Introduction
SERPINE2, a 43-kDa protein first found as a
neurite-promoting factor in the culture medium of glial cells
(1), was subsequently proven as a
member of the SERPIN family that has serine protease inhibitory
activity (2,3). It also belongs to the nexin protease
family according to its amino acid sequence, therefore also
commonly called protease nexin-1 (PN-1) (4). SERPINE2 mainly presents in the
extracellular matrix (ECM) and is secreted by many cell types,
including endothelial cells, fibroblasts, macrophages, platelets,
smooth muscle cells, chondrocytes, astrocytes and tumor cells
(5–9).
SERPINE2 binds to its specific target protein to form covalent
complexes, which are endocytosed and degraded by cells (10). Through this suicide pathway, SERPINE2
can regulate ECM by inhibiting and removing proteases in the local
milieu that degrade the ECM by hydrolyzation. As a serine protease
inhibitor, SERPINE2 is known to effectively inhibit thrombin,
urokinase plasminogen activator (uPA), tissue plasminogen activator
(tPA), plasmin, trypsin, XI factor, and prostasin, which is
involved in the regulation of blood coagulation and fibrinolysis
system (3,11,12).
Increasing evidence has shown that a balance between
the extra-cellular serine proteases and their cognate SERPINs plays
a key role in cancer progression, thus selected SERPINs might be
promising biomarkers for tumor diagnosis or prognosis (13). The potential values of SERPINE2 in the
prognosis of several cancers and its co-expression with target
proteases in the tumor microenvironment announce its role in tumor
progression. SERPINE2 is overexpressed in many cancers
including breast cancer (6),
pancreatic cancer, gastric cancer, colorectal cancer (14–16),
thyroid cancer (17), oral carcinoma
(18) and osteosarcoma (19), and contributes to tumor invasion and
metastasis. It has been identified that up-regulation of
SERPINE2 is related to oncogenic activation of RAS,
BRAF and MEK, which leads to the pro-neoplastic actions
of extracellular signal-regulated kinase (ERK) signaling in
intestinal epithelial cells (15).
SERPINE2 could also promote drug resistance in osteosarcoma
(19) and lymph node metastasis in
testicular germ cell tumors (20).
Fayard has described a novel pathway where SERPINE2, by binding to
lipoprotein receptor-related protein (LRP)-1, stimulates ERK
signaling and matrix metalloproteinase (MMP)-9 expressions and
promotes the metastatic spread of mammary tumors (6).
In contrast, we previously found that SERPINE2
presents in both normal and benign prostate hypertrophy tissues
while its level reduced in prostate carcinoma with a higher Gleason
score (21). The SERPINE2-mediated
inhibition of uPA serves to prevent prostate cell invasion and
metastasis, whereas MMP9 upregulates uPA and facilitates tumor cell
invasion through cleavage of SERPINE2 (22). We further showed that SERPINE2
inhibits Hedgehog signaling by reducing the hedgehog ligand Sonic
(SHH), which could downregulate downstream molecules, including
cyclin D (CCND) 1 and glioma-associated oncogene family zinc finger
1 (GLI1), and as a result inhibits tumor cell proliferation and
reduces the expression of B-cell CLL/lymphoma 2 (BCL2) to promote
apoptosis (23). SERPINE2 was
identified to exert pro-apoptosis activity in prostate carcinoma by
down-regulating X-linked inhibitor of apoptosis protein (XIAP)
through blocking the NF-κB signal pathway and changing the
stability of XIAP through inhibition of its phosphorylation
(8), indicating a protective role of
SERPINE2 in tumor progression.
In brain tumors, SERPINE2 plays a more complex role.
Pagliara showed that SERPINE2 affects glioma cell migration and
invasiveness through the regulation of uPA and MMP-9/2 expression
levels, contributing to the degradation of the ECM during tumor
invasion (24). However, another
study showed that SERPINE2 promoted pre-neoplastic lesion
progression to the medulloblastoma owing to aberrant Hedgehog
pathway activity independent of the SHH ligand (25).
The biological function of SERPINE2 in different
types of tumor appears diverse. In this study, we attempted to
determine the expression profiles of SERPINE2 through the
EST database, immunohistochemical staining of human tissue
microarrays that contains 24 common tumors and normal tissues. The
present study sought to systemically review the expression patterns
of SERPINE2 in a variety of human normal and tumor tissues,
therefore guide us to focus on studying the functional mechanism of
SERPINE2 in specific tumors.
Materials and methods
SERPINE2 transcript data
The SERPINE2 transcript data (UniGene
136630-Hs.38449) were obtained from the EST profile in the NIH
UniGene database (http://www.ncbi.nlm.nih.gov/UniGene/ESTProfileViewer),
represented by 1231 ESTs from 282 complementary DNA (cDNA)
libraries and corresponding to 4 reference sequences (different
isoforms), on January 1st, 2017. The cDNA libraries used for
generating the ESTs are themselves derived from a multitude of
different organs/tissues/cell types and developmental stages. EST
and cDNA sequences provide direct evidence for all of the sampled
transcripts, and they are currently the most important resources
for transcriptome exploration (26).
Data processing was performed using the GraphPad Prism 5
software.
Tissue microarray
Paraformaldehyde-fixed paraffin-embedded human
multiple organ tissue microarray slides were purchased from US
Biomax Inc. (catalog nos. FDA808c-1 and FDA808c-2; Rockville, MD,
USA). The FDA808c-1 slide contained 72 samples of 57 normal tissues
and 15 cancer adjacent tissues; the FDA808c-2 slide contained 13
normal tissues, 5 cancer adjacent tissues and 54 malignant tumor
tissues at different pathological stages.
Reagents and antibodies
Rabbit anti-human SERPINE2 monoclonal antibody (Ab)
was purchased from Protein Tech Group (Wuhan, China). The EnVision™
detection kit, including ChemMate™ EnVision+/HRP, rabbit/mouse and
DAB, was purchased from Goodbio Technology (Wuhan, China).
