Introduction
Cholangiocarcinoma is the second most common type of
primary liver malignancy globally (1), accounting for ~3% of gastrointestinal
tumors and 10–25% of all hepatobiliary malignancies. The incidence
rate of cholangiocarcinoma is increasing, particularly that of
intrahepatic cholangiocarcinoma (2).
However, despite advances in imaging, diagnosis and surgical
techniques, the 5-year overall survival rate for cholangiocarcinoma
is only 20–40% following potentially curative surgery, while the
median survival period is 22 months (3).
Natural products and their derivatives are valuable
chemical resources that may be used in the treatment and prevention
of cancer (4). Deoxypodophyllotoxin
(DPT), an analog of podophyllotoxin isolated from
Anthriscussylvestris roots, has been extensively studied due
to its multiple pharmacological activities. DPT exhibits antitumor
effects on HeLa cells, glioblastoma and non-small cell lung,
gastric and breast cancer (5–13), anti-inflammatory (14) and anti-viral (15) activities. There is an increasing
amount of evidence demonstrating that DPT inhibits proliferation in
cultured cancer cells by inducing apoptosis and or cell cycle
arrest (8–13).
To the best of our knowledge, the present study is
the first of its kind to examine the antitumor effect of DPT on
human cholangiocarcinoma cells, including its effects on cellular
growth and apoptosis rate, as well as the underlying
mechanisms.
Materials and methods
Materials
DPT was purchased from Yunnan Xili Pharmaceutical
Company (Kunming, China). The human cholangiocarcinoma QBC939 and
RBE cell cells were purchased from the Cell Bank of Type Culture
Collection of Chinese Academy of Sciences (Shanghai, China). All
cell culture reagents were obtained from Gibco; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). An Annexin V-fluorescein
isothiocyanate (FITC) apoptosis detection kit and propidium iodide
(PI)/RNase staining buffer were purchased from Calbiochem (EMD
Millipore, Billerica, MA, USA). MTT, dimethyl sulfoxide (DMSO),
Hoechst 33258, and antibodies were obtained from Sigma-Aldrich;
Merck KGaA (Darmstadt, Germany).
Cell lines
The human cholangiocarcinoma QBC939 and RBE cell
lines were cultured in RPMI-1640 medium, supplemented with 10%
(v/v) heat-inactivated fetal bovine serum, penicillin (100 U/ml)
and streptomycin (100 µg/ml). The cultures were maintained in a 5%
CO2 humidified atmosphere at 37°C.
Growth inhibitory evaluation
The MTT growth inhibition method was used to assess
the cytotoxicity of DPT as described previously (16). Briefly, QBC939 and RBE cells were
seeded in 96-well plates (4×103 cells/well). Following a
24 h incubation at 37°C to allow for attachment, the cells were
incubated with or without various concentrations of DPT (0, 0.05,
0.1, 0.5 and 1 µM) for indicated intervals (0, 24, 48 or 72 h).
Subsequently, 20 µl MTT dye solution (5 mg/ml in phosphate buffer;
pH 7.4) was added to each well and the cells were incubated for an
additional 4 h, prior to the addition of DMSO for color
development. Metabolic activity was quantified by measuring light
absorbance at 570 nm (17).
Flow cytometry for cell cycle
analysis
Following a 24 h incubation at 37°C to allow for
attachment, QBC939 and RBE cholangiocarcinoma cells
(1×106 cells/well) were seeded in 6-well plates and
treated with DPT for 48 h in a concentration range of 0–1 µM. The
cells were washed with PBS (pH 7.4) and fixed with 80% ice-cold
ethanol at 4°C overnight. The cells were subsequently treated with
80 mg/ml RNase and 50 mg/ml PI in the dark for 30 min, and analyzed
using a Coulter Epics XL Flow Cytometer (Beckman Coulter, Inc.,
Miami, FL, USA).
Hoechst 33258 staining
A fluorescent morphological assay (18) was performed to detect the apoptosis
induced by DPT. In total, 1×106 cells were seeded in
6-well plates and allowed to attach overnight. Thereafter the cells
were treated with 0.5 µM DPT or solvent control (Fresh medium
without serum) for 48 h. The cells were subsequently washed twice
with PBS and fixed using 4% formaldehyde for 15 min. Subsequently,
cells were washed in PBS and stained with 50 µl of Hoechst 33258
solution (50 ng/ml in PBS; Sigma-Aldrich; Merck KGaA) for 15 min at
4°C in the dark and subsequently examined using an Olympus FV1000
fluorescent microscope (Olympus Corporation, Tokyo, Japan) at 356
nm.
Analysis of apoptosis
Induction of apoptosis by DPT was assessed by the
binding of Annexin V to phosphatidylserine, which is externalized
to the outer leaflet of the plasma membrane early during the
induction of apoptosis. For Annexin V-FITC binding, QBC939 and RBE
cells were treated with DPT for 48 h, harvested and resuspended in
the binding buffer provided in the Annexin V-FITC apoptosis
detection kit. Cells were reacted with 5 µl Annexin V-FITC reagent
and 5 µl PI for 30 min at room temperature in the dark. Stained
cells were analyzed by flow cytometry.
