Introduction
Colorectal cancer (CRC) was the third leading cause
of cancer-associated mortality worldwide in 2015, despite the
advancements in the diagnosis and treatment (1). A total of ~50% of patients with colon
cancer will develop liver metastasis and the 5-year survival rate
for metastatic colon cancer is only 10–15% in the United States of
America (2,3). Metastasis is a major cause of mortality
in patients with colon cancer and is considered incurable due to a
lack of effective therapy (4). To
identify the novel melecules serving key roles in colon cancer, a
number of studies focused on determining the microRNA (miRNA/miR)
associated with this type of cancer (5–7).
Forkhead box O3 (FOXO3) is one of the most
comprehensively characterized members of the FOXO family of
transcription factors. It is a tumor suppressor, and is a potent
transcriptional activator which triggers the expression of a
program of genes involved in cell cycle arrest, DNA repair, hypoxia
and apoptosis (8,9). It has been identified that the FOXO3
transcription factor serves as a target of anti-tumor drugs in
different types of cancer, including breast cancer, chronic myeloid
leukemia and colon cancer (10,11). It
also affects the sensitivity of colon cancer cells to cisplatin
(11). FOXO3 activity may be
regulated by post-translational modifications, including
phosphorylation (12), but the
molecular mechanisms regulating the expression of the FOXO3
expression remain unclear.
miRs are naturally-occurring endogenous
single-stranded RNA usually measuring 18–24 nucleotides in length
(13). A total of >700 miRs have
been identified in the human genome. A function of microRNA is the
downregulation of target proteins expression by mRNA cleavage and
decay or translational repression (14). miRs have been implicated in the
regulation of gene expression essential for organ development,
cellular differentiation, homeostasis and those involved in tumor
occurance (15–17). miRNA expression is often dysregulated
in cancer tissues and they may function either as tumor suppressors
or oncogenes (oncomiRs) (18).
miR-155 has been demonstrated to be an oncogenic miRNA in certain
tumors, including liposarcoma and leukaemia (19–21).
miR-155 transgenic mice have also been demonstrated to develop
acute lymphocytic lymphoma or leukaemia (22). miR-155 is upregulated in gastric
cancer and serves as a prognosis biomarker in these patients
(23). Increased miR-155 expression
is involved in poor prognosis in pancreatic and lung cancers
(24,25), while in breast cancer miR-155
contributes to the progression of invasion (26,27). There
are numerous target genes of miR-155, including inositol
polyphosphate-5-phosphatase D and CCAAT/enhancer binding protein β,
suppressor of cytokine signaling 1 and FOXO3 (28). However, the function of miR-155 in
colon cancer is not understood, although it has been identified
that miR-155 is upregulated in colon cancer (29,30). It
remains unknown if miR-155 targets FOXO3 to effect colon cancer
cell viability.
In the present study, clinical tissues from patients
with colon cancer were collected and miR-155 and FOXO3 expression
levels were detected. It was identified that the expression levels
of miR-155 and FOXO3 were increased and decreased, respectively, in
colon cancer tissues and human cell lines. miR-155 was transfected
into colon carcinoma HT29 and SW620 cell lines, and it was
determined that FOXO3 expression was decreased at the protein level
in a dose-dependent manner, suggesting that they are negatively
associated. The sensitivity of colon carcinoma cells to
chemotherapy drugs cisplatin and paclitaxel was detected, and it
was demonstrated that miR-155 increased chemoresistance. The
present study suggests a novel pathway and target for colon cancer
therapy.
Materials and methods
Cell lines and reagents
Human colon carcinoma HT29 and SW620 cell lines were
purchased from The Cell Bank of Type Culture Collection of Chinese
Academy of Sciences (Shanghai, China) and were cultured in
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (Zhejiang Tianhang Biotechnology Co. Ltd, China), 100
units/ml penicillin and 100 g/ml streptomycin at 37°C in a
humidified chamber with 5% CO2. Samples from patients
with colon cancer and para-carcinoma tissues were collected by
radical colon resection from the People's Hospital of Rizhao City
(Rizhao, China). The present study was approved by the Ethical
Committee of the People's Hospital of Rizhao (Rizhao, China), all
patients were informed and written informed consent was obtained.
Cisplatin was purchased from Sigma-Aldrich; Merck KGaA (Darmstadt,
Germany). miR-155-5p mimic (C-3006 47-05-0005) and scrambled
control oligonucleotide (CN-001000-01-05; sequence,
5′-CCCUAUCACGAUUAGCAUUAAUU-3′) were purchased from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). The CellTiter 96®
Aqueous Non-Radioactive Cell Proliferation Assay kit was purchased
from Promega Corporation (Madison, WI, USA). All the PCR primers
were synthetized by Invitrogen (Thermo Fisher Scientific,
Inc.).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated using TRIzol®
(Life Technologies; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. For mRNA detection, cDNA for PCR was
obtained by RT-PCR following the manufacturer's protocol of a
Toyobo SYBR-Green RT-PCR Master Mix kit (Toyobo Life Science,
Osaka, Japan). cDNA was subjected to qPCR using the SYBR-Green PCR
reagents kit (Applied Biosystems; Thermo Fisher Scientific, Inc.)
on a ABI StepOnePlus (ABI) instrument (Thermo Fisher Scientific,
Inc.). For miR-155 detection, polyAs were added into RNA using
Escherichia coli polyA polymerase (Fermentas; Thermo Fisher
Scientific, Inc., Pittsburgh, PA, USA). A total of 2 µg tailed
total RNA was reverse transcribed with miR reverse transcriptase
primer (1 µM; Shanghai Genepharma Co., Ltd., Shanghai, China). qPCR
was performed with miR-155 forward (F) and reverse (R) primers, and
the thermocycler conditions were as follows: 95°C for 10 min; then
41 cycles at 95°C for 10 sec, 60°C for 30 sec and 72°C for 30 sec.
All primers used for qPCR analysis were synthesized by Invitrogen
(Thermo Fisher Scientific, Inc.), as follows: FoxO3 F,
AGTGGATGGTGCGCTGTGT; FoxO3 R, CTGTGCAGGGACAGGTTGT (31); GAPDH F, TGTGTCCGTCGTGGATCTGA; GAPDH R,
TTGCTGTTGAAGTCGCAGGAG; miR reverse transcriptase primer,
GCTGTCAACGATACGCTACGTAACGGCATGACAGTGTTTTTTTTTTTTTTTTTTTTTTTTN;
miR-155 F, TTAATGCTAATCGTGATAGGGGT; miR-155 R,
GCTGTCAACGATACGCTACGTAACG (20); U6
F, CTCGCTTCGGCAGCACA; and U6 R, AACGCTTCACGAATTTGCGT. The relative
amount of target mRNA was determined using the comparative
threshold (Cq) method by normalizing target mRNA Cq values to those
of GAPDH or U6 (32).
