Introduction
Cervical cancer is estimated to be the fourth most
frequent type of cancer among women worldwide (1) and the fourth leading cause of
cancer-related mortality (1–3). Environmental factors, living habits and
human papillomavirus (HPV) infection have been linked to the
development of cervical cancer (4–6).
Specifically, HPV infection is considered to be one of the most
important causal factors related to cervical cancer (7). However, HPV alone, appears to not be
sufficient for the development of cervical cancer (8), as only a small amount of HPV-infected
women finally develop cervical cancer (3,9).
A number studies have provided evidence of familial
clustering of cervical cancer, supporting the existence of genetic
effects (10–12). Moreover, several association studies
have demonstrated a number of genetic variants that possibly confer
susceptibility to cervical cancer by affecting immune responses,
DNA repair or viral cell entry and infection (3,13,14). However, uncertainty for the effect
size of genetic variants, particularly in different ethnic
backgrounds still exists, as the results of different genetic
studies have been conflicting (13).
Recently, Wang et al, genotyped 7,140 SNPs
across 305 genes that were involved in HPV infection, cell entry
and DNA repair, and reported that genes, among which general
transcription factor IIH subunit 4 (GTF2H4), deoxyuridine
triphosphatase (DUT), DNA meiotic recombinase 1
(DMC1), 2′-5-oligoadenylate synthetase 3 (OAS3),
sulfatase 1 (SULF1), interferon gamma (IFNG),
transmembrane channel like 6 (TMC6) and transmembrane
channel like 8 (TMC8) were associated with the risk of HPV
persistence and cervical pre-cancer/cancer (14). The loss of the expression of
DMC1 plays an important role in the development of cancers
in human tissues, including cervical cancer lines (15). The DUT enzyme influences nucleotide
metabolism by producing the immediate precursor of thymidine
nucleotides, dUMP, and consequently decreasing the intracellular
concentration of dUTP (16). As a
result, uracil cannot be incorporated into DNA (16). SULF1 is a heparin-degrading
endosulfatase, which desulfates heparan sulfate proteoglycans
(HSPGs) and blocks the binding of growth factors and their
receptors, inhibiting as a result, the activation of growth factors
and signaling pathways (17,18). OAS3 is induced during viral infection
and plays an important role on the antiviral intracellular innate
immune response (19). GTF2H4 is a
general transcription factor that interacts with factors important
in carcinogenesis and is involved in processes of DNA repair and
transcriptional control (20). IFNG
is regulatory cytokine, released by lymphocytes, that enhances
cellular immune responses via increased T-cell cytotoxicity and
natural killer (NK)-cell activity (21). The TMC6 and TMC8 genes
(also referred to as EVER1 and EVER2 genes), are
known for the development of Epidermodysplasia verruciformis, which
is associated with a high sensitivity to HPV infections (22). The TMC6 and TMC8 proteins appear to
regulate cellular zinc homeostasis in keratinocytes and lymphocyte
(23).
In a previous analysis in a population from Northern
Greece (3), we failed to detect a
significant effect of two SNPs of the EVER1/2 gene region
(rs2290907 and rs16970849) and the FAS-670 polymorphism
(rs1800682) on precancerous lesions and cervical cancer. This was
in contrast to a previous positive study by Castro et al
(24). FAS belongs to the family of
tumor necrosis factor (TNF) receptors (25,26). The
downregulation of FAS leads to resistance to death signals, a
phenomenon that has been observed in cervical cancer (27–30).
The present study was designed to replicate the
findings reported by Wang et al (14) and Castro et al (24) in a different, from our previous study
(3), Greek population of Central
Greece. In particular, we examined the effects of 10 SNPs
(rs1800682, rs5757133, rs3784621, rs4737999, rs12302655, rs2894054,
rs11177074, rs2290907, rs9893818 and [FAS, DMC1, DUT, SULF1,
OAS3, GTF2H4, IFNG, TMC6 and TMC8 (2 SNPs)] on the risk
of precancerous lesions and cervical cancer.
Materials and methods
Study population
A total of 608 women that had attended the
Obstetrics and Gynaecology Clinic of the University Hospital of
Larissa, Larissa, Greece participated in this study. The patient
group consisted of 316 women with a histopathologically confirmed
diagnosis of cervical cancer (n=17) or precancerous lesions, either
high grade (CIN2/3, n=100) or persistent low grade (CIN1, n=199).
The control group consisted of 292 age-matched women with normal
annual cervical cytology screening.
The local Ethics Review Board of the University
Hospital of Larissa approved the study protocol. Informed consent
was obtained from all individual participants included in the
study.
Isolation of DNA and genotyping
Genomic DNA was extracted from 200 µl of
EDTA-anti-coagulated whole blood, using a QIAamp® DNA
Blood Mini kit (Qiagen GmbH, Hilden, Germany) according to the
manufacturer's instructions. SNPs were genotyped with TaqMan
allele-specific PCR amplification technology on an ABI PRISM 7900
Sequence Detection System and analyzed with the Sequence Detection
Software (SDS 2.1) (both from Applied Biosystems, Foster City, CA,
USA) by laboratory personnel blinded to clinical status. In order
to assess genotyping reproducibility, initially observed SNP
allelic discrimination curves of all genotypes were confirmed by
direct DNA sequencing on an ABI PRISM 3100 genetic analyzer
(Applied Biosystems).
Statistical analysis
Hardy-Weinberg equilibrium was examined with the
exact test Power calculation analysis performed using the CaTS
Power Calculator (31).
Genotype-disease association analysis was performed with binary
logistic regression using the SNPStats platform (http://bioinfo.iconcologia.net/SNPstats/) (32). Odds ratios (ORs), 95% confidence
intervals (CIs) and P-values were calculated assuming the
co-dominant (genotypic) model (AA vs. Ab vs. bb) and the recessive
(AA + Ab vs. bb) modes of inheritance. Four phenotypic groups were
searched for the association with the analyzed SNPs compared to the
healthy controls: i) The cervical cancer group; ii) the group of
patients with high-grade precancerous lesions (CIN2/3); iii) the
group of patients with low-grade precancerous lesions (CIN1); and
iv) an overall patient group with abnormal cervical changes (either
cervical cancer or any type of precancerous lesions).
