Introduction
Human esophageal cancer (EC) is the third most
common digestive system malignancy worldwide, and is associated
with a high mortality rate (1). EC is
also one of the most common malignant tumors in China, ranking
third and fifth in causing male and female malignant tumors,
respectively (2). Esophageal squamous
cell carcinoma (ESCC) and esophageal adenocarcinoma (EA) are the
primary subtypes of EC; however, in China, ESCC accounts for
>90% of total cases (3). Despite
improved development of multimodal techniques including surgery,
chemotherapy and radiotherapy, the survival rate of patients with
ESCC remains poor owing to the high incidence of local invasion and
distant metastasis (3,4). Therefore, it is imperative to seek novel
therapeutic targets and effective biomarkers for ESCC.
Long non-coding RNAs (lncRNA) are non-protein-coding
transcripts >200 nucleotides (5,6). On the
basis of the proximity to nearby coding genes, lncRNAs are
categorized into 5 subclasses: Sense, antisense, intergenic,
bidirectional and intronic (7).
lncRNAs are involved in every aspect of cellular processes,
including cellular development, modulation of apoptosis, metastasis
and parental imprinting (8–10). Emerging evidence suggests that the
dysregulation of lncRNAs is linked to the development and
metastasis in several types of cancer, including EC (11–13). The
importance of lncRNAs in ESCC carcinogenesis has been recently
evidenced. Hundreds of ESCC-associated lncRNAs were identified,
some of which could be used as biomarkers for diagnosing or
prognosing ESCC (14–16). ESCC-associated lncRNAs remain to be
fully understood but research has progressed in this field: POU
class 3 homeobox 3 (POU3F3) promotes the malignant progression of
ESCC by binding the enhancer of zeste 2 polycomb repressive complex
2 subunit protein to promote the methylation of POU3F3 (17); colon cancer associated transcript 1
(non-protein coding) could modulate the histone methylation of the
promoter of sprouty RTK signaling antagonist 4 by binding polycomb
repressive complex 2 (PRC2) and suppressor of variegation 3–9
homolog 1 in the nucleus and promoted the expression of downstream
target gene homeobox B13 by adsorbing microRNA-7 in the cytoplasm
(18).
In a previous study, the novel lncRNA SET-binding
factor 2 (SBF2) antisense RNA1 (SBF2-AS1) was identified and was
demonstrated to be upregulated in non-small cell lung cancer
(NSCLC), and was also able to promote proliferation of NSCLC cells
(19). SBF2-AS1, located at the
11p15.1 locus, is a 2,708-nt antisense RNA to SBF2. A previous
study identified that SBF2-AS1 was able to bind with PRC2 and
guided PRC2 to the promoter of CDKN1A and therefore decreased
CDKN1A expression (19). However, the
exact role of SBF2-AS1 in ESCC remains unclear. Therefore, the
investigated the expression and biological function(s) of SBF2-AS1
in ESCC in vitro were investigated in the present study.
Materials and methods
Patients and tissue samples
All primary ESCC tissues and paired normal tissues
were collected from patients who had undergone surgery at the
Department of Thoracic Surgery, Cancer Institute of Jiangsu
Province (Nanjing, China) between May 2013 and May 2014. No
patients received radiotherapy or chemotherapy prior to surgical
resection. Experienced pathologists confirmed all tumors and paired
normal tissues, and clinical/pathological characteristics of each
patient were collected. Written informed consent was obtained from
all patients and the Ethics Committee of Cancer Institute of
Jiangsu Province approved the present study.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analyses
Total RNA was extracted from tissues or cultured
esophageal cell lines (HesEpiC; ScienCell Research Laboratories,
Inc., San Diego, CA, USA), KYSE-410 (Department of Radiology of The
First Affiliated Hospital of Nanjing Medical University, Jiangsu,
Nanjing, China), ECA-109 (Shanghai Institutes for Biological
Science, Shanghai, China) and TE-1 (Shanghai Institutes for
Biological Science, Shanghai, China) with TRIzol reagent (Thermo
Fisher Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol. The concentration and A260/280
ratio were measured using a NanoDrop 2000 spectrophotometer (Thermo
Fisher Scientific, Inc.). A 500 ng amount of total RNA was
reverse-transcribed in a final volume of 10 µl using random primers
under standard conditions using the Prime Script RT Master Mix
(Takara Biotechnology Co., Ltd., Dalian, China), according to the
manufacturer's protocol; the reverse transcription reactions were
performed at 25°C for 5 min, followed by 42°C for 60 min and 70°C
for 5 min. qPCR was performed using the SYBR Green Master Mix
(Thermo Fisher Scientific, Inc.) with 0.5 µl cDNA on a Veriti 96
Well Thermal Cycler (Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol, to determine relative levels of
SBF2-AS1. GAPDH and β-actin were used as internal controls.
Reaction mixtures were incubated at 95°C for 10 min, 40 cycles of
95°C for 15 sec, 60°C for 1 min and 72°C for 10 min. The fold
changes of individual genes were calculated using the
2−ΔΔCq method (20). The
following primer pairs were used for qPCR: SBF2-AS1 forward,
5′-CACGACCCAGAAGGAvGTCTAC-3′, and reverse,
5′-CCCGGTACCTTCCTGTCATA-3′; cyclin-dependent kinase 1A (CDKN1A)
forward, 5′-AGACCATGTGGACCTGTCACTG-3′, and reverse,
5′-GTTTGGAGTGGTAGAAATCTGTC-3′; GAPDH forward,
5′-CGCTCTCTGCTCTCCTGTTC-3′, and reverse,
5′-ATCCGTTGACTCCGACCTTCAC-3′; β-actin forward,
5′-CGCTCTCTGCTCCTCCTGTTC-3′, and reverse,
5′-ATCCGTTGACTCCGACCTTCAC-3′.
Nuclear mass separation
experiment
The subcellular localization of SBF2-AS1 was
detected using the PARIS™ kit (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. Firstly, the cells were
washed with ice-cold PBS twice. Subsequently, 1 ml ice-cold cell
lysis solution [containing 20 mmol/l HEPES (pH 7.5), 10 mmol/l KCl,
1.5 mmol/l MgCl2, 1 mmol/l EGTA, 1 mmol/l EDTA, 1 mmol/l
DTT, 0.1 mmol/l PMSF and 250 mmol/l sucrose] was added to the cells
and 3×106 cells were harvested with a cell scraper on
ice. The cell lysate was incubated on ice for 5 min. After
centrifuging at 500 × g for 5 min at 4°C, the precipitate
containing the nuclear RNA was isolated from the supernatant
containing the cytoplasmic RNA. Finally, the supernatant was
removed. The precipitate was washed with PBS twice and resuspended
using Nuclei EZ storage buffer (Thermo Fisher Scientific,
Inc.).
Cell culture
KYSE-410 cells were donated by the Department of
Radiology of The First Affiliated Hospital of Nanjing Medical
University (Jiangsu, Nanjing, China). ECA-109 and TE-1 cells were
purchased from Shanghai Institutes for Biological Science
(Shanghai, China). Normal human esophageal epithelial cell lines
(HEsEpiC) were purchased from ScienCell Research Laboratories, Inc.