Plasmid construction and
stable/transient transfection of miR-155
To construct the human FOXO3 recombined plasmid, the
FOXO3 gene (NCBI Reference Sequence: NM_001455.3) was cloned into
pCMV-tag2a vector (Agilent Technologies; Thermo Fisher Scientific,
Inc.). A human genomic fragment of 65 bp containing the miR-155
precursor DNA sequence (NCBI Reference Sequence: NR_030784.1) was
cloned into the pcDNA3.1(−)-myc-his vector (Invitrogen; Thermo
Fisher Scientific, Inc.). The recombinant plasmid was
pcDNA3.1-miR-155. The primers for construction were as follows;
underlined nucleotides represent BamH I and HindIII sites: miR-155
P1,
GATCCCTGTTAATGCTAATCGTGATAGGGGTTTTTGCCTCCAACTGACTCCTACATATTAGCATTAACAGA;
miR-155 P2,
AGCTTCTGTTAATGCTAATATGTAGGAGTCAGTTGGAGGCAAAAACCCCTATCACGATTAGCATTAACAGG;
FOXO3-P1, ATTAGGATCCATGGCAGAGGCACCGGCTTC; and FOXO3-P2,
GCAAAAGCTTTCCTGGCACCCAGCTCTGAG. To generate a cell line stably
expressing miR-155, HT29 and SW620 cells were transfected with 200
ng pcDNA3.1-miR-155 using Lipofectamine® 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Following 48 h
transfection, following 800 mg/ml G418 selection, the single clone
that over-expressed miR-155 was identified. For miR155 transient
transfection, miR-155 mimics (Invitrogen; Thermo Fisher Scientific,
Inc.) were used to transfect the 2 cell lines.
Luciferase assays
The online database search TargetScanHuman 6.2
(http://www.targetscan.org/), microRNA
(http://www.microrna.org) and miRTarBase
(http://mirtarbase.mbc.nctu.edu.tw/php/search.php)
were used to predict a potential target of miR-155 (Date of access,
January 2016). The wild type (WT) or mutant 3′-UTR of FOXO3
containing the putative miR-155 binding sites was synthesized and
inserted into pmirGLO Dual-Luciferase miRNA Target Expression
Vector (Promega Corporation) to generate the recombinant
constructs, pmirGLO-FOXO3-3′UTR-WT and pmirGLO-FOXO3-3′UTR-M,
respectively. miR-155 mimics or miR-inhibitor using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and incubated for 48 h. The primers for 3′UTR WT
and mutant were as follow; FOXO3-3′UTR-WT P1,
AAACTCTTTGCATAAAAAGCATTAGGCATAT; FOXO3-3′UTR-WT P2,
CTAGATATGCCTAATGCTTTTTATGCAAAGAGTTT; FOXO3-3′UTR-M P1,
AAACTCTTTGCATAAAAACGATAAGGCATAT; and FOXO3-3′UTR-M P2,
CTAGATATGCCTUAUGGTTTTTATGCAAAGAGTTT. Underlined nucleotides
represent PmeI and XbalI sites, respectively. Italicized
nucleotides represent the miR-155 target sequence. Bold italicized
nucleotides represent the sites of mutation. Cells were harvested
48 h following transfection for the measurement of luciferase
activity with a Dual-Luciferase Reporter Assay system (Promega
Corporation) according to manufacturer's protocol. Renilla
luciferase activity was applied as the method of normalization for
firefly luciferase activity.
Cell proliferation assays
The effect of miR-155-5p on cisplatin chemotherapy
sensitivity of colon cancer cell lines was detected using Cell
Titer 96® Aqueous Non-Radioactive Cell Proliferation
Assay kit (Promega Corporation) according to manufacturer's
protocol. Cells grown in regular media were plated on 96-well
plates (5,000 cells/well) were treated with cisplatin (0, 10, 50,
100 and 200 µM) at 37°C in a 5% CO2 humidified
atmosphere for 48 h. Subsequently, 20 µl combined MTS/phenazine
methosulfate solution was pipetted into each well and incubated for
4 h at 37°C. A total of 25 µl 10% SDS (Boster Biological
Thecnology, Pleasanton, CA, USA) was added to each well to stop the
reaction. Absorbance was detected at 490 nm using a 96-well plate
reader.
Western blot analysis
Tissues from the CT-26 tumors (2×106
cells/mouse; purchased from The Cell Bank of Type Culture
Collection of Chinese Academy of Sciences, Shanghai, China) were
collected and lysed cells by lysis buffer (Tris 20 mM, NaCl 150 mM,
1% Triton X-100 and 1% cocktail protease inhibitors). All of the
protein extraction process was carried out according to the product
specifiation. The BCA protein assay kit (Beyotime Institute of
Biotechnology, Haimen, China) was applied to detected the protein
concentration. Then total protein (30 µg) were separated by
electrophoresis on a 12% SDS-PAGE (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) and transferred onto nitrocellulose membranes
(Merck KGaA). The membranes were blocked in 5 % non-fat milk
solution for 2–4 h at 4°C, and then washed twice with TBST solution
(0.1% Tween-20). Membranes were incubated overnight at 4°C with the
following antibodies. FOXO3 polyclonal antibodies (1:1,000; Abcam,
Cambridge, UK; cat. no. ab23683) or β-actin (1:3,000, cat. no.
BM0626, Specificity: monoclonal mouse; Boster Biological
Technology, USA) monocolonal antibodies were used as primary
antibodies, followed by goat anti-rabbit (cat. no. A21020;
dilution: 1:1,000; AmyJet Scientific, Wuhan, China) or goat
anti-mouse secondary antibodies (cat. no. A21010; dilution:
1:1,000) conjugated with horseradish peroxidase (ProteinTech
Groups, Inc., Chicago, IL, USA) incubated for 2 h at 4°C. The
protein analysis was performed using Millipore ECL Western Blotting
Substrate on a UVP ChemiDoc-It imaging system (Chemidoc-IT 510;
Visionworks LS, UVP, LLC, USA).