Results
The characteristics of the 10 studied SNPs (gene,
chromosome, chromosomal position, minor allele and minor allele
frequencies) are presented in Table
I. The genotype call rate was ≥98.85%. All studied SNPs were
found to follow the Hardy-Weinberg equilibrium either in the cases
or the controls (exact test, P>0.01) (33). Genotype call rate and P-value (exact
test) for HWE, for each SNP, are presented and Table II.
 | Table I.Characteristics of SNPs genotyped in
the current study. |
Table I.
Characteristics of SNPs genotyped in
the current study.
| SNP | rs number | Gene | Chromosome | Chromosome
position | Minor allele | MAF CEU | MAF in our control
group |
|---|
| 1 | rs1800682 | FAS | 10 | 90739943 | C | 0.45 | 0.44 |
| 2 | rs2290907 | TNRC6C | 17 | 76093677 | C | 0.29 | 0.16 |
| 3 | rs16970849 | TMC8 | 17 | 73645503 | A | 0.14 | 0.04 |
| 4 | rs5757133 | DMC1 | 22 | 37277781 | T | 0.23 | 0.31 |
| 5 | rs3784621 | DUT | 15 | 46420384 | C | 0.46 | 0.20 |
| 6 | rs4737999 | SULF1 | 8 | 70680589 | A | 0.20 | 0.27 |
| 7 | rs9893818 |
TMC8/TMC6 | 17 | 73653762 | A | 0.04 | 0.00 |
| 8 | rs12302655 | OAS3 | 12 | 111858889 | A | 0.13 | 0.00 |
| 9 | rs2894054 | GTF2H4 | 6 | 30980253 | T | 0.09 | 0.12 |
| 10 | rs11177074 | IFNG | 12 | 66830701 | C | 0.14 | 0.07 |
| Table II.Genotype call rate and exact test for
HWE of each SNP in the current study. |
Table II.
Genotype call rate and exact test for
HWE of each SNP in the current study.
|
|
|
|
| Exact test
(P-value) for HWE |
|---|
|
|
|
|
|
|
|---|
| SNP | rs number | Gene | Genotype call rate
(%) | Controls | Cases |
|---|
| 1 | rs1800682 | FAS | 99.18 | 0.096 | 1 |
| 2 | rs2290907 | TNRC6C | 99.67 | 0.83 | 0.36 |
| 3 | rs16970849 | TMC8 | 99.34 | 1 | 1 |
| 4 | rs5757133 | DMC1 | 98.85 | 0.14 | 0.11 |
| 5 | rs3784621 | DUT | 99.34 | 0.85 | 0.6 |
| 6 | rs4737999 | SULF1 | 99.34 | 0.29 | 0.12 |
| 7 | rs9893818 |
TMC8/TMC6 | 99.34 | NA | NA |
| 8 | rs12302655 | OAS3 | 99.84 | 1 | 1 |
| 9 | rs2894054 | GTF2H4 | 99.67 | 0.089 | 0.78 |
| 10 | rs11177074 | IFNG | 98.85 | 0.38 | 0.71 |
The allelic and genotypic frequencies of the studied
SNPs in the control and the overall patient group, as well as in
the cervical cancer, high-grade precancerous lesion and low-grade
precancerous lesion groups are presented in Table III. Of note, as regards rs9893818,
all successfully genotyped participants (100%) carried the CC
genotype, whereas as regards rs12302655, >99.0% of the
participants carried the wild-type genotype.
| Table III.Allelic and genotype frequencies of
SNPs in healthy controls and in cases (cervical cancer cases, cases
with low grade and with high grade precancerous lesions). |
Table III.
Allelic and genotype frequencies of
SNPs in healthy controls and in cases (cervical cancer cases, cases
with low grade and with high grade precancerous lesions).
| SNP |
Genotypes/alleles | Controls n (%) | All cases n
(%) | Cervical cancer n
(%) | High-grade
precancerous lesions, n (%) | Low-grade
precancerous lesions, n (%) |
|---|
| rs1800682 |
|
|
|
|
|
|
|
Genotype | C/C | 63 (22) | 49 (16) | 2 (12) | 13 (13) | 34 (17) |
|
| T/C | 129 (44) | 150 (48) | 8 (47) | 50 (51) | 92 (47) |
|
| T/T | 98 (34) | 114 (36) | 7 (41) | 36 (36) | 71 (36) |
|
| Missed | 2 | 3 | 0 | 1 | 2 |
|
Allele | T | 325 (56) | 378 (60) | 22 (65) | 122 (62) | 234 (59) |
|
| C | 255 (44) | 248 (40) | 12 (35) | 76 (38) | 160 (41) |
| rs2290907 |
|
|
|
|
|
|
|
Genotype | C/C | 8 (3) | 4 (1) | 1 (6) | 1 (1) | 2 (01) |
|
| T/C | 79 (27) | 81 (26) | 3 (0.18) | 19 (0.19) | 59 (0.30) |
|
| T/T | 204 (70) | 230 (73) | 13 (0.76) | 80 (0.80) | 137 (0.69) |
|
| Missed | 1 | 1 | 0 | 0 | 1 |
|
Allele | T | 487 (84) | 541 (86) | 29 (85) | 179 (90) | 333 (84) |
|
| C | 95 (16) | 89 (14) | 5 (15) | 21 (10) | 63 (16) |
| rs16970849 |
|
|
|
|
|
|
|
Genotype | G/A | 24 (8) | 30 (10) | 0 (0) | 10 (10) | 20 (10) |
|
| G/G | 265 (92) | 285 (90) | 17 (100) | 90 (90) | 178 (90) |
|
| A/A | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
|
| Missed | 3 | 1 | 0 | 0 | 1 |
|
Allele | G | 554 (96) | 600 (95) | 34 (100) | 190 (95) | 376 (95) |