KYSE-410 cells and HEECs were grown in RPMI-1640 medium (Nanjing
KeyGen Biotech Co., Ltd., Nanjing, China). ECA-109 and TE-1 cells
were grown in Dulbecco's modified Eagle's medium (DMEM; Nanjing
KeyGen Biotech Co., Ltd.), supplemented with penicillin (80
U/ml)-streptomycin (0.08 mg/ml) and 10% fetal bovine serum (FBS;
Thermo Fisher Scientific, Inc.). All cells were grown at 37°C in a
humidified 5% CO2 atmosphere.
siRNA transfection
ESCC cells (70–80% confluence in 6-well plates) were
transfected with 100 nM specific small interfering RNA (siRNA) or
100 nM negative control (NC) siRNA using Lipofectamine®
RNA interference (RNAi) MAX (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Following
transfection, cells were cultured for 48 h at 37°C in a humidified
5% CO2 atmosphere. The following siRNA primer sequences
were used: SBF2-AS1 sense (204 site), 5′-CAGAAGGAGUCUACUGCUAAG-3′,
and antisense, 5′-UAGCAGUAGACUCCUUCUGGG-3′, SBF2-AS1 sense (1021
site), 5′-GCAAGCCUGCAUGGUACAUTT-3′, and antisense,
5′-AUGUACCAUGCAGGCUUGCTT-3′.
Cell proliferation assay
Cell proliferation was assayed using the Cell
Counting Kit-8 (CCK-8) assay (Promega Corporation, Madison, WI,
USA), according to the manufacturer's protocol. Transfected cells
were plated in 96-well plates (3,000 cells/well). Cell
proliferation was measured every 24 h in accordance with the
manufacturer's protocol. In brief, 10 µl CCK-8 solution was added
to each well and incubated for 2 h at 37°C, and each solution was
measured spectrophotometrically at 450 nm as previously described
(21).
Clonogenic assay
A total of 100 transfected cells were placed into a
fresh 6-well plate and maintained in 2 ml medium containing 10%
FBS, replacing the medium every 3 days. After 2 weeks, cells were
immobilized with 4% paraformaldehyde at room temperature for 30 min
and stained with 0.1% crystal violet at room temperature for 15
min. Visible colonies were counted using light microscopy and a
fluorescence microscope (magnification, ×40) (22).
In vitro cell migration and invasion
assays
ECA-109 cells transfected with 100 nM si-SBF2-AS1 or
si-NC was harvested 24 h after transfection. For migration assays,
transfected cells (3×105) were plated in the upper
chamber of the Transwell assay inserts (EMD Millipore, Billerica,
MA, USA) containing 200 µl serum-free DMEM with a membrane without
Matrigel (8-mm pores). Lower chambers were filled with DMEM
containing 10% FBS. After 24 h of incubation at 37°C in a 5%
CO2 incubator, cells on the filter surface were fixed
with 100% methanol at room temperature for 15 min and stained with
0.1% crystal violet at room temperature for 15 min, and images were
captured using an Axio Vert.A1 microscope (magnification, ×100;
Carl Zeiss AG, Oberkochen, Germany). Cell numbers were calculated
using five random fields for each chamber. For invasion assays,
transfected cells (5×105) were plated in the upper
chamber with a Matrigel-coated membrane (BD Biosciences, Franklin
Lakes, NJ, USA) in 500 µl serum-free DMEM with 750 µl 10% FBS in
DMEM in the lower chambers. Invasion function was determined using
the number of transmigrated cells following 48 h incubation at 37°C
in a 5% CO2 incubator.
Flow cytometric analysis
Transfected cells were harvested after 24 h of
incubation at 37°C in a 5% CO2 incubator. ECA-109 cells
were stained with Annexin V and propidium iodide (PI) using Annexin
V-fluorescein isothiocyanate/PI apoptosis detection kits (BD
Biosciences) and examined using flow cytometry (FACScan; BD
Biosciences). Cells used for cell cycle analysis were stained with
propidium oxide using the Cycle Test Plus DNA reagent kit (BD
Biosciences) and analyzed using FACScan (BD Biosciences) as
described previously (23).
Western blot assay
Proteins were extracted 48 h after transfection.
Cells in each well were placed in 100 µl lysis buffer containing 1
mM sodium vanadate, 0.5 mM dithiothreitol, 1 mM
phenylmethylsulfonyl fluoride, 2 mM leupeptin, 2 mM aprotinin and 2
mM pepstatin on ice for 30 min. Proteins (60 µl/lane) were resolved
using a 10% gel and SDS-PAGE and transferred onto a polyvinylidene
difluoride membrane. Membranes were blocked with 5% non-fat dry
milk in Tris-buffered saline for 2 h at room temperature. Primary
antibodies, including anti-CDKN1A (cat. no. sc-6246; 1:1,000; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA), anti-cyclin E (cat. no.
20808; 1:1,000; CST Biological Reagents Co., Ltd., Shanghai, China)
and anti-β-actin (cat. no. ab8227; 1:1,000; Abcam, Cambridge, UK)
were used. Membranes were incubated with primary antibodies at room
temperature overnight. Following washing with Tris-buffered saline,
the membrane was incubated with goat anti-rabbit immunoglobulin G
horseradish peroxidase-conjugated secondary antibody (cat. no.
ab6721; 1:10,000; Abcam) for 2 h at room temperature. Finally,
blots were visualized using enhanced chemiluminescence detection
(Thermo Fisher Scientific, Inc.), as described previously (24).
Bioinformatics analysis
Gene Set Enrichment Analysis (GSEA) was performed
using GSEA software (version 2.2.4; Broad Institute, Inc.,
Massachusetts Institute of Technology, Cambridge, MA, USA), and
gene signatures were downloaded from the Molecular Signatures
Database (MSigDB, version 4; Broad Institute, Inc.). The MSigDB
collects various types of gene sets and included 1,320 canonical
pathways derived from the pathway databases including BioCarta,
Kyoto Encyclopedia of Genes and Genomes, Pathway Interaction
Database and Reactome. The GSE53622 dataset was downloaded from the
Gene Expression Omnibus (GEO) and only ESCC tumor samples were used
to perform GSEA.
