Introduction
Liver cancer is one of the most common malignant
diseases globally (1). Between 2012
and 2017, numerous therapeutics of liver cancer have indicated
potential in clinical trials, particularly immunotherapies
(2). Cytokine-induced killer (CIK)
cells, which are the major histocompatibility complex-unrestricted
cytotoxic lymphocytes generated with tumor necrosis factor-α
(TNF-α), interferon-γ (IFN-γ), interleukin (IL)-2 and IL-12, serve
an important role in immunotherapy (3). CIK cell-based immunotherapy has been
indicated to be effective in the majority of tumors types, and is
currently studied in clinical trialsfrequently (4,5).
Programmed cell death-1 (PD-1) is a prominent
regulator of T-cell function, and is expressed on T cells following
chronic antigenic stimulation (6,7). Up
regulation of PD-1 was identified to be associated with the
suppression of T-cell function via mediating T cell apoptosis;
however, down regulation of PD-1 was revealed to have the ability
to reverse immune dysfunction (8–14).
Clinical studies targeting the PD-1-PD-L1 axis have demonstrated
effectiveness in a number of cancer types (15–17).
Engagement of PD-1 in T cells with its ligand PD-L1 in tumor cells
down regulates antitumor T-cell responses, and allows tumors to
escape these responses (18,19).
MicroRNAs (miRNAs), a class of highly conserved long
noncoding RNA, regulate gene expression negatively via targeting
the 3′ untranslated region (3′UTR) or coding region of the mRNA;
additionally, they participate in a number of biological processes,
including cell proliferation, differentiation, apoptosis and
tumorigenesis (20,21). A previous study indicated that miRNAs
are involved in regulation of the immune system and development of
immune cells (22).
The microRNA database (miRBase, release 21,
http://www.mirbase.org/cgi-bin/query.pl?terms=miR-374)
demonstrated that miR-374a and miR-374b may bind the 3′UTR region
of PD-1 mRNA (miRSVR score ≤0.1). Research has also indicated that
miR-374a and miR-374b could be expressed in cluster of
differentiation (CD) 4+T, CD8+T and natural killer (NK) cells
(23,24). In the present study, it was
demonstrated that miR-374b binds to the 3′UTR of PD-1 by using
Dual-Luciferase Reporter Assay. Therefore, miR-374b was selected as
the research target. In recent years, studies have indicated that
miR-374b affects the apoptosis ability of colorectal cancer cells
(25,26). miR-374b can accelerate cell
proliferation and the production of aberrant glycosylated
immunoglobulin (Ig) A1 in B cells (27), and it has the ability to participate
in the development of human osteosarcoma cells (28). Additionally, miR-374b can negatively
regulate C2C12 myoblast differentiation via targeting Myf6
(29), and inhibit proliferation and
promote apoptosis of T-cell lymphoblastic lymphoma (30). In addition, miR-374b is associated
with cisplatin resistance in pancreatic cancer cells (31), andit is capable of accelerating
invasion and metastasis of gastric cancer cells (32); however, the role of miR-374b in liver
cancer remains unclear. Therefore, it is essential to explore
miR-374b, which may become anovel promising therapeutic target for
liver cancer.
In the present study, miR-374b was predicted to
interact with PD-1 and was demonstrated to affect the
tumor-targeting capacity of CIK cells. The result indicated that
siPD-1 promoted the CIK secreting cytokine, but inhibited the
viability of HepG2 cells. In vivo study indicated that
siPD-1 decreased the tumor volume in liver cancer mouse models. In
conclusion, human CIK cells transfected with siPD-1 can target
liver cancer cells and enhance immunotherapy efficacy, and
therefore have a potential in the immunotherapy of liver
cancer.
Materials and methods
Cell lines and transfection
Liver cancer cell lines (HepG2, PLC and Huh7) were
purchased from American Type Culture Collection (Manassas, VA, USA)
and cultured in Dulbecco's modified Eagle's medium (DMEM; In
vitro gen; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
and normal hepatocytes (L-02 cells) were cultured in RPMI-1640
medium (In vitro gen; Thermo Fisher Scientific, Inc.). Each
medium contained 10% fetal calf serum (FCS; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) and 1% penicillin-streptomycin G (In
vitro gen; Thermo Fisher Scientific, Inc.). All cells were
incubated at 37°C in a humidified atmosphere of 5%
CO2.
miR-374b mimic, negative control (NC), miR-374b
inhibitor oligonucleotides and PD-1 siRNA were synthesized by
Shanghai Gene Pharma, Co., Ltd. (Shanghai, China) and the sequences
are as follows: miR-374b mimics, 5′-AUAUAAUACAACCUGCUAAGUG-3′; NC,
5′-UUCUCCGAACGUGUCACGUTT-3′; miR-374b inhibitor,
5′-CACUUAGCAGGUUGUAUUAUAU-3′; PD-1 siRNA,
5′-CCAGGAUGGUUCUUAGACUUU-3′.
In all experiments, the incubation was conducted at
37°C in a humidified atmosphere containing 5% CO2. CIK
cells were generated from peripheral blood mononuclear cells
(PBMCs) of healthy volunteers. A total of 2×104 cells in
the logarithmic phase were seeded into each well of a 6-well plate
in 2 ml of Opti-MEM I reduced serum medium (Life Technologies;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) and incubated
over night at 37°C in a humidified atmosphere of 5% CO2.
The next day, cells were transfected with 50 µM scramble siRNA
(negative control, NC), 50 µM PD-1 siRNAs, 50 nM miR-374b mimic, 50
nM negative control (NC) and 50 nM miR-374b inhibitor
oligonucleotides for 48 h using Lipofectamine® 2000
reagent (In vitro gen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol.
Preparation and identification of
human CIK cells
Human PBMCs were obtained from healthy donors via
Ficoll-Hypaque density centrifugation (3,000 × g for 30 min at
4°C), and then washed three times with PBS. Cells were resuspended
in 5 ml RPMI-1640 medium containing 1×106U/l human IFN-γ
(R&D Systems, Inc., Minneapolis, MN, USA; cat no., 285-IF) at a
concentration of ~3×106 cells/ml and incubated overnight
at 37°C in an atmosphere containing 5% CO2. After 24 h,
1,000 units/ml IL-2 (Chiron Corporation, Emeryville, CA, USA),
IL-1a (Chiron Corporation), 50 µg/l each of
allophycocyanin-conjugated anti-CD3 (cat. no., 553066; BD
Biosciences, Franklin Lakes, NJ, USA) and anti-CD28 (cat no.,
14-02281-86, eBioscience; Thermo Fisher Scientific, Inc.)
monoclonal antibodies (mAbs) were added. Fresh medium and fresh
IL-2 (cat no., 575406) were added every 2 days and the cells were
harvested on days 1,7, 14 and 21 and assessed using FACS
(FACSCalibur™; BD Biosciences, Franklin Lakes, NJ, USA) with
fluorescein isothiocyanate-conjugated anti-CD3 (cat no., 555274; BD
Biosciences) and phycoerythrin-CD56 (cat no., 561903; BD
Bioscience) using Flow Jo software (version 8.7.1; Flow Jo LLC,
Ashland). The protocol for the present study was approved by The
Ethical Review Committee of The First Affiliated Hospital of Hainan
Medical University (Hainan Province, China). Informed consent was
obtained from each person.