Immunohistochemistry
Immunohistochemical staining was carried out to
detect SERPINE2 in situ using an anti-SERPINE2 rabbit
anti-human monoclonal antibody according to the manufacturer's
protocol. Briefly, the slides were incubated at 60°C for 2 h,
followed by three 10-minute cycles of deparaffinization in
environmental protection dewaxing liquid and then hydration in a
graded ethanol series. The slides were pretreated to promote
antigen retrieval in a microwave oven for 15 min in sodium citrate,
pH 6.0. Endogenous peroxidase in the sections was blocked with
hydrogen peroxide (0.03%). Bovine serum albumin was added to
minimize the background. A 1:50 dilution of the SERPINE2 antibody
was applied to the sections, which were then incubated overnight at
4°C. Next, the sections were incubated with horseradish peroxidase
(HRP)-conjugated anti-mouse/rabbit IgG Ab secondary antibody. For
visualization, diaminobenzidine (DAB) was used as a chromogen
substrate. Both slides were counterstained with hematoxylin.
Negative controls were similarly processed by omitting the primary
antibody. Tissue microarrays were imaged at 200× and 400×
magnification.
Results
The transcriptional level of SERPINE2
differs among human tissues and organs and changes in the stages of
human body growth and development
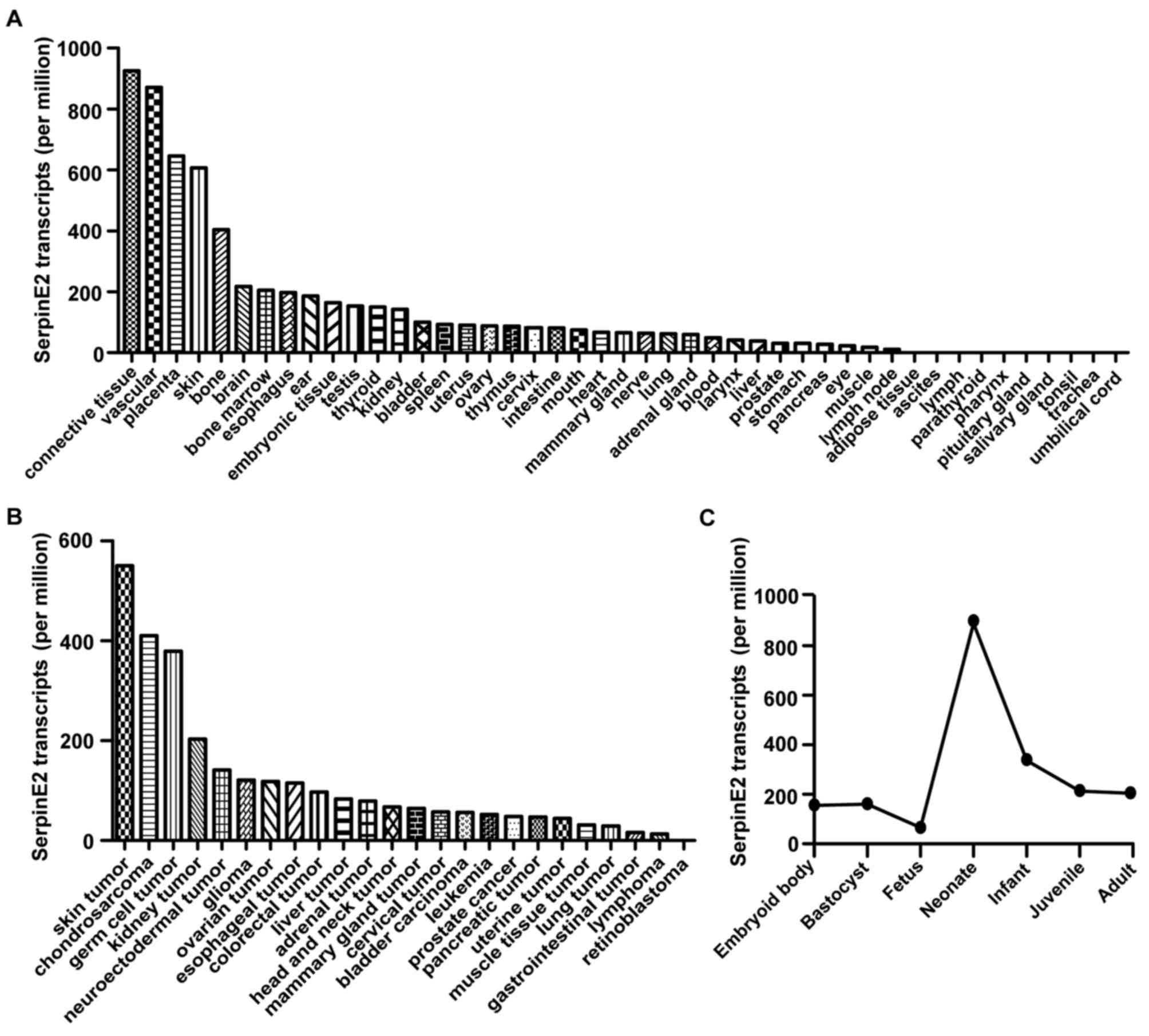
The EST profile in the NIH UniGene database shows
that SERPINE2 is widely expressed in human tissues and
organs. We found SERPINE2 transcripts in 77.78% (35 of 45)
of the tested tissues and organs (Fig.
1A). The expression of SERPINE2 varied among different
tissues. It was abundant in vascular connective tissue, placenta,
skin, and bone tissues, compared to bone marrow, esophagus, ear,
embryonic tissue, testis, thyroid, kidney and bladder tissues. No
transcript was found in umbilical cord, trachea, tonsil, salivary
gland, pituitary gland, pharynx, parathyroid, lymph, ascites and
adipose tissues. Beside its variation among different tissues,
SERPINE2 expression changed at different stages of
development.
It is readily detectable in normal tissues and
non-neoplastic tissues, as well as in tumor tissues according to
the EST database. Retinoblastoma was the only tumor among the 24
types of tumors without SERPINE2 transcript, indicating that
95.83% (23 of 24) of the tumor tissues tested expressed SERPINE2
(Fig. 1B). The expression of
SERPINE2 differs among different tumors and is significantly
higher in skin tumors, chondrosarcoma and germ cell tumors than the
other tumor types.
In addition, SERPINE2 is expressed throughout
all the stages of human growth, including the embryonic body,
blastocyst, fetus, neonate, infant, juvenile and adult stages. As
human body grows and develops, SERPINE2 expression begins to
climb at the fetus stage, reaches a peak at the neonate and remains
steady after entering juvenile period (Fig. 1C).
