Introduction
Head and neck squamous cell carcinoma (HNSCC)
includes epithelial malignancies of the oral cavity, oropharynx,
hypopharynx and larynx (1). Laryngeal
carcinoma originates from the larynx, which is divided into three
regions: The supraglottis, glottis and subglottis (2). Smoking is by far the principal risk
factor for laryngeal cancer, followed by alcohol consumption
(3). However, dietary factors,
including vitamin and fiber intake, are reported to be protective
factors for laryngeal cancer (3). In
addition to these factors, genetic factors contribute to the
development of laryngeal cancer (4).
Therefore, a more detailed understanding of the complex molecular
mechanisms leading to the development of laryngeal cancer is still
required.
Lacrimal proline-rich 4 (PRR4), also known as
nasopharyngeal carcinoma-associated proline-rich protein, was
discovered in 1995 by Dickson and Thiesse as a PRR protein (PRP)
synthesized in the acinar cells of the human lacrimal glands
(5). In previous studies, a decrease
in the PRR4 protein level in tear fluid was associated with
pathological conditions, including dry eye syndrome (DES),
thyroid-associated orbitopathy and diabetic proliferative
rethinopathy (6–8). It has also been demonstrated that the
gene encoding the PRR4 protein is highly expressed in the
human submucosal glands (9,10).
Submucosal glands occur in the upper respiratory
tract, visual and auditory systems, and the throat and intestines
of mammals. The majority of airway mucus is produced by the
submucosal glands. Salivary PRPs constitute approximately
two-thirds of the proteins secreted by the parotid gland (11,12). PRPs
are naturally unfolded, with no stable tertiary structure (13,14).
Salivary PRPs make up 70% of the proteins in saliva and are well
characterized (11,12). They have several functions including
the inhibition of calcium phosphate precipitation, binding
bacterial pathogens and binding to dietary tannins (15–17).
The development of novel techniques has allowed the
identification of novel macromolecules that may have critical
functions in the development and progression of cancer. In the
present study, a GeneFishing Assay (18) was performed in order to identify novel
genes that participate in laryngeal carcinogenesis. It was
identified that the PRR4 gene was one of the downregulated
genes, which may be due to the functional importance of PRR4
in laryngeal carcinogenesis. PRR4 mRNA expression levels in
the tumor and adjacent normal tissues from 90 patients with
laryngeal cancer were also investigated.
Materials and methods
Tissue samples
A total of 90 tumor tissues and matched
non-cancerous tissue samples were obtained from patients diagnosed
with laryngeal cancer undergoing surgery in the Department of
Otorhinolaryngology at the Cerrahpasa Medical Faculty, Istanbul
University (Istanbul, Turkey). A total of 87 men (96.7%) and 3
women (3.3%) were recruited in the present study. The age range of
patients was 39–81 years and the mean age at diagnosis was 60±9
years. The samples were fresh tissues obtained during surgery and
processed immediately. Tumor and normal tissue differentiation were
confirmed by a pathologist under a microscope. Only samples with a
tumor/stroma cell content >70% were included in the study as
tumor samples. Oral saliva was collected from 50 healthy
volunteers, including 25 smokers and 25 non-smokers. The subjects
did not eat or drink for 2 h prior to saliva collection, and their
mouth was rinsed with sterile MilliQ water (Merck KGaA, Darmstadt,
Germany). Saliva was collected into a DNase- and RNase-free 50-ml
Falcon tube. The Falcon tube was kept on ice during the collection
procedure. The saliva was processed immediately after
collection.
The present study was approved by the Cerrahpasa
Medical Faculty Ethics Committee (approval no.,
83045809/604.01/02-235918), and was performed in accordance with
the 2013 Declaration of Helsinki. Signed informed consent was
obtained from all patients prior to the study.
GeneFishing assay
RNA isolation and first-strand cDNA
synthesis
Total RNA was extracted from the tumor and adjacent
non-cancerous tissues of 4 patients using the miRCURY™ RNA
Isolation kit (Exiqon A/S, Vedbaek, Denmark) according to the
manufacturer's protocol. A total of 3 µg RNA was used for first
strand cDNA synthesis. The reaction conditions were as follows: 1
µM dT-ACP1 (provided in the GeneFishing™ DEG Premix kit; Seegene,
Inc., Seoul, South Korea), 1X reverse transcriptase buffer
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 0.5
mM dNTP, 20 U RNase inhibitor (Biomatik Corporation, Ontario,
Canada) and 200 U M-MLV reverse transcriptase (Invitrogen; Thermo
Fisher Scientific, Inc.) in a 20 µl final reaction volume. Reverse
transcription was performed at 42°C for 90 min and 94°C for 2 min.