Luciferase reporter assay
The database Target Scan (http://www.targetscan.org) was used to predict
potential targets for miR-374b. DNA fragments of the PD-1 3′UTR
containing the putative miR-374b binding site or mutated (Mut)
miR-374b binding site were amplified bypolymerase chain reaction
(PCR) using 2× Taq PCR Master Mix (Tiangen Biotech Co., Ltd.,
Beijing, China) from CIK cell genomic DNA. The thermocycling
conditions were as follows: 95°C for 5 mins, then 35 cycles of 95°C
for 30 secs, 57°C for 30 secs, 72°C for 1 min, followed by an
extension at 72°C for 10 min. The primers were as follows:
PD-1-XhoI 5′-CCGCTCGAGCAGTAAGCGGGCAGGC-3′ (forward),
PD-1-NotI5′-ATTTGCGGCCGCTCCTTAGCATGCTCTCATATTT-3′ (reverse);
PD-1-MUT 5′-CCTTCCCTGTGGTTCGCACTGGTTATAATTATAA-3′ (forward),
PD-1-MUT 5′-TTATAATTATAACCAGTGCGAACCACAGGGAAGG-3′ (reverse). The
DNA products were then inserted into the Pme I/Spe I sites of the
firefly luciferase coding region of the pMIR-report vector (Thermo
Fisher Scientific, Inc.). The plasmids were termed as wild-type
(pMIR-report-PD-1-WT) and Mut (pMIR-report-PD-1-Mut) sequences. The
mutation of UAAUAU to AUUAUA was introduced into the potential
miR-374b binding sites. A total of 8×104 cells) were
cultured in each well of a 96-well plate, and co-transfected with
miR-374b mimic and NC, and WT or Mut 3′-UTR of PD-1using
Lipofectamine 2000 reagent (In vitro gen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocolat 37°C
in an incubator with 5% CO2 for 48 h. The luciferase
activities were detected by Dual-Luciferase Reporter Assay ki
(Promega Corporation, Madison, WI, USA; cat. no., E1910). Cells
were collected, washed using PBS and lysed in Passive Lysis Buffer
(Promega Corporation) at 48 h following transfection. The
Dual-Luciferase Reporter Assay System (Promega Corporation) was
used to analyze the data. the Renilla luciferase reporter was used
as the internal control. Data are presented as the means ± standard
deviation (SD) of three independent experiments.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was isolated using TRIzol®
(In vitro gen; Thermo Fisher Scientific, Inc.), accordingly
to the manufacturer's instructions, and reversely transcribed to
cDNA via RT-PCR using a mi Script II RT Kit (Qiagen GmbH, Hilden,
Germany). The mRNA expression levels were detected using the
SYBR® Green PCR Master Mix kit (Clontech Laboratories,
Inc., Mountain view, CA, USA). The reaction system was performed in
a volume of 20 µl, and the thermo cycling conditions were as
follows: 95°C for 10 min, then 40 cycles of 95°C for 10 sec, 60°C
for 2 min, 72°C for 2 min, followed by an extension at 72°C for 10
mins. The target genes and controls were analyzed via RT-qPCR and
the reactions were performed on the ABI 7500 system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) with primers specific
for target genes: PD-1, forward: 5′-ATGACAGCGGCACCTACCT-3′,
reverse: 5′-CCTATTGTCCCTCGTGCG-3′; miR-374b-5p, forward:
5′-ACACTCCAGCTGGGATATAATACAACCTGCTA-3′, reverse:
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCACTTAGC-3′; PD-L1, forward:
5′-CCATCAAGTCCTGAGTGGTAAG-3′, reverse:
5′-TTGTTGTGTTGATTCTCAGTGTG-3′; GAPDH, forward:
5′-TGTTCGTCATGGGTGTGAAC-3′, reverse: 5′-ATGGCATGGACTGTGGTCAT-3′;
U6, forward: 5′-CTCGCTTCGGCAGCACA-3′, reverse:
5′-AACGCTTCACGAATTTGCGT-3′. The data were analyzed using
2−∆∆Cq (33). The relative
mRNA/miRNA expression levels were normalized to GAPDH/U6. All data
were presented as the mean ± SD of three independent
experiments.
Western blot analysis
Cell lysates were prepared in RIPA buffer (Beyotime
Institute of Biotechnology, Shanghai, China) via incubation for 20
min at 4°C. The protein concentrations were determined using a BCA
Protein Assay kit (Thermo Fisher Scientific, Inc.). Equal amounts
of total proteins (30 µg) were separated using 10% SDS-PAGE gels
based on the molecular weight of the objective proteins and
transferred onto a polyvinylidene difluoride membranes (Perkin
Elmer, Inc., Waltham, MA, USA). The PVDF membranes were blocked in
5% skim milk (BD Biosciences, Franklin Lakes, NJ, USA) for 2 h at
room temperature. Following this, the cells were incubated with
anti-GAPDH (dilution, 1:1,000; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA; cat. no. sc-365062) and anti-PD-1 antibody
(dilution, 1:1,000; Abcam, Cambridge, UK; cat no. ab52587) at 4°C
overnight. The blots were subsequently incubated with horseradish
peroxidase (HRP)-conjugated secondary antibodies (goat anti-mouse;
dilution, 1:2,000; cat no., SC-2005, and goat anti-rabbit;
dilution, 1:2,000, cat no., SC-2004) for 1 h at room temperature.
Finally, the proteins were detected using an ECL detection kit (EMD
Millipore, Billerica, MA, USA).
PD-1 expression on liver cancer cells
via flow cytometry
The liver cancer cell lines were cultured in DMEM
(In vitro gen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) with 10% FBS and the normal hepatocytes were cultured in
RPMI-1640 medium (In vitro gen; Thermo Fisher Scientific,
Inc.) with 10% FBS at 37°C in a humidified atmosphere of 5% CO2.
Liver cancer cell lines were centrifuged separately (100 × g for 5
mins at 4°C) and the concentration of cells was adjusted to
1×106 cell/ml by using phosphate-buffered saline (PBS,
cat. No. D-1408). Then, 1 ml cell suspension was obtained and
anti-human CD279 (PD-1) phycoerythrin (PE) monoclonal antibody
(dilution, 1:50; cat no., 12-9985-82; eBioscience; Thermo Fisher
Scientific, Inc.) was added, and cultured at 4°C for 30 min. Cells
were resuspended in 500 µl PBS for flow cytometry (FACSCalibur Flow
Cytometer; BD Biosciences) and analyzed with Flow Jo (version
4.5.4; Flow Jo LLC, Ashland). Mouse IgG1 K Iso type Control PE
antibody (dilution, 1:50; cat. no. 12-4301; eBioscience; Thermo
Fisher Scientific, Inc.) was used as a NC. All data are performed
as the mean ± SD of three independent experiments.