SERPINE2 is preferentially expressed
in gland-rich organs and adenocarcinomas
To investigate the expression profile of SERPINE2,
we conducted immunohistochemistry on two sets of tissue microarrays
(cat. nos. FDA808c-1 and FDA808c-2, from US Biomax Inc.) which
contained various normal tissues and tumor tissues. In consistency
with the EST profile, we found that the expression of SERPINE2 was
detectable in most of the tissues, especially enriched in glandular
organs, such as uterine endometrium, prostate, ovary, colon and
digestive organs. The cognate adenocarcinomas abundantly expressed
SERPINE2 (Fig. 2A). Skin tumors
(basal adenocarcinoma and malignant melanoma of the skin) as well
as liver tumors (hepatocellular carcinoma and hepatoblastoma)
exhibited even higher levels of SERPINE2 (Fig. 2B). Notably, the histological
expression of SERPINE2 in the skin basically matches its EST level.
SERPINE2 was plentiful in the pituitary glands, salivary glands and
tonsils (data not shown), in keeping with our previous observation
in other gland-rich organs, however not consistent with the EST
profile, suggesting post-transcriptional elements may take part in
the regulation in these organs. Taken together, SERPINE2 is
preferentially expressed in gland-rich organs, especially
adenocarcinomas.
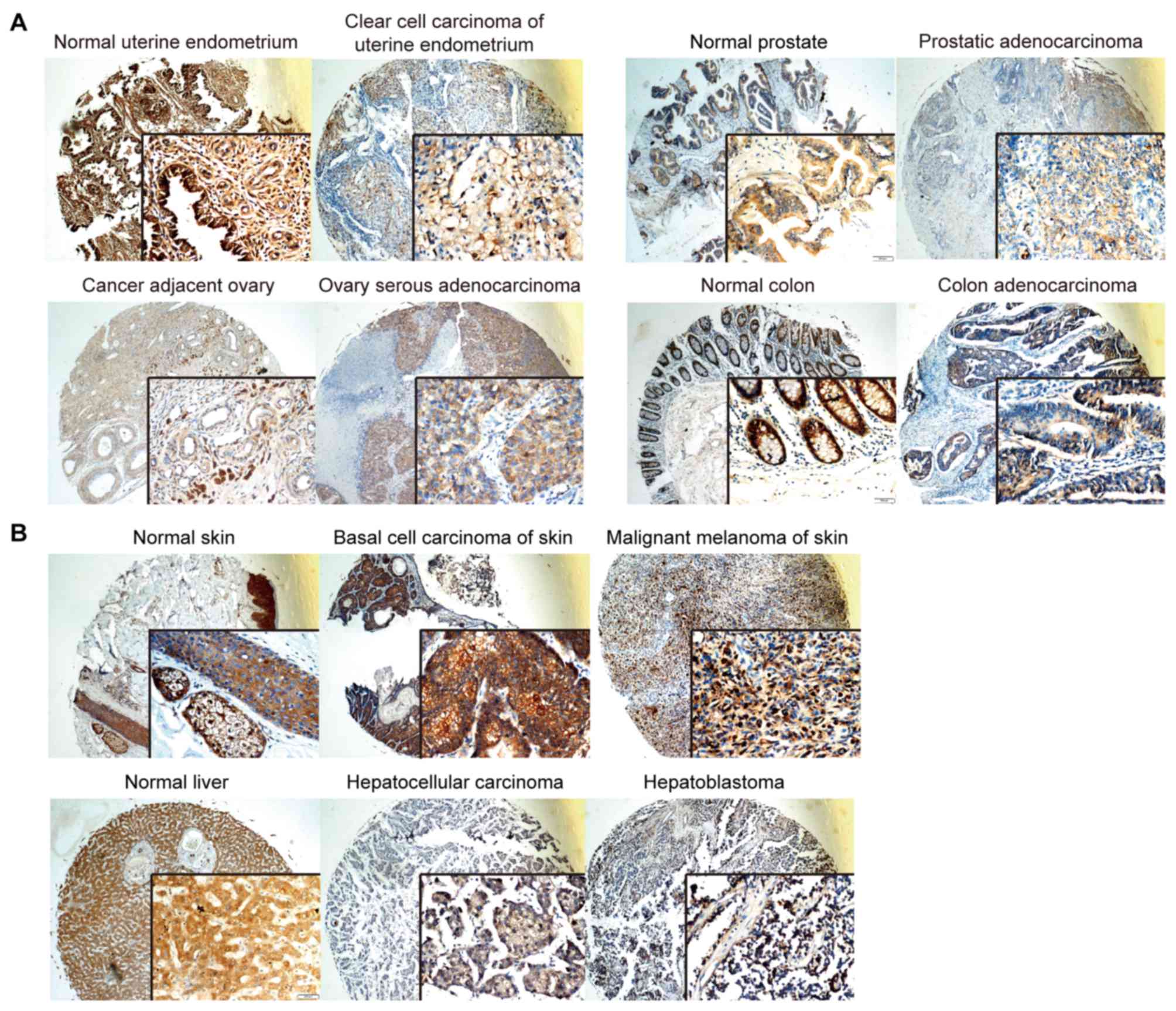 | Figure 2.In a number of organs, SERPINE2
protein levels are high in both tumor tissues and their normal
counterparts. SERPINE2 protein is stained in brown by
immunohistochemistry. (A) SERPINE2 protein expressions were
abundant in normal and adenocarcinoma tissues of uterine
endometrium, prostate, ovary and colon, which are gland-rich