Caspase 3 activity detection
Colon cancer cells treated with or without cisplatin
were were seeded at a density of 2×105 cells/well in a
6-well plate with 2 ml culture medium. Following 24 h incubation,
cells were treated with 200 µM cisplatin to induce apoptosis.
Casapse-3 activity was measured using a Caspase 3 Fluorometric
activity assay kit (cat no. C1115; Beyotime Institute of
Biotechnology) following the manufacturer's protocol.
Flow cytometry
Colon cancer cells treated with or without cisplatin
were harvested by 500 µl 0.25% trypsin and then centrifugation 5
min at room temperature (241.5 × g). A total of 500 µl binding
buffer (including precooled 70% ethanol and 0.5 mmol/l EDTA) was
added to each tube and incubated overnight at 4°C, then
intracellularly stained with phycoerythrin-anti human Ki67 mAb (cat
no. 350503; dilution, 1:20; BioLegend, Inc., San Diego, CA, USA) in
staining buffer (PBS containing 0.5% BSA) for 30 min at room
temperature in the dark. Samples were washed twice with PBS, and
analyzed by flow cytometry using a BD C6 auri flow cytometry and
Expo 32-ADC v. 1.2B software (both BD Biosciences, Franklin Lakes,
NJ, USA).
Animal model
Pathogen-free 10 weeks old male WT C57BL/6 mice
(mean weight:24.26±3.04 g, n=7) and pathogen-free 10-week-old male
miR-155 knockout mice (miR-155-/-) (mean weight, 24.75±2.93 g; n=6)
were obtained from Jackson Laboratory (Ben Harbour, ME, USA). The
animals were housed under standard conditions (18–29°C, 0.03%) in a
12 h light/dark cycle with access to water and food (ad libitum).
The mouse colon cancer CT26 cell line (2×106 cells per
mouse) was inoculated into the backs of the mice by hypodermic
inoculation (matrix glue was administered to the mice in order to
prevent the spread of the tumor, and multiple tumors were not
observed). A total of 2 days following tumor cell inoculation, mice
were treated with cisplatin (300 µM) every 2 days for 10 days by
intraperitoneal injection. The tumor sizes were measured every 3
days using calipers from 1st day of drug treatment to the 30th day,
and tumor volume was calculated as: Tumor volume=[length (a) ×
width (b)2)/2 (33). The
data are presented as the mean volume ± standard deviation (SD).
The Ethical Committee of the People's Hospital of Rizhao approved
all animal experimental procedures.
Statistical analysis
SPSS software (version 16; SPSS, Inc., Chicago, IL,
USA) was used for statistical analysis. All data were calculated as
the mean ± standard deviation. A one-way analysis of variance and
Student Newman-Keuls post-hoc test were used to analyze the results
between treated and control groups, and an unpaired two-tailed
Student's t-test was used to compare two groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-155 is negatively associated with
FOXO3 in colon cancer tissues
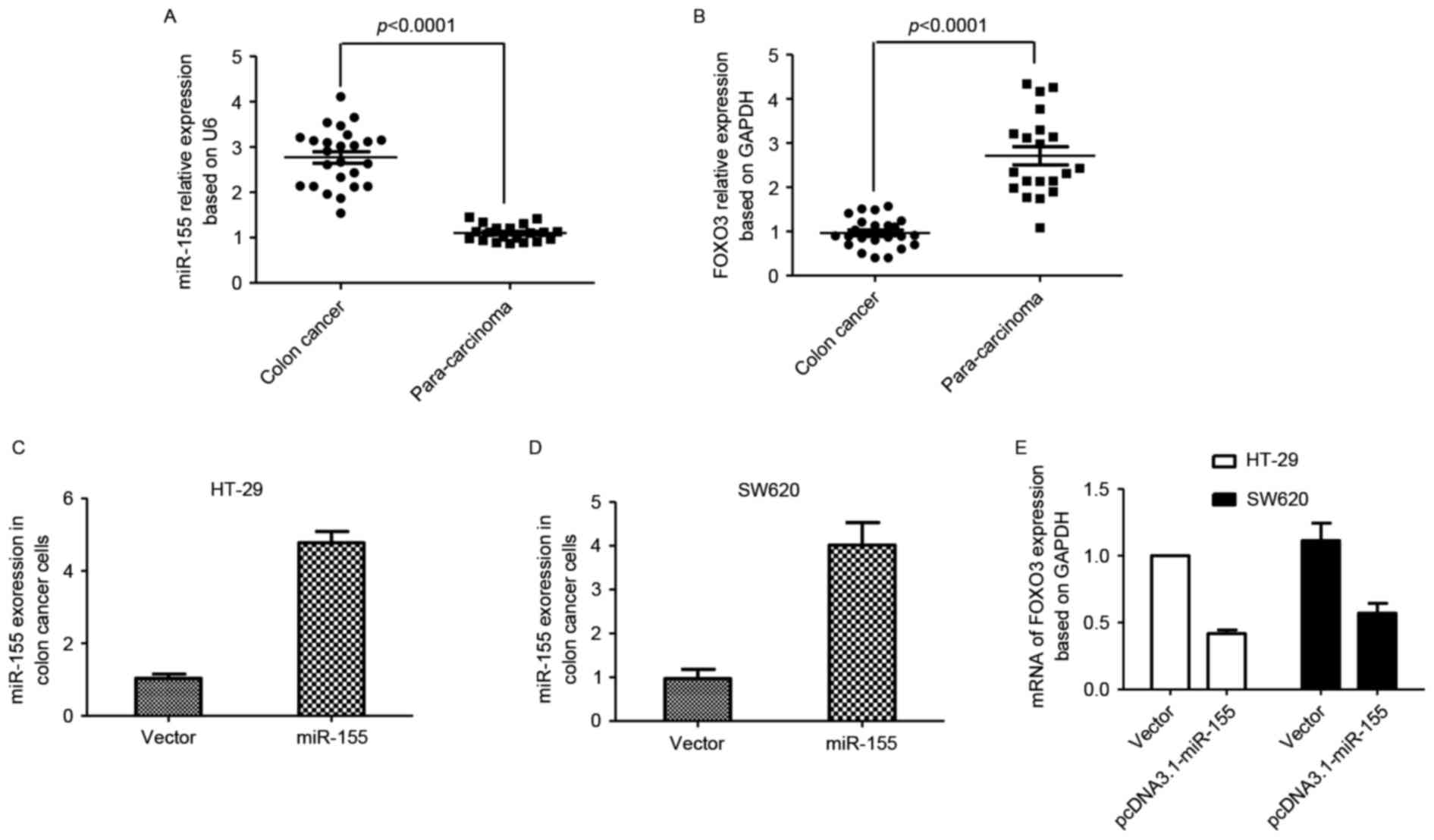
To investigate miR-155 expression in colon cancer
and para-carcinoma tissues, samples (colon cancer, n=25;
para-carcinoma, n=20) from Rizhao People's Hospital were collected.
