Introduction
Vincristine is one of the most common anticancer
drugs clinically used for the management of leukemia, lymphoma and
primary brain cancer (1–3). It can induce neurotoxicity that
manifests as painful neuropathy, which significantly decreases the
quality of life in patients (4–6). Although
efforts have been made to identify the pathogenesis of
vincristine-induced neuropathic pain, the mechanisms underlying
this remain largely unknown.
Peripheral sensory neurons are more susceptible to
vincristine toxicity than central nervous system (CNS) neurons
owing to the absence of blood-nerve barrier (7,8). Transient
receptor potential cation channel subfamily V member 1 (TRPV1) is a
ligand-gated non-selective cation channel which is prominently
expressed in nociceptive dorsal root ganglia (DRG) neurons
(9,10). Accumulating evidence has suggested
that TRPV1 receptor expression and function in DRG neurons are
greatly sensitized following tissue inflammation or nerve injury
(11,12). Blockade or knockdown of TRPV1
attenuates mechanic allodynia or thermal hyperalgesia in several
models of pain (13–15). However, whether TRPV1 contributes to
the development of vincristine-induced pain hypersensitivity
remains unclear.
Previous studies have demonstrated that glia
activation and pronociceptive substances, such as proinflammatory
interleukins and tumor necrosis factor (TNF-α), in the spinal cord
contribute to vincristine-induced neuropathic pain (16,17).
However, it remains unknown whether TNF-α is implicated in the
sensitization of primary sensory neurons in vincristine-induced
neuropathy. In addition, TNF-α is well-known to potentiate TRPV1
activity, leading to mechanical allodynia and thermal hyperalgesia
in DRGs (18,19). Therefore, the present study aimed to
investigate whether peripheral TNF-α is responsible for the
sensitization of TRPV1 following vincristine administration in
vivo. The results indicated that vincristine administration
induced the upregulation of TRPV1 protein expression and current
density in DRG nociceptive neurons. TNF-α sensitized TRPV1 by
promoting its expression, thus leading to mechanical allodynia and
thermal hyperalgesia in vincristine-treated rats. Taken together,
these findings suggest that targeting the TNF-α signaling pathway
may be an effective approach for treating vincristine-induced
pain.
Materials and methods
Animals
Experiments were performed on 195 adult male
Sprague-Dawley rats aged 2 months (weight, 220±20 g) from Beijing
Weitong Lihua Experimental Animal Co., Ltd. (Beijing, China). All
rats were housed in separated cages with free access to food and
water under a 12-h light/dark cycle (lights on from 8:00 a.m. to
8:00 p.m.). The room temperature was kept at 24±1°C, with a
relative humidity of 50±5%. All animal experimental procedures were
conducted in accordance with the guidelines of the International
Association for the Study of Pain (20) and were approved by the Animal
Experimentation Ethics Committee (AEEC) of Henan University
(Kaifeng, China). Vincristine was used to induce neuropathy as
previously described (20). Briefly,
75 µg/kg of vincristine sulfate (Hospira, Lake Forest, IL, USA) was
administered once daily intraperitoneally (i.p.) for 5 consecutive
days, paused for 2 days, then subsequently administered for another
5 consecutive days. For the control group, the same volume of
normal saline was administered by the same schedule.
Behavioral analysis
The 50% paw withdrawal threshold (PWT) and paw
withdrawal latency (PWL) were performed as described in our
previous studies (11,21). All behavioral experiments were
performed blinded to treatment group. For mechanical sensitivity,
all rats were placed in transparent boxes on an elevated metal mesh
floor and allowed a 20 min period for habituation prior to
examination. A series of von Frey hairs (0.41–15.1 g; Stoelting
Co., Wood Dale, IL, USA) were applied perpendicular to the plantar
surface for 3–5 sec of each hind paw. The 50% PWT was determined
using the Dixon's up-down method (22). For testing thermal nociception, all
rats were allowed to acclimate for a minimum of 30 min in a
transparent, square, bottomless acrylic box (22×12×22 cm). Then,
PWLs were measured with radiant light focused on the left plantar
surface of the hind paw. A maximal cutoff time of 30 sec was
applied to avoid unnecessary tissue damage. The measurements of PWL
were repeated three times with 3–5 min intervals between
consecutive tests.
The first behavioral experiment was performed to
examine whether vincristine induced nociceptive behaviors in normal
rats. Vincristine (75 µg/kg, i.p. injection) was administered once
a day for 5 days, paused for 2 days, then resumed for an additional
5 days. Pain behaviors were evaluated prior to drug injection and
on days 1, 3, 7, 10, 14, and 21 post-vincristine
administration.
To examine whether inhibition of TRPV1 with
capsazepine (CPZ) attenuated vincristine-induced nociceptive
behaviors in normal rats, effects of CPZ (30 mg/kg; i.p. injection)
were measured at 2, 6, 12, 24 and 48 h post-administration.
To explore whether inhibition of TNF-α synthesis
alleviated vincristine-induced nociceptive behaviors, the TNF-α
synthesis inhibitor, thalidomide (50 mg/kg), was injected i.p. 2 h
prior to vincristine administration once a day for 5 days, paused
for 2 days, then resumed for an additional 5 days, and pain
hypersensitivity was examined on day 14.
Western blot analysis
Under deep anesthesia with isoflurane (2–3% in 1
l/min oxygen), the rats L4-L5 DRGs were removed and stored at −80°C
until use. Samples were homogenized in ice-chilled lysis buffer
containing 50 mM Tris (pH 8.0), 150 mM NaCl, 1% NP40, 0.5% sodium
deoxycholate, 0.1% SDS and protease inhibitor cocktail. Then, the
homogenates were centrifuged at 10,000 × g for 10 min at 4°C.