First strand cDNA was diluted by adding 80 µl DNase-free water
prior to GeneFishing polymerase chain reaction (PCR).
GeneFishing PCR
A total of 20 different arbitrary annealing control
primers (ACPs) provided in the GeneFishing DEG Premix kit (Seegene
Inc.) were used for GeneFishing PCR. Diluted first-strand cDNA (50
ng) was used as a template in a reaction volume of 20 µl containing
0.5 µM arbitrary ACP (one type per reaction), 0.5 µM dT-ACP2 and 1X
SeeAmp™ ACP master mix. The reaction conditions were 94°C for 5
min, 50°C for 3 min, 72°C for 1 min, then 40 cycles of 94°C for 40
sec, 65°C for 40 sec and 72°C for 40 sec, and a final step at 72°C
for 5 min. GeneFishing PCR products were analyzed by
electrophoresis on 2% agarose gels, and all bands were quantified
using BioCapt analysis software (version 11.03; Vilber Lourmat
Deutschland GmbH, Eberhardzell, Germany). Bands were purified and
cloned if there was >10% difference in the band intensities
between the tumor and normal tissue. The differentially expressed
bands were extracted from the gels using the Zymoclean™ Gel DNA
Recovery kit (Zymo Research Corp., Irvine, CA, USA) according to
the manufacturer's protocol.
Cloning and sequencing
The purified PCR products were cloned into a TA
cloning vector using the TOPO TA Cloning kit for Sequencing
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The plasmid vector pCR™4-TOPO was used for
cloning. Subsequent to performing the cloning reaction, the plasmid
vector was transformed into One Shot® TOP10 Chemically
Competent E. coli (Thermo Fisher Scientific, Inc.) cells,
according to the manufacturer's protocol. E. coli cells were
spread onto Luria-Bertani (LB) agar plates containing 50 µg/ml
kanamycin and incubated overnight at 37°C. A total of 2–6 colonies
were selected and cultured overnight in LB medium containing 50
µg/ml kanamycin. The plasmid DNA was isolated using the PureLink™
Quick Plasmid Miniprep kit (Invitrogen; Thermo Fisher Scientific,
Inc.) and sequenced using an ABI Prism 3100-Avant™ Genetic Analyzer
(Applied Biosystems; Thermo Fisher Scientific, Inc.) using the ABI
Prism BigDye Terminator v3.1 Cycle Sequencing kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The DNA sequences were
analyzed by evaluating the data using the Basic Local Alignment
Search Tool program 2.4.0 (19).
Validation of the GeneFishing assay
data by RT-quantitative PCR (RT-qPCR)
Total RNA was extracted using the PureLink RNA Mini
kit (Ambion; Thermo Fisher Scientific, Inc.) from the 90 tumor and
non-cancerous adjacent tissues. Total RNA (400 ng) from each sample
was transcribed in a 20 µl reaction volume using the RevertAid
First Strand cDNA Synthesis kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. PRR4 gene
expression levels were analyzed by RT-qPCR using the LightCycler
480-II system (Roche Diagnostics GmbH, Mannheim, Germany). RT-qPCR
was performed in a final volume of 15 µl containing 1X master PCR
mix (SolGent, Inc., Daejeon, South Korea) with EvaGreen (Biotium,
Inc., Freemont, CA, USA), 600 nM gene-specific primers, cDNA and
nuclease free water. The β-actin gene was used as a housekeeping
gene for normalization of mRNA levels. The sequences of the primers
are presented in Table I. The PCR
cycling conditions were as follows: 95°C for 15 min, followed by 40
cycles at 95°C for 15 sec, 59°C for 30 sec and 72°C for 30 sec, and
a final 10 sec at 50°C. The relative quantification of mRNA levels
was calculated using the comparative 2−ΔΔCq method
(20). The expression levels of 12
differentially expressed RNAs in HNSCC or laryngeal cancer have
been analyzed in Gene Ontology, Serial Analysis of Gene Expression
and The Cancer Genome Atlas databases (21–23).
 | Table I.Sequences for the primers used
quantitative polymerase chain reaction in the present study. |
Table I.