|
| A | 24 (4) | 30 (5) | 0 (0) | 10 (5) | 20 (5) |
| rs5757133 |
|
|
|
|
|
|
|
Genotype | C/C | 142 (49) | 156 (50) | 11 (65) | 45 (45) | 100 (51) |
|
| C/T | 114 (39) | 120 (39) | 5 (29) | 43 (43) | 72 (37) |
|
| T/T | 34 (12) | 35 (0.11) | 1 (6) | 11 (11) | 23 (12) |
|
| Missed | 2 | 5 | 0 | 1 | 4 |
|
Allele | C | 398 (69) | 432 (69) | 27 (79) | 133 (67) | 272 (70) |
|
| T | 182 (31) | 190 (31) | 7 (21) | 65 (33) | 118 (30) |
| rs3784621 |
|
|
|
|
|
|
|
Genotype | C/C | 12 (4) | 14 (4) | 2 (12) | 3 (3) | 9 (5) |
|
| T/C | 92 (32) | 97 (31) | 4 (25) | 30 (30) | 63 (32) |
|
| T/T | 187 (64) | 202 (65) | 10 (62) | 66 (67) | 126 (64) |
|
| Missed | 1 | 3 | 1 | 1 | 1 |
|
Allele | T | 466 (80) | 501(80) | 24 (75) | 162 (82) | 315 (80) |
|
| C | 116 (20) | 125 (20) | 8 (25) | 36 (18) | 81 (20) |
| rs4737999 |
|
|
|
|
|
|
|
Genotype | A/A | 24 (8) | 13 (4) | 1 (6) | 6 (6) | 6 (3) |
|
| G/A | 106 (37) | 125 (40) | 7 (41) | 37 (37) | 81 (41) |
|
| G/G | 160 (55) | 176 (56) | 9 (53) | 57 (57) | 110 (56) |
|
| Missed | 2 | 2 | 0 | 0 | 2 |
|
Allele | G | 426 (73) | 477 (76) | 25 (74) | 151 (76) | 301 (76) |
|
| A | 154 (27) | 151 (24) | 9 (26) | 49 (24) | 93 (24) |
| rs9893818 |
|
|
|
|
|
|
|
Genotype | C/C | 290 (100) | 316 (100) | 17 (100) | 100 (100) | 197 (100) |
|
| C/A | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
|
| A/A | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
|
| Missed | 2 | 2 | 0 | 0 | 2 |
|
Allele | C | 580 (100) | 628 (100) | 34 (100) | 200 (100) | 394 (100) |
|
| A | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| rs12302655 |
|
|
|
|
|
|
|
Genotype | G/G | 292 (100) | 313 (99) | 17 (100) | 99 (99) | 197 (99) |
|
| G/A | 0 (0) | 2 (1) | 0 (0) | 1 (1) | 1 (1) |
|
| A/A | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
|
| Missed | 0 | 1 | 0 | 0 | 1 |
|
Allele | G | 584 (100) | 628 (99.7) | 34 (100) | 199 (100) | 395 (100) |
|
| A | 0 (0) | 2 (0.3) | 0 (0) | 1 (0) | 1 (0) |
| rs2894054 |
|
|
|
|
|
|
|
Genotype | C/C | 229 (79) | 246 (78) | 17 (100) | 78 (78) | 151 (76) |
|
| C/T | 54 (19) | 67 (21) | 0 (0) | 22 (22) | 45 (23) |
|
| T/T | 7 (02) | 3 (1) | 0 (0) | 0 (0) | 3 (2) |
|
| Missed | 2 | 0 | 0 | 0 | 0 |
|
Allele | C | 512 (88) | 559 (88) | 34 (100) | 178 (89) | 347 (87) |
|
| T | 68 (12) | 73 (12) | 0 (0) | 22 (11) | 51 (13) |
| rs11177074 |
|
|
|
|
|
|
|
Genotype | C/C | 0 (0) | 1 (0) | 0 (0) | 0 (0) | 1 (1) |
|
| T/C | 42 (15) | 52 (17) | 3 (18) | 18 (18) | 31 (16) |
|
| T/T | 245 (85) | 261 (83) | 14 (82) | 81 (0.82) | 166 (84) |
|
| Missed | 5 | 2 | 0 | 1 | 1 |
|
Allele | T | 532 (93) | 574 (91) | 31 (91) | 180 (0.91) | 363 (92) |
|
| C | 42 (7) | 54 (9) | 3 (9) | 18 (9) | 33 (8) |
Power analysis revealed that our study had a
statistical power of >80.0% to detect an genetic association
with an OR of 1.78, under the assumption of the multiplicative
model, a minor allele frequency of 5% (the lowest in cases for the
rs16970849), a type I error level of 0.05, in a sample size
consisting of 292 controls and 316 cases (data not shown).
Binary logistic regression analysis demonstrated a
significant effect of SULF1 rs4737999 on the risk of the
abnormal cervical changes. In particular, a significant difference
was observed between the controls and the overall patient group
(low-grade, high-grade and cervical cancer) considering the
recessive mode of inheritance (OR, 0.48; 95% CI, 0.24–0.96;
P=0.033). Individuals carrying the AA genotype had almost half a
risk of having cervical cancer, and low- or high-grade lesions
compared to those carrying either the GG or the GA genotypes.
Moreover, this effect was even more potent on the risk of low-grade
precancerous lesions (OR, 0.36; 95% CI, 0.14–0.92; P=0.042) and
(OR, 0.35; 95% CI, 0.14–0.87; P=0.014) in the co-dominant and
recessive models, respectively. Carriers of the rs4737999 AA
genotype were almost 3-fold less likely of having low-grade lesions
compared to carriers of the other genotypes. No other SNP was found
to alter the risk of any examined phenotype (Table IV).
| Table IV.Single locus analysis. |
Table IV.