Cytokine secretion assays
3×105 cells were seeded into a 6-well
microplate (cat no., 353046BD; Biosciences,) and incubated at 37°C
overnight. RPMI-1640 medium was added without FCS, and the
secretion of IFN-γ was measure dusing an enzyme-linked
immunosorbent assay (ELISA) kit (cat no., DY459; R&D Systems,
Inc.) after incubation for 72 h at 37°C according to the
manufacturer's instructions. All data are presented as the mean ±
SD of 3 independent experiments.
Cell counting kit-8 (CCK-8) assay
Cell viability was determined using a cell counting
kit-8 (CCK-8; cat no., CK04-11; Dojindo Molecular Technologies,
Inc., Rockville, MD, USA). Effect cells (CIK cells) and target
cells (HepG2 cells) with a 20:1 ratio of effect: target (E/T) cells
were seeded at a density of 6×103 in a 96-well plate
(Corning Life Sciences, Tewksbury, MA, USA) for 24 h at 37°C.
Following this, the cells were washed using PBS once. Then, 100 µl
RPMI-1640 medium and 10 µl CCK-8 was added to each well. Following
2 h at 37°C, the 96-well plate was measured at 450 nm using a
standard micro plate reader (Scientific™ Multiskan™ MK3; Thermo
Fisher Scientific, Inc.). The cell viability was calculated
according to the formula: Cell viability=[optical density (OD) of
the experimental sample/OD of the control group] ×100%. HepG2 cells
treated with an equivalent volume of PBS served as the negative
control group, and the experiment was repeated 3 times.
Cytotoxicity assay
A lactate dehydrogenase (LDH) release assay was used
to determine the cytotoxicity of the immune cells. The ratio of E/T
cells was 20:1. Target cells and effect cells were added to a
96-well culture plate with incomplete medium (DMEM/10% FBS/100 U/ml
Penicillin-Streptomycin) for triplicate wells and incubated for 24
h in a humidified atmosphere containing 5% CO2 at 37°C.
Following centrifugation of the suspension at 1,700 × g for 4 min
at 4°C, 50 µl was removed and mixed with 50 µl LDH substrate
solution (cat. no., L2402, Sigma-Aldrich; Merck KGaA) and then
incubated for 30 min at room temperature in the dark. Finally, 50
µl stop solution (50% dimethyl form amide and 20% sodium dodecyl
sulfate; pH 4.7) was added and the concentration of LDH in each
well was measured using an automatic ELISA reader (ELISA reader;
ASYS Hitech, GmbH, Austria). All data are performed as the mean ±
SD of three independent experiments.
Animals
A total of 24 female nude mice (6–8 weeks-old and
weighing 22–24 g) were purchased from Laboratory Animal Center of
Hainan Medical University. All mice were maintained in the
Laboratory Animal Center of Hainan Medical University at a
controlled temperature (25–28°C) and a humidity within 50–60%,
under a 12/12 h light/dark cycle, with ad libitum access to
sterile food and water. All the experiments were approved and
performed according to the Animal Ethics Committee of Hainan
Medical University.
Winn assay
A Winn assay was conducted to examine the tumor
growth-suppressing effect of CIK cells in mice (n=6). The E/T ratio
was 10:1. CIK, miR-374b inhibitor, PD-1 siRNA and HepG2
(CIK+Inhibitor+siRNA+HepG2) cells (1×106) or
CIK+Inhibitor+NC+HepG2 (control) cells (1×106) were
incubated for 12 h at 37°C, adjusted to a sugar concentration of
1.5 mg/ml, and then were subcutaneously injected into the leg of
nude mice. The volume of the tumors was measured on 7, 14, 21 and
28 days, respectively. Following the completion of the
administration, the mice were sacrificed at 28 days, with the
maximum tumor size measuring 360 mm2.
Adoptive transfer assay
To study the immunotherapy effect of CIK on liver
cancer, the mice (n=12) were randomly divided into two groups
according to the different treatments received: i)
Inhibitor+NC+HepG2; and ii) Inhibitor+siRNA+HepG2. The treated
HepG2 (1.5×107 cells/injection) cells were
subcutaneously infected into the leg of nude miceonce. A week
later, CIK cells (1.0×107) were intravenously injected
in the mice via their tail veinsonce a week. The volume of the
tumors was measured on days 7, 14, 21 and 28. Following the
completion of the administration, the mice were sacrificed at 28
days, with the maximum tumor size measuring 422 mm2.
Statistical analysis
Data are expressed as mean ± SD. One-way analysis of
variance and Dunnett's post-hoc test were calculated using the SPSS
software package version 11.5 (SPSS, Inc., Chicago, IL, USA).
P≤0.05 was considered to indicate statistically significant
difference.
Results
Preparing CIK cell bi-specific
antibodies against CD3 and CD56
Peripheral blood was collected from healthy donors.
On days 1,7, 14 and 21, mouse anti-human CD3 and CD56 mAbs were
used to analyze the phenotypes of the cells measured via flow
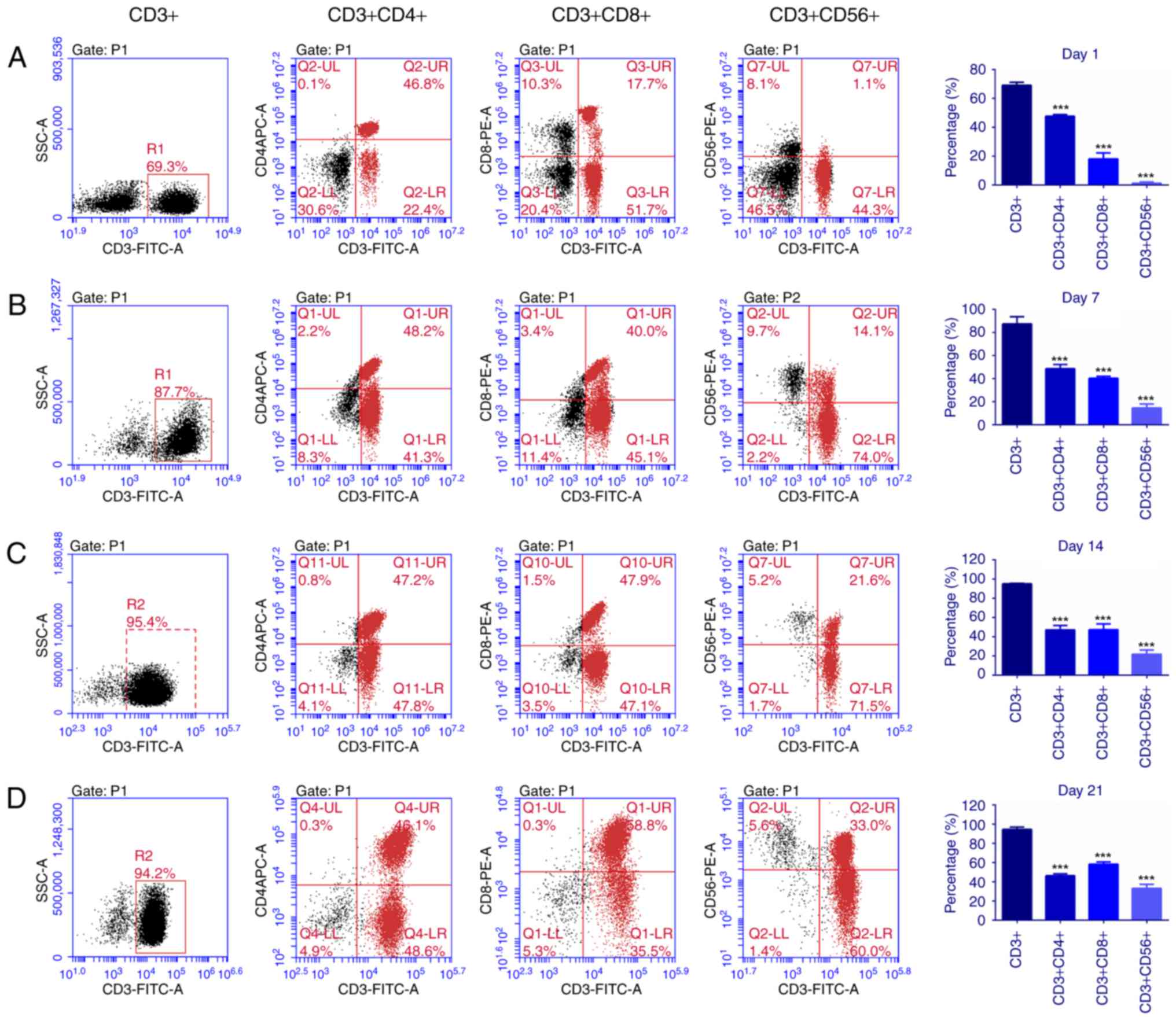
cytometry. The results in Fig. 1
depicted that following incubation with multiple cytokines for 1,7,
14 and 21 days, the percentage of CD3+ cells and CD3+CD56+ cells
significantly increased (P<0.001), whilst the percentage of CD4+
cells did not change significantly. In combination, the data
indicated that the CIK cells were mainly composed of T cells and NK
T cells.