Total RNA of tissues was extracted, and qPCR was performed. The
results indicated that miR-155 expression was increased in colon
cancer tissues compared with para-carcinoma tissues, while FOXO3
expression was decreased in colon cancer tissues compared with
para-carcinoma tissues (Fig. 1A and
B). The plasmid pcDNA3.1-miR-155 was transfected into colon
cancer HT29 and SW620 cell lines, and the mRNA expression of FOXO3
was also detected. FOXO3 mRNA levels decreased following miR-155
enhancement compared with the control group (Fig. 1C-E).
FOXO3 is a direct target of miR-155 in
colon cancer cells
Based on an online database search for targets of
miRNAs that were identified in previous studies (34,35), it
was suggested that FOXO3 is a potential target of miR-155. Then,
whether miR-155 mediated FOXO3 was determined. pmirGLO-UTR WT or M
plasmids were constructed and transfected into HT-29 cells. It was
demonstrated that reduced firefly luciferase expression indicated
the binding of miR-155 to the cloned miRNA target sequence, and the
mutant UTR group was not altered (Fig.
2A). This suggested that miR-155 may bind to position 1497–1503
of FOXO3 3′UTR and FOXO3 be directly targeted by miR-155 in human
colon cancer cells (Fig. 2B).
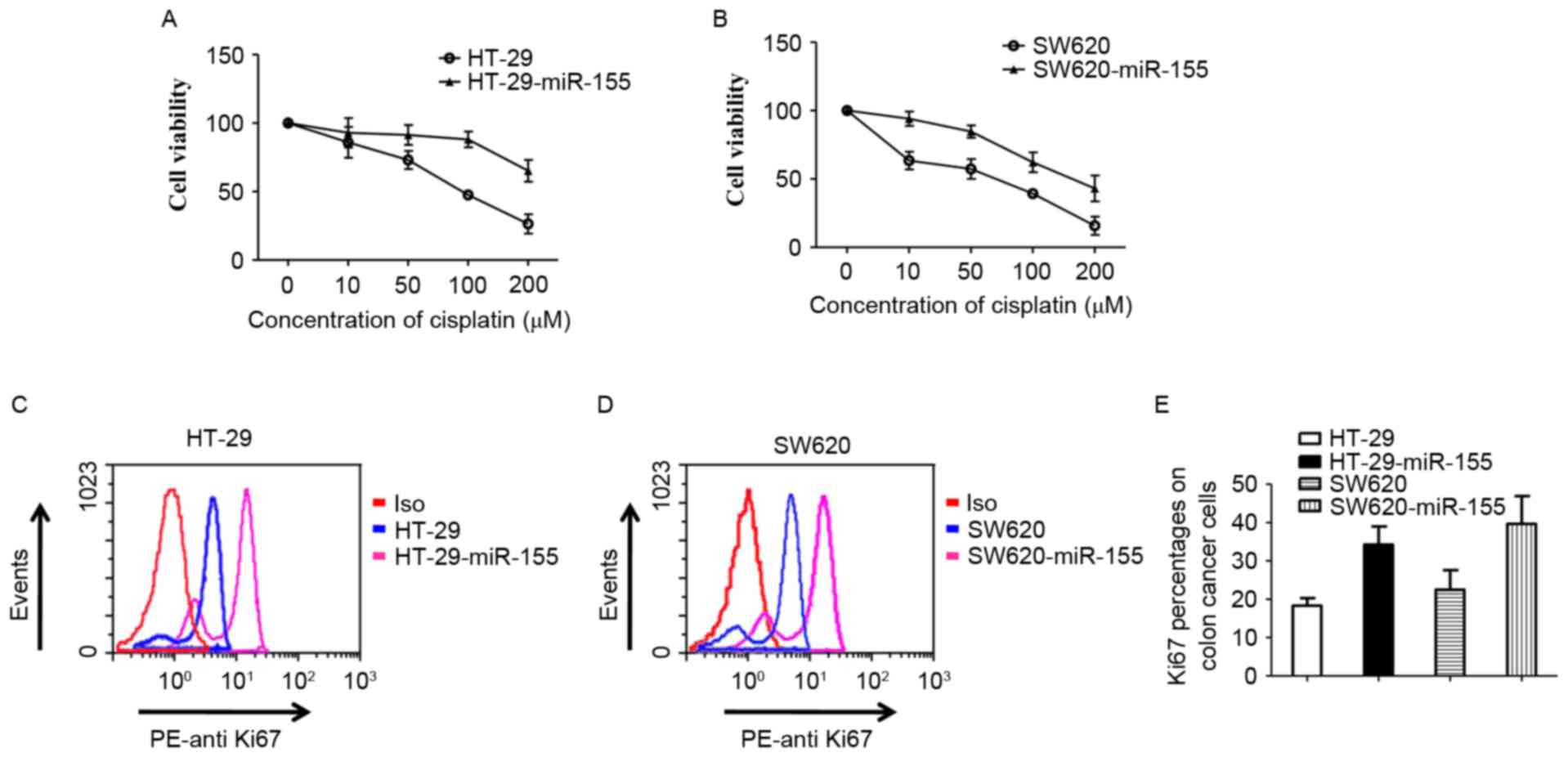
miR-155 decreases chemosensitivity to
cisplatin in colon cancer cells
Different concentrations of cisplatin (0, 10, 50,
100 and 200 µM) were used to treat HT29 and SW620 cells for 48 h.
The viability of cells was detected using a Non-Radioactive Cell
Proliferation Assay kit. HT29 cells demonstrated increased
sensitivity compared with HT29-miR-155 cells, which stabley
expressed high levels of miR-155 (Fig.