Western blot analysis
Following treatment with 0, 0.05, 0.1, 0.5 or 1 µM
DPT for 48 h, the cells were washed twice with PBS, and lysed using
radioimmunoprecipitation assay buffer (20 mM Tris, pH 7.5; 150 mM
NaCl; 1% Triton X-100; 2.5 mM sodium pyrophosphate; 1 mM EDTA; 1%
Na3CO4; 0.5 g/ml leupeptin; 1 mM
phenylmethanesulfonyl fluoride) on ice to obtain the protein.
Lysates were subsequently centrifuged at 13,400 × g for 15 min at
4°C. The supernatant was collected and total protein concentrations
were measured using a bicinchoninic acid assay (Pierce; Thermo
Fisher Scientific, Inc.). Mitochondria and cytosol were separated
by differential ultracentrifugation (19). A total of 30 µg protein lysate was
separated by 10% SDS-PAGE and electrophoretically transferred to a
polyvinylidene difluoride membrane (Immunobilon-P, 0.45 mm; EMD
Millipore, Billerica, MA, USA) using the TE 77 Semi-Dry Transfer
Unit (GE Healthcare Life Sciences, Buckinghamshire, UK). The blot
was blocked in blocking buffer (5% non-fat dry milk; 1% Tween-20 in
PBS) for 1 h at room temperature, incubated with the following
specific primary antibodies: Cyclin B (cat no. sc-166210),
cyclin-dependent kinase 1 (CDK1; cat no. sc-53219), Bcl-2
associated X protein (Bax; cat no. sc-80658), B-cell lymphoma-2
(Bcl-2; cat no. sc-509), cleaved caspase-3 (cat no. sc-271759),
cleaved caspase-8 (cat no. sc-5263), cleaved caspase-9 (cat no.
sc-17784) and β-actin (all the primary antibodies were purchased
from Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) at a
dilution of 1:1,000 overnight at 4°C. Subsequently, blots were
incubated with the corresponding horseradish peroxidase conjugated
secondary antibody (cat no. sc-2350, Santa Cruz Biotechnology,
Inc.) at a dilution of 1:2,000 for 1 h at room temperature. The
signal was visualized with the Enhanced Chemiluminescence Plus
detection system (GE Healthcare Life Sciences, Shanghai, China).
Protein bands were semi-quantified by densitometric analysis using
ImageJ 1.43 software (National Institutes of Health, Bethesda, MA,
USA). The densitometry readings of the bands were normalized
according to β-actin expression.
Statistical analysis
All experiments were repeated ≥3 times. The data are
presented as the mean ± standard deviation and processed using SPSS