Predicting that miR-374b binds to the
3UTR of PD-1 mRNA
To experimentally confirm that PD-1 is an authentic
target of miR-374b in CIK cells, the PD-1-WT and PD-1-Mut
luciferase reporter vectors were transfected into CIK cells in
combination with NC or miR-374b mimic. Following 48 h of
transfection, the results demonstrated that the luciferase activity
in the PD-1-WT with miR-374b mimic group was significantly reduced,
compared with the other three groups (P<0.01; Fig. 2A). The aforementioned data
demonstrated that PD-1 is a genuine target of miR-374b.
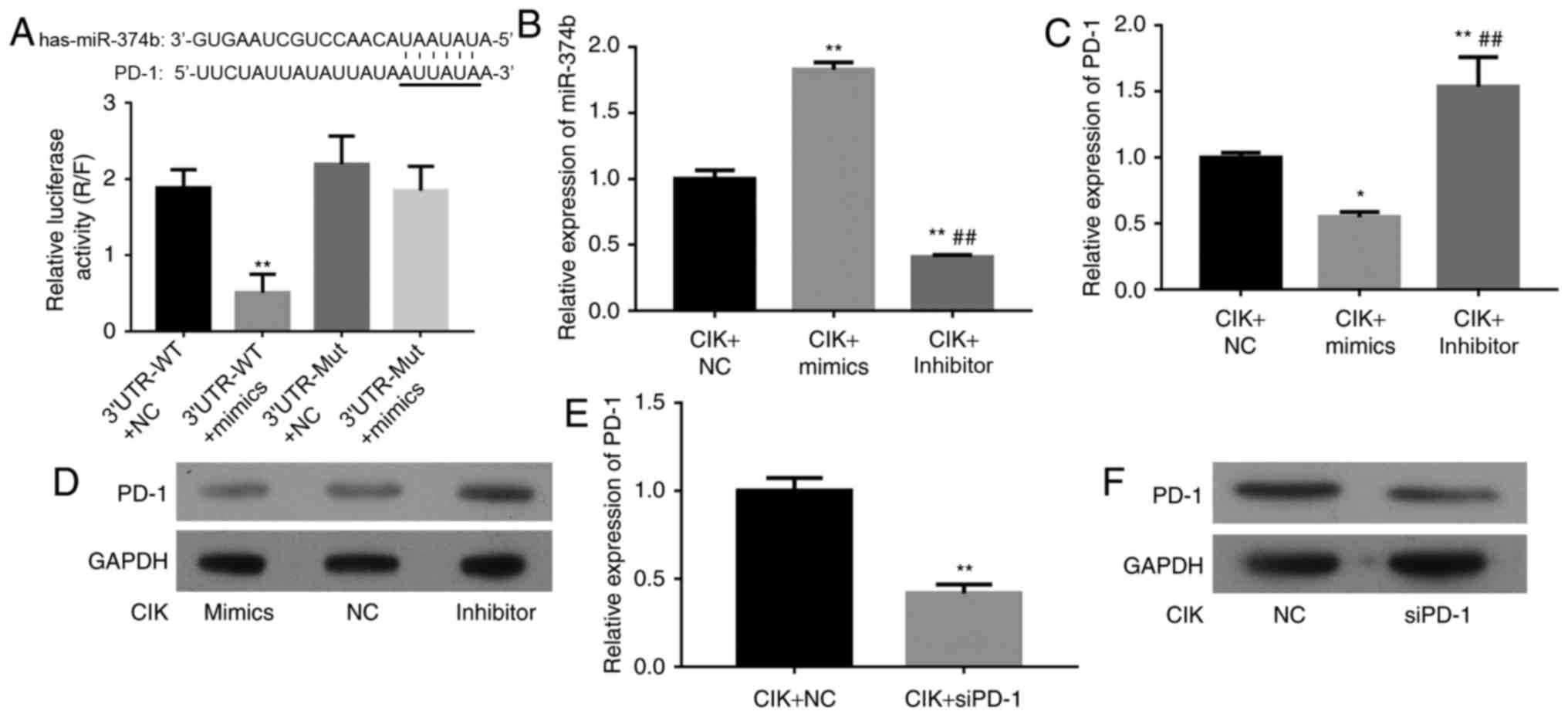 | Figure 2.The targeting association between
miR-374b and PD-1. (A) The target gene was predicted using the
Target Scan database. Following 48 h of transfection, the
luciferase activities of the four groups were detected by
dual-Luciferase reporter gene system. The results indicated that
the luciferase activity of PD-1-WT with miR-374b mimic group was
significantly reduced, compared with the other three groups
(**P<0.01). (B) CIK cells were transfected with NC, miR-374b
mimic or miR-374b inhibitor. The efficiency of this miR-374b mimic
or inhibitor was confirmed via qPCR assay. (C) Relative PD-1 mRNA
expression level in transfected CIK cells was confirmed via qPCR
(*P<0.05 vs. NC group; **P<0.01, compared with NC group;
##P<0.01, compared with miR-374b mimic group). (D)
PD-1 protein expression levels were confirmed via western blot
analysis. GAPDH was used as a reference. (E and F) CIK cells were
transfected with NC or siPD-1. The efficiency of siPD-1 was
confirmed via qPCR and western blot analysis (**P<0.01 vs. NC
group). WT, wild type; Mut, mutated; UTR, untranslated region; CIK,
cytokine-induced killer; NC, negative control; siPD-1, small
interfering programmed cell death-1; qPCR, quantitative polymerase
chain reaction; miR, microRNA; Inhibitor, miR-374b inhibitor. |
Effect of miR-374b on PD-1
expression
Due to the level of miR-374b being increased in CIK
cell lines, its role in CIK cells was investigated. A miR-374b
mimic was used to amplify the expression of miR-374b and a
synthetic inhibitor specific for miR-374b was employed to suppress
the expression of miR-374b in CIK cell lines. The efficiency of
this miR-374b mimic or inhibitor was confirmed using qPCR. The
relative miR-374b RNA expression was notably increased in the
miR-374b mimic transfected group (P<0.01, compared with the NC),
however significantly decreased in miR-374b inhibitor transfected
group (P<0.01, compared with the NC and miR-374b mimic groups;
Fig. 2B). The PD-1 expression levels
in the NC, miR-374b mimic and miR-374b inhibitor groups were
confirmed using qPCR and western blot analysis. The results
demonstrated that PD-1 expression levels in the miR-374b mimic
group were significantly reduced (P<0.05, compared with NC
group), however, significantly increased in miR-374b inhibitor
group. (P<0.01, compared with the NC and miR-374b mimic groups;
Fig. 2C and D).