Sequences for the primers used
quantitative polymerase chain reaction in the present study.
| Gene | Primer | Sequence |
|---|
| Proline-rich
4 | Forward |
5′-ACGAGGACACCGTCAACTCT-3′ |
| Reverse |
5′-TCAATGTCATGGCTTTCTGAAG-3′ |
| β-actin | Forward |
5′-CTCGCGCTACTCTCTCTTTCTGG-3′ |
| Reverse |
5′-GCTTACATGTCTCGATCCCACTTAA-3′ |
RNA isolation and cDNA synthesis from
saliva
Saliva samples were centrifuged at 11,000 × g for 20
min at 4°C to separate the supernatant from the cellular fraction.
The cell pellet was resuspended in 1 ml 1X PBS buffer, and RNA was
extracted from the cell pellet using the Hybrid-R Blood RNA kit
(GeneAll Biotechnology Co., Ltd., Seoul, South Korea) according to
the manufacturer's protocol. A total of 400 ng isolated RNA was
reverse-transcripted using the RevertAid First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.) according to the
manufactuer's protocol.
Statistical analysis
Statistical analyses were performed using SPSS
version 20 (IBM Corp., Armonk, NY, USA). A paired Student's t-test
or Pearson's χ2 test were used to calculate P-values.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Identification of differently
expressed genes
In order to identify the genes that were differently
expressed in tumor tissues compared with normal tissues, ACP-based
GeneFishing PCR was performed using a combination of 20 arbitrary
primers and two anchored oligo (dT) primers (dT-ACP1 and dT-ACP2).
A total of 4 pairs of tumor and normal tissues were used for the
GeneFishing assay.
A total of 27 differently expressed genes (DEGs)
were identified; of these, 15 DEGs were downregulated and 12 DEGs
were upregulated in tumor tissues compared with normal tissues.
Amongst the 27 DEGs, 12 DEGs were isolated, cloned, sequenced and
searched in GenBank if there was a difference of >10% in band
intensity. The isolated DEGs and their characteristics are
summarized in Table II. The
PRR4 gene was identified as one of the downregulated genes,
and due to the functional importance of PRR4 in the saliva,
PRR4 mRNA expression levels were further investigated in the
tumor tissues and adjacent normal tissues from 90 patients with
laryngeal cancer.
| Table II.Identified DEGs in the present
study. |
Table II.
Identified DEGs in the present
study.
| Annealing control
primer | DEG no. | Sequence homology
search | GeneBank accession
no. | Function |
|---|
| 2 | DEG4 | Human DNA sequence
from clone RP11-15N12 on chromosome 6, complete sequence (contains
solute carrier family 22 member 23 isoform a) | AL160398.27 | Transmembrane
protein that transports organic ions across cell membranes |
| 2 | DEG5 | Homo sapiens
tetraspanin 1, mRNA | NM_005727.3 | Transmembrane
protein that regulates cell adhesion, migration, proliferation and
differentiation |
| 3 | DEG7 | Homo sapiens
chromosome 1 clone RP5-1014C4, complete sequence | AC104456.2 |
|
| 4 | DEG9 | Human DNA sequence
from clone RP5-1148A21 on chromosome 6, complete sequence | AL135905.6 |
|
| 8 | DEG10 | Homo sapiens
mitochondrial ribosomal protein L53, mRNA | NM_053050.4 | Component of the
large subunit of the mitochondrial ribosome that is encoded by the
nuclear genome |
| 9 | DEG11 | Human DNA sequence
from clone RP4-668J24 on chromosome 6p25.1–25.3, complete
sequence | AL034346.31 |
|
| 10 | DEG14 | Homo sapiens
ribosomal protein S26, mRNA | NM_001029.3 | Ribosomal protein
that is a component of the 40S subunit |