3A). The same result was exhibited in SW620 cells (Fig. 3B). Antigen Ki-67 is a nuclear protein
that is associated with cellular proliferation and is a cellular
marker for proliferation (36). Ki67
was detected in HT29, SW620, HT29-miR-155 and SW620-miR-155 cells
by flow cytometry. The results indicated that ki67 expression was
increased in miR-155 high-expressed cells compared control cells
(Fig. 3C, D and E). These results
suggests that the miR-155-mediated decrease of chemosensitivity to
cisplatin in colon cancer cells may occur through targeting
FOXO3.
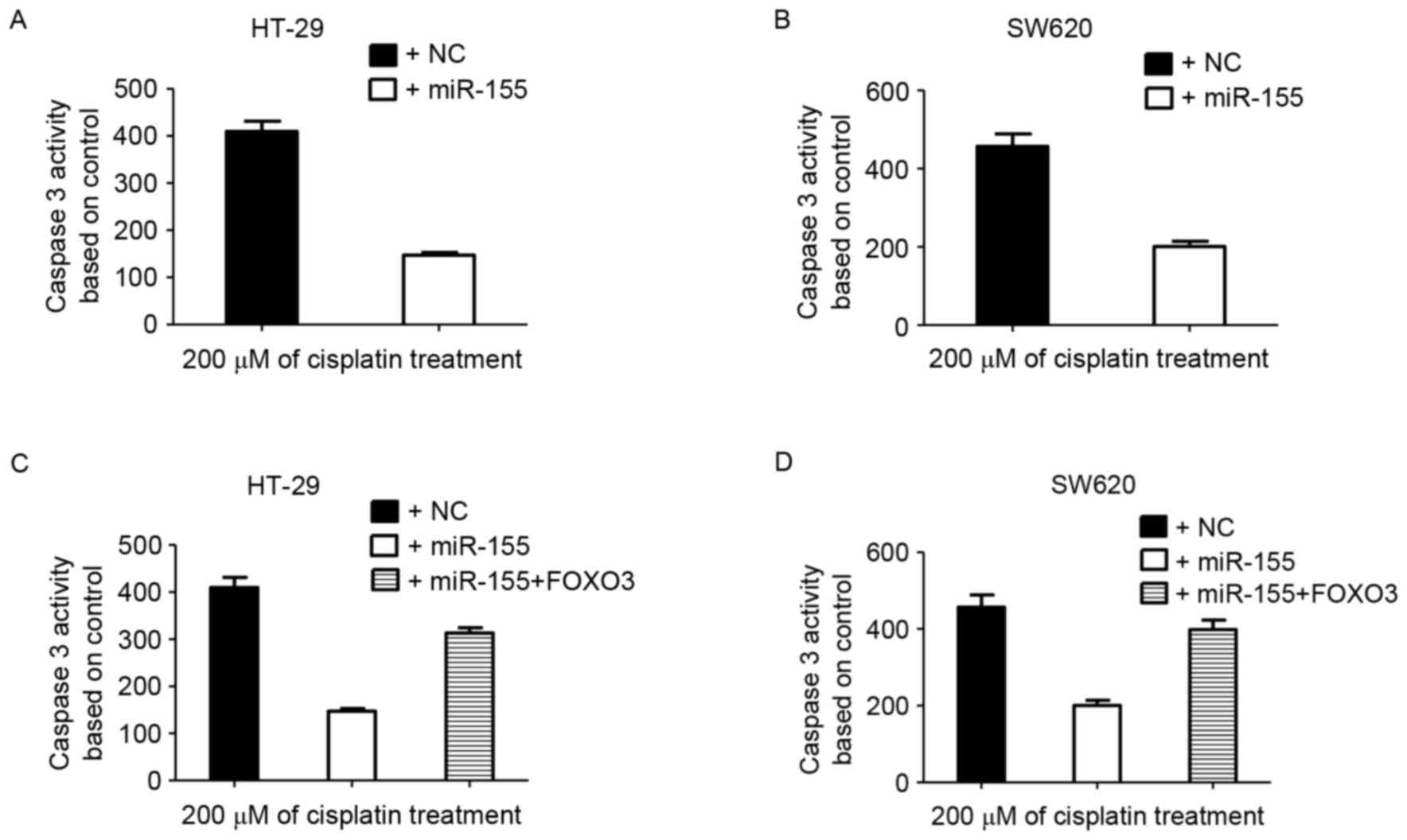
miR-155 blocks the caspase 3 activity
induced by cisplatin
Cisplatin may induce apoptosis in HT-29 and SW620
cells (37–39). miR-155 mimics were transfected into
these two cell lines to obtain HT-29-miR-155 and SW620-miR-155
cells. Colon cancer cells were treated with cisplatin for 48 h and
harvested for caspase 3 activity detection. High miR-155 expression
decreased the caspase 3 activity induced by cisplatin in HT-29 and
SW620 cells (Fig. 4A and B).
FOXO transcription factors have been indicated to
regulate apoptosis and cell cycle-associated genes (40–42). When
FOXO3 and miR-155 were co-transfected into colon cancer cells, it
was demonstrated that cisplatin efficiently induced caspase 3
activity (Fig. 4C and D). These data
suggested that miR-155 decreased colon cancer sensitivity to
cisplatin by targeting FOXO3.