Following centrifugation, the supernatant was collected and the
concentration of total protein was determined with a bicinchoninic
acid assay kit (Pierce; Thermo Fisher Scientific, Inc., Waltham,
MA, USA). Equal amount of protein samples were denatured and then
analyzed by western blotting.
Preparation of membrane fractions was performed in
DRG neurons as previously described (11,23). In
brief, L4 and L5 DRGs were harvested and homogenized in ice-cold
lysis buffer containing 10 mM Tris (pH 8.1), 0.3 M sucrose, 2 mM
EDTA, 1 mM DTT, 1 mM PMSF and protease inhibitor cocktail.
Homogenates were placed on ice for 1 h before centrifugation at
1,000 × g for 7 min at 4°C to remove nuclei and intact cells. The
pellet was rehomogenized and spun again under the same conditions.
The supernatants from the two low-speed spins were combined and
centrifuged at 120,000 × g for 1 h at 4°C. The pellet, containing
the total membrane fraction, was suspended in 0.2 M KCl/10 mM HEPES
(pH 7.4). To solubilize the membrane fraction, an equal volume of
5% Triton X-100/10 mM HEPES (pH 7.4) was added to the samples and
the suspension was kept on ice for 1 h. The unsolubilized material
was pelleted by centrifugation at 10,000 × g for 10 min at 4°C, and
the supernatant was analyzed by western blotting.
For western blot analysis, Equal amounts of protein
samples (40 µg) were separated through SDS-PAGE using 10% running
gels and then transferred to polyvinylidene fluoride membranes.
After blocking with 5% nonfat milk in TBST (20 mM Tris-HCl, pH 7.5,
150 mM NaCl and 0.05% Tween-20) for 60 min at room temperature, the
membrane was probed with the following primary antibodies overnight
at 4°C: anti-TRPV1 (1:200; cat. no. sc-28759; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), anti-β-actin (1:1,000; cat.
no. sc-47778; Santa Cruz Biotechnology, Inc.), anti-TNF-α (1:500;
cat. no. sc-52746; Santa Cruz Biotechnology, Inc.), and
anti-transferrin receptor (TfR; 1:1,000; cat. no. 13-6800; Thermo
Fisher Scientific, Inc.). The blots were washed in TBST and then
incubated in horseradish peroxidase-conjugated goat anti-rabbit
(1:1,000, sc-2054; Santa Cruz Biotechnology, Inc.)/mouse IgG
secondary antibody (1:1,000, sc-2973; Santa Cruz Biotechnology,
Inc.) for 1 h at room temperature. Protein bands were visualized
using an enhanced chemiluminescence detection kit (Pierce; Thermo
Fisher Scientific, Inc.) followed by autoradiography using
Hyperfilm MP (GE Healthcare Life Sciences, Little Chalfont, UK).
The average density of each band was quantified using Quality One
system (version 4.6.2; Bio-Rad Laboratories, Inc., Hercules, CA,
USA) and normalized with that of β-actin or TfR bands.
Acute dissociation of DRG neurons
Neurons were isolated from ipsilateral L4 and L5 DRG
of adult rats using previously described methods (11). Briefly, DRGs were removed from each
rat and minced in cold, oxygenated Dulbecco's modified Eagle's
medium (Gibco; Thermo Fisher Scientific, Inc.). The DRGs were
digested with collagenase type IA (3 mg/ml; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) for 45 min, followed by 0.25% trypsin
(Sigma-Aldrich; Merck KGaA) for another 12 min at 37°C. After
terminating the enzymatic treatment by adding DMEM/10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.), ganglia were
dissociated by trituration with fire-polished glass pipettes.
Isolated cells were plated on poly-D-lysine-coated glass coverslips
placed in 24-well sterile tissue culture plates, and kept in a 5%
CO2 incubator at 37°C for 2 h prior to patch clamp
recording.
Whole-cell patch recording of TRPV1
current in dissociated DRG neurons
Whole-cell patch recording was performed as
previously described (11). Two h
after plating, the dissociated DRG neurons were perfused with
extracellular solution containing 128 mM NaCl, 5.4 mM KCl, 1.8 mM
CaCl2, 5 mM MgCl2, 5.55 mM glucose, and 20 mM
HEPES, adjusted to pH 7.4 with NaOH. Patch pipettes had a
resistance of 5–8 MΩ when filled with internal solution containing
100 mM KCl, 10 mM EGTA, 40 mM HEPES, 5 mM MgCl2, and 1
mM Na2ATP, adjusted to pH 7.3 with KOH. Under
voltage-clamp recording, neurons were clamped at −70 mV, and series
resistance was compensated to >75%. The membrane capacitance was
read from the amplifier by patch master software (version 2×69;
HEKA Elektronik Dr Schulze GmbH, Lambrecht/Pfalz, Germany) to
determine the size of neurons and to calculate the current density.
The agonist-evoked TRPV1 currents were measured by adding capsaicin
(0.5 µM for 3 sec). Origin software 8.5 (OriginLab Corporation,
Northampton, MA, USA) was used for data analysis.
Statistical analysis
Statistical analyses were performed with GraphPad
Prism 5 for Windows (GraphPad Software, Inc., La Jolla, CA, USA).
All data were presented as mean ± standard error of the mean.
One-way analysis of variance (ANOVA) followed by Tukey's post hoc
test or two-way ANOVA followed by Bonferroni's post hoc test were
used for multiple comparisons. P<0.05 was considered to indicate
a statistically significant difference.