| 11 | DEG17 | Homo sapiens
chromosome 16 clone RP11-488I20, complete sequence | AC007353.5 |
|
| 12 | DEG20 | Homo sapiens
immunoglobulin heavy constant γ 1 (G1m marker), mRNA (cDNA clone
IMAGE:4851063) | BC018747.1 |
|
| 13 | DEG21 | Homo sapiens
proline-rich 4 (lacrimal), transcript variant 2, mRNA | NM_007244.2 | Secreted protein
which may have protective functions in the eye and the mouth |
| 15 | DEG22 | Homo sapiens
1-acylglycerol-3-phosphate O-acyltransferase 3, transcript variant
1, mRNA | NM_020132.4 | Acyltransferase
that converts lysophosphatidic acid into phosphatidic acid, which
is the second step in the de novo phospholipid biosynthetic
pathway |
| 17 | DEG23 | Homo sapiens
follistatin-like 1, mRNA | NM_007085.4 | Promotes
keratinocyte migration and wound repair |
Confirmation of ACP data by RT-qPCR
for PRR4
In order to confirm the expression pattern of DEG21
(PRR4 gene), its expression levels were analyzed using
RT-qPCR in 90 tumor samples and adjacent non-cancerous tissue
samples. The PRR4 transcript was detected in all tumor and
normal tissue samples except for 3 tumor tissues and 1 normal
tissue. However, PRR4 expression was significantly decreased
in 65 (72.2%) of the 90 tumor samples when compared with the paired
non-cancerous tissue (P<0.001; Table
III). Increased expression was observed in 24 tumor samples
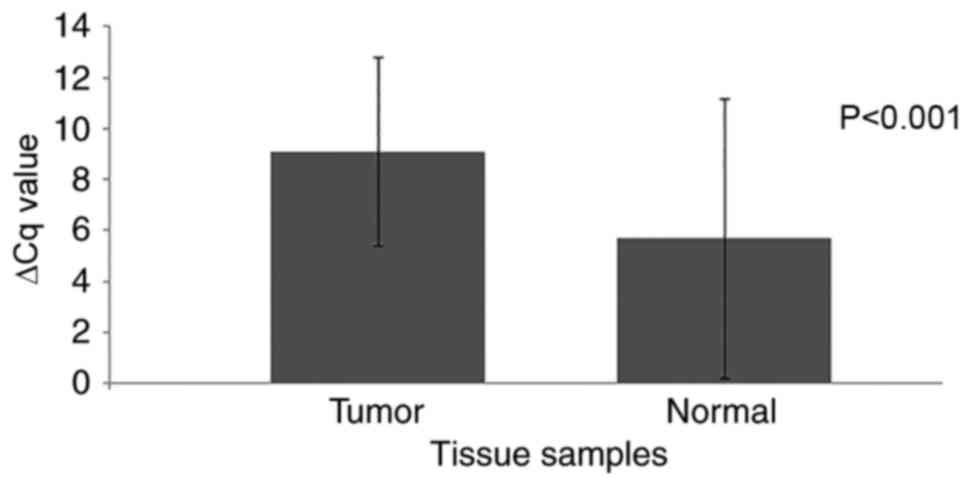
(26.7%), and no change was detected in 1 sample. The mean ΔCq
levels were 9.1±3.7 and 5.7±5.5 for the tumor and the normal tissue
samples, respectively, and a significant difference was identified
(P<0.001; Fig. 1; Table IV). This indicates an 11-fold
decrease in PRR4 expression in the tumor tissues compared
with the non-cancerous tissue. No association was identified with
any clinicopathological characteristic, including clinical stage,
histology, sex, age, histological grade and smoking status
(Table III).
| Table III.Distribution of PRR4 gene
expression and its association with clinicopathological
characteristics of the patients. |
Table III.
Distribution of PRR4 gene
expression and its association with clinicopathological
characteristics of the patients.
|
| PRR4 gene
expression, n (%) |
|
|---|
|
|
|
|
|---|
| Clinicopathological
parameter | Decreased | No change | Increased |
P-valuea |
|---|
| Clinical stage |
|
|
| 0.903 |
| Early
stage (I+II) | 7 (7.8) | 0 (0) | 3 (3.3) |
|
|
Advanced stage (III+IV) | 57 (63.3) | 1
(1.1) | 20 (22.2) |
|
|
Unknown | 1 (1.1) | 0 (0) | 1 (1.1) | NT |
| Histology |
|
|
| NT |
|
SCC | 64 (71.1) | 1
(1.1) | 21 (23.3) |
|
|
Non-SCC | 0 (0) | 0 (0) | 2 (2.2) |
|
|
Unknown | 1 (1.1) | 0 (0) | 1 (1.1) |
|
| Sex |
|
|
| 0.951 |
|
Female | 2 (2.2) | 0 (0) | 1 (1.1) |
|
|
Male | 63 (70) | 1
(1.1) | 23 (25.6) |
|
| Age |
|
|
| 0.5 |
|
≤50 | 8 (8.9) | 0 (0) | 5 (5.6) |
|
|
>50 | 57 (63.3) | 1
(1.1) | 18 (20) |
|
|
Unknown | 0 (0) | 0 (0) | 1 (1.1) | NT |
| Histological
grade |
|
|
| 0.327 |
| Low
grade (1+2) | 31 (34.4) | 0 (0) | 8 (8.9) |
|
| High
grade (3+4) | 30 (33.3) | 1
(1.1) | 14 (15.6) |
|
|
Unknown | 4 (4.4) | 0 (0) | 2 (2.2) | NT |
| Smoking |
|
|
| 0.815 |
|
Smoker | 53 (58.9) | 1
(1.1) | 20 (22.2) |
|
|
Non-smoker | 11 (12.2) | 0 (0) | 3 (3.3) |
|
|
Unknown | 1 (1.1) | 0 (0) | 1 (1.1) | NT |
| Total | 65 (72.2) | 1
(1.1) | 24 (26.7) | 0.001 |
| Table IV.Mean expression levels of the
PRR4 gene in tumor and normal tissues. |
Table IV.