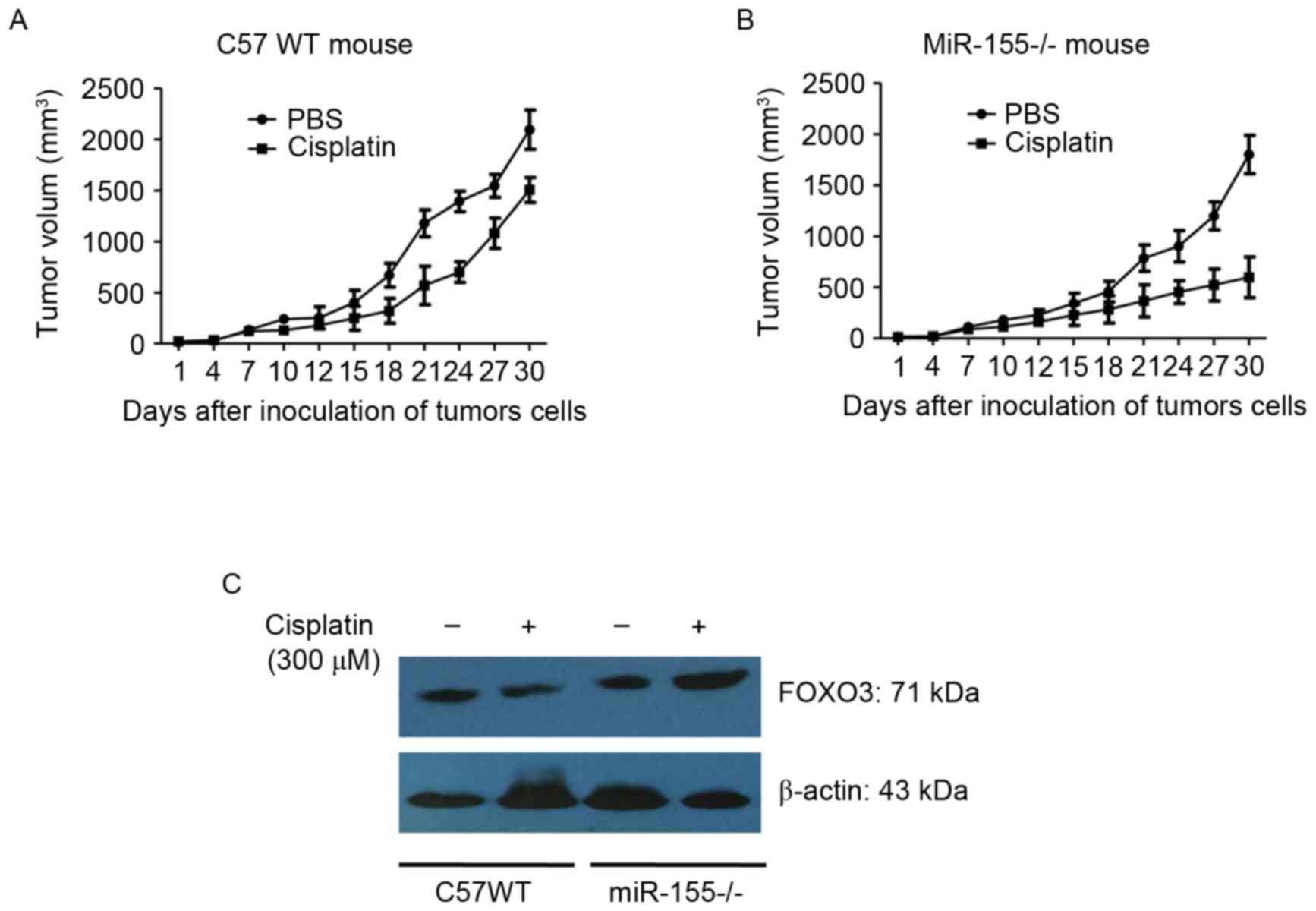
miR-155 promotes colon cancer growth
in mice
C57BL/6 (WT) and mir-155 knockout mice (miR-155-/-)
were divided into 4 groups at random. CT26 cells were inoculated
into the backs of the mice. The tumor-bearing mice were treated
with 300 µM cisplatin for 10 days. Tumor length (a) and width (b)
were measured using vernier calipers. Tumor volume was calculated
using the formula: V (mm3)=(a × b2)/2.
The results showed that: the tumor weight was
(0.7184±0.1547) g and (0.6728±0.1415) g in the control group of C57
WT mouse and miR-155-/- mouse, respectively, and the tumor weight
was (0.5649±0.1252) g and (0.4937±0.1096) g in the cisplatin group
of C57 WT mouse and miR-155-/- mouse, respectively. It was observed
that tumor growth was significantly reduced in the miR-155-/- mice
compared with the WT mice (Fig. 5A and
B). It was also identified that FOXO3 expression in tumor
tissues of miR-155-/- was increased compared with that of the WT
mice (Fig. 5C). The data additionally
suggested that miR-155 enhanced tumor growth and chemoresistance by
targeting the FOXO3 gene.
Discussion
Although the mechanism by miR-155 regulation is not
well understood, it has been suggested that the oncogenic microRNA
miR-155 is enhanced in numerous types of cancer (43–46).
However, miR-155 expression in colon cancer has not been
identified. In the present study, to the best of our knowledge, the
effect of miR-155 targeting FOXO3 gene in colon cancer cells was
demonstrated for the first time. Tissues from patients with colon
cancer were collected for miR-155 detection. The results indicated
that miR-155 was increased in the colon cancer tissues, but not in
para-carcinoma tissues. This result was in accordance with similar
previous studies that have detected miR-155 expression in colon
cancer (29,30).
As a prominent oncomiR, miR-155 may target the
mismatch repair genes MutL Homolog 1, MutS Homolog (MSH) 2 and MSH6
and contribute to the microsatellite instability (MSI) phenotype in
colon cancer (47). It downregulates
B-cell lymphoma (Bcl)-6 and causes V-Myc avian myelocytomatosis
viral oncogene homolog and Cyclin D1 upregulation, which promotes
cell proliferation (48). In renal
cancer cells, miR-155 may function as an oncogene by targeting BTB
domain and CNC homolog (49). In the
present study, miR-155 targets were predicted using TargetScanHuman
6.2, and it was identified that FOXO3 3′UTR was able to bind with
miR-155. This prediction was associated with the results revealed
by Zhang et al (50). In the
present study, FOXO3 expression in colon cancer was negatively
associated with miR-155 expression. To additionally demonstrate the
interaction between FOXO3 and miR-155, a recombinant plasmid
pmirGLO-FOXO3-3′UTR-WT expressing the FOXO3 3′UTR wild type and a
pmirGLO-FOXO3-3′UTR-M plasmid with the FOXO3 3′UTR with a small
number of mutant bases, were constructed. Then, miR-155 mimics and
inhibitors were co-transfected into HT-29 cells and luciferase
reporter assays demonstrated that miR-155 directly bound to the
FOXO3 3′UTR.
The functions of miR-155 in various types of cancer
are different: It has been suggested that miR-155 is associated
with the development of liver, leukemia, breast, lung and stomach
tumors (51–53). Conversely, miR-155 may serve to
prevent cancer in transgenic mice by promoting proper immune
function. To clarify the function of miR-155 in colon cancer, the
effect of miR-155 on chemoresistance was evaluated in the present
study. Cisplatin was used to treat HT-29, SW620, HT-29-miR-155 and
SW620-miR-155 cells. miR-155 increased the resistance of colon
cancer cells to cisplatin. This result is consistent with results
from Yu et al (54). However,
the novel observation of the present study was that in colon cancer
cells, miR-155 increased chemoresistance by targeting FOXO3. In the
miR-155-/- mice, CT26 cell growth was inhibited, and these cells
demonstrated an increased sensitivity to cisplatin compared with
the WT mice.
