD deficiency leads to persistent and widespread oxidative
damage and hepatocarcinogenesis later in life. Oncogene.
24:367–380. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jeon SH, Park JH and Chang SG: Expression
of antioxidant enzymes (Catalase, superoxide dismutase, and
glutathione peroxidase) in human bladder cancer. Korean J Urol.
48:921–926. 2007. View Article : Google Scholar
|
|
17
|
Kasapović J, Pejić S, Todorović A,
Stojiljković V and Pajović SB: Antioxidant status and lipid
peroxidation in the blood of breast cancer patients of different
ages. Cell Biochem Funct. 26:723–730. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sharma A, Tripathi M, Satyam A and Kumar
L: Study of antioxidant levels in patients with multiple myeloma.
Leuk Lymphoma. 50:809–815. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chung-man Ho J, Zheng S, Comhair SA,
Farver C and Erzurum SC: Differential expression of manganese
superoxide dismutase and catalase in lung cancer. Cancer Res.
61:8578–8585. 2001.PubMed/NCBI
|
|
20
|
Khan MA, Chen HC, Wan XX, Tania M, Xu AH,
Chen FZ and Zhang DZ: Regulatory effects of resveratrol on
antioxidant enzymes: A mechanism of growth inhibition and apoptosis
induction in cancer cells. Mol Cells. 35:219–225. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xingyuan Q, Lijing G, Weijie C, Mengyi S,
Bing L, et al: Optimized detection methods of fatty acids in the
Bohai seaweed by response surface method. Feed Res. 1:65–70.
2015.
|
|
22
|
Ge M, Chi X, Zhang A, Luo G, Sun G, Xie H
and Hei Z: Intestinal NF-E2-related factor-2 expression and
antioxidant activity changes in rats undergoing orthotopic liver
autotransplantation. Oncol Lett. 6:1307–1312. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yagi K: Simple assay for the level of
total lipid peroxides in serum or plasma. Methods Mol Biol.
108:101–106. 1998.PubMed/NCBI
|
|
24
|
Gutiérrez-Salinas J, García-Ortíz L,
Morales González JA, Hernández-Rodríguez S, Ramírez-García S,
Núñez-Ramos NR and Madrigal-Santillán E: In vitro effect of sodium
fluoride on malondialdehyde concentration and on superoxide
dismutase, catalase, and glutathione peroxidase in human
erythrocytes. ScientificWorldJournal. 2013:8647182013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Johnson K, Bruder ED and Raff H:
Adrenocortical control in the neonatal rat: ACTH- and
cAMP-independent corticosterone production during hypoxia. Physiol
Rep. 1:e000542013. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thumova M, Pech V, Froehlich O, Agazatian
D, Wang X, Verlander JW, Kim YH and Wall SM: Pendrin protein
abundance in the kidney is regulated by nitric oxide and cAMP. Am J
Physiol Renal Physiol. 303:F812–F820. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Geng L and Li Q: Expression and function
of heregulin-alpha and its receptors in mammary gland of mouse. Sci
China Life Sci. 53:1015–1024. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Geng L, Zhou W, Qu X, Chen W, Li Y, Liu C,
Sun J, Yu X, Wang H, Zhang Z, et al: Optimization of the
preparation of pectin from aloe using a box-behnken design.
Carbohydrate Polymers. 105:193–199. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ghoneum MH, Badr El-Din NK, Abdel Fattah
SM, Pan D and Tolentino L: Hydroferrate fluid, MRN-100, provides
protection against chemical-induced gastric and esophageal cancer
in Wistar rats. Int J Biol Sci. 11:295–303. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Deepalakshmi K, Mirunalini S, Krishnaveni
M and Arulmozhi V: In vitro and in vivo antioxidant potentials of
an ethanolic extract of Ganodermalucidum in rat mammary
carcinogenesis. Chin J Nat Med. 11:621–627. 2013.PubMed/NCBI
|
|
31
|
Periyasamy K, Baskaran K, Ilakkia A,
Vanitha K, Selvaraj S and Sakthisekaran D: Antitumor efficacy of
tangeretin by targeting the oxidative stress mediated on
7,12-dimethylbenz (a) anthracene-induced proliferative breast
cancer in Sprague-Dawley rats. Cancer Chemother Pharmacol.
75:263–272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Giorgio M, Trinei M, Migliaccio E and
Pelicci PG: Hydrogen peroxide: A metabolic by-product or a common
mediator of ageing signals? Nat Rev Mol Cell Biol. 8:722–728. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kowaltowski AJ, Castilho RF and Vercesi
AE: Mitochondrial permeability transition and oxidative stress.
FEBS Lett. 495:12–15. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Martinez-Useros J and Garcia-Foncillas J:
Obesity and colorectal cancer: Molecular features of adipose
tissue. J Transl Med. 14:212016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Piazza GA, Thompson WJ, Pamukcu R, Alila
HW, Whitehead CM, et al: Exisulind, a novel proapoptotic drug,
inhibits rat urinary bladder tumorigenesis. Cancer Res.