Effect of siPD-1 on CIK cells
The PD-1 expression levels were confirmed using qPCR
and western blot analysis. The relative PD-1 mRNA expression levels
and protein expression levels were significantly decreased in the
siPD-1 group, compared with the NC group (P<0.01; Fig. 2E and F).
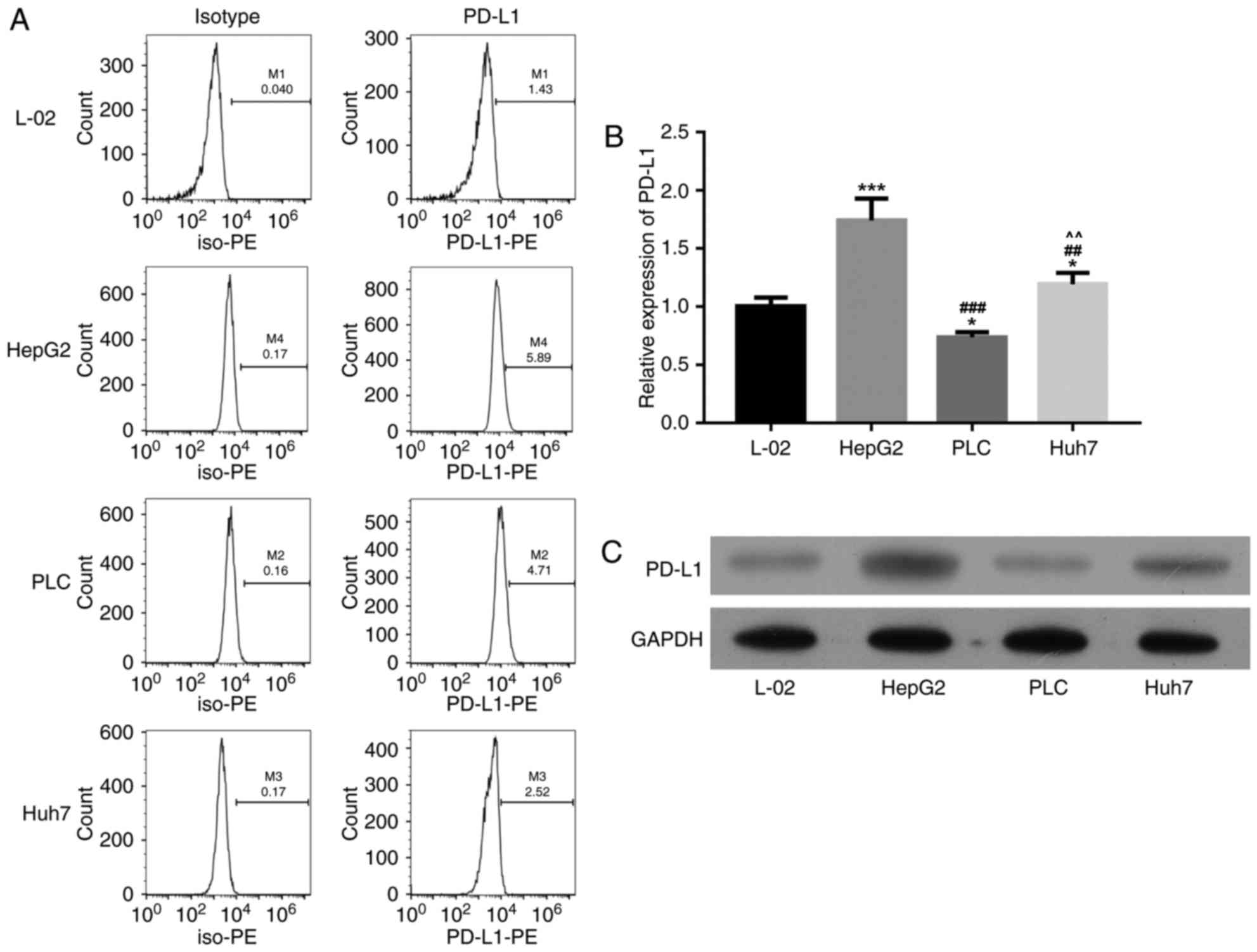
Selecting the high PD-1 expression
liver cancer cell line
Compared with the L-02 cells group, PD-1 expression
was significantly increased in HepG2 group (P<0.01), as
demonstrated via flow cytometry, RT-PCR and western blot analysis.
Consequently, HepG2 cells were selected for the following
transfecting experiments (Fig.
3).
Effect of PD-1 on IFN-γ production of
CIK cells
A synthetic siRNA targeting PD-1 was employed to
silence the expression level of PD-1 in CIK cells. The secretion of
IFN-γ was measured using an ELISA. The result demonstrated that
IFN-γ secretion was significantly increased in the
CIK+Inhibitor+siRNA group, compared with the CIK+Inhibitor+NC group
(P<0.01; Fig. 4A).