software (version 13.0; SPSS Inc., Chicago, IL, USA). Statistical
analyses were performed using either an unpaired or two-tailed
Student's t-test or one-way analysis of variance. Post-hoc analysis
between the groups was performed using Student-Newman-Keuls method.
P<0.05 was considered to indicate a statistically significant
difference.
Results
DPT inhibits the viability of QBC939
and RBE cells
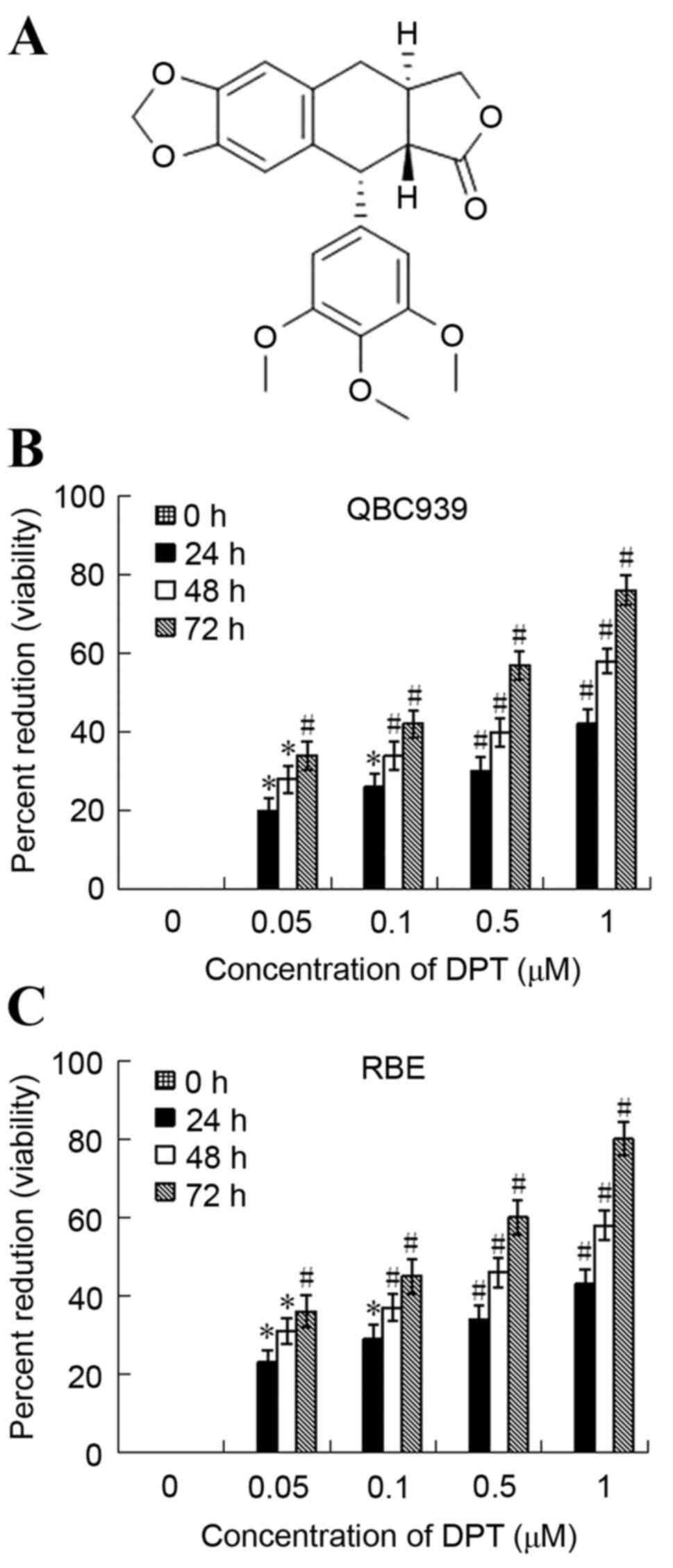
The growth inhibition effect of DPT on QBC939 and
RBE cellular viability was determined using an MTT assay. As
presented in Fig. 1, DPT induced a
dose and time-dependent inhibition of the cellular viability of
QBC939 and RBE cells following the treatment in vitro. The
50% growth inhibition concentration (IC50) of QBC939 was
estimated to be 1.186, 0.779 and 0.460 µM for 24, 48 and 72 h,
respectively. The IC50 of RBE was estimated to be 1.138,
0.726 and 0.405 µM for 24, 48 and 72 h, respectively.
DPT induced G2/M phase arrest in
QBC939 and RBE cells
The effect of DPT on cell cycle profile was analyzed
using flow cytometry. QBC939 and RBE cells were treated with 0,
0.05, 0.1, 0.5 and 1 µM of DPT for 48 h and their distribution in
the different phases of the cell cycle was calculated. The
population of cells in G2/M phase increased and that in S phase
decreased in a dose-dependent manner. For the QBC939 cells, the
percentage of cells in S phase decreased from 61.5±5.9 to
29.3±7.3%, while those in G2/M phase increased from 11.5±2.9 to
49.1±5.6% (Table I). For the RBE
cells, the percentage of cells in S phase decreased from 61.4±6.1
to 35.5±4.7%, while those in G2/M phase increased from 12.0±2.4 to
46.7±4.4% (Table II).
 | Table I.Effect of DPT on cell cycle profile of
QBC939 cells analyzed using FACScan analysis. |
Table I.
Effect of DPT on cell cycle profile of
QBC939 cells analyzed using FACScan analysis.
| DPT (µM) | G0/G1 | G2/M | S |
|---|
| 0 | 27.0±5.6 | 11.5±2.9 | 61.5±5.9 |
| 0.05 | 25.1±4.8a | 19.5±3.5a | 55.4±4.8a |
| 0.1 | 24.3±4.4a | 26.5±5.9b | 49.2±6.1a |
| 0.5 | 20.8±4.2b | 39.7±5.0b | 39.5±7.5b |
| 1 | 21.6±3.8b | 49.1±5.6b | 29.3±7.3b |
| Table II.Effect of DPT on cell cycle profile
of RBE cells by FACScan analysis. |
Table II.
Effect of DPT on cell cycle profile
of RBE cells by FACScan analysis.