Mean expression levels of the
PRR4 gene in tumor and normal tissues.
| Tissue type | PRR4 Cq
(mean ± SD) | β-actin Cq
(mean ± SD) | ΔCq (mean ±
SD) | ΔΔCq |
2−ΔΔCq | P-value |
|---|
| Tumor |
31.4±4.1 |
22.3±2.5 |
9.1±3.7 | 3.4 | 0.09 | <0.001 |
| Normal |
28.6±5.9 |
22.9±2.6 |
5.7±5.5 | 0 | 1 |
|
Expression of PRR4 in the control
group saliva
PRR4 expression was analyzed in the saliva of
healthy smoker and non-smoker groups, and the expression levels
were compared. The mean ΔCq values were 7.7±1.9 and 7.7±1.7 for the
smoker and the non-smoker groups, respectively (data not shown). No
significant differences were identified between the studied
groups.
Discussion
Cancer may be described as a disease of altered gene
expression. As a result of the up- or downregulation of different
genes, numerous genes are activated or silenced, which alters the
overall activity of the cell and supports tumor development
(24). Therefore, identifying
differentially expressed genes in tumor cells may help in
understanding the molecular mechanisms that underlie the
development and progression of cancer (25,26). To
date, by using different high-throughput technologies including
cDNA arrays and transcriptome analyses, numerous genes
differentially expressed in tumor cells have been identified
(27–29). However, it remains difficult to
determine the genetic events that function in tumorigenesis.
The PCR-based GeneFishing technology is a relatively
novel method that can be used to reveal differences in the gene
expression levels between two or more samples (18). In the present study, 27 differentially
expressed RNAs in laryngeal tumor samples were identified, and 12
of these were characterized by cloning and sequencing. Based on
Gene Ontology, Serial Analysis of Gene Expression and The Cancer
Genome Atlas database queries, no associations between these genes
and HNSCC or laryngeal cancer could be identified. However, the
effect of tetraspanin 1 and follistatin-like 1 in the progression
of other types of cancer have been reported (30,31).
PRR4 mRNA was discovered in 1995 by Dickinson
and Thiesse (5) in the acinar cells
of the human lacrimal gland. Transcriptome studies demonstrated
that PRR4 is highly expressed at the mRNA level in the
submucosal glands, including the parotid gland, and is considered
to be a potential biomarker for indicating the functional
efficiency of the gland (9,10). PRR4 is one of the salivary PRPs,
constituting approximately two-thirds of the proteins secreted by
human parotid glands (11,12). PRPs contain repetitive PRR sequences
or multiple tandem repeats with minor variations between repeated
sequences (32–34). Salivary PRPs have several functions,
but most are likely to serve a protective role by binding to
tannins via a repetitive domain in the epithelial surfaces
(15–17). Tannins are water-soluble polyphenols
that are present in a variety of plant-derived foods. It has been
reported that tannins are mutagenic and carcinogenic compounds;
animal experiments also revealed that the subcutaneous injection of
tannins resulted in tumor formation (35). Due to its function in binding to
pathogens and tannins, the PRR4 gene was selected for
further investigation in the present study.
