Introduction
Ovarian cancer, the second-leading cause of
gynecological malignancy, is a major cause of cancer-associated
mortality among women (1). Although
patients usually exhibit an acceptable initial response to
first-line therapy (cytoreductive surgery and combined
platinum-paclitaxel chemotherapy), the majority of cancer cases
recur, are invasive and result in patient mortality (2,3). The
underlying mechanism responsible for invasion and metastasis in
epithelial ovarian cancer remains unclear (4). Novel therapeutic agents are therefore
required to improve patient survival.
The epithelial-mesenchymal transition (EMT), which
causes epithelial cells to become similar mesenchymal cells with
increased motility, serves a notable role in embryonic development
(5). A notable hallmark of EMT is the
reduced or lack of expression of the epithelial marker E-cadherin
and induction of expression of the mesenchymal marker N-cadherin
(6), which promotes metastatic
disease (7). EMT is involved in
cancer progression, and represents the main event that promotes
tumor invasion and metastasis (8,9).
Therefore, targeting EMT may aid the development of potentially
effective therapies.
Hedgehog (Hh) signaling serves a critical role in
embryonic development and its abnormal induction is involved in
tumorigenesis. Aberrant Hh activation has been demonstrated in
breast, colon, and prostate cancer (10–12), and
is closely associated with self-renewal, invasion and migration of
cancer stem cells, promoting cancer progression (13,14). It
was reported that Hh signaling might induce EMT in pancreatic
cancer (9,15). In addition, Yue et al (16) reported that Hh also promoted EMT
progression in lung squamous cell carcinoma. Notably,
glioma-associated oncogene-1 (Gli-1), a major Hh signaling
effector, decreases levels of E-cadherin and increases those of
N-cadherin (17,18), indicating that Hh signaling is
important for EMT occurrence. Cyclopamine, a smoothened (Smo) Hh
signaling pathway inhibitor, is considered to be an anticancer
molecule; however, its clinical application is markedly hampered by
elevated costs and its instability in aqueous or polar solvents
(19). Therefore, developing
additional anticancer drugs remains critical.
Fructus viticis, the fruit of Vitex trifolia
L. (Verbenaceae), has been used in traditional Chinese medicine for
its anti-inflammatory effects for over a thousand years (20,21).
Casticin, a biologically active component of Fructus viticis
(20), possesses numerous biological
and pharmacological effects, including immunomodulatory,
anti-hyperprolactinemia, neuroprotective and anti-inflammatory
properties (22–25), and alters tumorigenesis and tumor
progression (26,27). Additionally, it has been demonstrated
that casticin suppresses EMT in hepatocellular carcinoma (28). However, studies assessing whether
casticin can suppress the EMT via Hh signaling in ovarian carcinoma
are scarce. Western blot analysis and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) were
used to evaluate the effect of casticin on Hh signaling, which
modulates EMT and cell migration in ovarian carcinoma.
Materials and methods
Cell culture and reagents
Human ovarian cancer SKOV3 cells were obtained from
the Cell Bank of Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China), and cultured in Dulbecco's modified
Eagle's medium (Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin, and 100 U/ml streptomycin
(Hyclone; GE Healthcare Life Sciences, Logan, UT, USA), in a
humidified atmosphere containing 5% CO2 at 37°C.
Casticin (purity ≥98%) was manufactured by Chengdu Biopurify
Phytochemicals Ltd. (Chengdu, China) as yellow crystals (molecular
weight, 374.3 Da). Cyclopamine was manufactured by Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). Rabbit anti-Gli-1 antibody
(1:1,000; cat. no. 2534) and Twist-related protein 1 (Twist1)
antibodies (1:1,000; cat. no. 46702) (both from Cell Signaling
Technology, Inc., Danvers, MA, USA), and rat anti-E-cadherin
(1:1,000; cat. no. SAB4503751) and mouse monoclonal anti-N-cadherin
(1:1,000; cat. no. C2542) (both from Sigma-Aldrich; Merck KGaA)
antibodies were used in the present study. Goat anti-rabbit lgG,
horseradish peroxidase (HRP)-linked antibody (cat. no. 7074;
1:1,000 dilution) and goat anti-mouse lgG, HRP-linked antibody
(cat. no. 7076; 1:1,000 dilution) were both purchased from Cell
Signaling Technology, Inc.
Determination of cell proliferation
and growth
Cell proliferation and growth were assessed by MTT
and Soft agar assays. For MTT assay, SKOV3 cells (10,000/well) were
seeded in 96-well plates and incubated for 24 h prior to treatment
with increasing casticin (0.0, 1.0, 2.0, 4.0 and 8.0 µM) or
cyclopamine amounts (0.0, 12.5, 25.0, 50.0 and 100.0 µM) for 48 h.
Subsequently, 20 µl MTT (5 mg/ml) was added for further 4 h.
Dimethyl sulfoxide was used to dissolve the resulting formazan
crystals; absorbance was read at 490 nm on Bio-Rad 550 (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The half-maximal inhibitory
concentration (IC50; causing 50% cell growth inhibition)
values were determined. Cell growth inhibition rate was derived as:
[(A490 control cells - A490 treated
cells)/A490 control cells] × 100.
Soft agar assay was performed using SKOV3 cells
(10,000/well) treated with or without casticin (2.0 µM) or
cyclopamine (25.0 µM) for 48 h and then cells were plated in 0.35%
agarose overlying a 0.6% agar layer. Cells were incubated at 37°C
for 27 days; at the end of the experiment, colonies were counted
under an inverted light microscope at a ×4 magnification (Olympus
Corportation, Tokyo, Japan).
Migration assay
When SKOV3 cells reached an optimal confluence
(80–90%) in a 6-well culture plate, the monolayer was gently and
slowly scratched with a clean 100 µl pipette tip across the center
of the plate. Following two washes with PBS, casticin (2.0 µM) or
cyclopamine (25.0 µM) were added at IC50 values. The
artificial wounds were imaged at 0 and 24 h, respectively, under an
inverted light microscope at a ×10 magnification (CKX41; Olympus
Corporation, Tokyo, Japan).
Transwell invasion assay
For the invasion assay, 1×105 SKOV3 cells
treated with casticin or cyclopamine were seeded in the top chamber
onto the Matrigel-coated membrane (24-well insert; pore size, 8 µm;
Corning Life Sciences, Corning, NY, USA). Cells were plated in DMEM
medium without serum or growth factors in the top chamber, and DMEM
medium supplemented with 10% FBS was used in the lower chamber. The
cells were incubated for 48 h and then cells that invaded via the
pores were removed with a cotton swab. Cells on the lower surface
of the membrane were fixed with 4% paraformaldehyde for 10 min at
25°C and stained with 10% Giemsa for 30 min at room temperature.