| DPT (µM) | G0/G1 | G2/M | S |
|---|
| 0 | 26.6±5.2 | 12.0±2.4 | 61.4±6.1 |
| 0.05 |
23.5±4.9a | 19.2±3.0a |
57.3±4.3a |
| 0.1 | 23.9±4.2a |
26.3±5.6b | 49.8±5.2a |
| 0.5 |
19.1±4.3b | 40.3±4.5b |
40.6±7.3b |
| 1 | 17.8±3.5b |
46.7±4.4b | 35.5±4.7b |
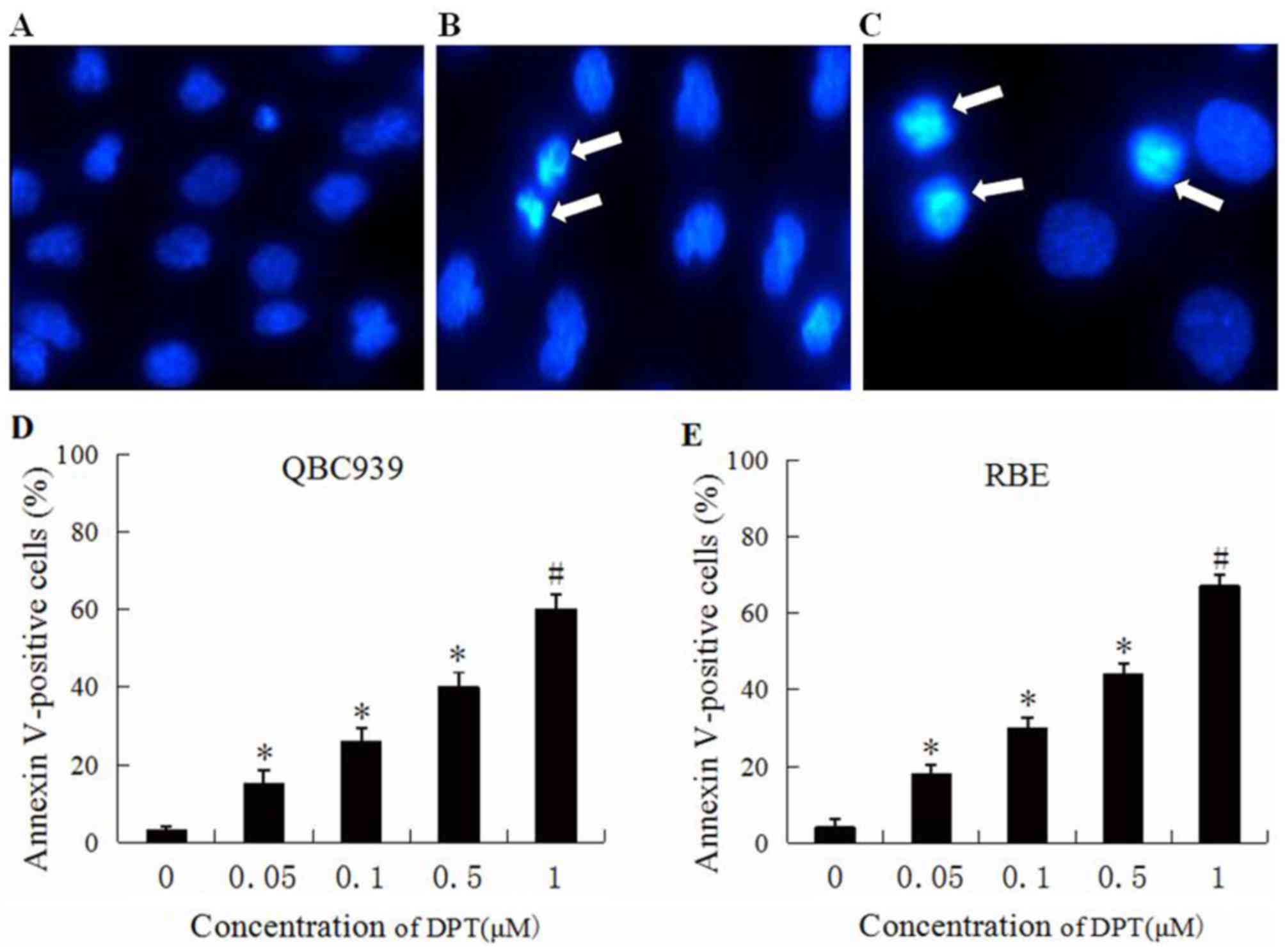
Morphological changes caused by
DPT
Control and DPT treated cells were stained with the
fluorescent dye Hoechst 33528 and visualized using a fluorescent
microscope. The control cells were normal, and the nuclei were
round and homogeneous (Fig. 2A),
whereas QBC939 (Fig. 2B) and RBE
(Fig. 2C) cells treated with DPT
exhibited cell shrinkage, chromatin condensation and nuclear
deformation and disassembly, which are all features indicative of
apoptosis.
Effects of DPT on QBC939 and RBE cell
apoptotic induction
The effects of DPT on cellular apoptosis were
analyzed using flow cytometry. Subsequent to QBC939 and RBE cells
being treated with various concentrations of DPT for 48 h, cells
were stained with Annexin V-FITC and PI. As presented in Fig. 2D, the percentage of Annexin V-FITC
binding QBC939 cells treated with DPT increased in a
concentration-dependent manner from 17% at 0.05 µM DPT to 60% at 1
µM DPT. As shown in Fig. 2E, the
percentage of Annexin V-FITC binding RBE cells treated with DPT
increased in concentration-dependent manner from 19% at 0.05 µM DPT
to 68% at 1 µM DPT.