organs. (B) SERPINE2 protein was enriched in normal skin, basal
adenocarcinoma and malignant melanoma of skin, as well as in normal
liver, hepatocellular carcinoma and hepatoblastoma. Magnification,
×200 (main image) and ×400 (box at bottom right). SERPINE2, serine
proteinase inhibitor, clade E member 2. |
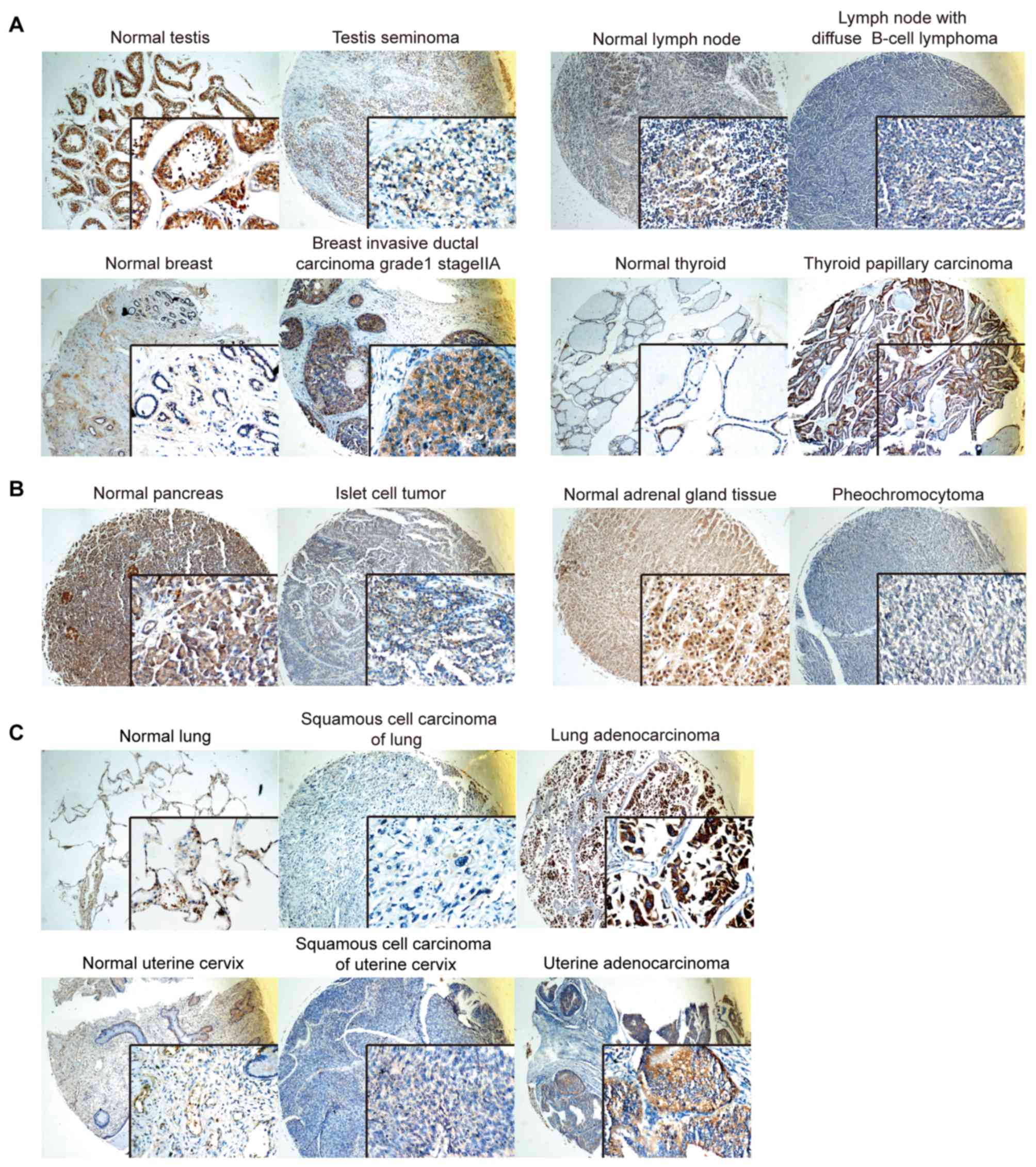
SERPINE2 expression differs in most
tumors and their counterparts of normal tissues
To understand the differential expression of
SERPINE2 in various types of tumors, we compared SERPINE2
expression between tumor and their corresponding normal tissues.
For instance, in testis seminoma and lymph nodes with diffuse B
cell lymphoma, SERPINE2 expression was remarkably lower than that
of their corresponding normal tissues (Fig. 3A). Likewise, SERPINE2 expression was
lower in colon adenocarcinoma (Fig.
2A) than that in its normal counterpart. However, breast
invasive ductal carcinoma and thyroid papillary carcinoma have
significantly higher expression SERPINE2 than their normal
counterparts (Fig. 3A). SERPINE2
expression was slightly lower in the benign tumors, such as islet
cell tumors and pheochromocytomas of the adrenal gland, in
comparison of the corresponding normal tissues (Fig. 3B).
Interestingly, SERPINE2 expressions were more
profound in adenocarcinomas of lung and uterine cervix than either
normal tissue or the squamous cell carcinomas originated from the
same organ (Fig. 3C). The same
scenario is also found in skin and uterus adenocarcinoma (data not
shown). In summary, SERPINE2 appears highly expressed in the
adenocarcinomatous tumor, but with lower levels in squamous cell
carcinoma of the same tissue origin, further supporting its
preference for glandular tissues.
