Introduction
According to the World Health Organization (WHO),
central nervous system (CNS) tumors can be subdivided into primary
and secondary tumors. Secondary tumors develop from cells that
spread to the brain from a cancer in another part of the body.
Primary brain tumors, including medulloblastoma, choroid plexus
papilloma, pituitary tumor, neurocytoma, germ cell tumor and glioma
in the majority of cases start from local cells of the brain
(1). Given the ability of brain
tumors to infiltrate and a high rate of recurrence, the prognosis
is usually poor even though multimodal therapies have been applied
(2). For example, it was reported
that the median survival is ≤15 months for patients with grade IV
glioma (glioblastoma, GBM) (2).
The cancer stem cell (CSC) hypothesis originates
from the hierarchical cellular organization within a tumor. Indeed,
CSCs could be enriched and expanded in vitro, and more
importantly they could initiate the formation of new tumors
following xenotransplantation (3–7). An
increasing number of studies suggested that purified CSCs grown
in vitro could be applied for studying the initiation and
progression of tumor as well as chemo- or radiotherapy resistance,
tumor relapse and testing for new therapeutic drugs (3,5). In
general, suspension spheroid-formation culture protocols are used
for CSC enrichment. Solid brain tumor mass is disassociated into
single cells and then grown in a non-adherent serum-free culture
with basic fibroblast growth factor (bFGF) and epidermal growth
factor (EGF) to trigger spheroid formation (6). By using this suspension culture
technique, CSCs have been successfully cultured from various brain
tumors, including glioblastoma, medulloblastoma and
oligodendroglioma (4,7–9). However,
the efficiency for initial growing tumorspheres from brain tumors
may be low and variable (10–12). Furthermore, the spheroid culture
method tends to enrich CSCs with specific molecular profiles,
including phosphatase and tensin homolog deficiency, wild-type
isocitrate dehydrogenase 1, amplified chromosome 7 and deleted
chromosome 10q (4,13).
Previously, two groups reported that they developed
a new adherent culture protocol for growing brain CSCs (14,15). In
this protocol, freshly resected glioblastomas from patients were
digested into single cells and then seeded onto laminin-coated
culture surface under standard neural stem cell culture conditions
(14,15). Through this advanced cell culture
protocol, brain CSCs could be enriched as an adherent monolayer,
which displayed a great potential for initiating the formation of
new tumors in vivo (14,15).
In the present study, the efficiency of deriving
brain CSCs was investigated by using this adherent culture method.
Whether the adherent culture method may be applied to a wide
spectrum of brain tumors or specifically to glioblastomas was also
examined. The results indicated that CSCs from glioblastomas but
not other types of brain tumors could be grown through adherent
culture and the derivation efficiency was ~50%. It was also found
that CD133+/Sox2+ cells within the
glioblastomas might be the original cells, which were enriched
under adherent culture conditions.
Materials and methods
Ethics statement
The entire study was approved by the Institutional
Review Board of the Shanghai Tenth People's Hospital (Shanghai,
China). All patients provided written informed consent to the
surgical procedures and gave permission for the use of resected
tissue specimens. All animal protocols were approved by the
Institutional Animal Care and Use Committee of Tongji University
(Shanghai, China). A total of 20 male Balb/c nude mice (18-20 g, 6
weeks old) purchased from Shanghai Slaccas Experimental Animal
Limited Company (Shanghai, China), were housed in specific
pathogen-free conditions throughout the experiments that were
automatically maintained at a temperature of 23±2°C, a relative
humidity of 45-65%, and a controlled 12/12 h light/dark cycle.
Animals had ad libitum access to food and water.
Brain tumor samples
Tumor specimens were obtained from 18 patients with
CNS tumors who underwent surgical procedures at the neurosurgery
department, the Shanghai Tenth People's Hospital. Tumors were
classified by histopathologic examination according to the WHO
classification, including the cytokeratin AE1/AE3, epithelial tumor
marker (16). The clinical profiles
of the patients were obtained from medical records, and patient
characteristics are listed in Table
I.
 | Table I.Adherent CSCs could only be enriched
from glioblastomas. |
Table I.