The number of cells invading via the membrane was counted under a
light microscope (magnification, ×20; three random fields per
well).
RT-qPCR
SKOV3 Cells were treated with casticin (2.0 µM) or
cyclopamine (25.0 µM) for 0, 3, 6, 12 and 24 h, then RNA was
extracted and cDNA prepared as previously reported (29). RT-qPCR was conducted with 1 µl cDNA on
MyiQ real-time PCR Instrument (Bio-Rad Laboratories, Inc.) using iQ
SYBR Green PCR Supermix (Bio-Rad Laboratories, Inc.), following the
manufacturer's instructions. The thermocycling conditions were as
follows: 95°C for 3 min, 95°C for 15 sec, and 60°C for 20 sec, for
40 cycles. Relative gene expression levels were determined by the
2−ΔΔCq method (30) with
GAPDH as an internal reference. The following primers were used:
Gli-1 forward, 5′-TTCCTACCAGAGTCCCAAGT-3′ and reverse,
5′-CCCTATGTGAAGCCCTATTT-3′; Twist1 forward,
5-CGGACAAGCTGAGCAAGATT-3′ and reverse, 5′-CCTTCTCTGGAAACAATGAC-3′;
N-cadherin forward, 5′-CGTGAAGGTTTGCCAGTGTG-3′ and reverse,
5′-GCGTTCTTTATCCCGCCGTT-3′; E-cadherin forward,
5′-CCTCAGGTCATAAACATCATTG-3′ and reverse,
5′-CGCCTCCTTCTTCATCATAGTAA-3′; GAPDH forward,
5′-GGTGGTCTCCTCTGACTTCAACA-3′ and reverse,
5′-GTTGCTGTAGCCAAATTCGTTGT-3′.
Western blotting
SKOV3 cells (1×106) treated with casticin
or cyclopamine (at IC50) for 48 h were lysed and
analyzed by western blot, as previously reported (31), using the aforementioned antibodies;
β-actin was employed as an internal control. Protein bands were
visualized by enhanced chemiluminescence kit (GE Healthcare,
Chicago, IL, USA), and visualized using Tanon-5500
chemiluminescence instrument (Tanon Inc., Shanghai, China). Data
were quantified by ImageJ 1.49v software (National Institutes of
Health, Bethesda, MD, USA).
Statistical analyses
Experiments were repeated ≥3 times, with data
expressed as the mean ± standard deviation. All data were analyzed
by SPSS 16.0 for Windows (SPSS Inc., Chicago, IL, USA). One-way
analysis of variance was used to perform group comparisons, with a
least significant difference test for post hoc multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
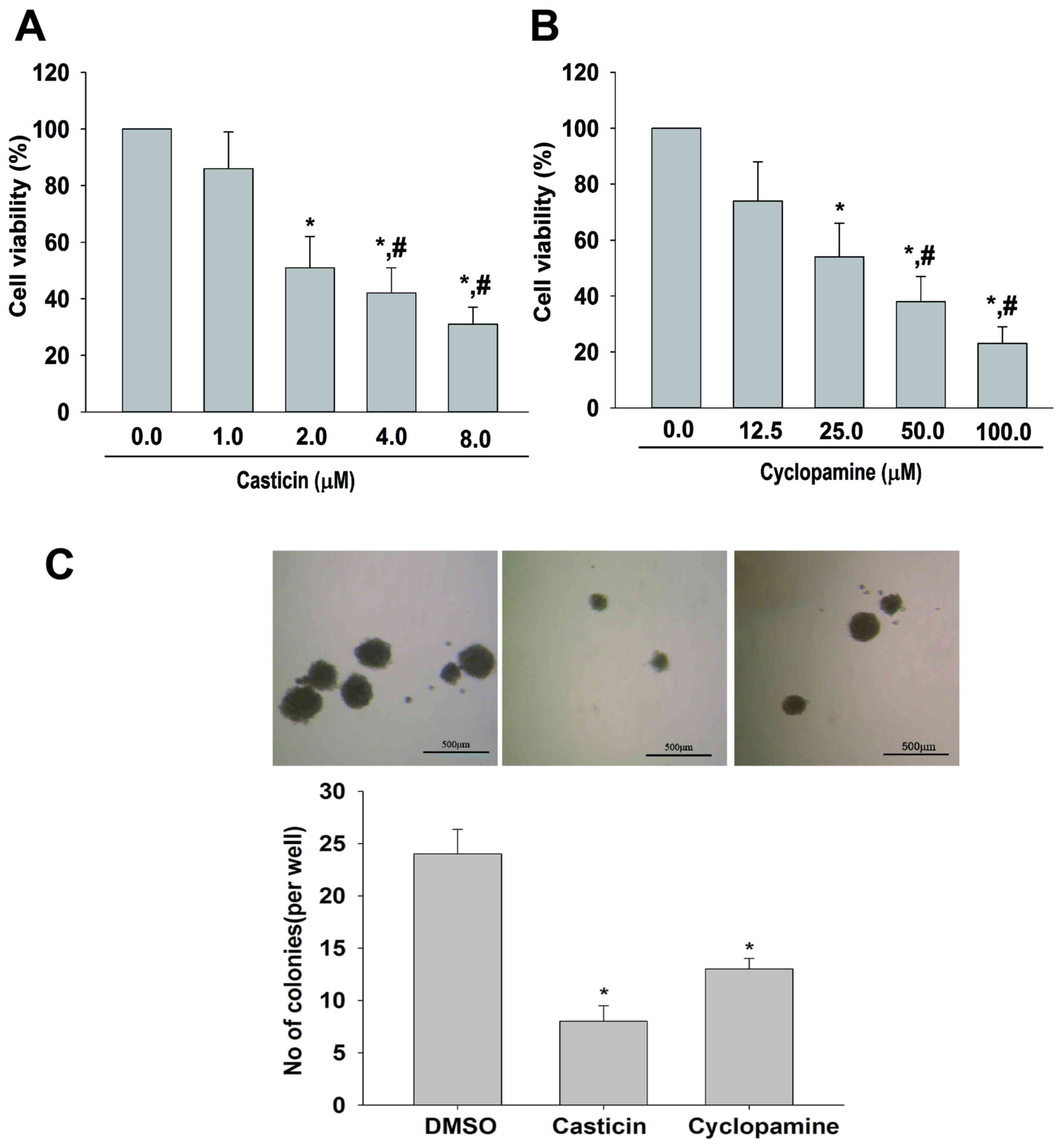
Casticin and cyclopamine inhibit SKOV3
cell proliferation and colony formation
The effects of casticin and cyclopamine on SKOV3
cell viability were determined by the MTT and soft agar assay. The
MTT assay revealed that casticin (Fig.