Effects of DPT on cell cycle
regulators
In order to reveal the mechanisms underlying the
G2/M arrest observed subsequent to the addition of DPT, western
blotting was used to determine the expression levels of cell
cycle-regulating proteins including Cyclin B1 and CDK1. The present
results revealed that DPT treatment resulted in a significant
reduction in the protein levels of Cyclin B1 and CDK1 in a dose
dependent manner (Fig. 3).
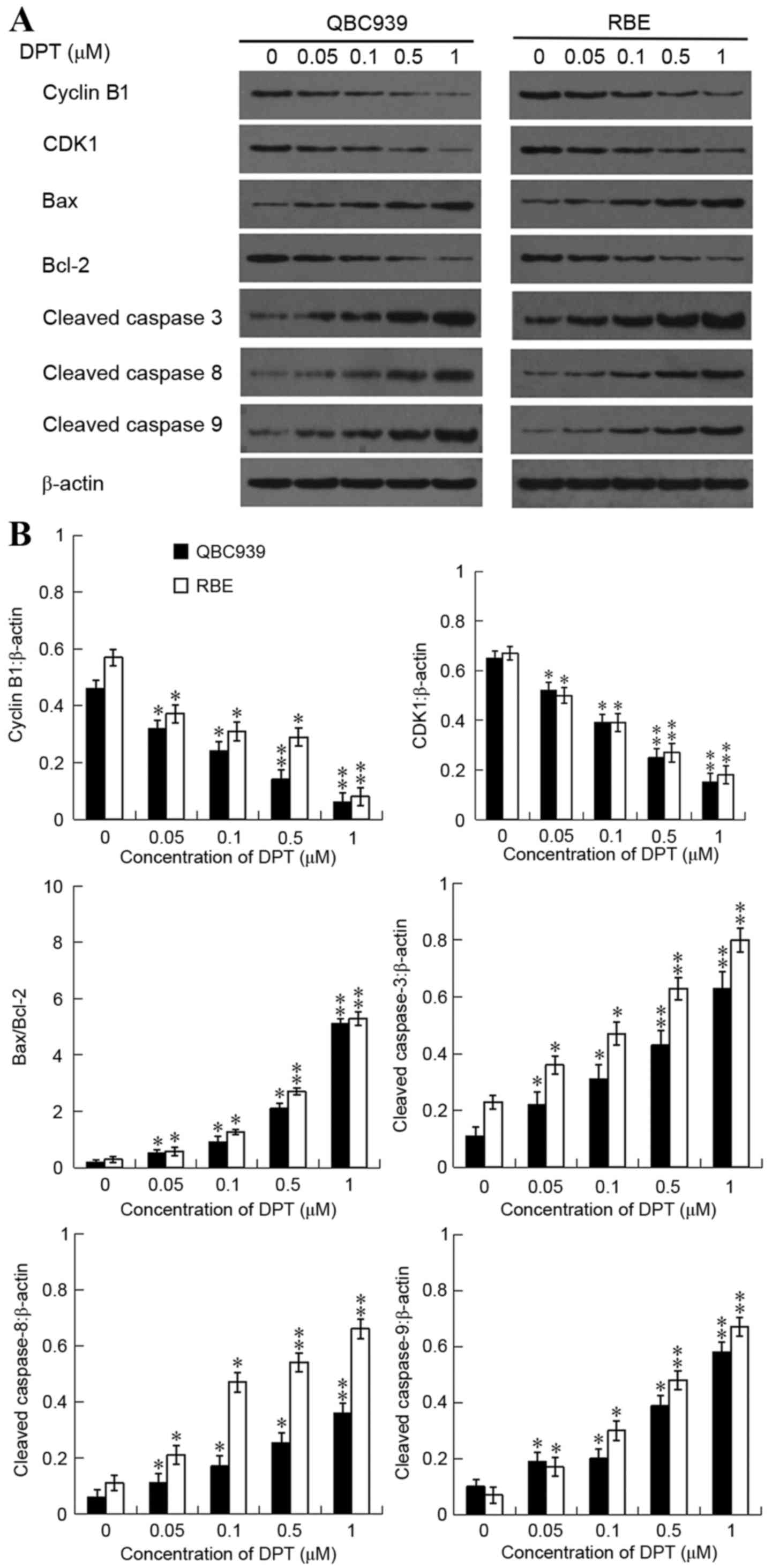 | Figure 3.Effect of DPT on protein expression
levels of Cyclin B1, CDK1, Bax, Bcl-2, cleaved caspase-3, cleaved
caspase-8 and cleaved caspase-9. QBC939 and RBE cells were
incubated either with or without DPT (0–1 µM) for 48 h and the
expression levels of Cyclin B1, CDK1, Bax, Bcl-2, cleaved
caspase-3, cleaved caspase-8 and cleaved caspase-9 were assessed by
western blot analysis. (A) Western blot analysis of Cyclin B1,
CDK1, Bax Bcl-2, cleaved caspase-3, cleaved caspase-8 and cleaved
caspase-9 expression levels, while β-actin was probed as the
protein loading control. (B) Band densitometry analysis of Cyclin
B1, CDK1, cleaved caspase-3, −8 and −9 expression levels normalized
to β-actin and the rations of Bax/Bcl-2 in QBC939 and RBE cells.