Statistical analysis
Differences between groups were assessed by a paired
two-tailed Student's t-test. One-way analysis of variance followed
by the Bonferroni post-hoc test or the nonparametric Kruskal-Wallis
test was applied to assess the relationship between circPRKCI
expression and other characteristics. The strength of the
association between continuous variables was tested with the
Spearman correlation. All statistical analyses were performed using
SPSS 20 software (Abbott Laboratories, Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Association between SBF2-AS1
expression and clinical characteristics
SBF2-AS1 expression levels were detected in 51
paired primary ESCC and adjacent non-cancerous tissues using
RT-qPCR. Results demonstrated that SBF2-AS1 was significantly
overexpressed in ESCC, with a mean upregulation of 8.82-fold
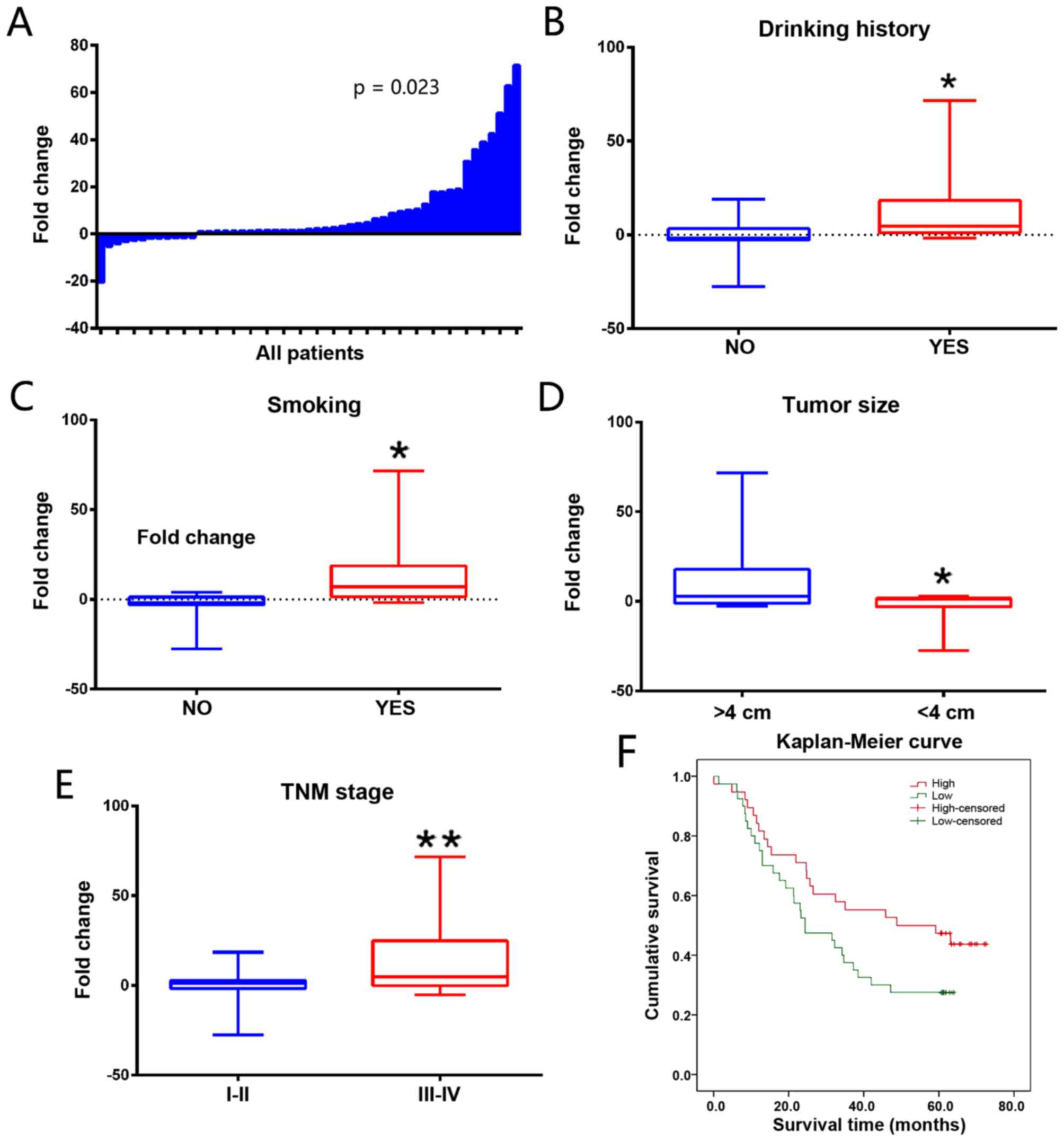
(P=0.023; Fig. 1A). As demonstrated
in Table I, drinking history, smoking
history, tumor size and TNM stage (P<0.05) were significantly
associated with the expression of SBF2-AS1. SBF2-AS1 expression
level was also increased in patients with a history of drinking
(9.927 vs. 4.507; P=0.030; Fig. 1B)
and smoking (10.938 vs. 4.232, P=0.032; Fig. 1C). Patients with a large tumor size
had an increased SBF2-AS1 expression (≥4 vs. <4 cm, 14.898 vs.
5.435; P=0.018; Fig. 1D). Patients
with ESCC exhibiting a TNM stage of III–IV demonstrated an
increased expression level of SBF2-AS1 compared with patients with
ESCC exhibiting with a TNM stage of I–II (TNM I–II vs. TNM III–IV,
1.302 vs. 15.475; P<0.01; Fig.
1E). The association between SBF2-AS1 expression level and
clinical characteristics is presented in Table I. The association between SBF2-AS1
expression and survival rate of ESCC patients was also evaluated.
In an online cohort of 60 ESCC patients, patients with increased
SBF2-AS1 expression levels had poorer survival rates compared with
those with lower expression levels of SBF2-AS1 [hazard ratio, 1.31;