1A) or cyclopamine (Fig. 1B)
inhibited SKOV3 cell growth in a dose-dependent manner.
IC50 values of casticin and cyclopamine were 2.18±0.37
and 27.61±2.16 µM, respectively. Thus, 2.0 µM casticin and 25 µM
cyclopamine were used to perform the soft agar assay. As presented
in Fig. 1C, the SKOV3 cells treated
with casticin or cyclopamine formed ~100% fewer colonies than the
control.
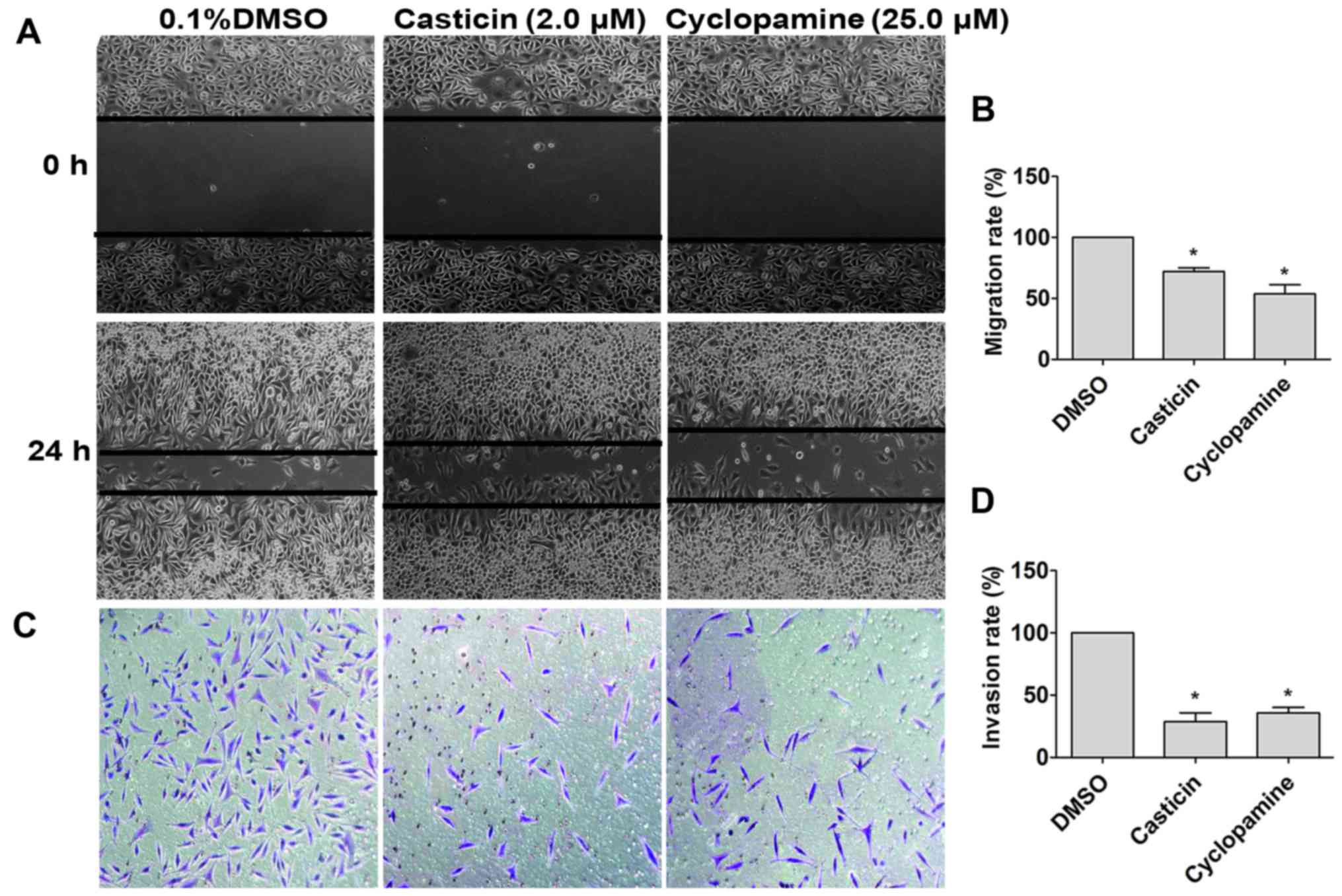
Casticin and cyclopamine inhibit SKOV3
cell migration and invasion
Wound-healing assays were performed to assess the
effect of casticin and cyclopamine on the migration of SKOV3 cells.
As presented in Fig. 2, treatment
with casticin (2.0 µM) or cyclopamine (25.0 µM) resulted in wider
wounds compared with the control group after 24 h of incubation.
Subsequently, quantification was conducted, which revealed that the
migration rate of the casticin (72%±1.73) and cyclopamine groups
(53.7%±4.37) were significantly lower compared with the control
(100%; P<0.05). To analyze whether casticin (2.0 µM) or
cyclopamine (25.0 µM) affected cell invasion further, a transwell
invasion assay was performed. The results demonstrated that there
was a marked difference in the invasion rate of cells that
underwent treatment with casticin (2.0 µM) or cyclopamine (25.0 µM;
28.8±7.1 or 35.8±4.4, respectively) compared with the control group
(100%; data not shown). These findings indicated that casticin and
cyclopamine inhibited the migration and invasion of the human
ovarian cancer SKOV3 cell line.