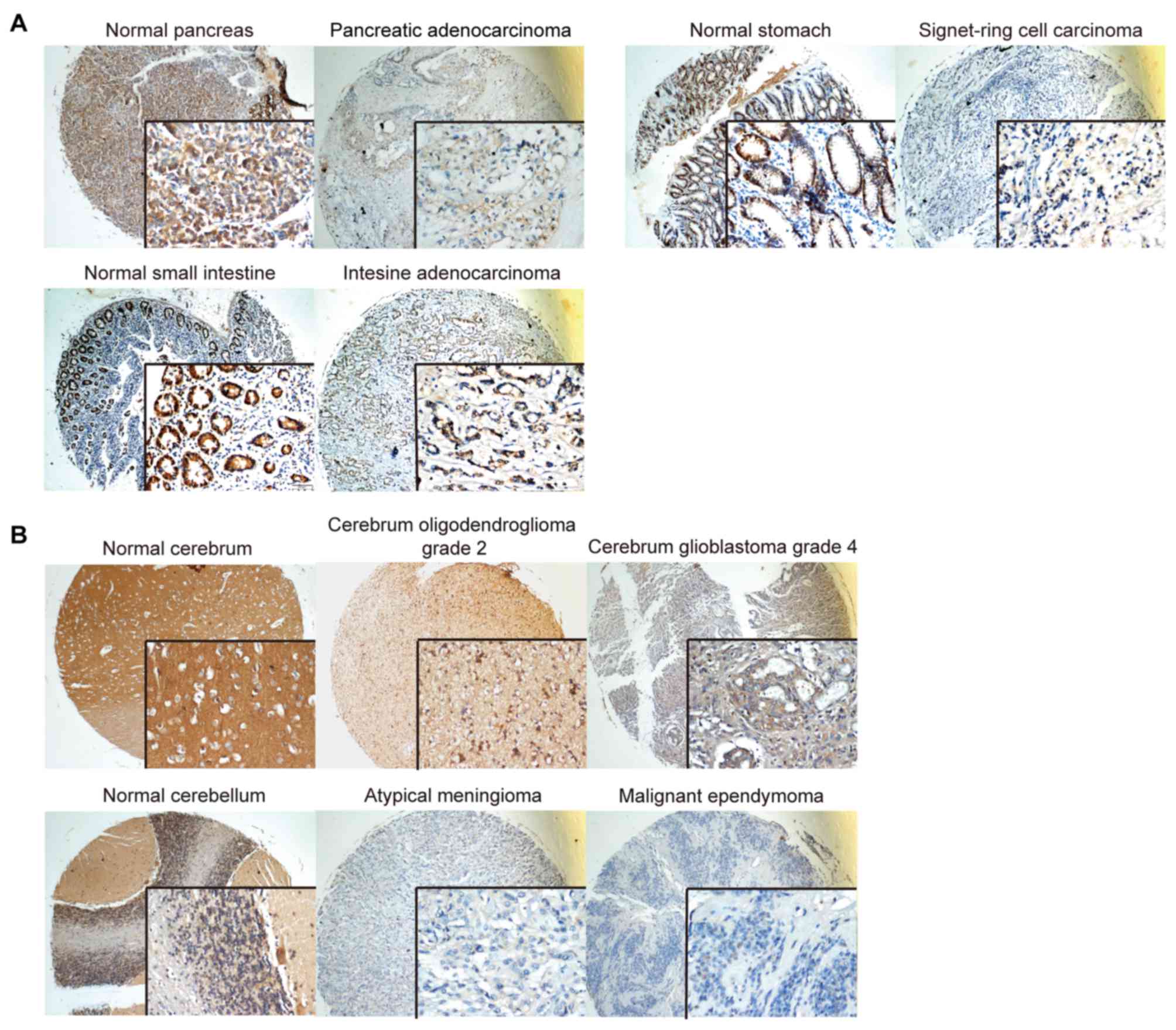
The role of SERPINE2 was originally highlighted in
the brain. Here, we observed high SERPINE2 abundance in the normal
cerebrum, normal cerebellum, cerebrum oligodendroglioma and
glioblastoma and extremely low levels in atypical meningioma and
malignant ependymoma (Fig. 4), all of
which are epithelial derived. There are low to medium levels of
SERPINE2 expression in mesenchymal tumors such as embryonal
rhabdomyosarcoma, osteosarcoma and intestine stromal sarcoma (data
not shown).
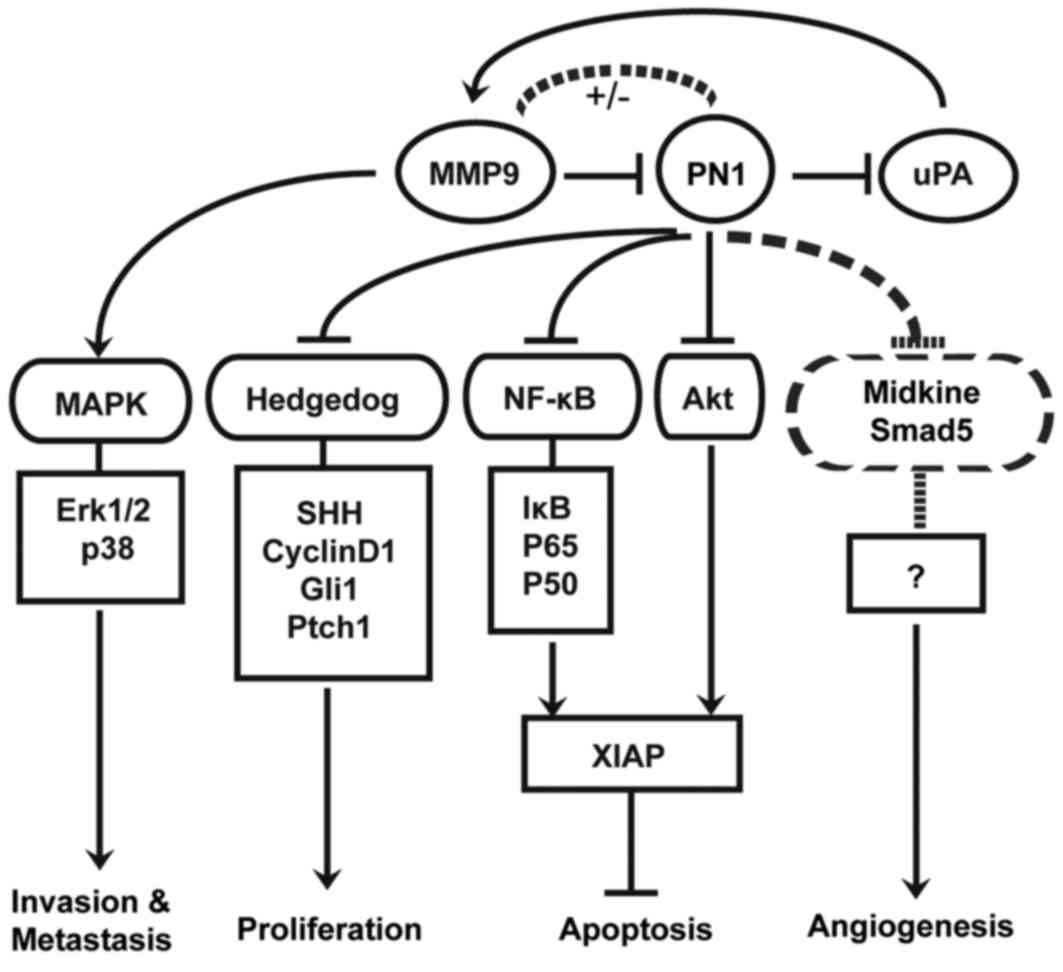
The molecular mechanisms how SERPINE2
regulates oncogenesis
Lastly, we provide a diagram to summarize the
established molecular mechanism how SERPINE2 regulates oncogenesis
(Fig. 5). SERPINE2 inhibits Hedgehog
signaling by down-regulating SHH and its down-stream targets
including PTCH1, GLI1 and CCND1, thus inhibiting proliferation of
prostate cancer cells and conferring protective activity (21). Previously, we found that SERPINE2 has
anti-angiogenesis activity in human prostate cancer. Recently,
SERPINE2 was found involved in angiogenesis through mediation of
midkine and SMAD5 pathways in the retina (27,28).