Results
Vincristine-evoked mechanical
allodynia and thermal hyperalgesia
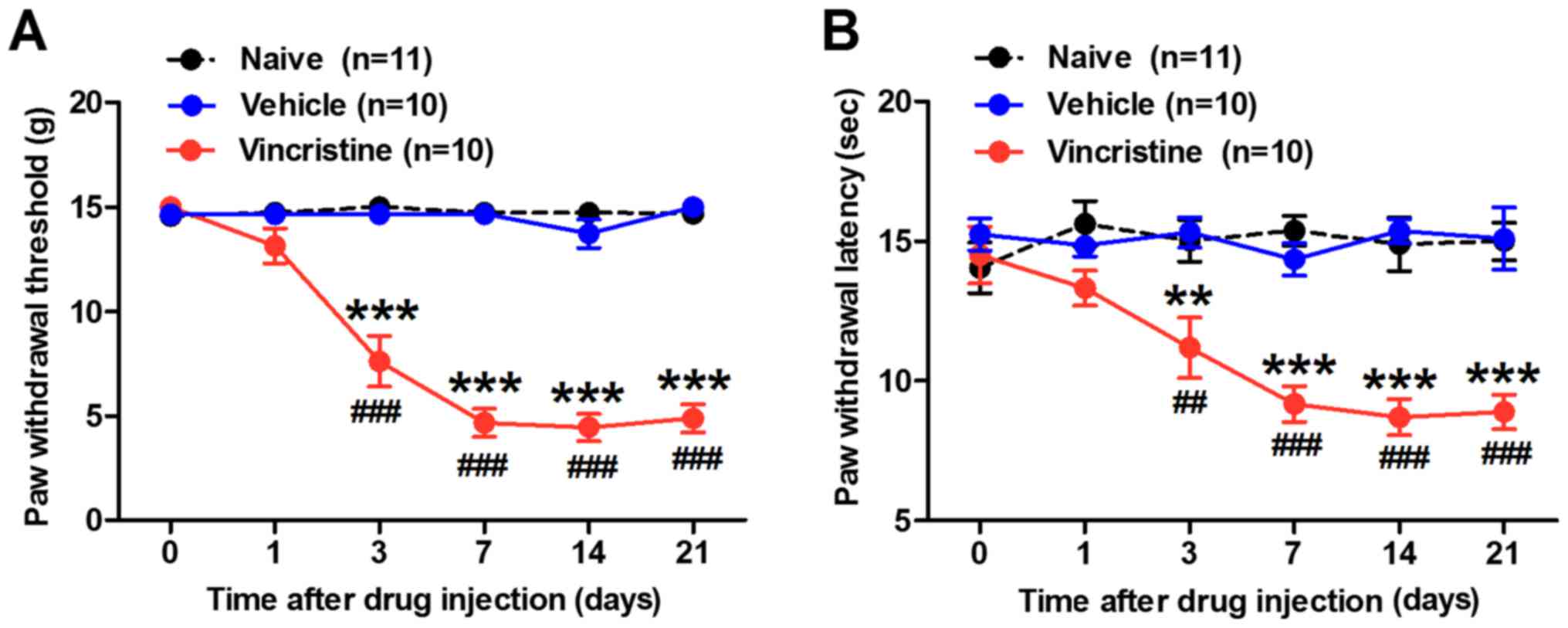
To confirm the establishment of the
vincristine-induced pain model, the 50% PWT and PWL to the radiant
heat stimulation were measured prior to vincristine administration
and on days 1, 3, 7, 10, 14, and 21 post-administration. Consistent
with previous reports (17,20), the results demonstrated that rats in
the vincristine-treated group exhibited significant mechanical
allodynia and thermal hyperalgesia (Fig.
1). For example, the 50% PWT to von Frey filaments in the left
hind paw in the vincristine-treated rats decreased from 15.00 g to
4.66±0.59 g on day 7 and 4.8±0.67 g on day 21 post-vincristine
administration (P<0.001; Fig. 1A).
The PWL in the vincristine-treated rats decreased from 14.50±1.02
sec to 9.17±0.64 sec on day 7 and 8.88±0.63 sec on day 21
post-administration (Fig. 1B). By
contrast, the 50% PWT and PWL of the left hind paw in the
vehicle-injected or naive rats exhibited no significant differences
during the time course of the experiments (Fig. 1A and B, respectively).