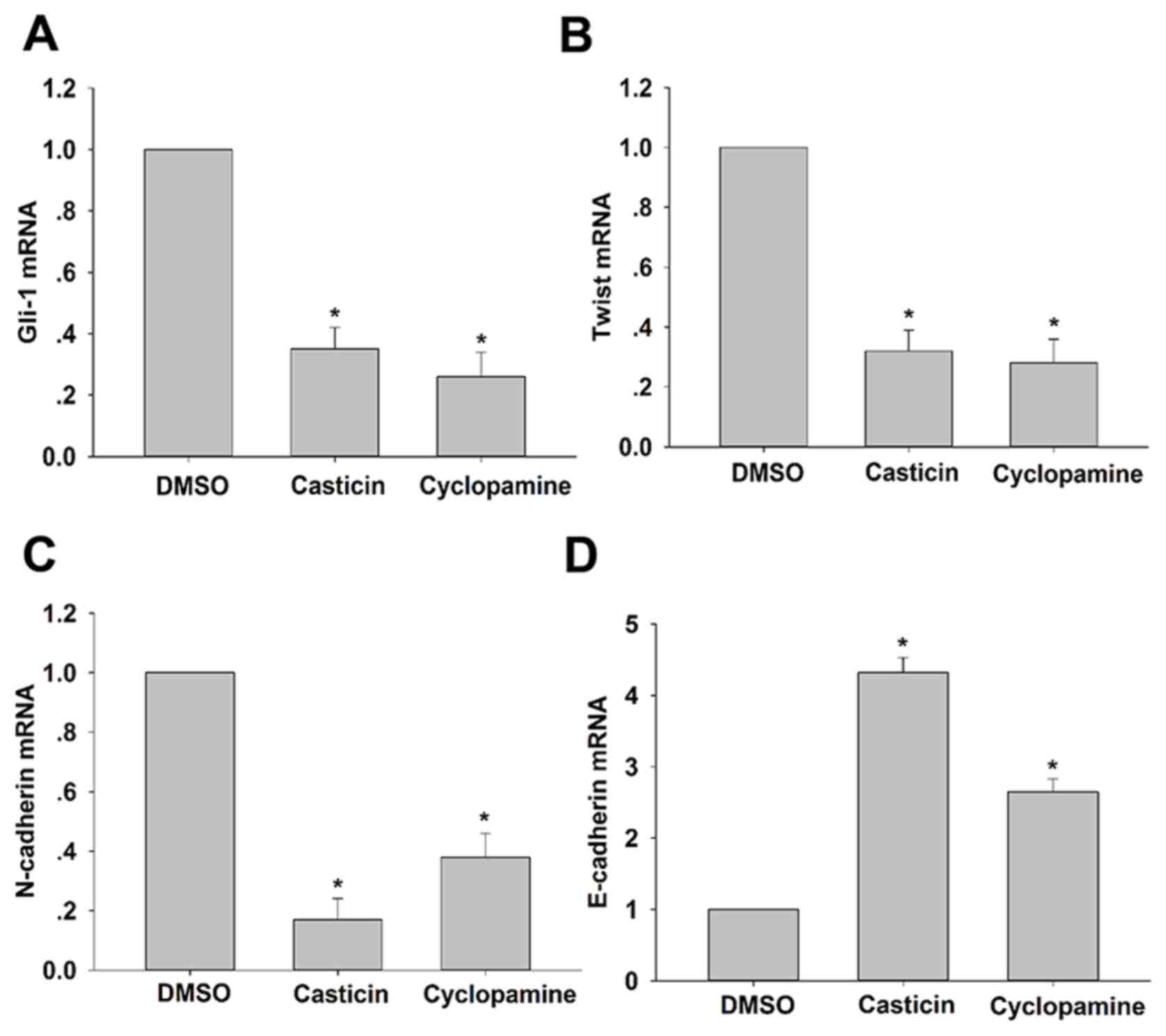
Casticin and cyclopamine alter Gli-1,
Twist1, E-cadherin and N-cadherin gene expression levels in SKOV3
cells
To assess whether casticin and cyclopamine affect
the mRNA expression of effectors of Hh signaling and EMT markers,
SKOV3 cells were treated for 24 h with complete medium containing
2.0 µM casticin and 25.0 µM cyclopamine, respectively, and were
analyzed by RT-qPCR. Compared with the control group, the mRNA
expression levels of Twist1 and N-cadherin were markedly decreased
(~32 and ~17%, respectively) in the casticin (2.0 µM) groups, and
decreased by ~30 and ~38% in the cyclopamine (25.0 µM) groups
(Fig. 3A-C). However, the mRNA
expression levels of E-cadherin were increased ~4-fold for
casticin-treated cells and ~2-fold for cyclopamine-treated cells
(Fig. 3D). In addition, the mRNA
expression levels of the Hh signaling effector, Gli-1, were also
decreased by 35 and 26% in the casticin (2.0 µM) and cyclopamine
(25.0 µM) groups, respectively. These results indicated the
inhibitory effects of casticin and cyclopamine on Hh signaling and
the EMT process.
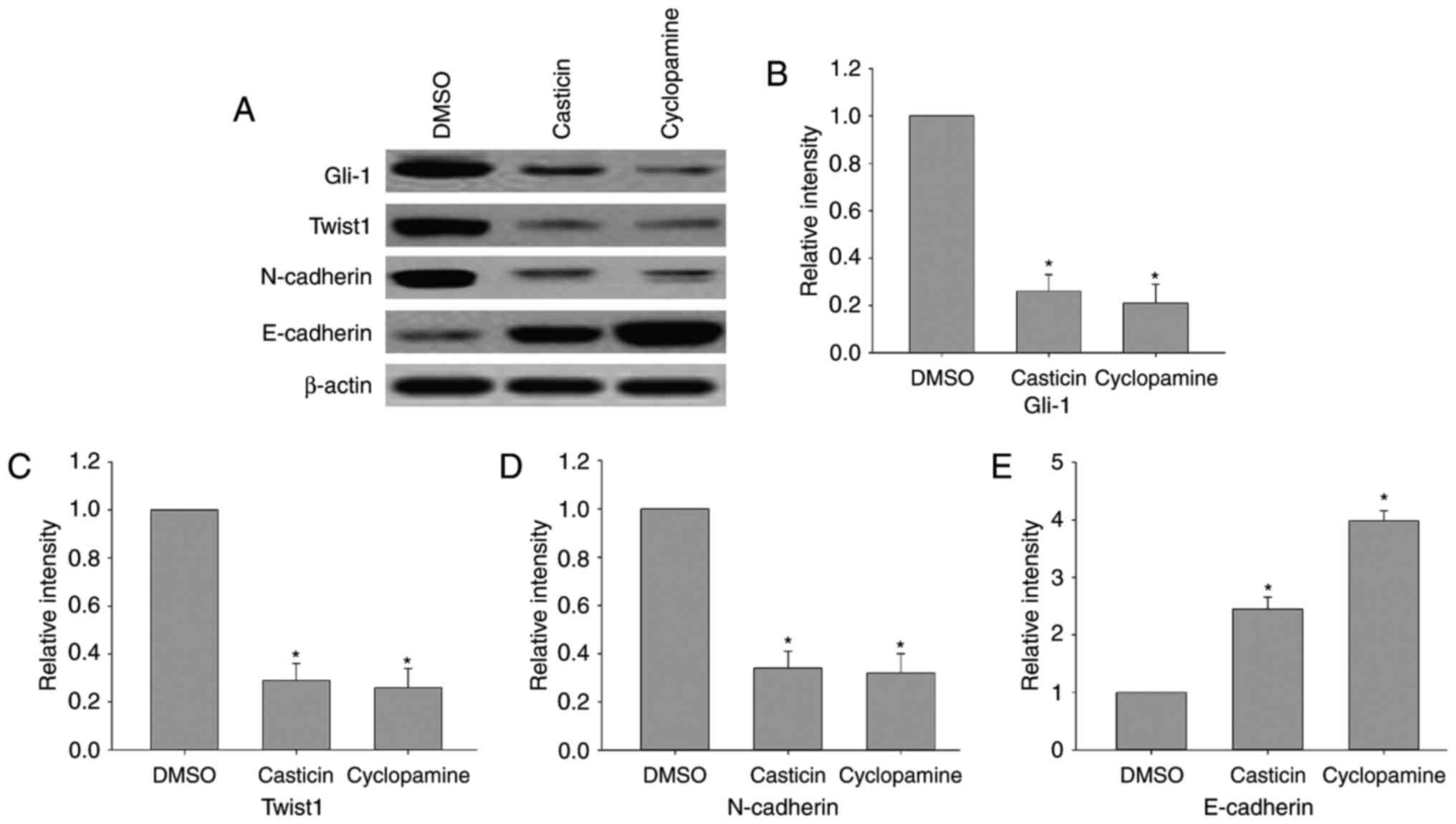
Effects of casticin and cyclopamine on
protein expression levels of Hh signaling effectors and EMT markers
in SKOV3 cells
Whether casticin and cyclopamine affect Hh signaling
pathway effectors and EMT markers was investigated at the protein
level. After 24 h of treatment, E-cadherin protein expression
levels were markedly increased in the casticin (2.0 µM) and
cyclopamine (25.0 µM) groups compared with in the control; however,
the protein expression levels of Gli-1, Twist1 and N-cadherin were
markedly reduced by casticin (2.0 µM) and cyclopamine (25.0 µM)
treatments, respectively, in comparison with the control group
(Fig. 4A). These findings indicated
that casticin and cyclopamine suppressed Hh signaling and the EMT
process.