Moreover, SERPINE2 promotes the apoptosis of
prostate cancer cells by reducing the transcriptional level of
XIAP through NF-κB signaling activator p65 and blunting XIAP
activity through Akt signaling. The dual regulation of XIAP can be
facilitated through inhibition of uPA (8). In glioma, SERPINE2 knock-down leads to
an increase in uPA that indirectly activates MMP9, and enhances
glioma cell migration and invasion via the Erk1/2 and p38 pathways
through the degradation of ECM components (24). However SERPINE2 was identified to
stimulate the Ras-ERK signaling pathway through syndecan-1-mediated
internalization in mouse embryonic fibroblasts (29). SERPINE2 was demonstrated to
inhibit MMP9 transcriptional activity in SERPINE2
deficient mice (30). By contrast,
SERPINE2 was reported to activate MMP9 expression and promote
cancer metastasis in mammary tumors (6). Indeed, since the interaction between
SERPINE2 and MMP9 is complex, the resultant effects can be
discrepant. Furthermore, SERPINE2 was shown to be involved in BRAF-
and MEK-induced tumorigenesis in colon epithelial cells and to
promote pancreatic cancer invasion by up-regulating ECM production
(14,15).
Discussion
The abundant expression of SERPINE2 in the glandular
tissues is prominent in our tissue microarray studies (Figs. 2A and 4A), suggesting its extracellular function
may associate with glandular secretion. Indeed, it has been
reported that SERPINE2 was highly expressed in the endometrium
during the secretory phase compared with the proliferative phase,
indicating that it may participate in tissue remodeling during
implantation in the placenta and uterus (31,32). A
high SERPINE2 level was found in the inhibition of cumulus
expansion and oocyte maturation (33). Interestingly, chromogranin A could
promote secretory granule biogenesis in endocrine cells through
up-regulation of the expression of SERPINE2 via a cAMP-protein
kinase A-Sp1 pathway to replenish released granules (34). In consistency with our results,
SERPINE2 was found significantly elevated in estrogen receptor (ER)
A-negative and in high-grade breast cancer (6,35). ER can
dimerize and bind to specific DNA sequences and estrogen response
elements to stimulate estrogen-target gene transcription. It
remains unknown whether the regulation of SERPINE2 transcription in
breast cancer is ER dependent. Because the involvement of hormones
in the progression of many tumors such as estrogen-dependent
mammary tumors has been well established, the interaction between
hormones and SERPINE2 in tumors may be worthy of further
investigation.
Our data show that adenocarcinomatous tissues of the
lung and uterine cervix highly express SERPINE2, in contrast to a
very scanty expression in squamous cell tumor originated from the
same organ (Fig. 3C). In the
meanwhile, a low SERPINE2 expression level was found in most
mesenchymal tumors (data not show). These findings suggest that
SERPINE2 may associate with tumor development of a certain
histological subtype. Due to such heterogeneity, the role of
SERPINE2 in different subtypes of tumor remains controversial. The
protective role of SERPINE2 in tumors was reported in prostate
carcinoma and glioma (8,23,24). These
studies provide substantial insights into the favorable function of
SERPINE2 against tumors by inhibition of tumor growth through the
hedgehog pathway and promotion of apoptosis through inhibiting XIAP
via the NF-κB pathway and Akt pathway. SERPINE2 can also prevent
invasion and metastasis by uPA degradation and lead to reduced
activation of the MAPK pathway through down-regulating MMP9. It is
also suggested that SERPINE2 may function as an anti-angiogenesis
factor by regulating midkine and SMAD5 (28).
On the other hand, Buchholz et al (14), reported that SERPINE2 expression was
trivial or absent in all normal pancreas and tissue samples of
chronic pancreatitis but abundant in most of the pancreatic
carcinoma as well as gastric and colorectal cancer. SERPINE2
overexpression enhances the invasive potential of pancreatic cancer
cells in nude mouse xenografts by altering ECM production and
organization within the tumors, thus contributing to the aggressive
phenotype of pancreatic cancer (36)
and the progression of colorectal cancer by down-regulation of
prostasin activity (37). In
osteosarcoma, SERPINE2 expression is higher in tissue from patients
with metastasis and a tumor-node-metastasis stage II–III, and
increases after chemotherapy. In vitro experiments show that
SERPINE2 can stimulate cell proliferation and promote
drug-resistance in osteosarcoma (19). Thus SERPINE2 appears pro-neoplastic in
such scenarios (15). Ultimately,
tissue specificity, individual genetic differences, and different
expression levels may all contribute to the complicated effects of
SERPINE2 in various types of tumors. Therefore, it may be
understandable that SERPINE2 can be protective or detrimental in
the process of tumor development.