Functional upregulation of TRPV1 in
DRGs in vincristine-treated rats
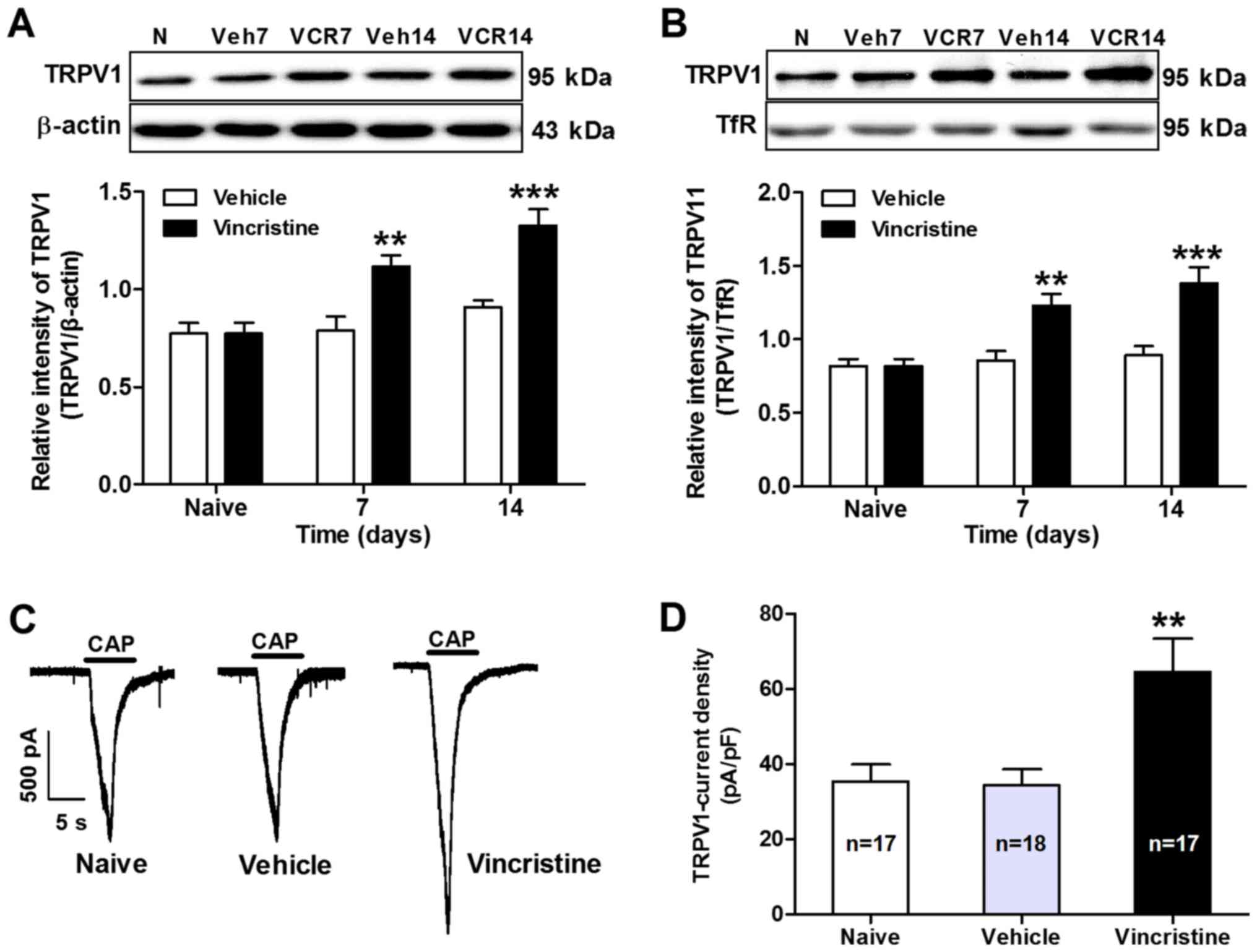
Functional upregulation of TRPV1 has been implicated
in different models of pathological pain (11,15,18). To
evaluate whether TRPV1 may contribute to vincristine-induced pain,
the total protein expression levels of TRPV1 in DRGs collected from
vehicle or vincristine-treated rats were examined. Using western
blot analysis, the results demonstrated that the expression levels
of TRPV1 total protein in L4 and L5 DRGs increased significantly
from day 7 to day 14 in vincristine-treated rats compared with
vehicle-treated rats (P<0.05; n=7; Fig. 2A). Considering that membrane insertion
of TRPV1 receptors in DRG neurons is important for the development
of pain hypersensitivity, expression of membrane-bound TRPV1
receptors in DRGs was examined following vincristine
administration. Using western blot analysis of membrane-specific
fractions, the results demonstrated that the membrane expression of
TRPV1 in L4 and L5 DRGs increased significantly from day 7 to day
14 in vincristine-treated rats compared with vehicle-treated rats
(P<0.01; Fig. 2B). To determine
the functional alteration of TRPV1 receptors in nociceptive DRG
neurons in vincristine-treated rats, the TRPV1 receptor currents in
DRG neurons were then measured with whole-cell patch clamp
recording. As expected, TRPV1 current density was also
significantly increased in DRG neurons from vincristine-treated
rats compared with vehicle-treated rats (64.64±8.81 in
vincristine-treated rats vs. 34.36±4.29 in vehicle-treated rats;
P<0.01; Fig. 2C and D).