A number of PRPs additionally function in the
regulation of transcription by binding to transcription factors
(36,37). However, their exact functions remain
unclear. The downregulation of the PRR4 mRNA in tear fluid
has been associated with pathological conditions including DES,
thyroid associated orbitopathy, Sjögren syndrome, blepharitis and
diabetic retinopathy (6–8,38–41). We hypothesized that laryngeal cells
may also express PRR4 mRNA depending on the anatomical
location in the larynx. Areas of the larynx have small glands,
termed the minor salivary glands, which produce mucus and saliva to
lubricate and moisten the area. Therefore, the expression of the
PRR4 gene was used for further validation. As a result of
expression analysis, it was observed that in the majority of tumor
tissues, PRR4 expression was downregulated.
PRR4 is a relatively novel protein and there are few
studies at present investigating the PRR4 gene in cancer.
Zinovyeva et al (42)
identified the expression of 80 genes downregulated in the
esophageal tumor cells compared with normal tissues using
suppression subtractive hybridization. Although one of the
identified genes was PRR4, the study did not select this
gene for further confirmation analysis. However, another PRP that
belonged to the same family (small proline-rich protein 3) was
selected for further analysis and its downregulation was associated
with esophageal squamous cell carcinoma. As a result of the pilot
study CapLC-ESI-Q-TOF, Casado et al (43) reported that PRR4 was the one of the
proteins present in the sputum of a non-smoker study group, whilst
it was absent from the sputum of the chronic obstructive pulmonary
disease group. In the same previous study, the authors demonstrated
that PRR4 was downregulated in healthy smokers. In the present
study, 82.2% of the patients with laryngeal cancer were smokers. As
a result of this, we hypothesized that the downregulation of
PRR4 in the tumor samples may be due to smoking. To test the
potential effect of smoking on PRR4 mRNA expression,
PRR4 mRNA expression levels in the saliva of 25 smokers and
25 non-smoking healthy subjects were investigated. However, no
difference was observed between the PRR4 levels of these
groups. Therefore, the results of the present study indicate that
the expression rate of the PRR4 gene was directly associated
with malignancy in laryngeal tumors. As a novel protein, and due to
its function in clearing bacterial pathogens and binding tannins,
it may be concluded that PRR4 functions in the progression
of laryngeal cancer and HNSCC.
Laryngeal cancer is a tumor of the upper
aerodigestive tract with a low overall survival rate (44). Therefore, there is an urgent need for
the earlier detection of laryngeal cancer and the identification of
theurapeutic target molecules. The results of the present study
suggest that the function of PRR4 in laryngeal tumor
warrants further study.
Acknowledgements
The present study was supported by the Scientific
Research Projects Coordination Unit of Istanbul University (grant
nos. 49005 and 20452).
Glossary
Abbreviations
Abbreviations:
|
HNSCC
|
head and neck squamous cell
carcinoma
|
|
PRR4
|
proline-rich 4
|
|
PRP
|
proline rich protein
|
|
DES
|
dry eye syndrome
|
|
ACP
|
annealing control primer
|
|
DEG
|
differently expressed gene
|
|
LB
|
Luria-Bertani
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
PBS
|
phosphate buffered saline
|
|
GO
|
gene ontology
|
|
SAGE
|
serial analysis of gene expression
|
|
TCGA
|
the cancer genome atlas
|
|
COPD
|
chronic obstructive pulmonary
disease
|
References
|
1
|
Vokes EE, Weichselbaum RR, Lippman SM and
Hong WK: Head and neck cancer. N Engl J Med. 328:184–194. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray I, Brennan P and Boffetta P:
Projections of alcohol- and tobacco-related cancer mortality in
Central Europe. Int J Cancer. 87:122–128. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Edefonti V, Bravi F, Garavello W, La
Vecchia C, Parpinel M, Franceschi S, Dal Maso L, Bosetti C,
Boffetta P, Ferraroni M and Decarli A: Nutrient-based dietary
patterns and laryngeal cancer: Evidence from an explatory analysis.
Cancer Epidemiol Biomarkers Prev. 19:18–27. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Loyo M and Pai S: The molecular genetics
of laryngeal cancer. Otolaryngol Clin North Am. 41:657–672. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dickinson DP and Thiesse M: A major human
lacrimal gland mRNA encodes a new proline-rich protein family
member. Invest Ophthalmol Vis Sci. 36:2020–2031. 1995.PubMed/NCBI
|
|
6
|
Grus FH, Podust VN, Bruns K, Lackner K, Fu
S, Dalmasso EA, Wirthlin A and Pfeiffer N: SELDI-TOF-MS ProteinChip
array profiling of tears from patients with dry eye. Invest
Ophthalmol Vis Sci. 46:863–876. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Matheis N, Okrojek R, Grus FH and Kahaly
GJ: Proteomics of tear fluid in thyroid-associated orbitopathy.