2001.61:3961–3968. PubMed/NCBI
|
|
36
|
Kool M, de Haas M, Scheffer GL, Scheper
RJ, van Eijk MJ, et al: Analysis of expression of cMOAT (MRP2),
MRP3, MRP4, and MRP5, homologues of the multidrug
resistance-associated protein gene (MRP1), in human cancer cell
lines. Cancer Res. 1997.57:3537–3547. PubMed/NCBI
|
|
37
|
Drees M, Zimmermann R and Eisenbrand G.:
3′,5′-Cyclic nucleotide phosphodiesterase in tumor cells as
potential target for tumor growth inhibition. Cancer Res.
1993.53:3058–3061. PubMed/NCBI
|
|
38
|
Ciardiello F, Pepe S, Bianco C,
Baldassarre G, Ruggiero A, et al: Down-regulation of RI alpha
subunit of cAMP-dependent protein kinase induces growth inhibition
of human mammary epithelial cells transformed by c-Ha-ras and
c-erbB-2 proto-oncogenes. Int. J. Cancer. 1993.53:438–443.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Singer AL, Sherwin RP, Dunn AS and
Appleman M.M.: Cyclic nucleotide phosphodiesterases in neoplastic
and nonneoplastic human mammary tissues. Cancer Res. 1976.36:60–66.
PubMed/NCBI
|
|
40
|
Cohen LA, Straka D and Chan PC: Cyclic
nucleotide phosphodiesterase activity in normal and neoplastic rat
mammary cells grown in monolayer culture. Cancer Res.
1976.36:2007–2012. PubMed/NCBI
|
|
41
|
Tinsley H.N..Gary B.D..Keeton A.B..Zhang
W..Abadi A.H..Reynolds R.C..Piazza G.A.: Sulindac sulfide
selectively inhibits growth and induces apoptosis of human breast
tumor cells by phosphodiesterase 5 inhibition, elevation of cyclic
GMP, and activation of protein kinase G. Mol. Cancer Ther.
2009.8:3331–3340. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Deguchi A and Thompson WJ: Weinstein IB
Activation of protein kinase G is sufficient to induce apoptosis
and inhibit cell migration in colon cancer cells. Cancer Res.
2004.64:3966–3973. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Carlson CC, Smithers SL, Yeh KA, Burnham
LL and Dransfield DT: Protein kinase A regulatory subunits in colon
cancer. Neoplasia. 1999.1:373–378. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tinsley H.N..Gary B.D..Keeton A.B..Zhang
W..Abadi A.H..Reynolds R.C..Piazza G.A.: Sulindac sulfide
selectively inhibits growth and induces apoptosis of human breast
tumor cells by phosphodiesterase 5 inhibition, elevation of cyclic
GMP, and activation of protein kinase G. Mol. Cancer Ther.
2009.8:3331–3340. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
DeRubertis FR and Craven PA: Sequential
alterations in the hepatic content and metabolism of cyclic AMP and
cyclic GMP induced by DL-ethionine: Evidence for malignant
transformation of liver with a sustained increase in cyclic AMP.
Metabolism. 1976.25:1611–1625. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lerner A and Epstein PM: Cyclic nucleotide
phosphodiesterases as targets for treatment of haematological
malignancies. Biochem. J. 2006.393:21–41. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang L, Murray F, Zahno A, Kanter JR,
Chou D, et al: Cyclic nucleotide phosphodiesterase profiling
reveals increased expression of phosphodiesterase 7B in chronic
lymphocytic leukemia. Proc. Natl. Acad. Sci. USA. 2008; 105:pp.
19532–19537. View Article : Google Scholar
|
|
48
|
Marko D, Pahlke G, Merz KH and Eisenbrand
G: Cyclic 3′,5′-nucleotide phosphodiesterases: Potential targets
for anticancer therapy. Chem. Res. Toxicol. 2000.13:944–948.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Aleksijevic A, Lang JM, Giron C, Stoclet
JC, Mayer S, et al: Alterations of peripheral blood lymphocyte
cyclic AMP and cyclic GMP in untreated patients with hodgkin's
disease. Clin. Immunol. Immunopathol. 1983.26:398–405. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Aleksijevic A, Lugnier C, Giron C, Mayer
S, Stoclet JC, et al: Cyclic AMP and cyclic GMP phosphodiesterase
activities in Hodgkin's disease lymphocytes. Int. J.
Immunopharmacol. 1987.9:525–531. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Heinonen PK and Metsa-Ketela T:
Prostanoids and cyclic nucleotides in malignant and benign ovarian
tumors. Med. Oncol. Tumor Pharmacother. 1988.5:11–15. PubMed/NCBI
|
|
52
|
Pertuit M, Barlier A, Enjalbert A and
Gérard C: Signalling pathway alterations in pituitary adenomas:
Involvement of Gsalpha, cAMP and mitogen-activated protein kinases.
J. Neuroendocrinol. 2009.21:869–877. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Michalides R..Griekspoor A..Balkenende
A..Verwoerd D..Janssen L..Jalink K..Floore A..Velds A..van't Veer
L..Neefjes J.: Tamoxifen resistance by a conformational arrest of
the estrogen receptor alpha after PKA activation in breast cancer.