*P<0.05 and **P<0.01 vs. control. DPT, deoxypodophyllotoxin.
CDK1, cyclin dependent kinase-1; Bax, Bcl-2 associated X protein;
Bcl-2, b-cell lymphoma-2. |
Effects of DPT on apoptosis
regulators
To examine the mechanisms underlying the apoptosis
observed following the addition of DPT, western blot analysis was
used to determine the expression levels of apoptosis-associated
proteins including Bax, Bcl-2, cleaved caspase-3, −8 and −9. DPT
treatment resulted to a significant reduction in the protein levels
of cleaved caspase-3, −8, and −9 in a dose-dependent manner
(Fig. 3). In addition, the expression
level of Bax in DPT-treated cells was greatly increased, and
accompanied by the downregulation of Bcl-2 expression levels
(Fig. 3). Therefore, the ratios of
Bax/Bcl-2 were notably increased in a dose-dependent manner.
Discussion
DPT has been widely used to treat numerous diseases
due to its anti-inflammatory and antitumoral effects (5–13).
However, the effect of DPT on human cholangiocarcinoma cells
remains unclear. The present study demonstrated that DPT exhibited
a significantly inhibitory effect on the proliferation of
cholangiocarcinoma cells (QBC939 and RBE) in a time and
concentration-dependent manner. The IC50 of DPT was
~1.186, 0.779 and 0.460 µM in QBC939 cells and 1.138, 0.726 and
0.405 µM in RBE cells for 24, 48 and 72 h, respectively. Additional
analysis revealed that DPT exerted clear cytotoxic activity against
human cholangiocarcinoma cells, primarily via cell cycle arrest at
the G2/M phase associated with the decrease of the Cyclin B1 and
CDK1 and apoptosis induction, which was verified by the typical
apoptotic morphological changes and the significant increase in
apoptotic cell populations. The present study also identified that
the apoptosis of cholangiocarcinoma cells induced by DPT involves
intrinsic and extrinsic pathways.
It is known that cellular growth and proliferation
of mammalian cells are mediated by cell cycle progression.
Furthermore, inhibition of the cell cycle has become an effective
strategy for eliminating cancer cells (20). The results of the present study
demonstrated that the percentage proportion was reduced in the S
phase cells and increased in the G2/M phase cells following
treatment with DPT in a dose-dependent manner, indicating that the
inhibitory effect of DPT on QBC939 and RBE cellular proliferation
is mediated by G2/M phase cell cycle arrest. Previously, DPT was
reported to induce inhibition of growth via cell cycle arrest in
the G2/M phase of different cancer cell lines including HeLa
leukemia (8–9) and non-small cell lung (10), gastric (11) and breast cancer (12) and glioblastoma (13) cells which is consistent with the
results obtained in the present study.
It is known that cyclin protein and cyclin-dependent
kinase (CDK) are two major components in cell cycle regulation
(9), which can combine into the
activated cyclin-CDK kinase complex to promote cell cycle
transport. Among CDKs that regulate cell cycle progression, Cyclin
B, in association with CDK1, governs cell cycle progression by
enhancing cell cycle distribution in the G2/M fraction (21). Entry into mitosis requires that
CDK1/Cyclin B complexes are activated by Cdc25C phosphatase, which
removes the inhibitory phosphorylation of CDK1 (22). To examine the mechanisms by which DPT
led to G2/M phase arrest in QBC939 and RBE cells, the present study
additionally investigated the status of key factors known to
regulate cell cycle progression. The present result srevealed that
DPT treatment significantly decreased the expression of Cyclin B1
and CDK1 suggesting that G2/M cell cycle arrest in
cholangiocarcinoma cells is caused, at least in part, by changes in
the Cyclin B1 and CDK1 protein levels.
Apoptosis is the process of programmed cell death,
and is coupled with a number of clear morphological changes and
cellular signaling pathways (23).
The activation of apoptosis has been regarded as a target in cancer
therapies (24). The morphological
characteristics of apoptosis include condensation of cytoplasm and
chromatin, chromosomal DNA fragmentation, and formation of
apoptotic bodies (25). Therefore,
the present study used Hoechst 33258 staining and flow cytometric
assays to measure DPT induced apoptosis. In DPT-treated QBC939 and
RBE cells, typical apoptotic changes of nuclear pyknosis and
karyorrhexis are observed by Hoechst 33258 staining. Meanwhile,
Annexin V staining results demonstrated that DPT significantly
induced the apoptosis of QBC939 and RBE cells in a dose dependent
manner (P<0.05). The results therefore suggest that DPT
inhibited the proliferation of the QBC939 and RBE cells partially
by inducing apoptosis.