Thyroid. 22:1039–1045. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Csősz É, Boross P, Csutak A, Berta A, Tóth
F, Póliska S, Török Z and Tőzsér J: Quantitative analysis of
proteins in the tear fluid of patients with diabetic retinopathy. J
Proteomics. 75:2196–2204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fischer AJ, Goss KL, Scheetz TE,
Wohlford-Lenane CL, Snyder JM and McCray PB Jr: Differential gene
expression in human conducting airway surface epithelia and
submucosal glands. Am J Respir Cell Mol Biol. 40:189–199. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lamkin MS, Arancillo AA and Oppenheim FG:
Temporal and compositional characteristics of salivary protein
adsorption to hydroxyapatite. J Dent Res. 75:803–808. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Edgar WM: Saliva: Its secretion,
composition and functions. Br Dent J. 172:305–312. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Azen EA, Amberger E, Fisher S, Prakobphol
A and Niece RL: PRB1, PRB2, and PRB4 coded polymorphisms among
human salivary concanavalin-A binding, II-1, and Po proline-rich
proteins. Am J Hum Genet. 58:143–153. 1996.PubMed/NCBI
|
|
13
|
Liu J, Faeder JR and Camacho CJ: Toward a
quantitative theory of intrinsically disordered proteins and their
function. Proc Natl Acad Sci USA. 106:pp. 19819–19823. 2009;
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wright PE and Dyson HJ: Intrinsically
unstructured proteins: Re-assessing the protein structure-function
paradigm. J Mol Biol. 293:321–331. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hagerman AE: Chemistry of Tannin-Protein
ComplexationChemistry and Significance of Condensed Tannins.
Hemingway RW, Karchesy JJ and Branham SJ: Plenum Press; New York:
pp. 323–333. 1989, View Article : Google Scholar
|
|
16
|
Hatton MN, Loomis RE, Levine MJ and Tabak
LA: Masticatory lubrication. The role of carbohydrate in the
lubricating property of a salivary glycoprotein-albumin complex.
Biochem J. 230:817–820. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Loomis RE, Bergey EJ, Levine MJ and Tabak
LA: Circular dichroism and fluorescence spectroscopic analyses of a
proline-rich glycoprotein from human parotid saliva. Int J Pept
Protein Res. 26:621–629. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hwang IT, Kim YJ, Kim SH, Kwak CI, Gu YY
and Chun JY: Annealing control primer system for improving
specificity of PCR amplification. Biotechniques. 35:1180–1184.
2003.PubMed/NCBI
|
|
19
|
Altschul SF, Gish W, Miller W, Myers EW
and Lipman DJ: Basic local alignment search tool. J Mol Biol.
215:403–410. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gene Ontology Consortium: Gene ontology
consortium: Going forward. Nucleic Acids Res. 43(Database Issue):
D1049–D1056. 2015.PubMed/NCBI
|
|
22
|
Lal A, Lash AE, Altschul SF, Velculescu V,
Zhang L, McLendon RE, Marra MA, Prange C, Morin PJ, Polyak K, et
al: A public database for gene expression in human cancers. Canc
Res. 59:5403–5407. 1999.
|
|
23
|
Tomczak K, Czerwinska P and Wiznerowicz M:
The cancer genome atlas (TCGA): An immeasurable source of
knowledge. Contemp Oncol (Pozn). 19:A68–A77. 2015.PubMed/NCBI
|
|
24
|
Boundless: Altered gene expression in
cancer. Boundless Biology. Boundless. 2016, https://www.boundless.com/biology/textbooks/boundless-biology-textbook/gene-expression-16/cancer-and-gene-regulation-118/altered-gene-expression-in-cancer-470-11690/
|
|
25
|
Golub TR, Slonim DK, Tamayo P, Huard C,
Gaasenbeek M, Mesirov JP, Coller H, Loh ML, Downing JR, Caligiuri
MA, et al: Molecular classification of cancer: Class discovery and
class prediction by gene expression monitoring. Science.
286:531–537. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vogelstein B and Kinzler KW: Cancer genes
and the pathways they control. Nat Med. 10:789–799. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Velculescu VE, Zhang L, Vogelstein B and
Kinzler KW: Serial analysis of gene expression. Science.