To conclude, the present study identified that
miR-155 may promote colon cancer growth and increase colon cancer
cells chemoresistance to cisplatin by directly targeting FOXO3.
This suggests a novel pathway for the treatment and cure of colon
cancer in the future.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Valeri N, Gasparini P, Fabbri M, Braconi
C, Veronese A, Lovat F, Adair B, Vannini I, Fanini F, Bottoni A, et
al: Modulation of mismatch repair and genomic stability by miR-155.
Proc Natl Acad Sci USA. 107:pp. 6982–6987. 2010; View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Geng L, Chaudhuri A, Talmon G, Wisecarver
JL, Are C..Brattain M and Wang J: MicroRNA-192 suppresses liver
metastasis of colon cancer. Oncogene. 33:5332–5340. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Eker B, Ozaslan E, Karaca H, Berk V,
Bozkurt O, Inanc M, Duran AO and Ozkan M: Factors affecting
prognosis in metastatic colorectal cancer patients. Asian Pac J
Cancer Prev. 16:3015–3021. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ozawa T, Kandimalla R, Gao F, Nozawa H,
Hata K, Nagata H, Okada S, Izumi D, Baba H, Fleshman J, et al: A
microrna signature associated with metastasis of T1 colorectal
tumors to lymph nodes. Gastroenterology. Nov 30–2017. View Article : Google Scholar
|
|
6
|
Bu P, Wang L, Chen KY, Srinivasan T,
Murthy PK, Tung KL, Varanko AK, Chen HJ, Ai Y, King S, et al: A
miR-34a-numb feedforward loop triggered by inflammation regulates
asymmetric stem cell division in intestine and colon cancer. Cell
Stem Cell. 18:189–202. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hur K, Toiyama Y, Okugawa Y, Ide S, Imaoka
H, Boland CR and Goel A: Circulating microRNA-203 predicts
prognosis and metastasis in human colorectal cancer. Gut.
66:654–665. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang L, Cai M, Gong Z, Zhang B, Li Y,
Guan L, Hou X, Li Q, Liu G, Xue Z, et al: Geminin facilitates FoxO3
deacetylation to promote breast cancer cell metastasis. J Clin
Invest. 127:2159–2175. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guan L, Zhang L, Gong Z, Hou X, Xu Y, Feng
X, Wang H and You H: FoxO3 inactivation promotes human
cholangiocarcinoma tumorigenesis and chemoresistance through
Keap1-Nrf2 signaling. Hepatology. 63:1914–1927. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Essafi A, Fernández de Mattos S, Hassen
YA, Soeiro I, Mufti GJ, Thomas NS, Medema RH and Lam EW: Direct
transcriptional regulation of Bim by FoxO3a mediates STI571-induced
apoptosis in Bcr-Abl-expressing cells. Oncogene. 24:2317–2329.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fernández de Mattos S, Villalonga P,
Clardy J and Lam EW: FOXO 3a mediates the cytotoxic effects of
cisplatin in colon cancer cells. Mol Cancer Ther. 7:3237–3246.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tsai KL, Sun YJ, Huang CY, Yang JY, Hung
MC and Hsiao CD: Crystal structure of the human FOXO3a-DBD/DNA
complex suggests the effects of post-translational modification.
Nucleic Acids Res. 35:6984–6994. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hammond SM: RNAi, microRNAs, and human
disease. Cancer Chemother Pharmacol. 58 Suppl 1:S63–S68. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim VN: MicroRNA biogenesis: Coordinated
cropping and dicing. Nat Rev Mol Cell Biol. 6:376–385. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mohammadi A, Mansoori B and Baradaran B:
The role of microRNAs in colorectal cancer. Biomed Pharmacother.
84:705–713. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Strubberg AM and Madison BB: MicroRNAs in
the etiology of colorectal cancer: Pathways and clinical
implications. Dis Model Mech. 10:197–214. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nagaraju GP, Madanraj AS, Aliya S, Rajitha
B, Alese OB, Kariali E, Alam A and El-Rayes BF: MicroRNAs as
biomarkers and prospective therapeutic targets in colon and
pancreatic cancers. Tumour Biol. 37:97–104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang P, Bill K, Liu J, Young E, Peng T,
Bolshakov S, Hoffman A, Song Y, Demicco EG, Terrada DL, et al:
miR-155 is a liposarcoma oncogene that targets casein kinase-1α and
enhancees β-catenin signaling. Cancer Res. 72:1751–1762. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Robertson ED, Wasylyk C, Ye T, Jung AC and
Wasylyk B: The oncogenic MicroRNA Hsa-miR-155-5p targets the
transcription factor ELK3 and links it to the hypoxia response.
PLoS One. 9:e1130502014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Garzon R, Heaphy CE, Havelange V, Fabbri
M, Volinia S, Tsao T, Zanesi N, Kornblau SM, Marcucci G, Calin GA,
et al: MicroRNA 29b functions in acute myeloid leukemia. Blood.
114:5331–5341. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Costinean S, Zanesi N, Pekarsky Y, Tili E,
Volinia S, Heerema N and Croce CM: Pre-B cell proliferation and
lymphoblastic leukemia/high-grade lymphoma in E(mu)-miR155
transgenic mice. Proc Natl Acad Sci USA. 103:pp. 7024–7029. 2006;
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen G, Tang Y, Wu JH and Liu FH: Role of
microRNAs in diagnosis and treatment of the pathogenesis of gastric
cancer. Int J Clin Exp Med. 7:5947–5957. 2014.PubMed/NCBI
|
|
24
|
Saito Y, Suzuki H, Matsuura M, Sato A,
Kasai Y, Yamada K, Saito H and Hibi T: MicroRNAs in hepatobiliary
and pancreatic cancers. Front Genet. 2:662011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Greither T, Grochola LF, Udelnow A,
Lautenschläger C, Würl P and Taubert H: Elevated expression of
microRNAs 155, 203, 210 and 222 in pancreatic tumors is associated
with poorer survival. Int J Cancer. 126:73–80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kong W, Yang H, He L, Zhao JJ, Coppola D,
Dalton WS and Cheng JQ: MicroRNA-155 is regulated by the
transforming growth factor beta/Smad pathway and contributes to
epithelial cell plasticity by targeting RhoA. Mol Cell Biol.