In mammalian cells, death receptor induced extrinsic
and mitochondria apoptosome mediated apoptotic intrinsic pathways
are the two major pathways involved in the initiation of apoptosis
(26). Bcl-2 family members and
caspases perform a central role in controlling the two pathways.
Mitochondria also perform a critical role in apoptosis induced by
chemotherapeutic agents (27).
Apoptosis is often associated with impaired mitochondrial adenine
nucleotide exchange, the alternative channels in the mitochondrial
outer membrane that permit the transit of adenine nucleotides
(28). This process can be regulated
by the Bcl-2 family of proteins that govern the release of
cytochrome c from the mitochondria (29). The increased ratio of Bax/Bcl-2
induces the loss of mitochondrial membrane potential leading to
permeability transition and cytochrome c release.
Furthermore, cytochrome c activates caspase-9, which cleaves
and activates the downstream effect of proteases, including
caspase-3, thus triggering apoptosis (30). The cell death receptor pathway,
mediated distinctively through active/cleaved caspase-8, performs
an important role in the maintenance of tissue homeostasis. In
addition, this pathway is characterized by binding cell death
ligands and cell death receptors, and subsequently activates
caspase-8, −3, −6 and −7 (31) and
cleaved poly ADP-ribose polymerase. A number of studies have shown
that DPT induces caspase-mediated apoptosis in a range of cancer
cells (8–13) and affects numerous factors in cellular
signal transduction pathways which are linked to the proapoptotic
(Bax) and antiapoptotic (Bcl-2) gene (9,10).
Accordingly, in the present study, the elevated ratio of Bax/Bcl-2
and cleavage of caspase-3, −8 and −9 significantly increased the
dose-dependent exposure to DPT. Generally, these findings indicated
that intrinsic and extrinsic pathways are involved in DPT induced
apoptosis.
In summary, the present results demonstrated, for
the first time, that DPT is a potent growth inhibitor of
cholangiocarcinoma cells. The growth inhibition is related to the
G2/M phase cell cycle arrest associated with the downregulation of
Cyclin B1 and CDK1 and the induction of apoptosis via the intrinsic
and extrinsic pathways. Thus, these results provide the basis for
DPT as a promising agent against cholangiocarcinoma. However, the
in vivo experiment and exact molecular mechanisms behind
this needs additional investigations.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Health
Department Scientific Research Foundation of Hunan Province (grant
no. 132012008).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Author's contributions
MX, SP and XF conceived and designed the study. MX,
YF, YZ and SL performed the experiments, analyzed the data and
wrote the paper. MX, SP and XF reviewed and edited the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Burger I, Hong K, Schulick R, Georgiades
C, Thuluvath P, Choti M, Kamel I and Geschwind JF: Transcatheter
arterial chemoembolization in unresectable cholangiocarcinoma:
Initial experience in a single institution. J Vasc Interv Radiol.
16:353–361. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gatto M, Bragazzi MC, Semeraro R, Napoli
C, Gentile R, Torrice A, Gaudio E and Alvaro D: Cholangiocarcinoma:
Update and future perspectives. Dig Liver Dis. 42:253–260. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hammill CW and Wong LL: Intrahepatic
cholangiocarcinoma: A malignancy of increasing importance. J Am
Coll Surg. 207:594–603. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yap TA and Workman P: Exploiting the
cancer genome: Strategies for the discovery and clinical
development of targeted molecular therapeutics. Annu Rev Pharmacol
Toxicol. 52:549–573. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ikeda R, Nagao T, Okabe H, Nakano Y,
Matsunaga H, Katano M and Mori M: Antiproliferative constituents in
umbelliferae plants. III. Constituents in the root and the ground
part of Anthriscus sylvestris Hoffm. Chem Pharm Bull.
46:871–874. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Masuda T, Oyama Y, Yonemori S, Takeda Y,
Yamazaki Y, Mizuguchi S, Nakata M, Tanaka T, Chikahisa L, Inabak Y
and Okada Y: Flow cytometric estimation on cytotoxic activity of
leaf extracts from seashore plants in subtropical Japan: Isolation,
quantification and cytotoxic action of (−)-deoxypodophyllotoxin.
Phytother Res. 16:353–358. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Muto N, Tomokuni T, Haramoto M, Tatemoto
H, Nakanishi T, Inatomi Y, Murata H and Inada A: Isolation of
apoptosis- and differentiation-inducing substances toward human
promyelocytic leukemia HL-60 cells from leaves of Juniperus
taxifolia. Biosci Biotechnol Biochem. 72:477–484. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yong Y, Shin SY, Lee YH and Lim Y:
Antitumor activity of deoxypodophyllotoxin isolated from
Anthriscus sylvestris: Induction of G2/M cell cycle arrest
and caspase-dependent apoptosis. Bioorg Med Chem Lett. 9:4367–4371.