As a glia-derived nexin, SERPINE2 is involved in
regulating astrocyte proliferation, neurite outgrowth, neuron
migration and localization, contributing to the development of
brain, and the regeneration and reconstruction of neurons (38–41). Our
studies of EST profile showed that SERPINE2 expression begins to
climb at early as the fetus stage, reaches a peak at the neonate
and plateaus after the juvenile stage (Fig. 1C), suggesting its roles in human
growth and development. The increased expression of SERPINE2 in the
neonate stage with the remarkable development of nervous systems
annouces its potential role in nervous system development. Indeed,
SERPINE2 may contribute to the development of the brain by
modulating the proliferation and differentiation of cerebellar
granular neuron precursors via the major mitogen SHH, as well as
via granule neuron migration and positioning (41). SERPINE2 may exert protective activity
in the pathalogical progression of intracranial hemorrhage
(42), cerebral ischemia (40), and Alzheimer's disease (43). Again, the role of SERPINE2 in tumor
progression in the brain seems to be a paradox. The downregulation
of SERPINE2 may activate the Erk1/2 and P38 signaling pathways and
lead to an increased invasion and metastasis of glioma cells, in
which SERPINE2 seems to play a pro-tumor role (24). In another study, however, SERPINE2 may
mediate an anti-tumor activity as its deficiency leads to the
reduced proliferation of medulloblastoma cells (25). Our tissue microarray shows that
SERPINE2 is expressed differentially in different types of brain
tumors: abundant in oligodendroglioma and glioblastoma and
extremely low in atypical meningioma and malignant ependymoma
(Fig. 4).
Our study reveals that expression and function of
SERPINE2 may vary in different kinds of tumors. To date, the
research on anti-neoplastic mechanisms are more comprehensive than
pro-neoplastic ones. The distinctive expression pattern found in
this study may help us to understand the function of SERPINE2 in
specific tumor types, therefore guide the research of SERPINE2 as a
potential tumor marker for diagnosis and prognosis.
Acknowledgements
The present study was kindly funded by the National
Natural Science Foundation of China (81101572, 81372326) and the
Young Distinguish Talents grant of Tongji Hospital.
References
|
1
|
Barde YA, Lindsay RM, Monard D and Thoenen
H: New factor released by cultured glioma cells supporting survival
and growth of sensory neurones. Nature. 274:8181978. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Guenther J, Nick H and Monard D: A
glia-derived neurite-promoting factor with protease inhibitory
activity. EMBO J. 4:1963–1966. 1985.PubMed/NCBI
|
|
3
|
Stone SR, Nick H, Hofsteenge J and Monard
D: Glial-derived neurite-promoting factor is a slow-binding
inhibitor of trypsin, thrombin and urokinase. Arch Biochem Biophys.
252:237–244. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gloor S, Odink K, Guenther J, Nick H and
Monard D: A glia-derived neurite promoting factor with protease
inhibitory activity belongs to the protease nexins. Cell.
47:687–693. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mansilla S, Boulaftali Y, Venisse L,
Arocas V, Meilhac O, Michel JB, Jandrot-Perrus M and Bouton MC:
Macrophages and platelets are the major source of protease nexin-1
in human atherosclerotic plaque. Arterioscler Thromb Vasc Biol.
28:1844–1850. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fayard B, Bianchi F, Dey J, Moreno E,
Djaffer S, Hynes NE and Monard D: The serine protease inhibitor
protease nexin-1 controls mammary cancer metastasis through
LRP-1-mediated MMP-9 expression. Cancer Res. 69:5690–5698. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rosenblatt DE, Cotman CW, Nieto-Sampedro
M, Rowe JW and Knauer DJ: Identification of a protease inhibitor
produced by astrocytes that is structurally and functionally
homologous to human protease nexin-I. Brain Res. 415:40–48. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
McKee CM, Ding Y, Zhou J, Li C, Huang L,
Xin X, He J, Allen JE, El-Deiry WS, Cao Y, et al: Protease nexin 1
induces apoptosis of prostate tumor cells through inhibition of
X-chromosome-linked inhibitor of apoptosis protein. Oncotarget.
6:3784–3796. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Santoro A, Conde J, Scotece M, Abella V,
Lois A, Lopez V, Pino J, Gomez R, Gomez-Reino JJ and Gualillo O:
SERPINE2 Inhibits IL-1α-Induced MMP-13 Expression in Human
Chondrocytes: Involvement of ERK/NF-κB/AP-1 Pathways. PLoS One.
10:e01359792015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Crisp RJ, Knauer DJ and Knauer MF: Roles
of the heparin and low density lipid receptor-related
protein-binding sites of protease nexin 1 (PN1) in urokinase-PN1
complex catabolism. The PN1 heparin-binding site mediates complex
retention and degradation but not cell surface binding or
internalization. J Biol Chem. 275:19628–19637. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Boulaftali Y, Ho-Tin-Noe B, Pena A, Loyau
S, Venisse L, François D, Richard B, Arocas V, Collet JP,
Jandrot-Perrus M and Bouton MC: Platelet protease nexin-1, a serpin
that strongly influences fibrinolysis and thrombolysis.
Circulation. 123:1326–1334. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kitamura K and Tomita K: Regulation of
renal sodium handling through the interaction between serine
proteases and serine protease inhibitors. Clin Exp Nephrol.
14:405–410. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zheng D, Chen H, Davids J, Bryant M and
Lucas A: Serpins for diagnosis and therapy in cancer. Cardiovasc
Hematol Disord Drug Targets. 13:123–132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Buchholz M, Biebl A, Neesse A, Wagner M,
Iwamura T, Leder G, Adler G and Gress TM: SERPINE2 (protease nexin
I) promotes extracellular matrix production and local invasion of
pancreatic tumors in vivo. Cancer Res. 63:4945–4951.
2003.PubMed/NCBI
|
|
15
|
Bergeron S, Lemieux E, Durand V, Cagnol S,
Carrier JC, Lussier JG, Boucher MJ and Rivard N: The serine
protease inhibitor serpinE2 is a novel target of ERK signaling
involved in human colorectal tumorigenesis. Mol Cancer. 9:2712010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang K, Wang B, Xing AY, Xu KS, Li GX and
Yu ZH: Prognostic significance of SERPINE2 in gastric cancer and
its biological function in SGC7901 cells. J Cancer Res Clin Oncol.
141:805–812. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stępień T, Brożyna M, Kuzdak K, Motylewska
E, Komorowski J, Stępień H and Ławnicka H: Elevated concentrations
of SERPINE2/protease nexin-1 and secretory leukocyte protease
inhibitor in the serum of patients with papillary thyroid cancer.
Dis Markers. 2017:49621372017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gao S, Krogdahl A, Sørensen JA, Kousted
TM, Dabelsteen E and Andreasen PA: Overexpression of protease
nexin-1 mRNA and protein in oral squamous cell carcinomas. Oral
Oncol. 44:309–313. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mao M and Wang W: SerpinE2 promotes
multiple cell proliferation and drug resistance in osteosarcoma.