Single locus analysis.
|
|
| All cases (n=316)
vs. controls (n=292) | Low-grade (n=199)
vs. controls (n=292) | High-grade (n=100)
vs. controls (n=292) | Cancer (n=17) vs.
controls (n=292) |
|---|
|
|
|
|
|
|
|
|---|
| SNP | Genotype | OR (95% CI) | P-value | OR (95% CI) | P-value | OR (95% CI) | P-value | OR (95% CI) | P-value |
|---|
| rs1800682 |
|
|
|
|
|
|
|
|
|
|
Codominant | T/T | 1.00 | 0.16 | 1.00 | 0.47 | 1.00 | 0.15 | 1.00 | 0.54 |
|
| T/C | 1.00
(0.70–1.43) |
| 0.98
(0.66–1.48) |
| 1.06
(0.64–1.74) |
| 0.87
(0.30–2.48) |
|
|
| C/C | 0.67
(0.42–1.06) |
| 0.74
(0.44–1.25) |
| 0.56
(0.28–1.14) |
| 0.44
(0.09–2.21) |
|
|
Recessive | T/T-T/C | 1.00 | 0.056 | 1.00 | 0.22 | 1.00 | 0.054 | 1.00 | 0.84 |
|
| C/C | 0.67
(0.44–1.01) |
| 0.75
(0.47–1.19) |
| 0.54
(0.29–1.04) |
| 0.48
(0.11–2.16) |
|
| rs2290907 |
|
|
|
|
|
|
|
|
|
|
Codominant | T/T | 1.00 | 0.37 | 1.00 | 0.33 | 1.00 | 0.12 | 1.00 | 0.57 |
|
| T/C | 0.91
(0.63–1.31) |
| 1.11
(0.74–1.66) |
| 0.61
(0.35–1.08) |
| 0.60
(0.17–2.15) |
|
|
| C/C | 0.44
(0.13–1.49) |
| 0.37
(0.08–1.78) |
| 0.32
(0.04–2.59) |
| 1.96
(0.23–16.89) |
|
|
Recessive | T/T-T/C | 1.00 | 0.19 | 1.00 | 0.16 | 1.00 | 0.27 | 1.00 | 0.51 |
|
| C/C | 0.45
(0.14–1.53) |
| 0.36
(0.08–1.72) |
| 0.36
(0.04–2.89) |
| 2.21
(0.26–18.77) |
|
| rs16970849 |
|
|
|
|
|
|
|
|
|
|
| G/G | 1.00 | 0.6 | 1.00 | 0.5 | 1.00 | 0.61 | 1.00 | 0.091 |
|
| G/A | 1.16
(0.66–2.04) |
| 1.24
(0.67–2.31) |
| 1.23
(0.56–2.66) |
| 0.00 (0.00-NA) |
|
| rs5757133 |
|
|
|
|
|
|
|
|
|
|
Codominant | C/C | 1.00 | 0.95 | 1.00 | 0.86 | 1.00 | 0.77 | 1.00 | 0.42 |
|
| T/C | 0.96
(0.68–1.35) |
| 0.90
(0.61–1.33) |
| 1.19
(0.73–1.93) |
| 0.57
(0.19–1.68) |
|
|
| T/T | 0.94
(0.55–1.58) |
| 0.96
(0.53–1.73) |
| 1.02
(0.48–2.18) |
| 0.38
(0.05–3.04) |
|
|
Recessive | C/C-T/C | 1.00 | 0.86 | 1.00 | 0.98 | 1.00 | 0.87 | 1.00 | 0.42 |
|
| T/T | 0.95
(0.58–1.58) |
| 1.01
(0.57–1.77) |
| 0.94
(0.46–1.94) |
| 0.47
(0.06–3.66) |
|
| rs3784621 |
|
|
|
|
|
|
|
|
|
|
Codominant | T/T | 1.00 | 0.97 | 1.00 | 0.97 | 1.00 | 0.84 | 1.00 | 0.4 |
|
| T/C | 0.98
(0.69–1.38) |
| 1.02
(0.69–1.50) |
| 0.92
(0.56–1.52) |
| 0.81
(0.25–2.66) |
|
|