Time-dependent effect of casticin
treatment on mRNA expression levels of Gli-1 and EMT markers
To determine the time course of casticin activity on
mRNA expression, SKOV3 cells were exposed to 2.0 µM casticin at 0,
3, 6, 12, and 24 h, and Gli-1, Twist1, E-cadherin, and N-cadherin
mRNA expression levels were quantitated by RT-qPCR. Gli-1, Twist1
and N-cadherin mRNA expression levels were decreased as time
progressed (Fig. 5A-C), with
significant alterations in expression levels detected at 24 h,
which were 19, 14 and 16% of the expression at 0 h, respectively.
E-cadherin levels increased (Fig. 5D)
as incubation time increased.
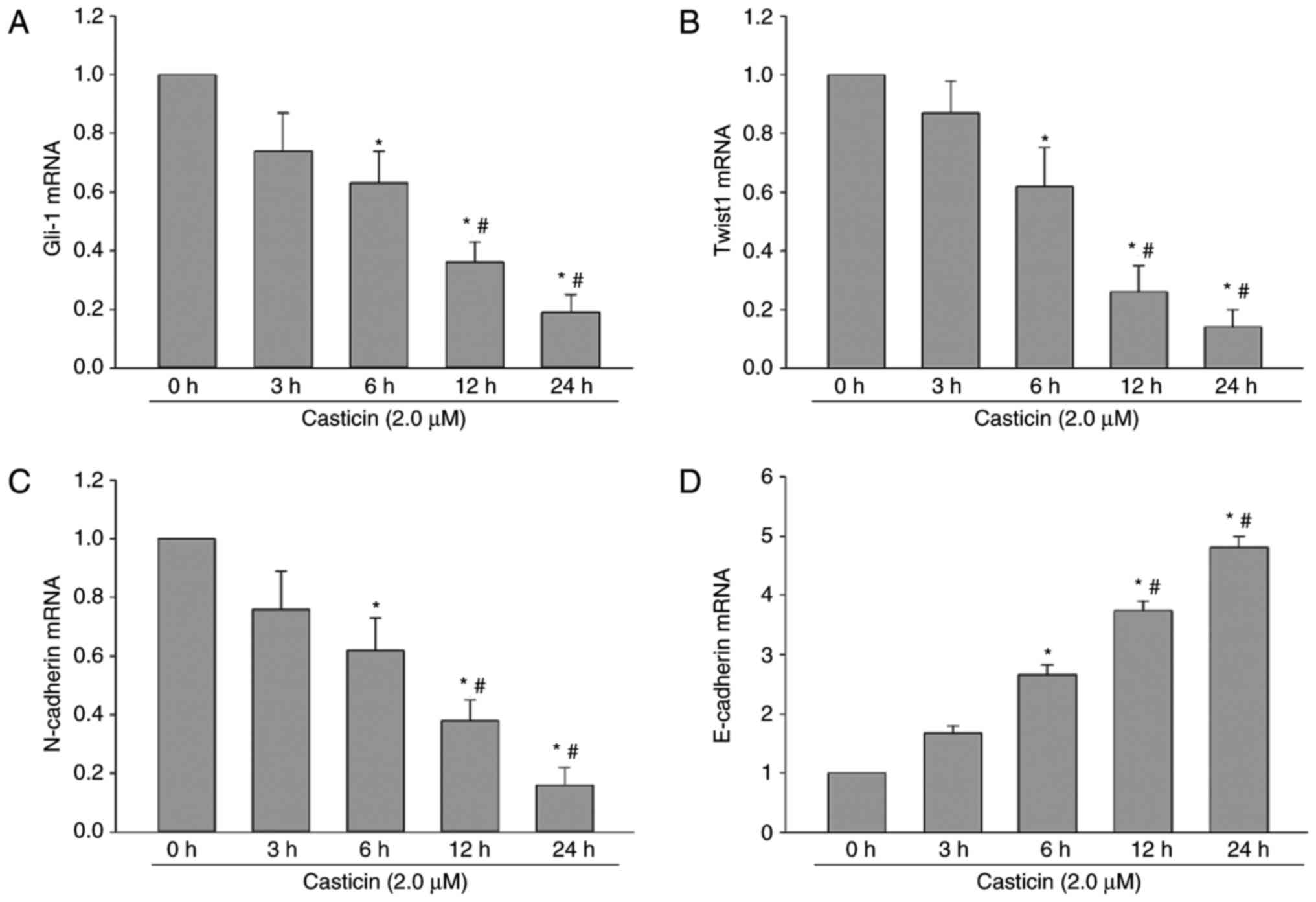 | Figure 5.Time-dependent effect of casticin
treatment on Gli-1, Twist1, N-cadherin and E-cadherin mRNA
expression levels. SKOV3 cells were cultured with casticin (2.0 µM)
for 0, 3, 6, 12 and 24 h, and analyzed by reverse
transcription-quantitative polymerase chain reaction for relative
mRNA expression levels of (A) Gli-1, (B) Twist1, (C) N-cadherin and
(D) E-cadherin. GAPDH was used for normalization. *P<0.01 vs.