95% confidence interval (CI), 1.090–1.568; P=0.016; Fig. 1F].
 | Table I.Association between SBF2-AS1
expression and clinicopathological characteristics. |
Table I.
Association between SBF2-AS1
expression and clinicopathological characteristics.
| Characteristic | Number of
patients | Percentage | Fold-change | P-value |
|---|
| Age, years |
|
|
| 0.298 |
|
<60 | 17 | 33.3 | 10.897 |
|
|
>60 | 34 | 66.7 | 7.78 |
|
| Sex |
|
|
| 0.270 |
|
Male | 35 | 80.3 | 9.861 |
|
|
Female | 16 | 19.7 | 7.232 |
|
| Smoker status |
|
|
| 0.032 |
|
Yes | 18 | 41.2 | 10.938 |
|
| No | 33 | 58.8 | 4.232 |
|
| Drinking
history |
|
|
| 0.030 |
|
Yes | 32 | 78.4 | 9.927 |
|
| No | 19 | 21.6 | 4.507 |
|
| Family history of
ESCC |
|
|
| 0.222 |
|
Yes | 4 | 7.8 | 3.804 |
|
| No | 47 | 92.2 | 9.179 |
|
| Location of the
tumor within the esophagus |
|
|
| 0.102 |
|
Upper | 9 | 17.6 | 11.612 |
|
|
Middle | 30 | 58.8 | 9.782 |
|
|
Lower | 12 | 23.5 | 4.058 |
|
| Tumor size, cm |
|
|
| 0.018 |
|
<4 | 37 | 72.5 | 14.898 |
|
|
>4 | 14 | 27.5 | 5.435 |
|
| Tumor
differentiation (according to the differences between tumor cells
and normal esophageal epithelial cells) |
|
|
| 0.101 |
| Poor
(differences, large) | 30 | 58.8 | 13.712 |
|
|
Moderate (differences,
moderate) | 7 | 13.7 | 9.747 |
|
| Well
(differences, minor) | 14 | 27.5 | 8.163 |
|
| Lymph node
metastasis |
|
|
| 0.727 |
|
N0-N1 | 18 | 35.3 | 7.504 |
|
|
N2-N3 | 33 | 64.7 | 9.422 |
|
| TNM stage |
|
|
| <0.010 |
|
I–II | 22 | 43.1 | 1.208 |
|
|
III–IV | 29 | 56.9 | 14.485 |
|
Expression characteristics of SBF2-AS1
in ESCC cell lines
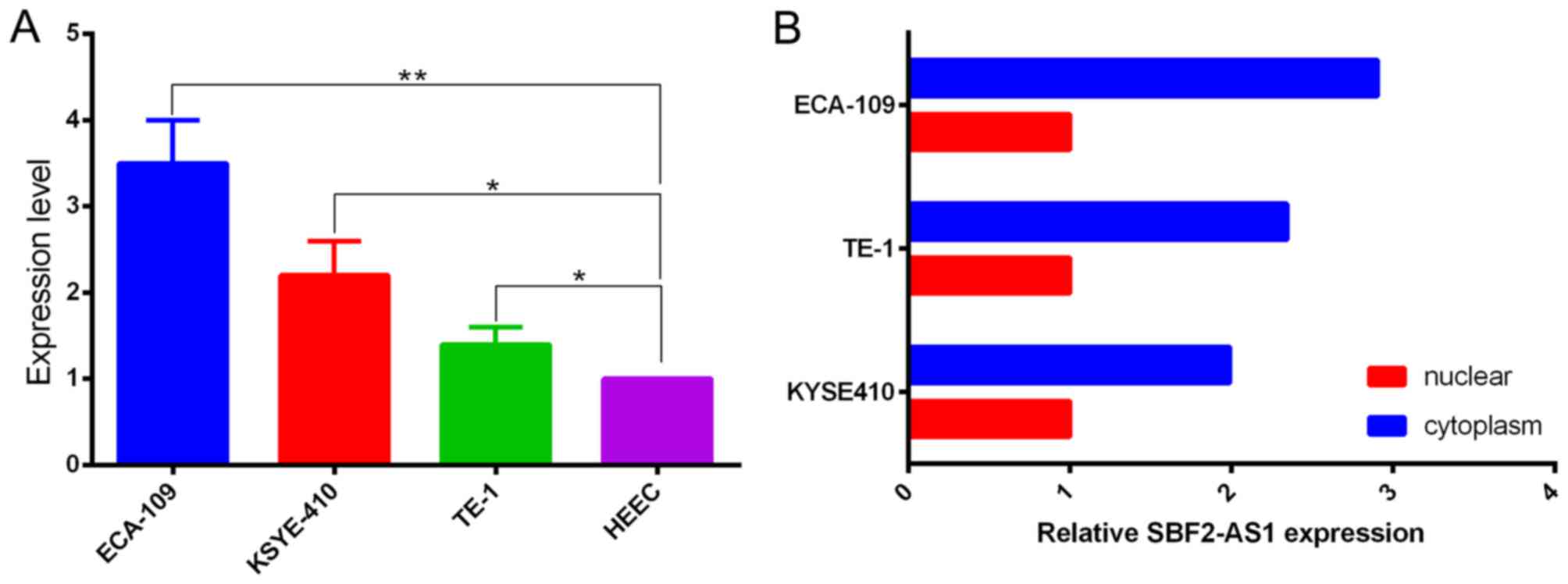
Prior to the functional study of SBF2-AS1 in
vitro, expression in ESCC cell lines was measured using
RT-qPCR. When normalized to HEEC cell lines, results demonstrated
that the expression level of SBF2-AS1 was upregulated in the ESCC
cells analyzed (Fig. 2A). The
expression level of SBF2-AS1 was highest in ECA-109 cells,