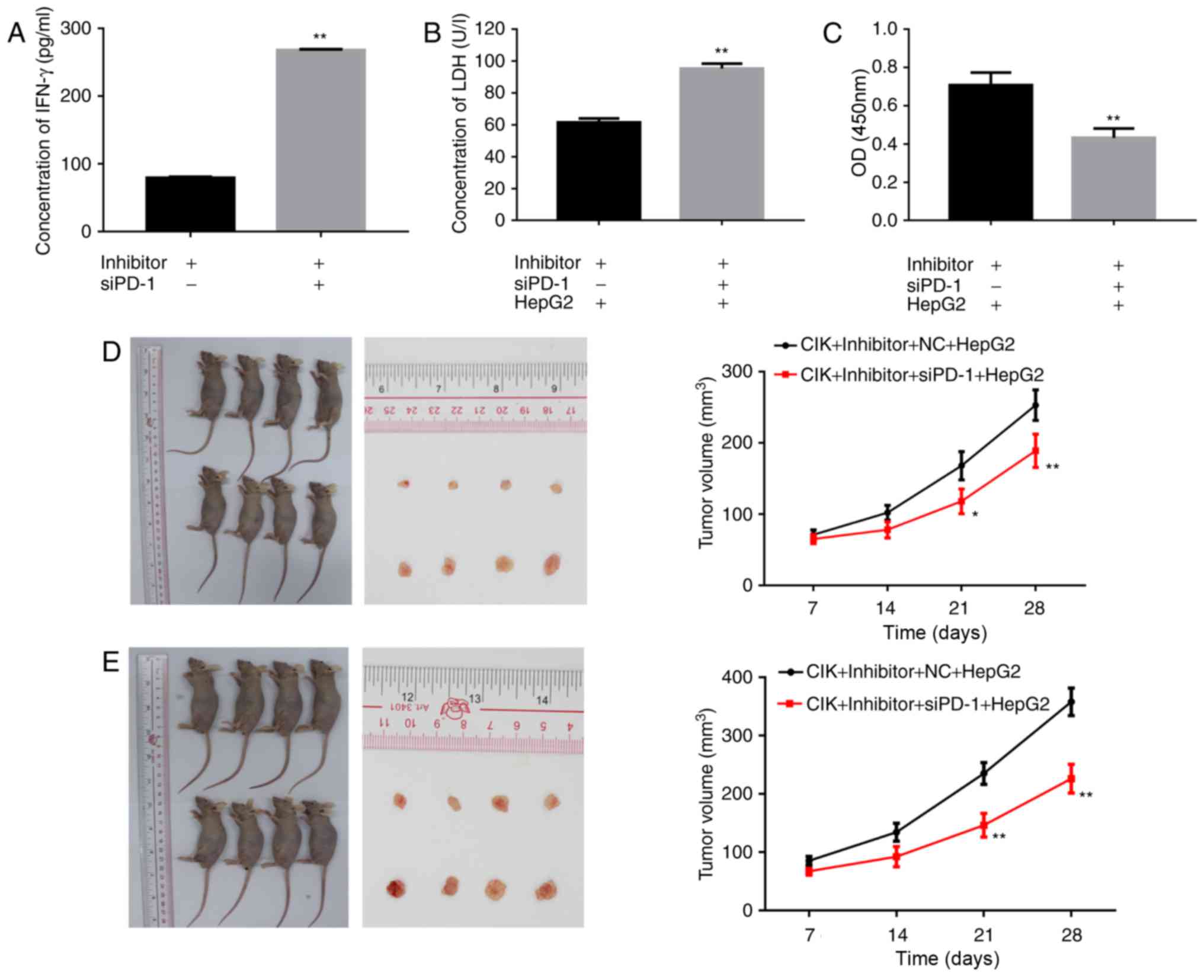 | Figure 4.Effect of PD-1 on IFN-γ production,
cell viability, cytotoxicity and antitumor effect in CIK cells
in vitro and in vivo. A synthetic siRNA targeting
PD-1 was employed to silence the expression level of PD-1 in CIK
cells. (A) An ELISA was used to measure the expression of IFN-γ in
CIK cells. The bands were quantified (**P<0.01 vs.
CIK+Inhibitor+NC group). (B) ALDH release assay was used to
determine the cytotoxicity of the CIK on HepG2 cells (**P<0.01
vs. CIK+Inhibitor+NC+HepG2 group). (C) The Cell Counting Kit-8
assay was performed to visualize the proliferation of HepG2 cells
with CIK (**P<0.01 vs. CIK+Inhibitor+NC+HepG2 group). (D) Winn
assay and (E) adoptive transfer assay were used to examine the
antitumor effect of siPD-1 transfected CIK cells in nude mice. The
volume of the tumors in different groups is depicted
(*P<0.05,**P<0.01 vs. CIK+Inhibitor+NC+HepG2 group). IFN,
interferon; siPD-1, small interfering programmed cell death-1; NC,
negative control; CIK, cytokine-induced killer; Inhibitor, miR-374b
inhibitor; LDH, lactate dehydrogenase; OD, optical density. |
Effects of siPD-1 on cytotoxicity of
CIK cells
A LDH release assay was used to determine the
cytotoxicity of the immune cells. the concentration of LDH in the
CIK+Inhibitor+siRNA+HepG2 group was significantly increased,
compared with the CIK+Inhibitor+NC+HepG2 group (P<0.01; Fig. 4B).
Effects of siPD-1 transfected CIK on
HepG2 cell viability
Following this, PD-1 siRNA was designed and
transfected into CIK cells to knockdown the endogenous expression
of PD-1. A CCK-8 assay was performed to visualize the proliferation
of the HepG2 cells. the viability of HepG2 cells in the
CIK+Inhibitor+siRNA+HepG2 group was significantly decreased,
compared with the CIK+Inhibitor+NC+HepG2 group (P<0.01; Fig. 4C).
Antitumor effect of siPD-1 transfected
CIK cells in nude mice
Winn assay and adoptive transfer assay were used to
examine the antitumor effect of siPD-1 transfected CIK cells in
nude mice. The result demonstrated that the volume of the tumors in
CIK+Inhibitor+siRNA+HepG2 group was notably reduced, compared with
the CIK+Inhibitor+NC+HepG2 group (P<0.05 and P<0.01 at
different time points; Fig. 4D and
E).
Discussion
In liver cancer, the activities of liver- and
tumor-infiltrating T cells are restricted due to the presence of
multiple immune suppression mechanisms underlying the hepatic
microenvironment (34,35). Particularly, the PD-1 signaling
pathway negatively regulates antitumor immunity, resulting in lower
proliferation, IFN-γ production and cytotoxicity by T cells
(36,37). PD-1 is an immune checkpoint inhibitor
primarily expressed in CD8+ T lymphocytes, and frequently co-opted
via cancer cells to escape immune surveillance (38). Additionally, as a regulator, PD-1
suppresses proliferation and cytokine production of T cells
(39). Recently, numerous studies
were regarding the functional roles of PD-1 in T lymphocytes
(40–42). In addition, previous studies indicated
that the anti-PD-1 antibody has the ability to block this
checkpoint and induce the regression of several tumor types,
including liver cancer, in preclinical studies (43,44). A
number of studies revealed the potential role of miRNAs in cancer
via affecting the cell proliferation, migration and invasion,
through the regulation of different target genes (45–49).
Differential expression levels of miRNAs have been indicated to
contribute to the initiation and progression of liver cancer
(50,51). A previous study reported that miR-4717
suppressed PD-1 expression on lymphocytes in individuals with
rs10204525 genotype GG via the interaction with single-nucleotide
polymorphisms in the 3′UTR of PD-1 gene, and the suppression of
PD-1 expression was associated with increased production of TNF-α
and IFN-γ (52).