Adherent CSCs could only be enriched
from glioblastomas.
|
|
|
|
|
|
|
Pathological examination (paraffin
section/immunohistochemistry/immunofluorescence) |
|---|
|
|
|
|
|
|
|
|
|---|
| Patient no. | Age | Sex | Tumor
localization | Clinical
diagnosis | Growth of CSCs | GFAP | p53 | Vim | S100 | Ki67 (%) | CGA | SYN | AE1/AE3 | EMA | EGFR | CD34 |
|---|
| 1 | 17 | M | Right insula | Germ cell
tumors | No | – | ++ | N/A | + | 90 | – | – | – | – | N/A | + |
| 2 | 64 | M | Cerebellum | Astrocytoma (WHO
II) | No | + | N/A | + | + | 2 | N/A | N/A | – | N/A | N/A | + |
| 3 | 71 | M | Left
temporal-occipital | Glioblastoma (WHO
IV) | Yes | + | N/A | + | + | 30 | – | – | – | N/A | N/A | + |
| 5 | 61 | M | Left temporal
lobe | Astrocytoma (WHO
III) | No | + | N/A | + | + | 30 | N/A | N/A | – | – | N/A | + |
| 6 | 62 | M | Frontal lobe | Glioblastoma (WHO
I) | Yes | + | N/A | + | + | 5 | N/A | N/A | – | – | N/A | + |
| 7 | 46 | M | Frontal lobe | Glioblastoma (WHO
III) | No | +/– | + | + | + | 50 | – | +/– | N/A | + | N/A | + |
| 8 | 14 | F | Lateral
ventricle | Ependymoma (WHO
II) | No | + | + | + | + | 4 | N/A | +/– | – | N/A | N/A | N/A |
| 9 | 21 | F | Bilateral
ventricle | Neurocytoma (WHO
II) | No | + | + | N/A | + | 15 | + | + | N/A | N/A | – | + |
| 10 | 46 | F | Left frontoparietal
lobe | Glioblastoma (WHO
IV) | No | + | N/A | N/A | + | 25 | – | N/A | – | – | N/A | + |
| 11 | 74 | M | Corpus
callosum | Glioblastoma (WHO
IV) | No | + | ++ | N/A | – | 30 | – | – | N/A | – | + | N/A |
| 12 | 32 | M | Corpus
callosum | Astrocytoma (WHO
III) | No | + | ++ | N/A | N/A | 10 | N/A | N/A | N/A | N/A | N/A | N/A |
| 13 | 44 | F | Thalamus | Glioblastoma (WHO
IV) | No | + | + | + | + | 20 | – | – | – | – | ++ | N/A |
| 14 | 37 | M | Right frontal
lobe | Astrocytoma (WHO
III) | No | + | ++ | + | + | 3 | N/A | N/A | – | + | + | + |
| 15 | 61 | M | Left frontal
lobe | Glioblastoma (WHO
IV) | Yes | + | N/A | + | + | 30 | – | – | – | – | + | + |
| 16 | 60 | M | Right temporal
lobe | Glioblastoma (WHO
IV) | No | + | N/A | N/A | + | 40 | – | – | – | – | N/A | + |
| 17 | 39 | M | Frontal lobe | Astrocytoma (WHO
I) | No | + | +/– | + | + | 1 | – | – | – | – | N/A | + |
| 18 | 54 | M | Right frontal
lobe | Glioblastoma (WHO
IV) | Yes | + | + | + | + | 25 | N/A | N/A | N/A | – | N/A | + |
| 19 | 65 | F | Right temporal
lobe | Metastatic
lung | No | – | +/– | – | + | 1 | – | – | + | + | N/A | + |
|
|
|
|
| adenocarcinoma |
Isolation of primary brain CSCs
The standard CSC enrichment protocols (14,15) were
followed, which resemble the culture conditions for normal neural
stem cells. The tumor tissues were minced with a scissor and then
digested with accutase (A11105; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) for 20 min. The cells were then triturated
into single cells and plated on culture surfaces precoated with
laminin for ≥3 h at 37°C. The formula for the culture medium
(DMEM/F12/N2/B27) was, DMEM/F12 (catalog no. 11330; Gibco; Thermo
Fisher Scientific, Inc.), 1× N2 (catalog no. 17502; Gibco; Thermo
Fisher Scientific, Inc.), 1× B27 supplement (catalog no. 17504;
Gibco; Thermo Fisher Scientific, Inc.) and 10 ng/ml EGF (catalog
no. E9644; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 10 ng/ml
bFGF (catalog no. 100-18B; PeproTech, Inc., Rocky Hill, NJ, USA).