Gli-1, Twist1, N-cadherin and E-cadherin expression levels at 0 h;
#P<0.01 vs. Gli-1, Twist1, N-cadherin and E-cadherin
at 6 h. Gli-1, glioma-associated oncogene-1; Twist1, Twist related
protein 1. |
Time-dependent effect of casticin on
protein expression levels of EMT markers and Hh signaling
effectors
To evaluate the time course of casticin action on
protein expression, Gli-1, Twist1, E-cadherin and N-cadherin
protein expression levels were determined. The results of the
present study were consistent with those of the RT-qPCR analysis:
Time-dependent increases in E-cadherin protein levels, concomitant
with reduced Gli-1, Twist1 and N-cadherin levels were observed
(Fig. 6). Notably, the alterations
were most pronounced at 24 h, indicating that casticin may inhibit
EMT and Hh signaling to a greater degree over time.
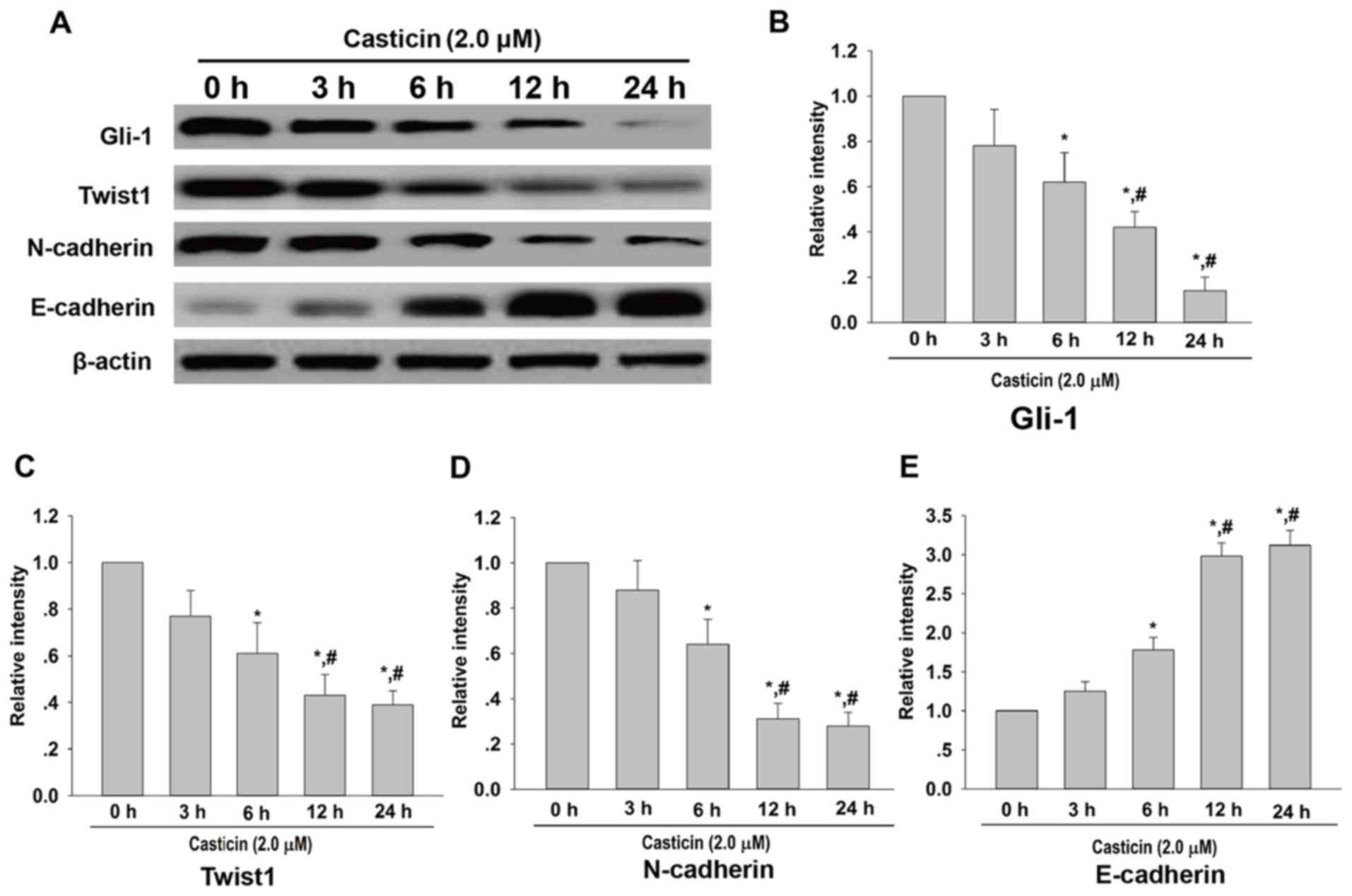 | Figure 6.Time-dependent effect of casticin on
Gli-1, Twist1, N-cadherin and E-cadherin protein expression levels.
SKOV3 cells were cultured with DMEM medium containing casticin (2.0
µM) for 0, 3, 6, 12 and 24 h. (A) Western blotting was used to
analyze the protein expression levels of Gli-1 Twist1, N-cadherin
and E-cadherin, with β-actin used as the internal control. (B-E)
Densitometry analysis of Gli-1, Twist1, N-cadherin and E-cadherin
protein quantitation, respectively. *P<0.01 vs. levels at 0 h;
#P<0.01 vs. levels at 6 h. DMSO, dimethyl sulfoxide;
Gli-1, glioma-associated oncogene-1; Twist1, Twist related protein
1. |
Discussion
Casticin, an anti-inflammatory molecule (32), inhibits numerous cancer cells
(27,28,33).
Previous studies demonstrated that casticin inhibited EMT in liver
cancer stem cells (28) and induced
apoptosis in ovarian cancer cells (29). These findings demonstrated that: i)
SKOV3 cell viability and migration were inhibited by casticin; ii)
casticin suppressed Gli-1 expression; and iii) Twist and N-cadherin
protein and mRNA expression were reduced by casticin; and iv)
casticin treatment resulted in enhanced expression of E-cadherin
mRNA and protein. It should be noted that casticin also inhibits
cell proliferation and EMT in other types of cancer cell, including
hepatocellular carcinoma (28),
indicating its therapeutic potential.
The Hh family includes sonic Hh, desert Hh and
Indian Hh (34). The Hh gene directly
affects embryonic development (35).