CPZ alleviates mechanical pain
hypersensitivity in vincristine-treated rats
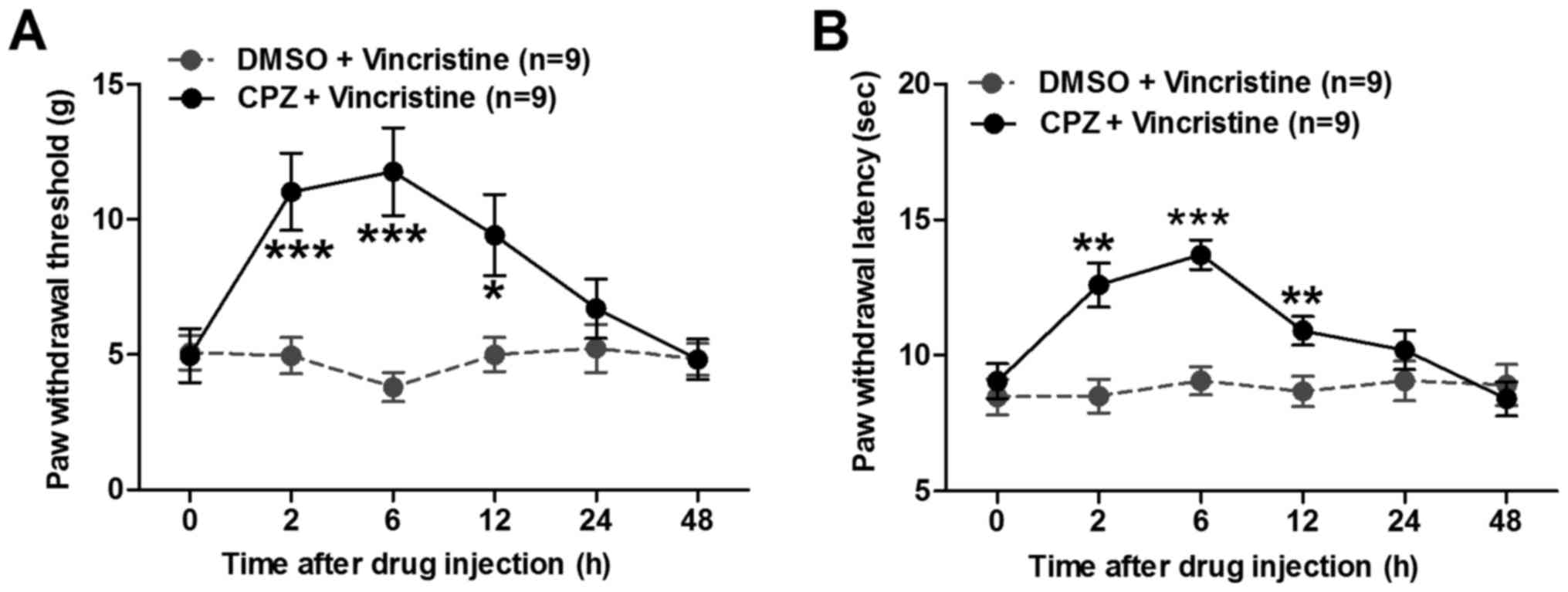
To examine whether the functional potentiation of
TRPV1 may have induced pain hypersensitivity in vincristine-treated
rats, the present study examined whether CPZ, a TRPV1 receptor
antagonist, could attenuate vincristine-induced pain in rats. CPZ
or vehicle (DMSO) was intraperitoneally delivered to
vincristine-treated rats on day 14, and both the PWT and PWL were
measured at 2, 6, 12, 24 and 48 h post-CPZ or vehicle
administration. As illustrated in Fig.
3, administration of CPZ could prominently reverse the
vincristine-induced decrease in both PWT (Fig. 3A) and PWL (Fig. 3B). These data provided direct evidence
that TRPV1 receptors contribute to vincristine-induced pain
hypersensitivity in rats.
Sensitization of TRPV1 by TNF-α
orchestrates the development of vincristine-induced pain
Previous studies have demonstrated that TNF-α is a
key mediator in inflammatory and neuropathic hyperalgesia (24,25). An
increase of TNF-α protein levels in the spinal cord has been
observed following administration of vincristine in rats, and
inhibition of TNF-α synthesis attenuated neuropathic pain caused by
vincristine (16,17). To explore whether TNF-α has a role in
promoting functional upregulation of TRPV1 in DRG neurons in
vincristine-treated rats, the present study first examined the
expression levels of TNF-α in the DRGs in vincristine-treated rats
using western blot analysis. As presented in Fig. 4A and B, expression of TNF-α in L4 and
L5 DRGs increased markedly from day 7 to day 14 in
vincristine-treated rats compared with vehicle-treated rats. Next,
the TNF-α synthesis inhibitor thalidomide (50 mg/kg) was
administered to the rats 2 h prior to vincristine administration
and TRPV1 expression was examined on day 14. Using western blot
analysis, the results demonstrated that the total (Fig. 4C) and membrane-bound (Fig. 4D) TRPV1 expression levels in L4 and L5
DRGs decreased significantly on day 14 post-thalidomide
administration. In addition, the pain behaviors were evaluated in
the thalidomide-treated rats. The results demonstrated that
thalidomide administration significantly inhibited the
vincristine-induced decreases in PWT (11.07±1.51 g in
thalidomide+vincristine-treated rats vs. 4.87±0.65 g in vincristine
alone-treated rats; P<0.001; Fig.
4E) and in PWL (11.98±0.99 sec in
thalidomide+vincristine-treated rats vs. 8.32±0.39 sec in
vincristine alone-treated rats; P<0.01; Fig. 4F) in vincristine-treated rats on day
14. These results indicated that TNF-α has a positive role in
promoting functional upregulation of TRPV1 in DRG neurons in
vincristine-induced neuropathic rats.