28:6773–6784. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Neilsen PM, Noll JE, Mattiske S, Bracken
CP, Gregory PA, Schulz RB, Lim SP, Kumar R, Suetani RJ, Goodall GJ
and Callen DF: Mutant p53 drives invasion in breast tumors through
up-regulation of miR-155. Oncogene. 32:2992–3000. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Willimott S and Wagner SD: miR-125b and
miR-155 contribute to BCL2 repression and proliferation in response
to CD40 ligand (CD154) in human leukemic B-cells. J Biol Chem.
287:2608–2617. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang M, Zhang P, Li Y, Liu G, Zhou B, Zhan
L, Zhou Z and Sun X: The quantitative analysis by stem-loop
real-time PCR revealed the microRNA-34a, microRNA-155 and
microRNA-200c overexpression in human colorectal cancer. Med Oncol.
29:3113–3118. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kara M, Yumrutas O, Ozcan O, Celik OI,
Bozgeyik E, Bozgeyik I and Tasdemir S: Differential expressions of
cancer-associated genes and their regulatory miRNAs in colorectal
carcinoma. Gene. 567:81–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Renault VM, Thekkat PU, Hoang KL, White
JL, Brady CA, Kenzelmann Broz D, Venturelli OS, Johnson TM, Oskoui
PR, Xuan Z, et al: The pro-longevity gene FoxO3 is a direct target
of the p53 tumor suppressor. Oncogene. 30:3207–3221. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Naito S, von Eschenbach AC, Giavazzi R and
Fidler IJ: Growth and metastasis of tumor cells isolated from a
human renal cell carcinoma implanted into different organs of nude
mice. Cancer Res. 46:4109–4115. 1986.PubMed/NCBI
|
|
34
|
Min M, Peng L, Yang Y, Guo M, Wang W and
Sun G: MicroRNA-155 is involved in the pathogenesis of ulcerative
colitis by targeting FOXO3a. Inflamm Bowel Dis. 20:652–659. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ling N, Gu J, Lei Z, Li M, Zhao J, Zhang
HT and Li X: microRNA-155 regulates cell proliferation and invasion
by targeting FOXO3a in glioma. Oncol Rep. 30:2111–2118. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Scholzen T and Gerdes J: The Ki-67
protein: From the known and the unknown. J Cell Physiol.
182:311–322. 2010. View Article : Google Scholar
|
|
37
|
Hu XJ, Xie MY, Kluxen FM and Diel P:
Genistein modulates the anti-tumor activity of cisplatin in MCF-7
breast and HT-29 colon cancer cells. Arch Toxicol. 88:625–635.
2014.PubMed/NCBI
|
|
38
|
Serova M, Calvo F, Lokiec F, Koeppel F,
Poindessous V, Larsen AK, Laar ES, Waters SJ, Cvitkovic E and
Raymond E: Characterizations of irofulven cytotoxicity in
combination with cisplatin and oxaliplatin in human colon, breast,
and ovarian cancer cells. Cancer Chemother Pharmacol. 57:491–499.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lacour S, Micheau O, Hammann A, Drouineaud
V, Tschopp J, Solary E and Dimanche-Boitrel MT: Chemotherapy
enhances TNF-related apoptosis-inducing ligand DISC assembly in
HT29 human colon cancer cells. Oncogene. 22:1807–1816. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Burgering BM: A brief introduction to
FOXOlogy. Oncogene. 27:2258–2262. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Arden KC: FoxO: Linking new signaling
pathways. Mol Cell. 14:416–418. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen Q, Ganapathy S, Singh KP, Shankar S
and Srivastava RK: Resveratrol induces growth arrest and apoptosis
through activation of FOXO transcription factors in prostate cancer
cells. PLoS One. 5:e152882010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Calin GA, Liu CG, Sevignani C, Ferracin M,
Felli N, Dumitru CD, Shimizu M, Cimmino A, Zupo S, Dono M, et al:
MicroRNA profiling reveals distinct signatures in B cell chronic
lympho cytic leukemias. Proc Natl Acad Sci USA. 101:pp.
11755–11760. 2004; View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tili E, Croce C and Michaille J: miR-155:
On the crosstalk between inflammation and cancer. Int Rev Immunol.
28:264–284. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Iorio MV, Ferracin M, Liu CG, Veronese A,
Spizzo R, Sabbioni S, Magri E, Pedriali M, Fabbri M, Campiglio M,
et al: MicroRNA gene expression deregulation in human breast
cancer. Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:pp. 2257–2261.
2006; View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Valeri N, Gasparini P, Fabbri M, Braconi
C, Veronese A, Lovat F, Adair B, Vannini I, Fanini F, Bottoni A, et
al: Modulation of mismatch repair and genomic stability by miR-155.
Proc Natl Acad Sci USA. 107:pp. 6982–6987. 2010; View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sandhu SK, Volinia S, Costinean S, Galasso
M, Neinast R, Santhanam R, Parthun MR, Perrotti D, Marcucci G,
Garzon R and Croce CM: miR-155 targets histone deacetylase 4
(HDAC4) and impairs transcrip tional activity of B-cell lymphoma 6
(BCL6) in the Emu-miR-155 transgenic mouse model. Proc Nat Acad Sci
USA. 109:pp. 20047–20052. 2012; View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li S, Chen T, Zhong Z, Wang Y, Li Y and
Zhao X: microRNA-155 silencing inhibits proliferation and migration
and induces apoptosis by upregulating BACH1 in renal cancer cells.
Mol Med Rep. 5:949–954. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang P, Bill K, Liu J, Young E, Peng T,
Bolshakov S, Hoffman A, Song Y, Demicco EG, Terrada DL, et al:
miR-155 is a liposarcoma oncogene that targets casein kinase-1α and
enhances b-catenin signaling. Cancer Res. 72:1751–1762. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lv L, An X, Li H and Ma L: Effect of
miR-155 knockdown on the reversal of doxorubicin resistance in
human lung cancer A549/dox cells. Oncol Lett. 11:1161–1166. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Vigorito E, Kohlhaas S, Lu D and Leyland
R: miR-155: An ancient regulator of the immune system. Immunol Rev.
253:146–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Higgs G and Slack F: The multiple roles of
microRNA-155 in onco genesis. J Clin Bioinforma. 3:172013.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yu DD, Lv MM, Chen WX, Zhong SL, Zhang XH,
Chen L, Ma TF, Tang JH and Zhao JH: Role of miR-155 in drug
resistance of breast cancer. Tumour Biol. 36:1395–1401. 2015.
View Article : Google Scholar : PubMed/NCBI
|