270:484–487. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
van Baal JW, Milana F, Rygiel AM,
Sondermeijer CM, Spek CA, Bergman JJ, Peppelenbosch MP and
Krishnadath KK: A comparative analysis by SAGE of gene expression
profiles of esophageal adenocarcinoma and esophageal squamous cell
carcinoma. Cell Oncol. 30:63–75. 2008.PubMed/NCBI
|
|
29
|
DeRisi J, Penland L, Brown PO, Bittner ML,
Meltzer PS, Ray M, Chen Y, Su YA and Trent JM: Use of a cDNA
microarray to analyse gene expression patterns in human cancer. Nat
Genet. 14:457–460. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu Z, Luo T, Nie M, Pang T, Zhang X, Shen
X, Ma L, Bi J, Wei G, Fong G and Xue X: TSAPN1 functions as an
oncogene in gastric cancer and is downregulated by miR-573. FEBS
Lett. 589:1988–1994. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ni X, Cao X, Wu Y and Wu J: FSTL1
supresses tumor cell proliferation, invasion and survival in
non-small cell lung cancer. Oncol Rep. 39:13–20. 2018.PubMed/NCBI
|
|
32
|
Boze H, Marlin T, Durand D, Pérez J,
Vernhet A, Canon F, Sarni-Manchado P, Cheynier V and Cabane B:
Proline-rich salivary proteins have extended conformations. Biophys
J. 99:656–665. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bennick A: Structural and genetic aspects
of proline-rich proteins. J Dent Res. 66:457–461. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Williamson MP: The structure and function
of proline-rich regions in proteins. Biochem J. 297:249–260. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stoltz DR: Carcinogenes and mutagenes in
the Environment. 3. Stich HF: CRC Press; Boca Raton, Florida: pp.
751982
|
|
36
|
Mermod N, O'Neill EA, Kelly TJ and Tjian
R: The proline-rich transcriptional activator of CTF/NF-I is
distinct from the replication and DNA binding domain. Cell.
58:741–753. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gessler M, Poustka A, Cavenee W, Neve RL,
Orkin SH and Bruns GA: Homozygous deletion in Wilms tumours of a
zinc-finger gene identified by chromosome jumping. Nature.
343:774–778. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Aluru SV, Agarwal S, Srinivasan B, Iyer
GK, Rajappa SM, Tatu U, Padmanabhan P, Subramanian N and
Narayanasamy A: Lacrimal proline-rich 4 (LPRR4) protein in the tear
fluid is a potential biomarker of dry eye syndrome. PLoS One.
7:e519792012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Perumal N, Funke S, Pfeiffer N and Grus
FH: Characterization of lacrimal proline-rich protein 4 (PRR4) in
human tear proteome. Proteomics. 14:1698–1709. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tsai PS, Evans JE, Green KM, Sullivan RM,
Schaumberg DA, Richards SM, Dana MR and Sullivan DA: Proteomic
analysis of human meibomian gland secretions. Br J Ophthalmol.
90:372–377. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Koo BS, Lee DY, Ha HS, Kim JC and Kim CW:
Comparative analysis of the tear protein expression in blepharitis
patients using two-dimensional electrophoresis. J Proteome Res.
4:719–724. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zinovyeva MV, Monastyrskaya GS, Kopantzev
EP, Vinogradova TV, Kostina MB, Sass AV, Filyukova OB, Uspenskaya
NY, Sukhikh GT and Sverdlov ED: Identification of some human genes
oppositely regulated during esophageal squamous cell carcinoma
formation and human embryonic esophagus development. Dis Esophagus.
23:260–270. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Casado B, Iadarola P, Pannell LK, Luisetti
M, Corsico A, Ansaldo E, Ferrarotti I, Boschetto P and Baraniuk JN:
Protein expression in sputum of smokers and chronic obstructive
pulmonary disease patients: A pilot study by CapLC-ESI-Q-TOF. J
Proteome Res. 6:4615–4623. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ferlay J, Steliarova-Foucher E,
Lortet-Tieulent J, Rosso S, Coebergh JW, Comber H, Forman D and
Bray F: Cancer incidence and mortality patterns in Europe:
Estimates for 40 countries in 2012. Eur J Cancer. 49:1374–1403.
2013. View Article : Google Scholar : PubMed/NCBI
|