In the present study, it was predicted that miR-374b
may target the 3′UTR of PD-1 mRNA. Transfection of CIK cells from
individuals with miR-374b mimics significantly suppressed PD-1 mRNA
and protein expression in CIK cells. By contrast, the miR-374b
inhibitor significantly up regulated the PD-1 expression. In order
to investigate the mechanisms underlying the antitumor effects of
PD-1 in CIK cells, in vitro and in vivo studies were
performed. In vitro, PD-1 high expression CIK cells were
obtained through transfecting miR-374b inhibitor into CIK cells.
Following this, a siRNA targeting PD-1 was constructed and
transfected it to CIK cells to examine the antitumor effects of
PD-1. HepG2 cells with high expression of PD-1 were selected as the
target cells for the following transfection study. The results
demonstrated that IFN-γ secretion and the concentration of LDH were
significantly increased in the CIK+Inhibitor+siRNA group, compared
with the CIK+Inhibitor+NC group; however, the viability of liver
cancer cell line HepG2 in the CIK+Inhibitor+siRNA+HepG2 group had
significantly decreased, compared with the HepG2 cells in the
CIK+Inhibitor+NC+HepG2 group.
In vivo, Winn assay and adoptive transfer
assay were used to examine the antitumor effect of siPD-1
transfected CIK cells in nude mice. The result demonstrated that
mice inoculated with HepG2 cells and miR-374b inhibitor transfected
CIK cells with PD-1 knocked down had a tumor volume significantly
reduced, compared with the control group. The data demonstrated
that PD-1is closely associated with the promotion of tumor
growth.
These observations support the conclusion that the
targeted killing effect of CIK cells was associated with the down
regulation of PD-1 gene expression. The novel data provides a
deeper understanding of the role of PD-1 in immune regulation and
has significant implications for cancer immune therapies targeting
PD-1 on CIK cells; therefore, the results in the present study have
provided novel target and genetic information, which may be
employed for designing new therapeutic modalities against liver
cancer. However, further studies are required to validate the
interacted role of miR-374b and PD-1 expression and the effect of
PD-1 on liver cancer growth in vivo using various liver
cancer cell lines. Furthermore, it will be necessary to further
explore the functions and mechanisms underlying miR-374b and PD-1
on CIK cells in liver cancer.
Acknowledgements
The present study was supported by the Key Research
and Development Project of Hainan Province from the Hainan Province
Science and Technology Department (grant no. ZDYF2016107).
References
|
1
|
El-Serag HB: Epidemiology of viral
hepatitis and hepatocellular carcinoma. Gastroenterology.
142:1264–1273.e1261. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sawyers CL, Abate-Shen C, Anderson KC,
Barker A, Baselga J, Berger NA, Foti M, Jemal A, Lawrence TS, Li
CI, et al: AACR Cancer progress report 2013. Clin Cancer Res.
19:S4–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang FS, Liu MX, Zhang B, Shi M, Lei ZY,
Sun WB, Du QY and Chen JM: Antitumor activities of human autologous
cytokine-induced killer (CIK) cells against hepatocellular
carcinoma cells in vitro and in vivo. World J Gastroenterol.
8:464–468. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li XD, Xu B, Wu J, Ji M, Xu BH, Jiang JT
and Wu CP: Review of Chinese clinical trials on CIK cell treatment
for malignancies. Clin Transl Oncol:. 14:102–108. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schmidt-Wolf IG, Negrin RS, Kiem HP, Blume
KG and Weissman IL: Use of a SCID mouse/human lymphoma model to
evaluate cytokine-induced killer cells with potent antitumor cell
activity. J Exp Med. 174:139–149. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen L: Co-inhibitory molecules of the
B7-CD28 family in the control of T-cell immunity. Nat Rev Immunol.
4:336–347. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Freeman GJ, Long AJ, Iwai Y, Bourque K,
Chernova T, Nishimura H, Fitz LJ, Malenkovich N, Okazaki T and
Byrne MC: Engagement of the PD-1 immunoinhibitory receptor by a
novel B7 family member leads to negative regulation of lymphocyte
activation. J Exp Med. 192:1027–1034. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bengsch B, Martin B and Thimme R:
Restoration of HBV-specific CD8+ T cell function by PD-1 blockade
in inactive carrier patients is linked to T cell differentiation. J
Hepatol. 61:1212–1219. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Boni C, Fisicaro P, Valdatta C, Amadei B,
Di Vincenzo P, Giuberti T, Laccabue D, Zerbini A, Cavalli A,
Missale G, et al: Characterization of hepatitis B virus
(HBV)-specific T-cell dysfunction in chronic HBV infection. J
Virol. 81:4215–4225. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fisicaro P, Valdatta C, Massari M, Loggi
E, Biasini E, Sacchelli L, Cavallo MC, Silini EM, Andreone P,
Missale G and Ferrari C: Antiviral intrahepatic T-cell responses
can be restored by blocking programmed death-1 pathway in chronic
hepatitis B. Gastroenterology. 138:682–693, 693.e1-4. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mühlbauer M, Fleck M, Schütz C, Weiss T,
Froh M, Blank C, Schölmerich J and Hellerbrand C: PD-L1 is induced
in hepatocytes by viral infection and by interferon-alpha and
-gamma and mediates T cell apoptosis. J Hepatol. 45:520–528. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng G, Li S, Wu W, Tan X, Chen Y and Chen
Z: PD-1 upregulation is associated with HBV-specific T cell
dysfunction in chronic hepatitis B patients. Mol Immunol.
45:963–970. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tzeng HT, Tsai HF, Liao HJ, Lin YJ, Chen
L, Chen PJ and Hsu PN: PD-1 blockage reverses immune dysfunction
and hepatitis B viral persistence in a mouse animal model. PloS
One. 7:e391792012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ye P, Weng ZH, Zhang SL, Zhang JA, Zhao L,
Dong JH, Jie SH, Pang R and Wei RH: Programmed death-1 expression
is associated with the disease status in hepatitis B virus
infection. World J Gastroenterol. 14:4551–4557. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hamid O, Robert C, Daud A, Hodi FS, Hwu
WJ, Kefford R, Wolchok JD, Hersey P, Joseph RW, Weber JS, et al:
Safety and tumor responses with lambrolizumab (anti-PD-1) in
melanoma. N Engl J Med. 369:134–144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Topalian SL, Hodi FS, Brahmer JR,
Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD,
Sosman JA, Atkins MB, et al: Safety, activity, and immune
correlates of anti-PD-1 antibody in cancer. N Engl J Med.
366:2443–2454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Topalian SL, Sznol M, McDermott DF, Kluger
HM, Carvajal RD, Sharfman WH, Brahmer JR, Lawrence DP, Atkins MB,
Powderly JD, et al: Survival, durable tumor remission, and
long-term safety in patients with advanced melanoma receiving
nivolumab. J Clin Oncol. 32:1020–1030. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Duraiswamy J, Freeman GJ and Coukos G:
Therapeutic PD-1 pathway blockade augments with other modalities of
immunotherapy T-cell function to prevent immune decline in ovarian
cancer. Cancer Res. 73:6900–6912. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Topalian SL, Drake CG and Pardoll DM:
Targeting the PD-1/B7-H1(PD-L1) pathway to activate anti-tumor
immunity. Curr Opin Immunol. 24:207–212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gozuacik D, Akkoc Y, Ozturk DG and Kocak
M: Autophagy-Regulating microRNAs and Cancer. Front Oncol.