| C/C | 1.08
(0.49–2.39) |
| 1.11
(0.46–2.72) |
| 0.71
(0.19–2.59) |
| 3.12
(0.61–15.85) |
|
|
Recessive | T/T-T/C | 1.00 | 0.83 | 1.00 | 0.82 | 1.00 | 0.62 | 1.00 | 0.19 |
|
| C/C | 1.09
(0.49–2.39) |
| 1.11
(0.46–2.68) |
| 0.73
(0.20–2.63) |
| 3.32
(0.68–16.29) |
|
| rs4737999 |
|
|
|
|
|
|
|
|
|
|
Codominant | G/G | 1.00 | 0.096 | 1.00 | 0.042 | 1.00 | 0.75 | 1.00 | 0.89 |
|
| G/A | 1.07
(0.77–1.50) |
| 1.11
(0.76–1.62) |
| 0.98
(0.61–1.59) |
| 1.17
(0.42–3.25) |
|
|
| A/A | 0.49
(0.24–1.00) |
| 0.36
(0.14–0.92) |
| 0.70
(0.27–1.80) |
| 0.74
(0.09–6.11) |
|
|
Recessive | G/G-G/A | 1.00 | 0.033 | 1.00 | 0.014 | 1.00 | 0.45 | 1.00 | 0.71 |
|
| A/A | 0.48
(0.24–0.96) |
| 0.35
(0.14–0.87) |
| 0.71
(0.28–1.78) |
| 0.69
(0.09–5.45) |
|
| rs9893818 |
| NA |
| NA |
| NA |
| NA |
|
| rs12302655 |
|
|
|
|
|
|
|
|
|
|
| G/G | 1.00 | 0.1 | 1.00 | 0.18 | 1.00 | 0.098 | NA | NA |
|
| G/A | NA (0.00-NA) |
| NA (0.00-NA) |
| NA (0.00-NA) |
| NA |
|
| rs2894054 |
|
|
|
|
|
|
|
|
|
|
Codominant | C/C | 1.00 | 0.28 | 1.00 | 0.46 | 1.00 | 0.1 | 1.00 | 0.02 |
|
| T/C | 1.15
(0.77–1.72) |
| 1.26
(0.81–1.97) |
| 1.20
(0.68–2.09) |
| 0.00 (0.00-NA) |
|
|
| T/T | 0.40
(0.10–1.56) |
| 0.65
(0.17–2.55) |
| 0.00 (0.00-NA) |
| 0.00 (0.00-NA) |
|
|
Recessive | C/C-T/C | 1.00 | 0.15 | 1.00 | 0.48 | 1.00 | 0.041 | 1.00 | 0.37 |
|
| T/T | 0.39
(0.10–1.51) |
| 0.62
(0.16–2.42) |
| 0.00 (0.00-NA) |
| 0.00 (0.00-NA) |
|
| rs11177074 |
|
|
|
|
|
|
|
|
|
|
Codominant | T/T | 1.00 | 0.42 | 1.00 | 0.39 | 1.00 | 0.41 | 1.00 | 0.74 |
|
| T/C | 1.16
(0.75–1.81) |
| 1.09
(0.66–1.80) |
| 1.30
(0.71–2.38) |
| 1.25
(0.34–4.54) |
|
|
| C/C | NA (0.00-NA) |
| NA (0.00-NA) |
| NA |
| NA |
|
|
Recessive | T/T-T/C | 1.00 | 0.25 | 1.00 | 0.18 | NA | NA | NA | NA |
|
| C/C | NA (0.00-NA) |
| NA (0.00-NA) |
| NA |
| NA |
|
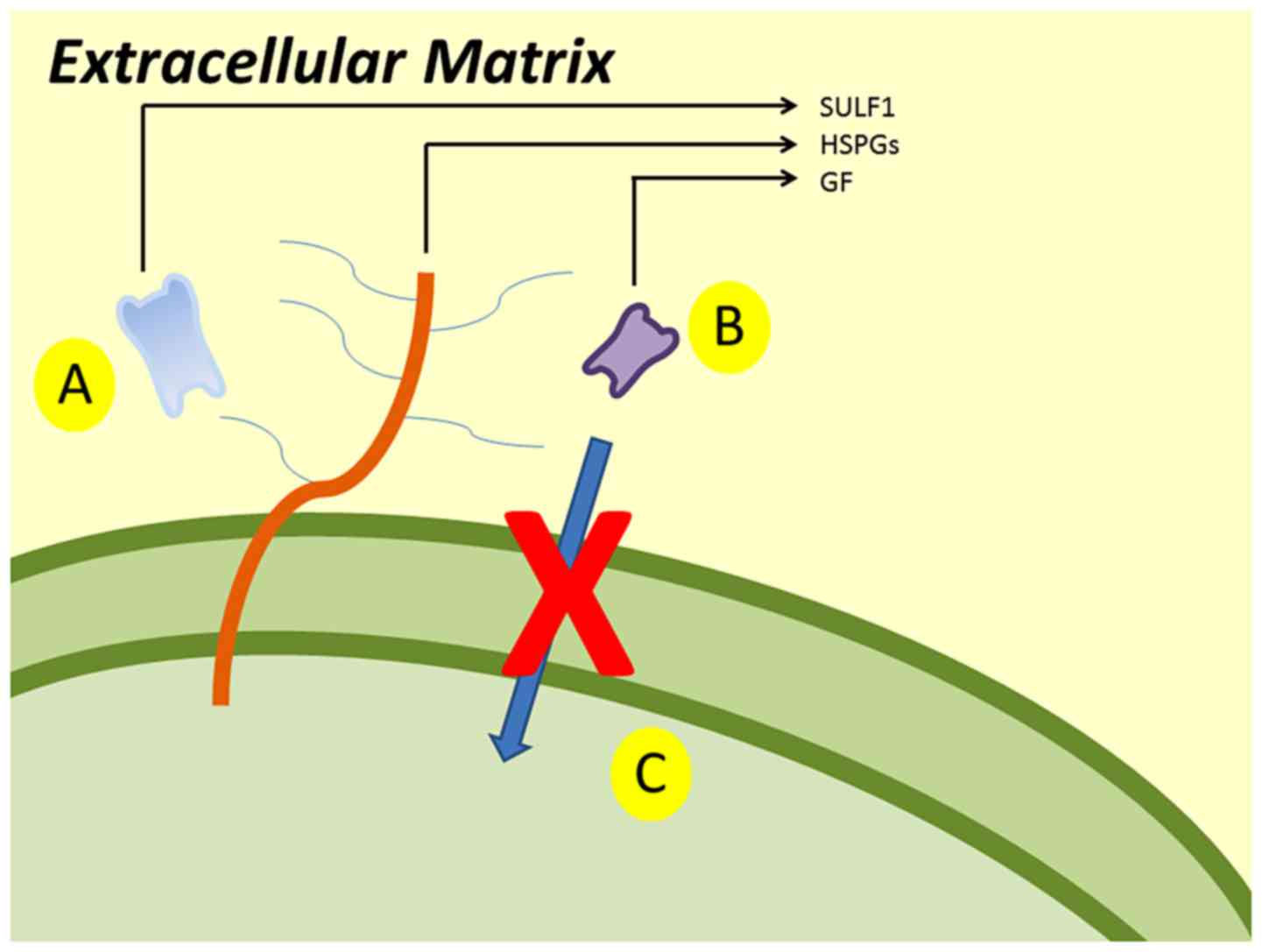
The main mechanism of SULF1 gene is presented
in Fig. 1. SULF1 gene encodes
the SULF1 protein. SULF1 is a heparin-degrading endosulfatase,
which desulfates HSPGs and blocks the binding of growth factors and
their receptors. Consequently, it inhibits the activation of growth
factor and the signaling pathways.