Gli-1, a potent inducer of downstream target genes, is itself a
transcriptional target of Hh signaling (36); however, Gli-1 induction prevents
E-cadherin expression in kidney epithelium (37). Additionally, downregulation of Gli-1
induces the invasion and migration of ovarian cancer cells
(38), and Gli-1 expression in
advanced ovarian cancer may be associated with poor survival
(39). It is reported that tumor
progression requires a variety of phenotypical alterations that
decrease intercellular adhesion, with EMT serving a key role
(40). During EMT, epithelial cell
markers are lost, whereas mesenchymal markers, including
N-cadherin, are expressed. The Hh signaling pathway is closely
associated with EMT. It was previously demonstrated that casticin
could inhibit the migration and invasion of hepatocellular
carcinoma cells by suppressing EMT (28). However, reports that demonstrate these
effects in ovarian cancer cells via the Hh signaling pathway are
scarce.
In the present study, cyclopamine, an inhibitor of
the Hh signaling pathway that directly binds to Smo, was used as
positive control. Cyclopamine has been demonstrated to exert
effects on numerous types of tumor, including lung, ovarian, and
breast cancer (41–43). However, poor oral solubility hinders
its clinical development. As aforementioned, casticin may inhibit
EMT in SKOV3 cells via the suppression of Hh pathway activity.
Casticin exhibited a lower IC50 value than
cyclopamine.
The underlying molecular mechanism of casticin in
the Hh signaling pathway requires further investigation. The
findings of the present study revealed that the metastatic
potential of SKOV3 cells incubated with casticin was decreased
compared with those incubated with the control. In addition, Gli-1,
Twist1 and N-cadherin expression levels were also decreased,
whereas those of E-cadherin were increased. This may be due to
Gli-1-mediated activation of Twist1, an EMT-inducing transcription
factor (44); thus, levels of EMT
markers may be affected when Gli-1 expression is reduced. Hence,
further investigation is required to understand the clinical
efficacy of casticin.
The results of the present study revealed that
casticin inhibited ovarian cancer cell migration in vitro by
repressing Hh signaling and EMT. Further in vivo studies are
necessary to improve the understanding of the relevance of Hh
signaling and EMT in ovarian cancer stem-like cells.
Acknowledgements
The present study was funded by a Project of the
National Natural Science Foundation of China (grant nos. 30760248,
31400311 and 81172375), the Project of Scientific Research Fund of
Hunan Provincial Education Department (grant no. 14C0707), the
Project of Hunan Provincial Natural Science Foundation (grant no.
13JJ3061) and the Scientific Research Fund of Hunan Normal
University (grant nos. 140668 and 140666).
References
|
1
|
Hunn J and Rodriguez GC: Ovarian cancer:
Etiology, risk factors, and epidemiology. Clin Obstet Gynecol.
55:3–23. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chari ST, Yadav D, Smyrk TC, DiMagno EP,
Miller LJ, Raimondo M, Clain JE, Norton IA, Pearson RK, Petersen
BT, et al: Study of recurrence after surgical resection of
intraductal papillary mucinous neoplasm of the pancreas.
Gastroenterology. 123:1500–1507. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lim SB, Yu CS, Jang SJ, Kim TW, Kim JH and
Kim JC: Prognostic significance of lymphovascular invasion in
sporadic colorectal cancer. Dis Colon Rectum. 53:377–384. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Radisky DC: Epithelial-mesenchymal
transition. J Cell Sci. 118:4325–4326. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hotz HG, Hotz B and Buhr HJ: Genes
associated with epithelial-mesenchymal transition: Possible
therapeutic targets in ductal pancreatic adenocarcinoma? Anticancer
Agents Med Chem. 11:448–454. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mell LK, Meyer JJ, Tretiakova M, Khramtsov
A, Gong C, Yamada SD, Montag AG and Mundt AJ: Prognostic
significance of E-cadherin protein expression in pathological stage
I–III endometrial cancer. Clin Cancer Res. 10:5546–5553. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun XD, Liu XE and Huang DS: Curcumin
reverses the epithelial-mesenchymal transition of pancreatic cancer
cells by inhibiting the Hedgehog signaling pathway. Oncol Rep.
29:2401–2407. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lei J, Ma J, Ma Q, Li X, Liu H, Xu Q, Duan
W, Sun Q, Xu J, Wu Z and Wu E: Hedgehog signaling regulates hypoxia
induced epithelial to mesenchymal transition and invasion in
pancreatic cancer cells via a ligand-independent manner. Mol
Cancer. 12:662013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brechbiel J, Miller-Moslin K and Adjei AA:
Crosstalk between hedgehog and other signaling pathways as a basis
for combination therapies in cancer. Cancer Treat Rev. 40:750–759.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feldmann G, Fendrich V, McGovern K, Bedja
D, Bisht S, Alvarez H, Koorstra JB, Habbe N, Karikari C, Mullendore
M, et al: An orally bioavailable small-molecule inhibitor of
Hedgehog signaling inhibits tumor initiation and metastasis in
pancreatic cancer. Mol Cancer Ther. 7:2725–2735. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bao C, Namgung H, Lee J, Park HC, Ko J,
Moon H, Ko HW and Lee HJ: Daidzein suppresses tumor necrosis
factor-α induced migration and invasion by inhibiting hedgehog/Gli1
signaling in human breast cancer cells. J Agric Food Chem.
62:3759–3767. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Takebe N, Harris PJ, Warren RQ and Ivy SP:
Targeting cancer stem cells by inhibiting Wnt, Notch, and Hedgehog
pathways. Nat Rev Clin Oncol. 8:97–106. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim EJ, Sahai V, Abel EV, Griffith KA,
Greenson JK, Takebe N, Khan GN, Blau JL, Craig R, Balis UG, et al:
Pilot clinical trial of hedgehog pathway inhibitor GDC-0449
(vismodegib) in combination with gemcitabine in patients with
metastatic pancreatic adenocarcinoma. Clin Cancer Res.
20:5937–5945. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang F, Ma L, Zhang Z, Liu X, Gao H,
Zhuang Y, Yang P, Kornmann M, Tian X and Yang Y: Hedgehog signaling
regulates Epithelial-mesenchymal transition in pancreatic cancer
Stem-like cells. J Cancer. 7:408–417. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yue D, Li H, Che J, Zhang Y, Tseng HH, Jin
JQ, Luh TM, Giroux-Leprieur E, Mo M, Zheng Q, et al: Hedgehog/Gli
promotes epithelial-mesenchymal transition in lung squamous cell
carcinomas. J Exp Clin Cancer Res. 33:342014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kong Y, Peng Y, Liu Y, Xin H, Zhan X and
Tan W: Twist1 and Snail link Hedgehog signaling to tumor-initiating
cell-like properties and acquired chemoresistance independently of
ABC transporters. Stem Cells. 33:1063–1074. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu P, Yang J, Hou Y, Zhang H, Zeng Z, Zhao
L, Yu T, Tang X, Tu G, Cui X and Liu M: LncRNA expression
signatures of twist-induced epithelial-to-mesenchymal transition in
MCF10A cells. Cell Signal. 26:83–93. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang J, Garrossian M, Gardner D,
Garrossian A, Chang YT, Kim YK and Chang CW: Synthesis and
anticancer activity studies of cyclopamine derivatives. Bioorg Med
Chem Lett. 18:1359–1363. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin S, Zhang H, Han T, Wu JZ, Rahman K and
Qin LP: In vivo effect of casticin on acute inflammation. Zhong Xi
Yi Jie He Xue Bao. 5:573–576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee SM, Lee YJ, Kim YC, Kim JS, Kang DG
and Lee HS: Vascular protective role of vitexicarpin isolated from
Vitex rotundifolia in human umbilical vein endothelial cells.