Mol Med Rep. 14:881–887. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nagahara A, Nakayama M, Oka D, Tsuchiya M,
Kawashima A, Mukai M, Nakai Y, Takayama H, Nishimura K, Jo Y, et
al: SERPINE2 is a possible candidate promotor for lymph node
metastasis in testicular cancer. Biochem Biophys Res Commun.
391:1641–1646. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
McKee CM, Xu D, Cao Y, Kabraji S, Allen D,
Kersemans V, Beech J, Smart S, Hamdy F, Ishkanian A, et al:
Protease nexin 1 inhibits hedgehog signaling in prostate
adenocarcinoma. J Clin Invest. 122:4025–4036. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu D, McKee CM, Cao Y, Ding Y, Kessler BM
and Muschel RJ: Matrix metalloproteinase-9 regulates tumor cell
invasion through cleavage of protease nexin-1. Cancer Res.
70:6988–6998. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
McKee CM, Xu D and Muschel RJ: Protease
nexin 1: A novel regulator of prostate cancer cell growth and
neo-angiogenesis. Oncotarget. 4:1–2. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pagliara V, Adornetto A, Mammi M, Masullo
M, Sarnataro D, Pietropaolo C and Arcone R: Protease Nexin-1
affects the migration and invasion of C6 glioma cells through the
regulation of urokinase plasminogen activator and matrix
metalloproteinase-9/2. Biochim Biophys Acta. 1843:2631–2644. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vaillant C, Valdivieso P, Nuciforo S, Kool
M, Schwarzentruber-Schauerte A, Méreau H, Cabuy E, Lobrinus JA,
Pfister S, Zuniga A, et al: Serpine2/PN-1 is required for
proliferative expansion of pre-neoplastic lesions and malignant
progression to medulloblastoma. PLoS One. 10:e01248702015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Boguski MS and Schuler GD: ESTablishing a
human transcript map. Nat Genet. 10:369–371. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Selbonne S, Azibani F, Iatmanen S,
Boulaftali Y, Richard B, Jandrot-Perrus M, Bouton MC and Arocas V:
In vitro and in vivo antiangiogenic properties of the serpin
protease nexin-1. Mol Cell Biol. 32:1496–1505. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Selbonne S, Francois D, Raoul W,
Boulaftali Y, Sennlaub F, Jandrot-Perrus M, Bouton MC and Arocas V:
Protease nexin-1 regulates retinal vascular development. Cell Mol
Life Sci. 72:3999–4011. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li X, Herz J and Monard D: Activation of
ERK signaling upon alternative protease nexin-1 internalization
mediated by syndecan-1. J Cell Biochem. 99:936–951. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu D, Suenaga N, Edelmann MJ, Fridman R,
Muschel RJ and Kessler BM: Novel MMP-9 substrates in cancer cells
revealed by a label-free quantitative proteomics approach. Mol Cell
Proteomics. 7:2215–2228. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chern SR, Li SH, Lu CH and Chen EI:
Spatiotemporal expression of the serine protease inhibitor,
SERPINE2, in the mouse placenta and uterus during the estrous
cycle, pregnancy, and lactation. Reprod Biol Endocrinol. 8:1272010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee RK, Fan CC, Hwu YM, Lu CH, Lin MH,
Chen YJ and Li SH: SERPINE2, an inhibitor of plasminogen
activators, is highly expressed in the human endometrium during the
secretory phase. Reprod Biol Endocrinol. 9:382011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lu CH, Lee RK, Hwu YM, Lin MH, Yeh LY,
Chen YJ, Lin SP and Li SH: Involvement of the serine protease
inhibitor, SERPINE2, and the urokinase plasminogen activator in
cumulus expansion and oocyte maturation. PLoS One. 8:e746022013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Koshimizu H, Cawley NX, Kim T, Yergey AL
and Loh YP: Serpinin: A novel chromogranin A-derived, secreted
peptide up-regulates protease nexin-1 expression and granule
biogenesis in endocrine cells. Mol Endocrinol. 25:732–744. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Martin KJ, Patrick DR, Bissell MJ and
Fournier MV: Prognostic breast cancer signature identified from 3D
culture model accurately predicts clinical outcome across
independent datasets. PLoS One. 3:e29942008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li Y and Lu YY: Applying a highly specific
and reproducible cDNA RDA method to clone garlic up-regulated genes
in human gastric cancer cells. World J Gastroenterol. 8:213–216.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Selzer-Plon J, Bornholdt J, Friis S,
Bisgaard HC, Lothe IM, Tveit KM, Kure EH, Vogel U and Vogel LK:
Expression of prostasin and its inhibitors during colorectal cancer
carcinogenesis. BMC Cancer. 9:2012009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cavanaugh KP, Gurwitz D, Cunningham DD and
Bradshaw RA: Reciprocal modulation of astrocyte stellation by
thrombin and protease nexin-1. J Neurochem. 54:1735–1743. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lindner J, Guenther J, Nick H, Zinser G,
Antonicek H, Schachner M and Monard D: Modulation of granule cell
migration by a glia-derived protein. Proc Natl Acad Sci USA. 83:pp.
4568–4571. 1986; View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mirante O, Price M, Puentes W, Castillo X,
Benakis C, Thevenet J, Monard D and Hirt L: Endogenous protease
nexin-1 protects against cerebral ischemia. Int J Mol Sci.
14:16719–16731. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vaillant C, Michos O, Orolicki S, Brellier
F, Taieb S, Moreno E, Té H, Zeller R and Monard D: Protease nexin 1
and its receptor LRP modulate SHH signalling during cerebellar
development. Development. 134:1745–1754. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wu H, Zhao R, Qi J, Cong Y, Wang D, Liu T,
Gu Y, Ban X and Huang Q: The expression and the role of protease
nexin-1 on brain edema after intracerebral hemorrhage. J Neurol
Sci. 270:172–183. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Vaughan PJ, Su J, Cotman CW and Cunningham
DD: Protease nexin-1, a potent thrombin inhibitor, is reduced
around cerebral blood vessels in Alzheimer's disease. Brain Res.
668:160–170. 1994. View Article : Google Scholar : PubMed/NCBI
|