Discussion
In the present study, we tried to replicate the
findings of previous studies regarding the role of SNPs in DNA
repair, viral infection and cell entry, and their effects on the
risk of cervical cancer and precancerous lesions (14,24). In
addition, we re-examined an independent Greek cohort in order to
determine the influence of the rs1800682 (FAS), rs2290907
(TMC6) and rs16970849 (TMC8) gene variants (3). In the present study, we found that a
specific variant of the SULF1 gene, rs4737999, was
associated with a significantly decreased risk of developing
precancerous lesions and cervical cancer.
The SULF1 gene is located at the 8q13.3
region. It encodes the homonymous protein, a heparin-degrading
endosulfatase, which desulfates HSPGs and blocks the binding of
growth factors and their receptors and as a result it inhibits the
activation of growth factor and the signaling pathways (Fig. 1) (17,18). The
expression of SULF1 appears to be stable in normal tissues,
whereas it is downregulated in several tumor cells (34). Moreover, the proliferation and
migration of tumor cells can be inhibited by the re-expression of
the SULF1 gene (35).
In the study by Wang et al (14), the SULF1 gene reached a
statistically significant threshold for the CIN3/Cancer group
compared to the controls (P=0.0030). Moreover, the SULF1
gene was also associated with HPV persistence (P=0.005).
Additionally, according to SNP-Based association analysis, when the
CIN3/Cancer group was compared to the control group, 3 out of the
77 examined SULF1 SNPs (rs4737999, rs4284050 and rs10108002)
achieved statistical significance (P-value trend <0.05). In this
analysis of Wang et al, the strongest association was
reported for the rs4737999; this polymorphism also was associated
with precancerous lesions and cervical cancer in the present
study.
A number of SNPs of the SULF1 gene have been
found to influence the risk of cancer. The AA genotype of r3802278,
a SNP located in the 3′-untranslated region (3′-UTR) of the
SULF1 gene, was found to play a protective role against
breast cancer (36). Moreover,
rs2623047, a 5′-upstream gene variant in SULF1, has been
associated with an increased risk of breast cancer, as well as with
an earlier age of onset and the survival of ovarian cancer
(37). Finally, rs6990375, a 3′ prime
UTR variant, has been associated with earlier age of ovarian cancer
(38). The SNP rs4737999, that
reached a statistically significant threshold in the present study,
is an intronic non-coding variant located between exons 13 and 14.
Therefore, the SULF1 gene may represent an important locus
linked to tumorigenesis, as SNPs located in the 5′-upstream region,
in the 3′-UTR region or even in the middle of the gene have been
found to alter the risk of cancer.
A number of studies have reported that the
FAS-670 gene promoter polymorphism is associated with
cervical carcinogenesis (39–43). Moreover, the expression of the
FAS/FASL genes and the CD95-CD95 ligand (FAS/FASL)
interaction seem to confer susceptibility to the development of
cervical cancer (3). However, the
present study failed to detect any significant effect of the
FAS gene SNPs on the risk of cervical cancer or any
precancerous lesion. This is in accordance with the results of our
previous study in another Greek cohort (3). It is possible that ethnic differences in
FAS gene variability may account for the different results
among populations (44).
In conclusion, the present study confirms the
finidngs of previous reports regarding the role of SULF1 in
the risk of precancerous lesions and cervical cancer. This
association may have prognostic and pharmacogenetic implications to
precancerous lesions or cervical cancer, as SULF1 may be
considered as a therapeutic target or biomarker (45,46). Our
findings need to be replicated in other populations of other ethnic
backgrouns and in experimental models, in order to elucidate the
possible role of polymorphic variants of the SULF1 gene in
the pathophysiology of mechanism of tumorigenesis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
Eda and VS were involved in the conceptualization of
the study, data curation, formal analysis and investigation,
methodology, project administration, resources, software, study
supervision, validation, writing of the original draft and writing,
reviewing and editing the manuscript. AG, EP, MK, GX, ED, GG and
ENK provided the patient blood samples and clinical information,
and were involved in data validation, and in the writing, reviewing
and editing of the manuscript. DAS and AT were involved in data
investigation and validation, as well as in the writing, reviewing
and editing of the manuscript. AD was involved in the
conceptualization of the study, data curation, formal analysis and
investigation, methodology, project administration, resources,
software, study supervision, validation, writing of the original
draft and writing, reviewing and editing the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures performed in studies involving human
participants were in accordance with the ethical standards of the
institutional and/or national research committee and with the 1964
Helsinki declaration and its later amendments or comparable ethical
standards. The local Ethics Review Board of the University Hospital
of Larissa approved the study protocol. Informed consent was
obtained from all individual participants included in the
study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SNP
|
single nucleotide polymorphism
|
|
FAS
|
Fas cell surface death receptor
|
|
TNRC6C
|
trinucleotide repeat containing 6C
|
|
TMC8
|
transmembrane channel like 8
|
|
DMC1
|
DNA meiotic recombinase 1
|
|
DUT
|
deoxyuridine triphosphatase
|
|
SULF1
|
sulfatase 1
|
|
OAS3
|
2′-5-oligoadenylate synthetase 3
|
|
GTF2H4
|
general transcription factor IIH
subunit 4
|
|
IFNG
|
interferon gamma
|
|
HPV
|
human papillomavirus
|
|
TMC6
|
transmembrane channel like 6
|
|
OR
|
odds ratio
|
|
CI
|
confidence interval
|
|
TNF
|
tumor necrosis factor
|
|
HSPGs
|
heparan sulfate proteoglycans
|
|
3′-UTR
|
3′-untranslated region
|
|
CD95
|
cluster of differentiation 95
|
|
FASL
|
Fas ligand
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pavlidou E, Daponte A, Egea R, Dardiotis
E, Hadjigeorgiou GM, Barbadilla A and Agorastos T: Genetic
polymorphisms of FAS and EVER genes in a Greek population and their
susceptibility to cervical cancer: A case control study. BMC
Cancer. 16:9232016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schoell WM, Janicek MF and Mirhashemi R:
Epidemiology and biology of cervical cancer. Semin Surg Oncol.