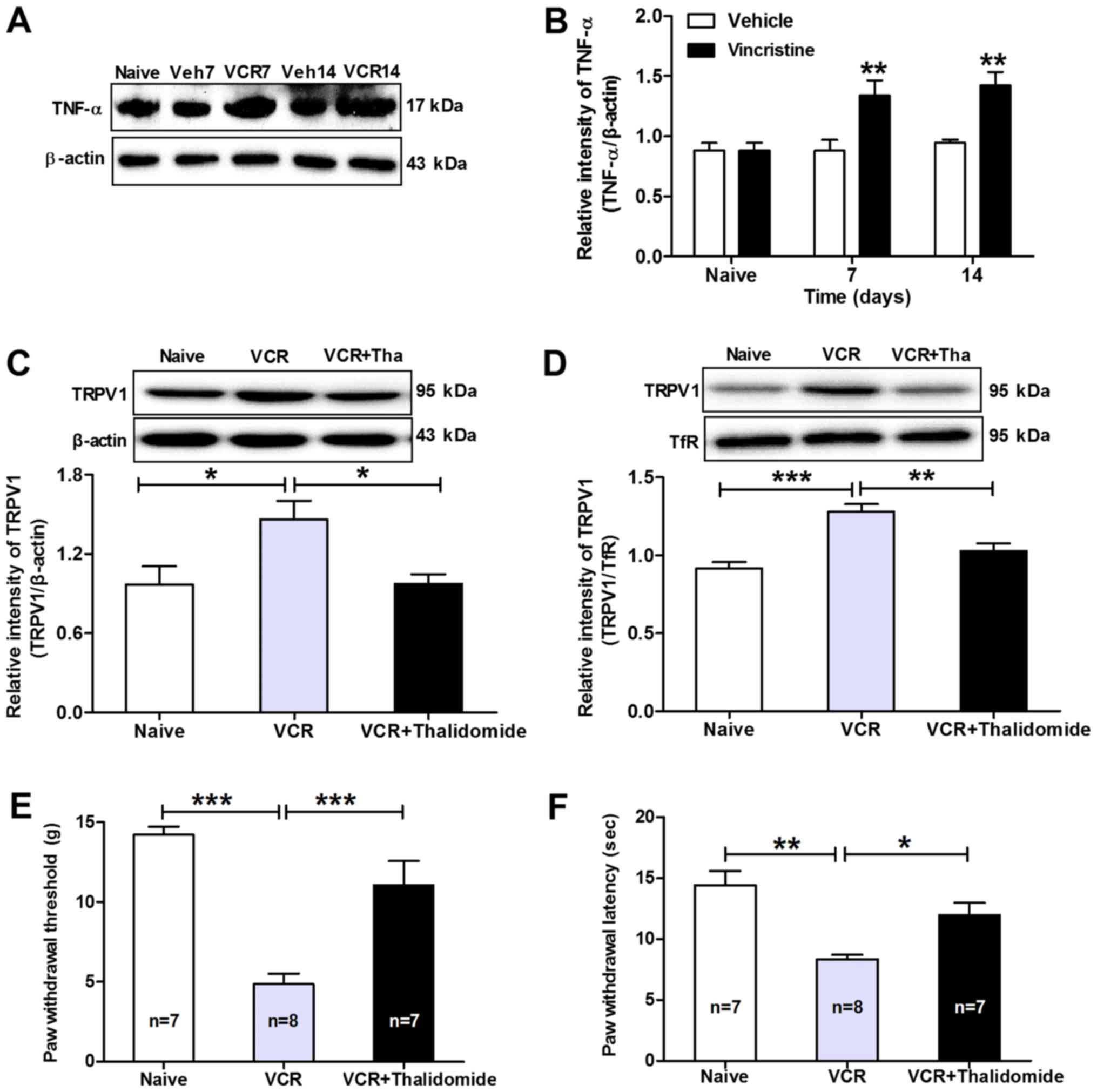 | Figure 4.TNF-α promotes functional
upregulation of TRPV1 in DRG neurons in vincristine-treated rats.
(A) Representative images and (B) quantification from western blot
analysis of TNF-α protein expression levels in the DRGs of
vincristine-treated rats. TNF-α protein expression levels increased
significantly from day 7 to day 14 in vincristine-treated rats
compared with vehicle-treated rats. **P<0.01 compared with
vehicle-treated group (n=5/group). (C) Western blot analysis of
TRPV1 total and (D) membrane-bound protein expression levels in
ipsilateral L4 and L5 DRGs following treatment with thalidomide in
vincristine-treated rats. β-actin and TfR were used as internal
controls for total and membrane-bound protein, respectively.
*P<0.05, **P<0.01 and ***P<0.001, with comparisons
indicated by brackets (n=5/group). (E) Effects of pre-treatment
with thalidomide on the mechanical allodynia and (F) thermal
hyperalgesia of vincristine-treated rats. Note that thalidomide
pre-treatment significantly prevented the vincristine-induced
decreases in paw withdrawal threshold and paw withdrawal latency in
vincristine rats. *P<0.05, **P<0.01 and ***P<0.001, with
comparisons indicated by brackets. TNF, tumor necrosis factor;
TRPV1, transient receptor potential cation channel subfamily V
member 1; DRG, dorsal root ganglions; TfR, transferrin receptor;
Veh, vehicle; VCR, vincristine. |
Discussion
Pain is an early and especially disabling symptom of
vincristine-induced peripheral neurotoxicity, which reduces the
quality of daily life in cancer patients (26,27). DRGs
are hypothesized to be one of the preferential sites in which
chemotherapy neurotoxicity occurs because of the absence of
blood-nerve barrier (7,8). As is well-known, one of the pathologic
mechanisms of pain is associated with sensitization of primary
sensory neurons. Cumulative evidence indicates that TRPV1
expression and function in DRGs is greatly sensitized following
tissue inflammation or nerve injury, thus leading to peripheral
sensitization and pain hypersensitivity (12,14).
Therefore, the present study examined the changes in TRPV1
expression and function in DRG sensory neurons. As expected, the
results revealed that the expression as well as the function of
TRPV1 receptors was significantly increased in DRGs in
vincristine-treated rats. Furthermore, administration of the TRPV1
antagonist CPZ significantly attenuated vincristine-induced
mechanic allodynia and thermal hyperalgesia. Collectively, these
results indicated that TRPV1 contributes to vincristine-induced
pain sensation. To the best of our knowledge, this is the first
report providing a direct link between functional upregulation of
TRPV1 and vincristine-induced pain behaviors.