Inflammation. 35:584–593. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mesaik MA, Azizuddin, Murad S, Khan KM,
Tareen RB, Ahmed A, Atta-ur-Rahman and Choudhary MI: Isolation and
immunomodulatory properties of a flavonoid, casticin from Vitex
agnus-castus. Phytother Res. 23:1516–1520. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ye Q, Zhang QY, Zheng CJ, Wang Y and Qin
LP: Casticin, a flavonoid isolated from Vitex rotundifolia,
inhibits prolactin release in vivo and in vitro. Acta Pharmacol
Sin. 31:1564–1568. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ling Y, Zhu J, Fan M, Wu B, Qin L and
Huang C: Metabolism studies of casticin in rats using
HPLC-ESI-MS(n). Biomed Chromatogr. 26:1502–1508. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Velpandian T, Gupta P, Ravi AK, Sharma HP
and Biswas NR: Evaluation of pharmacological activities and
assessment of intraocular penetration of an ayurvedic polyherbal
eye drop (Itone™) in experimental models. BMC Complement Altern
Med. 13:12013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu F, Cao X, Liu Z, Guo H, Ren K, Quan M,
Zhou Y, Xiang H and Cao J: Casticin suppresses self-renewal and
invasion of lung cancer stem-like cells from A549 cells through
down-regulation of pAkt. Acta Biochim Biophys Sin (Shanghai).
46:15–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He G, Cao X, He M, Sheng X, Wu Y and Ai X:
Casticin inhibits self-renewal of liver cancer stem cells from the
MHCC97 cell line. Oncol Lett. 7:2023–2028. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
He M, Cao XC, He GC, Sheng XF, Ai XH and
Wu YH: Casticin inhibits epithelial-mesenchymal transition of liver
cancer stem cells of the SMMC-7721 cell line through downregulating
Twist. Oncol Lett. 7:1625–1631. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang L, Cao XC, Cao JG, Liu F, Quan MF,
Sheng XF and Ren KQ: Casticin induces ovarian cancer cell apoptosis
by repressing FoxM1 through the activation of FOXO3a. Oncol Lett.
5:1605–1610. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ning Y, Li Q, Xiang H, Liu F and Cao J:
Apoptosis induced by 7-difluoromethoxyl-5,4′-di-n-octyl genistein
via the inactivation of FoxM1 in ovarian cancer cells. Oncol Rep.
27:1857–1864. 2012.PubMed/NCBI
|
|
32
|
Liou CJ, Len WB, Wu SJ, Lin CF, Wu XL and
Huang WC: Casticin inhibits COX-2 and iNOS expression via
suppression of NF-kB and MAPK signaling in
lipopolysaccharide-stimulated mouse macrophages. J Ethnopharmacol.
158:310–316. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou Y, Tian L, Long L, Quan M, Liu F and
Cao J: Casticin potentiates TRAIL-induced apoptosis of gastric
cancer cells through endoplasmic reticulum stress. PLoS One.
8:e588552013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Katoh Y and Katoh M: Comparative genomics
on Sonic hedgehog orthologs. Oncol Rep. 14:1087–1090.
2005.PubMed/NCBI
|
|
35
|
Nusslein-Volhard C and Wieschaus E:
Mutations affecting segment number and polarity in Drosophila.
Nature. 287:795–801. 1980. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee J, Platt KA, Censullo P and Ruiz i
Altaba A: Gli1 is a target of Sonic hedgehog that induces ventral
neural tube development. Development. 124:2537–2552.
1997.PubMed/NCBI
|
|
37
|
Li X, Deng W, Nail CD, Bailey SK, Kraus
MH, Ruppert JM and Lobo-Ruppert SM: Snail induction is an early
response to Gli1 that determines the efficiency of epithelial
transformation. Oncogene. 25:609–621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen Q, Xu R, Zeng C, Lu Q, Huang D, Shi
C, Zhang W, Deng L, Yan R, Rao H, et al: Down-regulation of Gli
transcription factor leads to the inhibition of migration and
invasion of ovarian cancer cells via integrin β4-mediated FAK
signaling. PLoS One. 9:e883862014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ciucci A, De Stefano I, Vellone VG, Lisi
L, Bottoni C, Scambia G, Zannoni GF and Gallo D: Expression of the
glioma-associated oncogene homolog 1 (gli1) in advanced serous
ovarian cancer is associated with unfavorable overall survival.
PLoS One. 8:e601452013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen T, You Y, Jiang H and Wang ZZ:
Epithelial-mesenchymal transition (EMT): A biological process in
the development, stem cell differentiation, and tumorigenesis. J
Cell Physiol. 232:3261–3272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yue D, Li H, Che J, Zhang Y, Tseng HH, Jin
JQ, Luh TM, Giroux-Leprieur E, Mo M, Zheng Q, et al: Hedgehog/Gli
promotes epithelial-mesenchymal transition in lung squamous cell
carcinomas. J Exp Clin Canc Res. 33:342014. View Article : Google Scholar
|
|
42
|
Cho H, Lai TC and Kwon GS: Poly(ethylene
glycol)-block-poly(ε-caprolactone) micelles for combination drug
delivery: Evaluation of paclitaxel, cyclopamine and gossypol in
intraperitoneal xenograft models of ovarian cancer. J Control
Release. 166:1–9. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Che J, Zhang FZ, Zhao CQ, Hu XD and Fan
SJ: Cyclopamine is a novel Hedgehog signaling inhibitor with
significant anti-proliferative, anti-invasive and anti-estrogenic
potency in human breast cancer cells. Oncol Lett. 5:1417–1421.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ray A, Meng E, Reed E, Shevde LA and
Rocconi RP: Hedgehog signaling pathway regulates the growth of
ovarian cancer spheroid forming cells. Int J Oncol. 39:797–804.
2011.PubMed/NCBI
|