16:203–211. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boda D, Docea AO, Calina D, Ilie MA,
Caruntu C, Zurac S, Neagu M, Constantin C, Branisteanu DE,
Voiculescu V, et al: Human papilloma virus: Apprehending the link
with carcinogenesis and unveiling new research avenues (Review).
Int J Oncol. 52:637–655. 2018.PubMed/NCBI
|
|
6
|
Libra M, Scalisi A, Vella N, Clementi S,
Sorio R, Stivala F, Spandidos DA and Mazzarino C: Uterine cervical
carcinoma: Role of matrix metalloproteinases (Review). Int J Oncol.
34:897–903. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bosch FX, Lorincz A, Muñoz N, Meijer CJLM
and Shah KV: The causal relation between human papillomavirus and
cervical cancer. J Clin Pathol. 55:244–265. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baritaki S, Sifakis S, Huerta-Yepez S,
Neonakis IK, Soufla G, Bonavida B and Spandidos DA: Overexpression
of VEGF and TGF-beta1 mRNA in Pap smears correlates with
progression of cervical intraepithelial neoplasia to cancer:
Implication of YY1 in cervical tumorigenesis and HPV infection. Int
J Oncol. 31:69–79. 2007.PubMed/NCBI
|
|
9
|
Castellsagué X and Muñoz N: Chapter 3:
Cofactors in human papillomavirus carcinogenesis - role of parity,
oral contraceptives, and tobacco smoking. J Natl Cancer Inst
Monogr. 2003:20–28. 2003. View Article : Google Scholar
|
|
10
|
Magnusson PK, Lichtenstein P and
Gyllensten UB: Heritability of cervical tumours. Int J Cancer.
88:698–701. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mammas IN, Zafiropoulos A and Spandidos
DA: Involvement of the ras genes in female genital tract cancer.
Int J Oncol. 26:1241–1255. 2005.PubMed/NCBI
|
|
12
|
Koffa M, Koumantakis E, Ergazaki M,
Malamoumitsi V and Spandidos D: Detection of ras gene-mutations and
hpv in lesions of the human female reproductive-tract. Int J Oncol.
5:189–195. 1994.PubMed/NCBI
|
|
13
|
Chen X, Jiang J, Shen H and Hu Z: Genetic
susceptibility of cervical cancer. J Biomed Res. 25:155–164. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang SS, Gonzalez P, Yu K, Porras C, Li Q,
Safaeian M, Rodriguez AC, Sherman ME, Bratti C, Schiffman M, et al:
Common genetic variants and risk for HPV persistence and
progression to cervical cancer. PLoS One. 5:e86672010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Harada H, Nagai H, Tsuneizumi M, Mikami I,
Sugano S and Emi M: Identification of DMC1, a novel gene in the TOC
region on 17q25.1 that shows loss of expression in multiple human
cancers. J Hum Genet. 46:90–95. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mol CD, Harris JM, McIntosh EM and Tainer
JA: Human dUTP pyrophosphatase: Uracil recognition by a beta
hairpin and active sites formed by three separate subunits.
Structure. 4:1077–1092. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ji W, Yang J, Wang D, Cao L, Tan W, Qian
H, Sun B, Qian Q, Yin Z, Wu M, et al: hSulf-1 gene exhibits
anticancer efficacy through negatively regulating VEGFR-2 signaling
in human cancers. PLoS One. 6:e232742011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu H, Fu X, Ji W, Liu K, Bao L, Yan Y, Wu
M, Yang J and Su C: Human sulfatase-1 inhibits the migration and
proliferation of SMMC-7721 hepatocellular carcinoma cells by
downregulating the growth factor signaling. Hepatol Res.
43:516–525. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thamizhmani R and Vijayachari P:
Association of dengue virus infection susceptibility with
polymorphisms of 2′-5′-oligoadenylate synthetase genes: a
case-control study. Braz J Infect Dis. 18:548–550. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gervais V, Lamour V, Jawhari A, Frindel F,
Wasielewski E, Dubaele S, Egly JM, Thierry JC, Kieffer B and
Poterszman A: TFIIH contains a PH domain involved in DNA nucleotide
excision repair. Nat Struct Mol Biol. 11:616–622. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun Y, Lu Y, Pen Q, Li T, Xie L, Deng Y
and Qin A: Interferon gamma +874 T/A polymorphism increases the
risk of cervical cancer: Evidence from a meta-analysis. Tumour
Biol. 36:4555–4564. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Orth G: Host defenses against human
papillomaviruses: Lessons from epidermodysplasia verruciformis.
Curr Top Microbiol Immunol. 321:59–83. 2008.PubMed/NCBI
|
|
23
|
Lazarczyk M, Dalard C, Hayder M, Dupre L,
Pignolet B, Majewski S, Vuillier F, Favre M and Liblau RS: EVER
proteins, key elements of the natural anti-human papillomavirus
barrier, are regulated upon T-cell activation. PLoS One.
7:e399952012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Castro FA, Ivansson EL, Schmitt M,
Juko-Pecirep I, Kjellberg L, Hildesheim A, Gyllensten UB and
Pawlita M: Contribution of TMC6 and TMC8 (EVER1 and EVER2) variants
to cervical cancer susceptibility. Int J Cancer. 130:349–355. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Contreras DN, Krammer PH, Potkul RK, Bu P,
Rossi JL, Kaufmann AM, Gissmann L and Qiao L: Cervical cancer cells
induce apoptosis of cytotoxic T lymphocytes. J Immunother.