therefore ECA-109 was selected as the experimental cell line.
Confirmation of the subcellular localization of SBF2-AS1 was
achieved by measuring expression levels of SBF2-AS1 in nuclear and
cytoplasmic fractions of ECA-109, TE-1 and KYSE410 cell lines.
Results demonstrated that SBF2-AS1 was primarily distributed within
the cytoplasm (Fig. 2B).
SBF2-AS1 promotes the proliferation of
ESCC cell lines in vitro
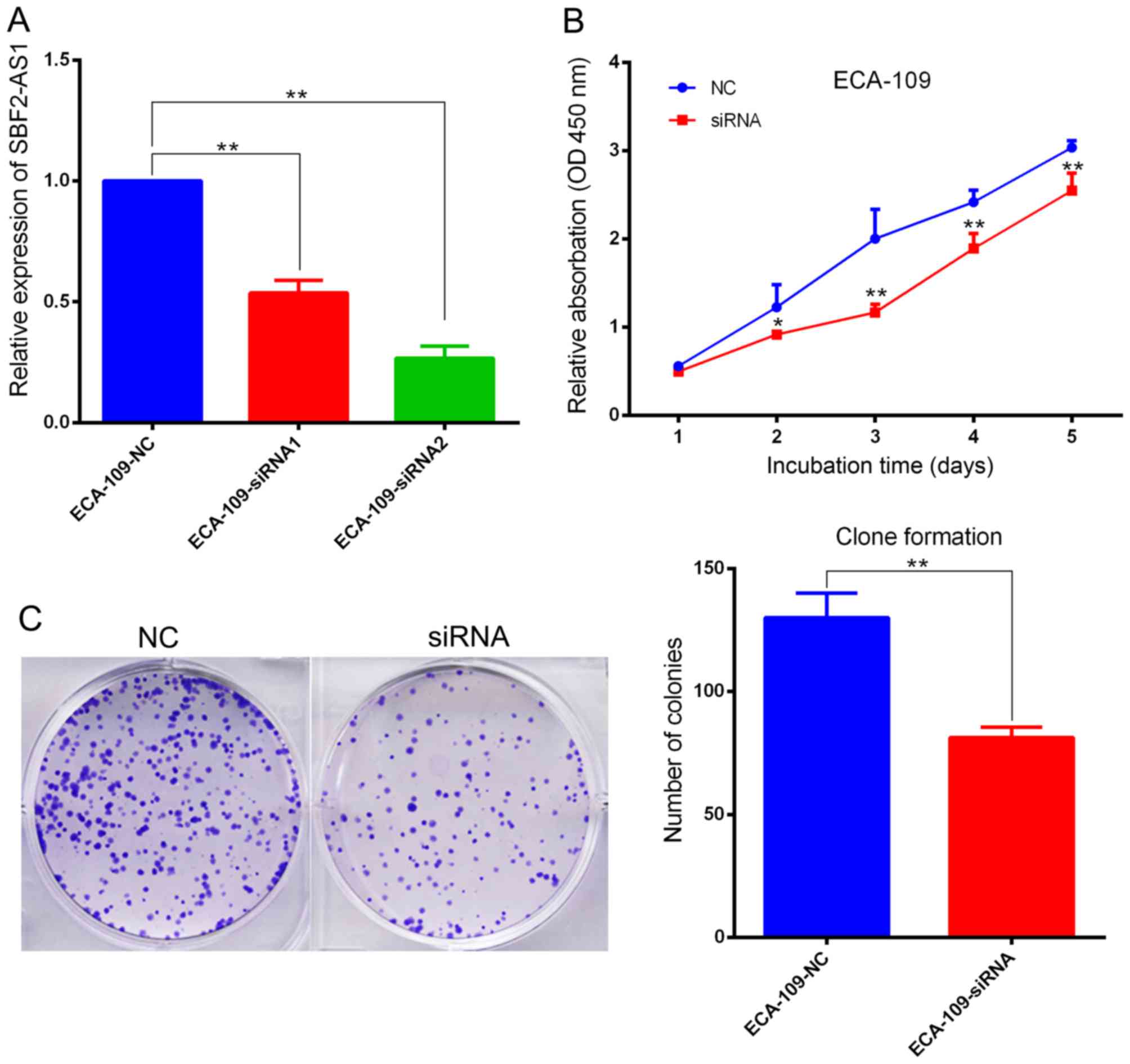
Investigation of the biological function(s) of
SBF2-AS1 was achieved by targeting SBF2-AS1 at the 204 (siRNA1) and
1021 (siRNA2) sites using two designer siRNAs as described
previously (14). Results indicate
that siRNA2 had improved inhibition efficacy, therefore it was used
in subsequent cell function experiments (Fig. 3A). ECA-109 cells were transfected with
si-SBF2-AS1 or NC siRNA. At 36 h post-treatment, SBF2-AS1
expression was effectively knocked down (Fig. 3A). The CCK-8 assay demonstrated that
knockdown of SBF2-AS1 significantly inhibited (P<0.01) cell
proliferation in the ECA-109 cell line (Fig. 3B). Furthermore, the colony formation
assay revealed that siRNA treatment significantly inhibited
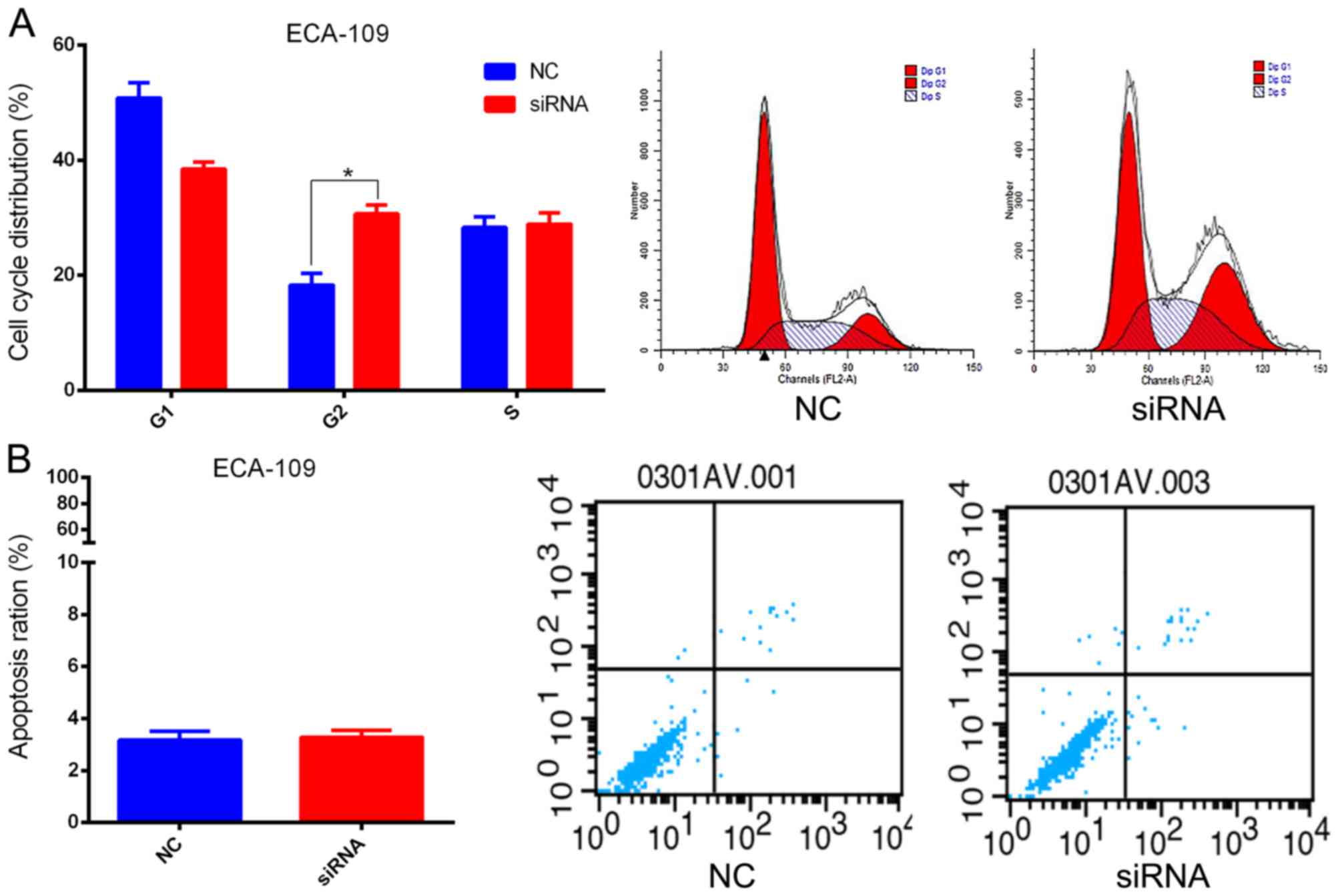
(P=0.007) the ability of ESCC cells to form colonies (Fig. 3C). Results generated using flow
cytometry demonstrated that cell cycle progression of ECA109 cells
was arrested at the G2 phase following treatment with
siRNA-2 (Fig. 4A). However, silencing
of SBF2-AS1 did not affect the proportion of apoptotic cells
(Fig. 4B).
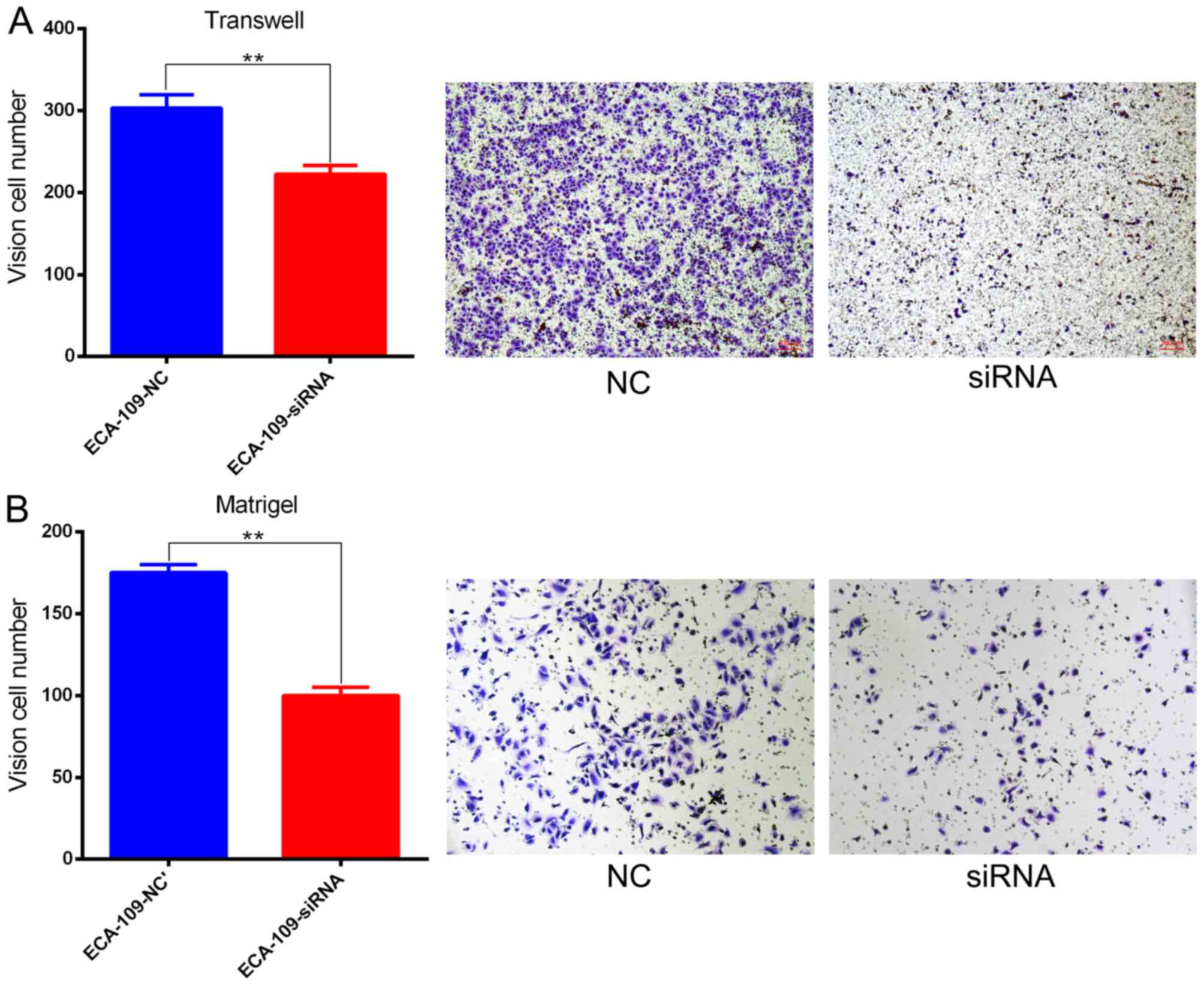
Effect of SBF2-AS1 on ESCC cell
migration and invasion
Migration and invasion are important cancer
progression events. The potential for SBF2-AS1 to have a direct
role in cell invasion within ESCC was investigated using Transwell
and Matrigel assays. Results demonstrated that siRNA treatment
significantly impaired (P=0.006) migration capacity compared with
NC (Fig. 5A). Knockdown of SBF2-AS1
also inhibited the invasive ability of ESCC cells (Fig. 5B).