7:652017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jasinski-Bergner S, Mandelboim O and
Seliger B: The role of microRNAs in the control of innate immune
response in cancer. J Natl Cancer Inst. 106(pii):
dju2572014.PubMed/NCBI
|
|
23
|
Atarod S and Dickinson AM: MicroRNAs: The
Missing Link in the Biology of Graft-Versus-Host Disease? Front
Immunol. 4:4202013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu XM and Zhang HJ: miRNAs as new
molecular insights into inflammatory bowel disease: Crucial
regulators in autoimmunity and inflammation. World J Gastroenterol.
22:2206–2218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gong H, Cao Y, Han G, Zhang Y, You Q, Wang
Y and Pan Y.: p53/microRNA-374b/AKT1 regulates colorectal cancer
cell apoptosis in response to DNA damage. Int J Oncol.
50:1785–1791. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu X, Li S, Xu X, Wu S, Chen R, Jiang Q,
Li Y and Xu Y: The potential value of miR-1 and miR-374b as
biomarkers for colorectal cancer. Int J Clin Exp Pathol.
8:2840–2851. 2015.PubMed/NCBI
|
|
27
|
Hu S, Bao H, Xu X, Zhou X, Qin W, Zeng C
and Liu Z: Increased miR-374b promotes cell proliferation and the
production of aberrant glycosylated IgA1 in B cells of IgA
nephropathy. FEBS Lett. 589:4019–4025. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liao YY, Tsai HC, Chou PY, Wang SW, Chen
HT, Lin YM, Chiang IP, Chang TM, Hsu SK, Chou MC, et al: CCL3
promotes angiogenesis by dysregulation of miR-374b/VEGF-A axis in
human osteosarcoma cells. Oncotarget. 7:4310–4325. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ma Z, Sun X, Xu D, Xiong Y and Zuo B:
MicroRNA, miR-374b, directly targets Myf6 and negatively regulates
C2C12 myoblasts differentiation. Biochem Biophys Res Commun.
467:670–675. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Qian D, Chen K, Deng H, Rao H, Huang H,
Liao Y, Sun X, Lu S, Yuan Z, Xie D and Cai Q: MicroRNA-374b
suppresses proliferation and promotes apoptosis in T-cell
lymphoblastic lymphoma by repressing AKT1 and Wnt-16. Clin Cancer
Res. 21:4881–4891. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schreiber R, Mezencev R, Matyunina LV and
McDonald JF: Evidence for the role of microRNA 374b in acquired
cisplatin resistance in pancreatic cancer cells. Cancer Gene Ther.
23:241–245. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie J, Tan ZH, Tang X, Mo MS, Liu YP, Gan
RL, Li Y, Zhang L and Li GQ: MiR-374b-5p suppresses RECK expression
and promotes gastric cancer cell invasion and metastasis. World J
Gastroenterol. 20:17439–17447. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Crispe IN: Hepatic T cells and liver
tolerance. Nat Rev Immunol. 3:51–62. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Protzer U, Maini MK and Knolle PA: Living
in the liver: Hepatic infections. Nat Rev Immunol. 12:201–213.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McDermott DF and Atkins MB: PD-1 as a
potential target in cancer therapy. Cancer Med. 2:662–673.
2013.PubMed/NCBI
|
|
37
|
Thomson AW and Knolle PA:
Antigen-presenting cell function in the tolerogenic liver
environment. Nat Rev Immunol. 10:753–766. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Odorizzi PM, Pauken KE, Paley MA, Sharpe A
and Wherry EJ: Genetic absence of PD-1 promotes accumulation of
terminally differentiated exhausted CD8+ T cells. J Exp Med.
212:1125–1137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Spranger S, Koblish HK, Horton B, Scherle
PA, Newton R and Gajewski TF: Mechanism of tumor rejection with
doublets of CTLA-4, PD-1/PD-L1, or IDO blockade involves restored
IL-2 production and proliferation of CD8(+) T cells directly within
the tumor microenvironment. J Immunother Cancer. 2:32014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Linedale R, Schmidt C, King BT, Ganko AG,
Simpson F, Panizza BJ and Leggatt GR: Elevated frequencies of CD8 T
cells expressing PD-1, CTLA-4 and Tim-3 within tumour from
perineural squamous cell carcinoma patients. PloS One.
12:e01757552017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu H, Wang Y, Zeng Q, Zeng YQ, Liang CL,
Qiu F, Nie H and Dai Z: Suppression of allograft rejection by
CD8+CD122+PD-1+ Tregs is dictated by their Fas ligand-initiated
killing of effector T cells versus Fas-mediated own apoptosis.
Oncotarget. 8:24187–24195. 2017.PubMed/NCBI
|
|
42
|
Schmittnaegel M, Rigamonti N, Kadioglu E,
Cassará A, Wyser Rmili C, Kiialainen A, Kienast Y, Mueller HJ, Ooi
CH, Laoui D and De Palma M: Dual angiopoietin-2 and VEGFA
inhibition elicits antitumor immunity that is enhanced by PD-1
checkpoint blockade. Sci Transl Med. 9(pii): eaak96702017.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Daskivich TJ and Belldegrun A: Words of
wisdom. Re: Safety, activity, and immune correlates of anti-PD-1
antibody in cancer. Eur Urol. 67:816–817. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hato T, Goyal L, Greten TF, Duda DG and
Zhu AX: Immune checkpoint blockade in hepatocellular carcinoma:
current progress and future directions. Hepatology. 60:1776–1782.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yongchun Z, Linwei T, Xicai W, Lianhua Y,
Guangqiang Z, Ming Y, Guanjian L, Yujie L and Yunchao H:
MicroRNA-195 inhibits non-small cell lung cancer cell
proliferation, migration and invasion by targeting MYB. Cancer
Lett. 347:65–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Aleckovic M and Kang Y: Regulation of
cancer metastasis by cell-free miRNAs. Biochim Biophys Acta.
1855:24–42. 2015.PubMed/NCBI
|
|
48
|
Di Leva G, Garofalo M and Croce CM:
MicroRNAs in cancer. Annu Rev Pathol. 9:287–314. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang Y, Yang P and Wang XF:
Microenvironmental regulation of cancer metastasis by miRNAs.
Trends Cell Biol. 24:153–160. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mao B and Wang G: MicroRNAs involved with
hepatocellular carcinoma (Review). Oncol Rep. 34:2811–2820. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Giordano S and Columbano A: MicroRNAs: New
tools for diagnosis, prognosis, and therapy in hepatocellular
carcinoma? Hepatology. 57:840–847. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang G, Li N, Li Z, Zhu Q, Li F, Yang C,
Han Q, Lv Y, Zhou Z and Liu Z: microRNA-4717 differentially
interacts with its polymorphic target in the PD1 3′ untranslated
region: A mechanism for regulating PD-1 expression and function in
HBV-associated liver diseases. Oncotarget. 6:18933–18944.
2015.PubMed/NCBI
|