2009. View Article : Google Scholar
|
|
9
|
Shin SY, Yong Y, Kim CG, Lee YH and Lim Y:
Deoxypodophyllotoxin induces G2/M cell cycle arrest and apoptosis
in HeLa cells. Cancer Lett. 287:231–239. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu M, Jiang Z, Duan H, Sun L, Zhang S,
Chen M, Wang Y, Gao Q, Song Y, Zhu X and Zhang L:
Deoxypodophyllotoxin triggers necroptosis in human non-small cell
lung cancer NCI-H460 cells. Biomed Pharmacother. 67:701–706. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang YR, Xu Y, Jiang ZZ, Guerram M, Wang
B, Zhu X and Zhang LY: Deoxypodophyllotoxin induces G2/M cell cycle
arrest and apoptosis in SGC-7901 cells and inhibits tumor growth in
vivo. Molecules. 20:1661–1675. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Benzina S, Harquail J, Jean S, Beauregard
AP, Colguhoun CD, Carroll M, Bos A, Gray CA and Robichaud GA:
Deoxypodophyllotoxin isolated from Juniperus communis
induces apoptosis in breast cancer cells. Anticancer Agents Med
Chem. 15:79–88. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mounia G, Jiang ZZ, Sun LX, Zhu X and
Zhang LX: Antineoplastic effects of deoxypodophyllotoxin, a potent
cytotoxic agent of plant origin, on glioblastoma U-87 MG and SF126
cells. Pharmacol Rep. 67:245–252. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee SH, Son MJ, Ju HK, Lin CX, Moom TC,
Choi HG, Son JK and Chang HW: Dual inhibition of cyclooxygenases-2
and 5-lipoxygenase by deoxypodophyllotoxin in mouse bone
narrow-derivd mast cells. Biol Pharm Bill. 27:786–788. 2004.
View Article : Google Scholar
|
|
15
|
Sudo K, Konno K, Shigeta S and Yokota T:
Inhibitory effects of podophyllotoxin derivatives on herpes simplex
virus replication. Antivir Chem Chemother. 9:263–267. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Niemann-Jönsson A, Ares MP, Yan ZQ, Bu DX,
Fredrikson GN, Brånén L, Pörn-Ares I, Nilsson AH and Nilsson J:
Increased rate of apoptosis in intimal arterial smooth muscle cells
through endogenous activation of TNF receptors. Arterioscler Thromb
Vasc Biol. 21:1909–1914. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Meier P, Finch A and Evan G: Apoptosis in
development. Nature. 407:796–801. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lizotte E, Tremblay A, Allen BG and Fiset
C: Isolation and characterization of subcellular protein fractions
from mouse heart. Anal Biochem. 345:47–54. 2015. View Article : Google Scholar
|
|
20
|
Cho JH, Lee JG, Yang YI, Kim JH, Ahn JH,
Baek NI, Lee KT and Choi JH: Eupatilin, a dietary flavonoid,
induces G2/M cell cycle arrest in human endometrial cancer cells.
Food Chem Toxicol. 49:1737–1744. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Taylor WR and Stark GR: Regulation of the
G2/M transition by p53. Oncogene. 20:1803–1815. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Smits VJ and Medema RH: Checking out the
G(2)/M transition. Biochim Biophys Acta. 1519:1–12. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang R, Liu A, Ma X, Li L, Su D and Liu J:
Sodium tanshinone IIA sulfonate protects cardiomyocytes against
oxidative stress-mediated apoptosis through inhibiting JNK
activation. J Cardiovasc Pharmacol. 51:396–401. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Neto CC, Amoroso JW and Liberty AM:
Anticancer activities of cranberry phytochemicals: An update. Mol
Nutr Food Res. 52 Suppl 1:S18–S27. 2008.PubMed/NCBI
|
|
26
|
Wei H and John JK: Anticancer therapy
targeting the apoptotic pathway. Oncology. 4:721–729.
2003.PubMed/NCBI
|
|
27
|
Delivani P and Martin SJ: Mitochondrial
membrane remodeling apoptosis: An inside story. Cell Death Differ.
13:2007–2010. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vander Heiden MG, Chandel NS, Williamson
EK, Schumacker PT and Thompson CB: Bcl-xL regulates the membrane
potential and volume homeostasis of mitochondria. Cell. 91:627–637.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Green DR: Apoptotic pathways: Ten min to
dead. Cell. 121:671–674. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu X, Yue P, Zhou Z, Khuri FR and Sun SY:
Death receptor regulation and celecoxib-induced apoptosis in human
lung cancer cells. J Natl Cancer Inst. 96:1769–1780. 2004.
View Article : Google Scholar : PubMed/NCBI
|