Silencing SBF2-AS1 upregulates CDKN1A
expression in ESCC
To explore the potential mechanism of SBF2-AS1 in
ESCC GSEA analysis in an ESCC cohort was performed. According to
the GSEA results, SBF2-AS1 was significantly associated with genes
involved in Gene Ontology items of cell cycle (P=0.032), the
mitotic phase of cell cycle (P=0.018) and the cell cycle phase
(P=0.027). Notably, CDKN1A was identified in all three gene sets,
and was negatively associated with SBF2-AS1 (Fig. 6A-C). In addition, GSEA also revealed
that genes that were regulated by PRC2 were significantly
associated with SBF2-AS1 (Fig. 6D;
P<0.05). Thus, it was hypothesized that SBF2-AS1 may also
downregulate CDKN1A expression in ESCC cells. Expression levels of
CDKN1A were investigated in ECA-109 cells following the silencing
of SBF2-AS1. Results demonstrated that mRNA and protein levels of
CDKN1A were significantly increased (P<0.05) in ECA-109 cells
(Fig. 6E and F). Western blot
analysis further confirmed that cyclin E (CCNE1), a known
downstream gene of CDKN1A signaling, was also downregulated
following SBF2-AS1 silencing (Fig.
6F). Furthermore, in the third-party ESCC cohort (GSE53622),
SBF2-AS1 and CDKN1A were negatively correlated (R=−0.179;
P<0.01; Fig. 6G). Therefore, these
results indicate that SBF2-AS1 was able to decrease CDKN1A
expression in ESCC.
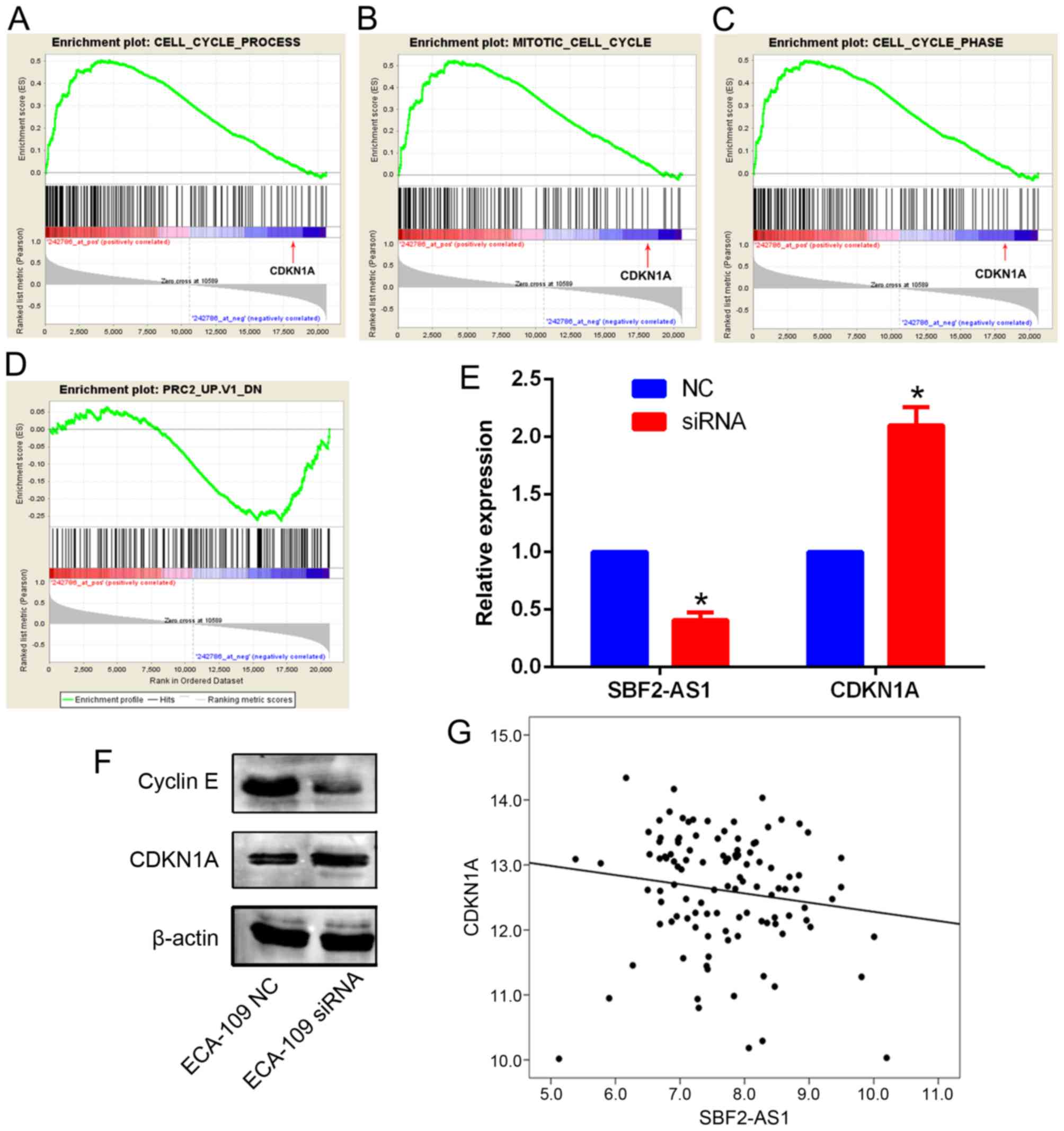 | Figure 6.GSEA results indicate that genes
associated with (A) cell cycle process, (B) mitotic cell cycle and
(C) cell cycle phase are positively enriched with SBF2-AS1, and
among these genes, CDKN1A was significantly negatively associated
with SBF2-AS1 (indicated by the red arrow). Genes regulated by PRC2
were negatively associated with SBF2-AS1. (D) Silencing of SBF2-AS1
increased the CDKN1A expression level in ECA-109 cells. (E) Cyclin
E expression was decreased following SBF2-AS1 silencing, which is
consistent with the (F) upregulation of CDKN1A. (G) SBF2-AS1 was
negatively associated with CDKN1A in the GSE53622 dataset
(P<0.01 vs. NC groups). *P<0.05. GSEA, Gene Set Enrichment
Analysis; SBF2-AS1, SET-binding factor 2 antisense RNA1; CDKN1A,
cyclin-dependent kinase 1A; PRC2, Polycomb repressive complex 2;
NC, negative control; siRNA, small interfering RNA. |
Discussion
Numerous lncRNAs have been identified as critical
regulators of gene expression, and serve several key roles in
tumorigenesis; therefore, there is a growing interest in the role
of lncRNAs in cancer (25). Multiple
types of lncRNA have been identified as associated with the
development and progression of ESCC, including pist, colon
cancer-associated transcript 2, homeobox transcript antisense RNA
and metastasis-associated lung adenocarcinoma transcript 1
(26–31). SBF2-AS1 is a novel lncRNA transcribed
on chromosome 11p15.1 and, previously, its function and molecular
mechanism in NSCLC has been reported (14). However, the function and molecular
mechanism of SBF2-AS1 in ESCC remains unclear. In the present
study, results demonstrated that the expression level of SBF2-AS1
was associated with smoking and drinking history, tumor size and
TNM stage. Previous studies confirmed that smoking and drinking are
significantly poor prognostic risk factors of ESCC (32,33). Chang
et al (34) revealed that
within a Chinese population sex, age, smoking and drinking status
are associated with an increased risk of ESCC in 9,805 cases and
10,493 controls. Tai et al (35) reported that women who consumed >158
g alcohol/week had a 20.58-fold increased risk (95% CI,
1.72–245.62; P=0.02) of developing ESCC compared with those who had
never consumed alcohol, following adjustment for other covariates.