Cancer Cell. 2004.5:597–605. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Shacter E and Weitzman SA: Chronic
inflammation and cancer. Oncology (Williston Park). 16:217–26,
229–232. 2002.PubMed/NCBI
|
|
55
|
Khansari N, Shakiba Y and Mahmoudi M:
Chronic inflammation and oxidative stress as a major cause of
age-related diseases and cancer. Recent Pat Inflamm Allergy Drug
Discov. 3:73–80. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Toyoshi I, Noriyuki S and Yasutaka M:
Bilirubin as an important physiological modulator of oxidative
stress and chronic inflammation in metabolic syndrome and diabetes:
A new aspect on old molecule. Diabetology International. 7:338–341.
2016. View Article : Google Scholar
|
|
57
|
Ahmed A, Redmond HP and Wang JH: Links
between Toll-like receptor 4 and breast cancer. Oncoimmunology.
2:e229452013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ahmed A, Wang JH and Redmond HP: Silencing
of TLR4 increases tumor progression and lung metastasis in a murine
model of breast cancer. Ann Surg Oncol. 20 Suppl 3:S389–S396. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Mai CW, Kang YB and Pichika MR: Should a
Toll-like receptor 4 (TLR-4) agonist or antagonist be designed to
treat cancer? TLR-4: Its expression and effects in the ten most
common cancers. Onco Targets Ther. 6:1573–1587. 2013.PubMed/NCBI
|
|
60
|
Ghochikyan A, Pichugin A, Bagaev A,
Davtyan A, Hovakimyan A, Tukhvatulin A, Davtyan H, Shcheblyakov D,
Logunov D, Chulkina M, et al: Targeting TLR-4 with a novel
pharmaceutical grade plant derived agonist, Immunomax®,
as a therapeutic strategy for metastatic breast cancer. J Transl
Med. 12:3222014. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yang H, Wang B, Wang T, Xu L, He C, Wen H,
Yan J, Su H and Zhu X: Toll-like receptor 4 prompts human breast
cancer cells invasiveness via lipopolysaccharide stimulation and is
overexpressed in patients with lymph node metastasis. PLoS One.
9:e1099802014. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Delerive P, De Bosscher K, Besnard S,
VandenBerghe W, Peters JM, Gonzalez FJ, Fruchart JC, Tedgui A,
Haegeman G and Staels B: Peroxisome proliferator-activated receptor
a negatively regulates the vascular inflammatory gene response by
negative cross-talk with transcription factors NF-kappaB and AP-1.
J Biol Chem. 274:32048–32054. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Peters JM, Shah YM and Gonzalez FJ: The
role of peroxisome proliferator-activated receptors in
carcinogenesis and chemoprevention. Nat Rev Cancer. 12:181–195.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Takeda S, Ikeda E, Su S, Harada M, Okazaki
H, Yoshioka Y, Nishimura H, Ishii H, Kakizoe K, Taniguchi A, et al:
Δ (9)-THC modulation of fatty acid 2-hydroxylase (FA2H) gene
expression: Possible involvement of induced levels of PPARα in
MDA-MB-231 breast cancer cells. Toxicology. 326:18–24. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Schoonjans K, Staels B and Auwerx J: The
peroxisome proliferator activated receptors (PPARS) and their
effects on lipid metabolism and adipocyte differentiation. Biochim
Biophys Acta. 1302:93–109. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Desvergne B and Wahli W: Peroxisome
proliferator-activated receptors: Nuclear control of metabolism.
Endocr Rev. 20:649–688. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Shigeto T, Yokoyama Y, Xin B and Mizunuma
H: Peroxisome proliferator-activated receptor alpha and gamma
ligands inhibit the growth of human ovarian cancer. Oncol Rep.
18:833–840. 2007.PubMed/NCBI
|
|
68
|
Drukala J, Urbanska K, Wilk A, Grabacka M,
Wybieralska E, Del Valle L, Madeja Z and Reiss K: ROS accumulation
and IGF-IR inhibition contribute to fenofibrate/PPARalpha-mediated
inhibition of glioma cell motility in vitro. Mol Cancer. 9:1592010.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Grabacka M, Plonka PM, Urbanska K and
Reiss K: Peroxisome proliferator-activated receptor alpha
activation decreases metastatic potential of melanoma cells in
vitro via down-regulation of Akt. Clin Cancer Res. 12:3028–3036.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhou J, Zhang S, Xue J, Avery J, Wu J,
Lind SE and Ding WQ: Activation of peroxisome
proliferator-activated receptor α (PPARα) suppresses
hypoxia-inducible factor-1α (HIF-1α) signaling in cancer cells. J
Biol Chem. 287:35161–35169. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Yessoufou A1, Atègbo JM, Attakpa E,
Hichami A, Moutairou K, Dramane KL and Khan NA: Peroxisome
proliferator-activated receptor-alpha modulates insulin gene
transcription factors and inflammation in adipose tissues in mice.
Mol Cell Biochem. 323:101–111. 2009. View Article : Google Scholar : PubMed/NCBI
|