A growing number of studies have demonstrated that
TNF-α serves a vital role in the development of pathological pain
(16,28,29). For
example, TNF-α is overexpressed in the spinal cord or DRGs when
chronic pain happens, while administration of TNF-α elicits ectopic
discharges in DRG neurons and pain hypersensitivity in normal
animals (16,30–32). In
addition, treatment with a TNF-α antibody or TNF receptor
antagonist ameliorates pain hypersensitivity in several models of
pathological pain (16,30–32). In
agreement with these reports, the present study also demonstrated
that TNF-α was upregulated in DRGs in vincristine-treated rats. In
addition, the present results demonstrated that thalidomide, an
inhibitor of TNF-α synthesis, attenuated vincristine-induced TRPV1
total and membrane protein expression. Obviously, TNF-α not only
promotes TRPV1 expression at a translational level, but may also
induce the directed TRPV1 trafficking to the surface membrane in
DRGs in vincristine-treated rats. Consistent with the present
results, emerging evidence has suggested that TNF-α overexpression
in sensory neurons results in increased cyclin-dependent kinase 5
activity which phosphorylates TRPV1 at Thr406 and
promotes its surface localization (19,33). In
addition to regulating TRPV1 membrane translocation, TNF-α has been
reported to increase TRPV1 total protein levels via a translational
regulation. For example, long-term incubation of DRG neurons with
TNF-α significantly increases the proportion of
TRPV1-immunoreactive neurons through the activation of
extracellular signal-regulated kinase signaling pathway (34). Hence, it can be concluded that TNF-α
sensitizes TRPV1 by increasing TRPV1 expression and by mobilizing
new channels to the neuronal surface in vincristine-induced pain
hypersensitivity. Taken together, the present results suggested
that TNF-α could sensitize TRPV1 by promoting its expression, thus
leading to mechanical allodynia and thermal hyperalgesia in
vincristine-treated rats. These findings may enhance the current
understanding of the pathophysiological mechanisms underlying
vincristine-induced pain.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Science
Foundation of Henan (grant no. 162300410039) and the Program for
Science and Technology, Department of Education of Henan Province
(grant no. 16A350013).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
YW, WL and JH conducted the electrophysiological
studies and participated in the design of the study. CF, HH and JH
participated in the behavioral test and performed the statistical
analysis. JW, XL and SW performed the western blotting, and part of
the behavioral test. SQX participated in the design of the study.
DF conceived of the study, participated in its design and
coordination, and drafted the article. TL performed the statistical
analysis and participated in the preparation of the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experimental procedures were carried out
in accordance with the guidelines of the International Association
for the Study of Pain and were approved by the Animal
Experimentation Ethics Committee of Henan University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nieder C, Mehta MP and Jalali R: Combined
radio- and chemotherapy of brain tumours in adult patients. Clin
Oncol (R Coll Radiol). 21:515–524. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Raj TA, Smith AM and Moore AS: Vincristine
sulfate liposomal injection for acute lymphoblastic leukemia. Int J
Nanomedicine. 8:4361–4369. 2013.PubMed/NCBI
|
|
3
|
Eden T, Pieters R and Richards S;
Childhood Acute Lymphoblastic Leukaemia Collaborative Group
(CALLCG), : Systematic review of the addition of vincristine plus
steroid pulses in maintenance treatment for childhood acute
lymphoblastic leukaemia-an individual patient data meta-analysis
involving 5,659 children. Br J Haematol. 149:722–733. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Quasthoff S and Hartung HP:
Chemotherapy-induced peripheral neuropathy. J Neurol. 249:9–17.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sioka C and Kyritsis AP: Central and
peripheral nervous system toxicity of common chemotherapeutic
agents. Cancer Chemother Pharmacol. 63:761–767. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hu C, Zhao YT, Zhang G and Xu MF:
Antinociceptive effects of fucoidan in rat models of
vincristine-induced neuropathic pain. Mol Med Rep. 15:975–980.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Allen DT and Kiernan JA: Permeation of
proteins from the blood into peripheral nerves and ganglia.
Neuroscience. 59:755–764. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Old EA, Nadkarni S, Grist J, Gentry C,
Bevan S, Kim KW, Mogg AJ, Perretti M and Malcangio M: Monocytes
expressing CX3CR1 orchestrate the development of
vincristine-induced pain. J Clin Invest. 124:2023–2036. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mitchell K, Bates BD, Keller JM, Lopez M,
Scholl L, Navarro J, Madian N, Haspel G, Nemenov MI and Iadarola
MJ: Ablation of rat TRPV1-expressing Adelta/C-fibers with
resiniferatoxin: Analysis of withdrawal behaviors, recovery of
function and molecular correlates. Mol Pain. 6:942010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han Q, Kim YH, Wang X, Liu D, Zhang ZJ,
Bey AL, Lay M, Chang W, Berta T, Zhang Y, et al: SHANK3 deficiency
impairs heat hyperalgesia and TRPV1 signaling in primary sensory
neurons. Neuron. 92:1279–1293. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fang D, Kong LY, Cai J, Li S, Liu XD, Han
JS and Xing GG: Interleukin-6-mediated functional upregulation of
TRPV1 receptors in dorsal root ganglion neurons through the
activation of JAK/PI3K signaling pathway: Roles in the development
of bone cancer pain in a rat model. Pain. 156:1124–1144.
2015.PubMed/NCBI
|
|
12
|
Labuz D, Spahn V, Celik MÖ and Machelska
H: Opioids and TRPV1 in the peripheral control of neuropathic
pain-Defining a target site in the injured nerve.
Neuropharmacology. 101:330–340. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morales-Lázaro SL, Llorente I,
Sierra-Ramírez F, López-Romero AE, Ortíz-Rentería M, Serrano-Flores
B, Simon SA, Islas LD and Rosenbaum T: Inhibition of TRPV1 channels
by a naturally occurring omega-9 fatty acid reduces pain and itch.