Taghavi et al (36)
demonstrated that cigarette smoking was significantly associated
with p53 overexpression in ESCC cases (odds ratio, 3.64; 95% CI,
1.32–10.02; P=0.010). Therefore, it is suggested that SBF2-AS1 may
be associated with the occurrence and development of EC.
The biological function of SBF2-AS1 in vitro
was also analyzed using siRNA-mediated silencing in ECA109 cell
lines. Results demonstrated that following the knockdown of
SBF2-AS1, ESCC proliferative, migratory and invasive ability was
significantly suppressed. Flow cytometric analysis revealed that
the cell cycle was arrested in the G2 phase following
knockdown of SBF2-AS1; however, the proportion of apoptotic cells
was not affected. Evidence indicates that SBF2-AS1 may function as
an oncogenic gene in ESCC. In previous studies, it was reported
that SBF2-AS2 was able to bind to suppressor of zeste 12 homolog
(SUZ12), a core component of PRC2 (19,37–39). The
enrichment of SUZ12 subsequently induces the inhibition of
trimethylation of histone 3 Lys27 in the promoter region
of CDKN1A, and therefore downregulates the expression of CDKN1A
(19). Bioinformatical analysis in
the present study demonstrated that CDKN1A expression was
negatively associated with SBF2-AS1. Furthermore, GSEA revealed
that genes that were regulated by PRC2 were also significantly
associated with SBF2-AS1. Therefore, the results of the present
study confirm that CDKN1A was negatively regulated by SBF2-AS1 at
the RNA and protein levels. Nuclear and cytoplasmic fraction data
also demonstrated that SBF2-AS1 was distributed in the nucleus and
cytoplasm, which indicates that SBF2-AS1 may serve multiple
functions at the transcriptional and post-transcriptional levels.
According to our previous study (19), SBF2-AS1 may also transcriptionally
downregulate the expression of CDKN1A via SUZ12 binding within the
nucleus, as confirmed by the present study. CDKN1A, a
cyclin-dependent kinase inhibitor, serves an important role in cell
cycle arrest, and is therefore regarded as a tumor suppressor gene
(40). It is also reported that
CCNE1, a known downstream target gene of CDKN1A, was downregulated
following SBF2-AS1 silencing; however, the potential
post-transcriptional mechanism(s) of SBF2-AS1 require further
investigation.
To conclude, the results of the present study
demonstrate that SBF2-AS1 is significantly upregulated in ESCC, and
silencing SBF2-AS1 inhibits the proliferative and invasive ability
of ESCC cells. SBF2-AS1 may be a novel biomarker in the diagnosis
of ESCC, and a potential therapeutic target of ESCC; however,
further research in this area is required.
Acknowledgements
The present study was funded by the Natural Science
Foundation of China (grant nos. 81372321, 81572261, 81472200,
81501977 and 81702377), The Innovation Capability Development
Project of Jiangsu Province (grant no. BM2015004) and the Human
Resource Summit Grant of Jiangsu Province (grant nos. WSN-023 and
WSN-040).
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ESCC
|
esophageal squamous cell carcinoma
|
|
EA
|
esophageal adenocarcinoma
|
|
ncRNA
|
non-coding RNA
|
|
lncRNA
|
long non-coding RNA
|
|
SBF2-AS1
|
SET-binding factor 2 antisense RNA
1
|
|
CCK-8
|
Cell Counting Kit-8
|
|
PRC2
|
Polycomb repressive complex 2
|
|
CDKN1A
|
cyclin-dependent kinase inhibitor
1A
|
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clini. 66:115–132. 2016. View Article : Google Scholar
|
|
3
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kimura H, Konishi K, Arakawa H, Oonishi I,
Kaji M, Maeda K, Yabushita K, Tsuji M and Miwa A: Number of lymph
node metastases influences survival in patients with thoracic
esophageal carcinoma: Therapeutic value of radiation treatment for
recurrence. Dis Esophagus. 12:205–208. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ponting CP, Oliver PL and Reik W:
Evolution and Functions of Long Noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Derrien T, Johnson R, Bussotti G, Tanzer
A, Djebali S, Tilgner H, Guernec G, Martin D, Merkel A, Knowles DG,
et al: The GENCODE v7 catalogue of human long non-coding RNAs:
Analysis of their structure, evolution and expression. Genome Res.
22:1775–1789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cesana M, Cacchiarelli D, Legnini I,
Santini T, Sthandier O, Chinappi M, Tramontano A and Bozzoni I: A
long noncoding RNA controls muscle differentiation by functioning
as a competing endogenous RNA. Cell. 147:358–369. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Amaral PP, Dinger ME and Mattick JS:
Non-coding RNAs in homeostasis, disease and stress responses: An
evolutionary perspective. Brief Funct Genomics. 12:254–278. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li JY, Ma X and Zhang CB: Overexpression
of long non-coding RNA UCA1 predicts a poor prognosis in patients
with esophageal squamous cell carcinoma. Int J Clin Exp Pathol.
7:7938–7944. 2014.PubMed/NCBI
|
|
12
|
Tong YS, Zhou XL, Wang XW, Wu QQ, Yang TX,
Lv J, Yang JS, Zhu B and Cao XF: Association of decreased
expression of long non-coding RNA LOC285194 with chemoradiotherapy
resistance and poor prognosis in esophageal squamous cell
carcinoma. J Transl Med. 12:2332014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hao Y, Wu W, Shi F, Dalmolin RJ, Yan M,
Tian F, Chen X, Chen G and Cao W: Prediction of long noncoding RNA
functions with co-expression network in esophageal squamous cell
carcinoma. BMC Cancer. 15:1682015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cao W, Wu W, Shi F, Chen X, Wu L, Yang K,
Tian F, Zhu M, Chen G, Wang W, et al: Integrated analysis of long
noncoding RNA and coding RNA expression in esophageal squamous cell
carcinoma. Int J Genomics. 2013:4805342013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Shi X, Yang W, Lu Z, Wang P, Chen Z
and He J: Transcriptome profiling of lncRNA and co-expression
networks in esophageal squamous cell carcinoma by RNA sequencing.