23:67–74. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dybikowska A, Sliwinski W, Emerich J and
Podhajska AJ: Evaluation of Fas gene promoter polymorphism in
cervical cancer patients. Int J Mol Med. 14:475–478.
2004.PubMed/NCBI
|
|
27
|
Butler LM, Hewett PJ, Butler WJ and Cowled
PA: Down-regulation of Fas gene expression in colon cancer is not a
result of allelic loss or gene rearrangement. Br J Cancer.
77:1454–1459. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee SH, Shin MS, Park WS, Kim SY, Dong SM,
Pi JH, Lee HK, Kim HS, Jang JJ, Kim CS, et al: Alterations of Fas
(APO-1/CD95) gene in transitional cell carcinomas of urinary
bladder. Cancer Res. 59:3068–3072. 1999.PubMed/NCBI
|
|
29
|
Shimonishi T, Isse K, Shibata F, Aburatani
I, Tsuneyama K, Sabit H, Harada K, Miyazaki K and Nakanuma Y:
Up-regulation of fas ligand at early stages and down-regulation of
Fas at progressed stages of intrahepatic cholangiocarcinoma reflect
evasion from immune surveillance. Hepatology. 32:761–769. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang Q, Wang J and Hu Y: FAS-670 gene
polymorphism and cervical carcinogenesis risk: A meta-analysis.
Biomed Rep. 1:889–894. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Skol AD, Scott LJ, Abecasis GR and Boehnke
M: Joint analysis is more efficient than replication-based analysis
for two-stage genome-wide association studies. Nat Genet.
38:209–213. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Solé X, Guinó E, Valls J, Iniesta R and
Moreno V: SNPStats: A web tool for the analysis of association
studies. Bioinformatics. 22:1928–1929. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ziegler A, Van Steen K and Wellek S:
Investigating Hardy-Weinberg equilibrium in case-control or cohort
studies or meta-analysis. Breast Cancer Res Treat. 128:197–201.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Narita K, Staub J, Chien J, Meyer K, Bauer
M, Friedl A, Ramakrishnan S and Shridhar V: HSulf-1 inhibits
angiogenesis and tumorigenesis in vivo. Cancer Res. 66:6025–6032.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lai J, Chien J, Staub J, Avula R, Greene
EL, Matthews TA, Smith DI, Kaufmann SH, Roberts LR and Shridhar V:
Loss of HSulf-1 up-regulates heparin-binding growth factor
signaling in cancer. J Biol Chem. 278:23107–23117. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou Q, Jiang Y, Yin W, Wang Y and Lu J:
Single-nucleotide polymorphism in microRNA-binding site of SULF1
target gene as a protective factor against the susceptibility to
breast cancer: A case-control study. Onco Targets Ther.
9:2749–2757. 2016.PubMed/NCBI
|
|
37
|
Okolicsanyi RK, Faure M, Jacinto JM,
Chacon-Cortes D, Chambers S, Youl PH, Haupt LM and Griffiths LR:
Association of the SNP rs2623047 in the HSPG modification enzyme
SULF1 with an Australian Caucasian breast cancer cohort. Gene.
547:50–54. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Han CH, Huang YJ, Lu KH, Liu Z, Mills GB,
Wei Q and Wang LE: Polymorphisms in the SULF1 gene are associated
with early age of onset and survival of ovarian cancer. J Exp Clin
Cancer Res. 30:52011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tamandani Kordi DM, Sobti RC and Shekari
M: Association of Fas-670 gene polymorphism with risk of cervical
cancer in North Indian population. Clin Exp Obstet Gynecol.
35:183–186. 2008.PubMed/NCBI
|
|
40
|
Ueda M, Terai Y, Kanda K, Kanemura M,
Takehara M, Yamaguchi H, Nishiyama K, Yasuda M and Ueki M: Fas gene
promoter −670 polymorphism in gynecological cancer. Int J Gynecol
Cancer. 16 Suppl 1:179–182. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nunobiki O, Ueda M, Toji E, Yamamoto M,
Akashi K, Sato N, Izuma S, Torii K, Tanaka I, Okamoto Y, et al:
Genetic polymorphism of cancer susceptibility genes and HPV
infection in cervical carcinogenesis. Pathol Res Int.
2011:3640692011. View Article : Google Scholar
|
|
42
|
Lai HC, Lin WY, Lin YW, Chang CC, Yu MH,
Chen CC and Chu TY: Genetic polymorphisms of FAS and FASL
(CD95/CD95L) genes in cervical carcinogenesis: An analysis of
haplotype and gene-gene interaction. Gynecol Oncol. 99:113–118.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lai HC, Sytwu HK, Sun CA, Yu MH, Yu CP,
Liu HS, Chang CC and Chu TY: Single nucleotide polymorphism at Fas
promoter is associated with cervical carcinogenesis. Int J Cancer.
103:221–225. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tan SC, Ismail MP, Duski DR, Othman NH and
Ankathil R: FAS c.-671A>G polymorphism and cervical cancer risk:
A case-control study and meta-analysis. Cancer Genet. 211:18–25.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hammond E, Khurana A, Shridhar V and
Dredge K: The role of heparanase and sulfatases in the modification
of heparan sulfate proteoglycans within the tumor microenvironment
and opportunities for novel cancer therapeutics. Front Oncol.
4:1952014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hur K, Han TS, Jung EJ, Yu J, Lee HJ, Kim
WH, Goel A and Yang HK: Up-regulated expression of sulfatases
(SULF1 and SULF2) as prognostic and metastasis predictive markers
in human gastric cancer. J Pathol. 228:88–98. 2012.PubMed/NCBI
|