Nat Commun. 7:130922016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao X, Zhao XT, Xu LC, Yue LP, Liu FY,
Cai J, Liao FF, Kong JG, Xing GG, Yi M and Wan Y: Shp-1
dephosphorylates TRPV1 in dorsal root ganglion neurons and
alleviates CFA-induced inflammatory pain in rats. Pain.
156:597–608. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu Q, Zhang XM, Duan KZ, Gu XY, Han M, Liu
BL, Zhao ZQ and Zhang YQ: Peripheral TGF-β1 signaling is a critical
event in bone cancer-induced hyperalgesia in rodents. J Neurosci.
33:19099–19111. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kiguchi N, Maeda T, Kobayashi Y, Saika F
and Kishioka S: Involvement of inflammatory mediators in
neuropathic pain caused by vincristine. Int Rev Neurobiol.
85:179–190. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shen Y, Zhang ZJ, Zhu MD, Jiang BC, Yang T
and Gao YJ: Exogenous induction of HO-1 alleviates
vincristine-induced neuropathic pain by reducing spinal glial
activation in mice. Neurobiol Dis. 79:100–110. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Constantin CE, Mair N, Sailer CA,
Andratsch M, Xu ZZ, Blumer MJ, Scherbakov N, Davis JB, Bluethmann
H, Ji RR and Kress M: Endogenous tumor necrosis factor alpha
(TNFalpha) requires TNF receptor type 2 to generate heat
hyperalgesia in a mouse cancer model. J Neurosci. 28:5072–5081.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rozas P, Lazcano P, Piña R, Cho A, Terse
A, Pertusa M, Madrid R, Gonzalez-Billault C, Kulkarni AB and
Utreras E: Targeted overexpression of tumor necrosis factor-α
increases cyclin-dependent kinase 5 activity and TRPV1-dependent
Ca2+ influx in trigeminal neurons. Pain. 157:1346–1362.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kahng J, Kim TK, Chung EY, Kim YS and Moon
JY: The effect of thioctic acid on allodynia in a rat
vincristine-induced neuropathy model. J Int Med Res. 43:350–355.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang H, Fang D, Kong LY, Jin ZR, Cai J,
Kang XJ, Wan Y and Xing GG: Sensitization of neurons in the central
nucleus of the amygdala via the decreased GABAergic inhibition
contributes to the development of neuropathic pain-related
anxiety-like behaviors in rats. Mol Brain. 7:722014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chaplan SR, Bach FW, Pogrel JW, Chung JM
and Yaksh TL: Quantitative assessment of tactile allodynia in the
rat paw. J Neurosci Methods. 53:55–63. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tyrrell L, Renganathan M, Dib-Hajj SD and
Waxman SG: Glycosylation alters steady-state inactivation of sodium
channel Nav1.9/NaN in dorsal root ganglion neurons and is
developmentally regulated. J Neurosci. 21:9629–9637.
2001.PubMed/NCBI
|
|
24
|
Chen Y, Zhang Y, Huo Y, Wang D and Hong Y:
Adrenomedullin mediates tumor necrosis factor-α-induced responses
in dorsal root ganglia in rats. Brain Res. 1644:183–191. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu J.E.X..Liu H, Li F, Cao Y, Tian J and
Yan J: Tumor necrosis factor-alpha is a potential diagnostic
biomarker for chronic neuropathic pain after spinal cord injury.
Neurosci Lett. 595:30–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sisignano M, Baron R, Scholich K and
Geisslinger G: Mechanism-based treatment for chemotherapy-induced
peripheral neuropathic pain. Nat Rev Neurol. 10:694–707. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bosilkovska M, Ing Lorenzini K,
Uppugunduri CR, Desmeules J, Daali Y and Escher M: Severe
vincristine-induced neuropathic pain in a CYP3A5 nonexpressor with
reduced CYP3A4/5 activity: Case study. Clin Ther. 38:216–220. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gong SS, Li YX, Zhang MT, Du J, Ma PS, Yao
WX, Zhou R, Niu Y, Sun T and Yu JQ: Neuroprotective effect of
matrine in mouse model of vincristine-induced neuropathic pain.
Neurochem Res. 41:3147–3159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Walters ET: Neuroinflammatory
contributions to pain after SCI: Roles for central glial mechanisms
and nociceptor-mediated host defense. Exp Neurol. 258:48–61. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kochukov MY: Tumor necrosis factor-alpha
(TNF-alpha) enhances functional thermal and chemical responses of
TRP cation channels in human synoviocytes. Mol Pain. 5:492009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sacerdote P, Franchi S, Gerra G, Leccese
V, Panerai AE and Somaini L: Buprenorphine and methadone
maintenance treatment of heroin addicts preserves immune function.
Brain Behav Immun. 22:606–613. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sasaki M, Hashimoto S, Sawa T and Amaya F:
Tumor necrosis factor-alpha induces expression of C/EBP-beta in
primary afferent neurons following nerve injury. Neuroscience.
279:1–9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu J, Du J, Yang Y and Wang Y:
Phosphorylation of TRPV1 by cyclin-dependent kinase 5 promotes
TRPV1 surface localization, leading to inflammatory thermal
hyperalgesia. Exp Neurol. 273:253–262. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hensellek S, Brell P, Schaible HG, Bräuer
R and Segond von Banchet G: The cytokine TNFalpha increases the
proportion of DRG neurones expressing the TRPV1 receptor via the
TNFR1 receptor and ERK activation. Mol Cell Neurosci. 36:381–391.
2007. View Article : Google Scholar : PubMed/NCBI
|