Tumour Biol. 37:13091–13100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Chen Z, Tian L, Zhou C, He MY, Gao
Y, Wang S, Zhou F, Shi S, Feng X, et al: LncRNA profile study
reveals a three-lncRNA signature associated with the survival of
patients with oesophageal squamous cell carcinoma. Gut.
63:1700–1710. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tong YS, Wang XW, Zhou XL, Liu ZH, Yang
TX, Shi WH, Xie HW, Lv J, Wu QQ and Cao XF: Identification of the
long non-coding RNA POU3F3 in plasma as a novel biomarker for
diagnosis of esophageal squamous cell carcinoma. Mol Cancer.
14:32015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang E, Han L, Yin D, He X, Hong L, Si X,
Qiu M, Xu T, De W, Xu L, et al: H3K27 acetylation activated-long
non-coding RNA CCAT1 affects cell proliferation and migration by
regulating SPRY4 and HOXB13 expression in esophageal squamous cell
carcinoma. Nucleic Acids Res. 45:1–3101. 2017.PubMed/NCBI
|
|
19
|
Lv J, Qiu M, Xia W, Liu C, Xu Y, Wang J,
Leng X, Huang S, Zhu R, Zhao M, et al: High expression of long
non-coding RNA SBF2-AS1 promotes proliferation in non-small cell
lung cancer. J Exp Clin Cancer Res. 35:752016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
VanGuilder HD, Vrana KE and Freeman WM:
Twenty-five years of quantitative PCR for gene expression analysis.
Biotechniques. 44:619–626. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qiu M, Xu Y, Wang J, Zhang E, Sun M, Zheng
Y, Li M, Xia W, Feng D, Yin R and Xu L: A novel lncRNA, LUADT1,
promotes lung adenocarcinoma proliferation via the epigenetic
suppression of p27. Cell Death Dis. 6:e18582015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang X, Sun Q, Chen C, Yin R, Huang X,
Wang X, Shi R, Xu L and Ren B: ZYG11A serves as an oncogene in
non-small cell lung cancer and influences CCNE1 expression.
Oncotarget. 7:8029–8042. 2016.PubMed/NCBI
|
|
23
|
Wang J, Qiu M, Xu Y, Li M, Dong G, Mao Q,
Yin R and Xu L: Long noncoding RNA CCAT2 correlates with smoking in
esophageal squamous cell carcinoma. Tumour Biol. 36:5523–5528.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang X, Zhang Z, Qiu M, Hu J, Fan X, Wang
J, Xu L and Yin R: Glypican-5 is a novel metastasis suppressor gene
in non-small cell lung cancer. Cancer Lett. 341:265–273. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liz J and Esteller M: lncRNAs and
microRNAs with a role in cancer development. Biochim Biophys Acta.
1859:169–176. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wei G, Luo H, Sun Y, Li J, Tian L, Liu W,
Liu L, Luo J, He J and Chen R: Transcriptome profiling of
esophageal squamous cell carcinoma reveals a long noncoding RNA
acting as a tumor suppressor. Oncotarget. 6:17065–17080. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang X, Xu Y, He C, Guo X, Zhang J, He C,
Zhang L, Kong M, Chen B and Zhu C: Elevated expression of CCAT2 is
associated with poor prognosis in esophageal squamous cell
carcinoma. J Surg Oncol. 111:834–839. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen FJ, Sun M, Li SQ, Wu QQ, Ji L, Liu
ZL, Zhou GZ, Cao G, Jin L, Xie HW, et al: Upregulation of the long
non-coding RNA HOTAIR promotes esophageal squamous cell carcinoma
metastasis and poor prognosis. Mol Carcinog. 52:908–915. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ge XS, Ma HJ, Zheng XH, Ruan HL, Liao XY,
Xue WQ, Chen YB, Zhang Y and Jia WH: HOTAIR, a prognostic factor in
esophageal squamous cell carcinoma, inhibits WIF-1 expression and
activates Wnt pathway. Cancer Sci. 104:1675–1682. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li X, Wu Z, Mei Q, Guo M, Fu X and Han W:
Long non-coding RNA HOTAIR, a driver of malignancy, predicts
negative prognosis and exhibits oncogenic activity in oesophageal
squamous cell carcinoma. Br J Cancer. 109:2266–2278. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hirata H, Hinoda Y, Shahryari V, Deng G,
Nakajima K, Tabatabai ZL, Ishii N and Dahiya R: Long noncoding RNA
MALAT1 promotes aggressive renal cell carcinoma through Ezh2 and
interacts with miR-205. Cancer Res. 75:1322–1331. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Saeki H, Ohno S, Araki K, Egashira A,
Kawaguchi H, Ikeda Y, Morita M, Kitamura K and Sugimachi K: Alcohol
consumption and cigarette smoking in relation to high frequency of
p53 protein accumulation in oesophageal squamous cell carcinoma in
the Japanese. Br J Cancer. 82:1892–1894. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin S, Wang X, Huang C, Liu X, Zhao J, Yu
IT and Christiani DC: Consumption of salted meat and its
interactions with alcohol drinking and tobacco smoking on
esophageal squamous-cell carcinoma. Int J Cancer. 137:582–589.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang J, Huang Y, Wei L, Ma B, Miao X, Li
Y, Hu Z, Yu D, Jia W, Liu Y, et al: Risk prediction of esophageal
squamous-cell carcinoma with common genetic variants and lifestyle
factors in Chinese population. Carcinogenesis. 34:1782–1786. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tai SY, Wu IC, Wu DC, Su HJ, Huang JL,
Tsai HJ, Lu CY, Lee JM and Wu MT: Cigarette smoking and alcohol
drinking and esophageal cancer risk in Taiwanese women. World J
Gastroenterol. 16:1518–1521. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Taghavi N, Biramijamal F, Sotoudeh M,
Moaven O, Khademi H, Abbaszadegan MR and Malekzadeh R: Association
of p53/p21 expression with cigarette smoking and prognosis in
esophageal squamous cell carcinoma patients. World J Gastroenterol.
16:4958–4967. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kogo R, Shimamura T, Mimori K, Kawahara K,
Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, et al:
Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin
modification and is associated with poor prognosis in colorectal
cancers. Cancer Res. 71:6320–6326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Klattenhoff CA, Scheuermann JC, Surface
LE, Bradley RK, Fields PA, Steinhauser ML, Ding H, Butty VL, Torrey
L, Haas S, et al: Braveheart, a long non-coding RNA required for
cardiovascular lineage commitment. Cell. 52:570–583. 2013.
View Article : Google Scholar
|
|
40
|
Yang W, Du WW, Li X, Yee AJ and Yang BB:
Foxo3 activity promoted by non-coding effects of circular RNA and
Foxo3 pseudogene in the inhibition of tumor growth and
angiogenesis. Oncogene. 35:3919–3931. 2016. View Article : Google Scholar : PubMed/NCBI
|