The cells were attached overnight, and cell debris together with
floating red blood cells and tumor cells were removed on the next
day. The established CSC lines were regularly passaged every 5-6
days by using accutase for digestion.
Mice brain tissue collection
Male C57BL/6 nude mice (6 weeks) were purchased from
Slaccas Experimental Animals LLC, Shanghai Slaccas (Shanghai,
China). The mice were deeply anesthetized with avertin
(Sigma-Aldrich; Merck KGaA) and fixed with 4% paraformaldehyde in
PB for 15 min via transcardial perfusion. The brains were excised
and dehydrated in gradient sucrose. The frozen sections were cut at
40 µm in thickness using a freezing microtome (Leica Microsystems,
Inc., Buffalo Grove, IL, USA).
Immunostaining
The expression levels of CD133, Nestin, Sox2 and
GFAP in brain tumor tissues, normal mice brain or cultured cells
were analyzed by immunofluorescence staining. The resected brain
tumors were fixed with 4% paraformaldehyde (PFA) for 3 h,
dehydrated by sucrose and sectioned using a microtome. The cultured
cells were plated on laminin-coated coverslips and subsequently
were used for immunostaining. The sections or 4% PFA
fixed-coverslips with cultured cells were blocked with 10% normal
donkey serum (Beijing Biodee Biotechnology Co., Ltd., Beijing,
China) and 0.1% Triton X-100/PBS for 1 h at room temperature
followed by incubation with primary antibodies overnight at 4°C. On
the next day, after adequate washing, the tissue sections or
coverslips were incubated with the appropriate fluorescence-labeled
secondary antibodies for 40 min at room temperature. The antibodies
used in this study were: Anti-CD133 (1:50; catalog no. ARH4033;
Antibody Revolution Inc., San Diego, USA), anti-Nestin (1:500;
catalog no. MAB5326; EMD Millipore, Billerica, MA, USA), anti-Sox2
(1:500; catalog no. AF2018; R&D Systems, Inc., Minneapolis, MN,
USA), anti-GFAP (1:1,000; catalog no. z0334; Dako A/S, Glostrup,
Denmark), Peroxidase-conjugated donkey anti-goat (1:1,000, catalog
no. 705035003; Jackson ImmunoResearch Inc., West Grove, PA, USA),
Peroxidase-conjugated goat anti-mouse (1:1,000, catalog no.
115035003; Jackson ImmunoResearch Inc.) and Peroxidase-conjugated
goat anti-rabbit (1:1,000, catalog no. 111035003; Jackson
ImmunoResearch Inc.). The nuclei were counterstained with Hoechst
solution for 5 min at room temperature. The images were taken with
a confocal microscope (SP-5; Leica Microsystems, Inc.).
Statistical analysis
The data were statistically analyzed using
Statistical Package for the Social Sciences (SPSS) statistical
(version 21) software from SPSS, Inc., (Chicago, IL, USA) and using
one-way analysis of variance followed by Tukey's test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Enrichment of CSCs from clinically
resected glioblastomas through adherent culture
A total of 18 surgically resected brain tumors were
mechanically grinded followed by digestion with accutase. Single
cell mixtures were plated down onto laminin-coated surface in
DMEM/F12/N2/B27 medium supplied with bFGF and EGF. A total of 4
adherent lines were then successfully established from these 18
tumors. As indicated in Table I, all
4 tumors that were able to generate adherent cultures were primary
glioblastomas (grade IV astrocytoma). While other tumor subtypes,
including germ cell tumor (patient no. 1), ependymoma (patient no.
8), neurocytoma (patient no. 9), low-grade astrocytoma (patient
nos. 2, 5, 12, 14 and 17), and secondary brain tumor metastasized
from lung adenocarcinoma (patient no. 19), were not able to
generate adherent cultures. This suggests that the adherent culture
paradigm favors enriching CSCs from glioblastomas. A total of 4 out
of 8 glioblastomas generated adherent CSCs, and therefore the
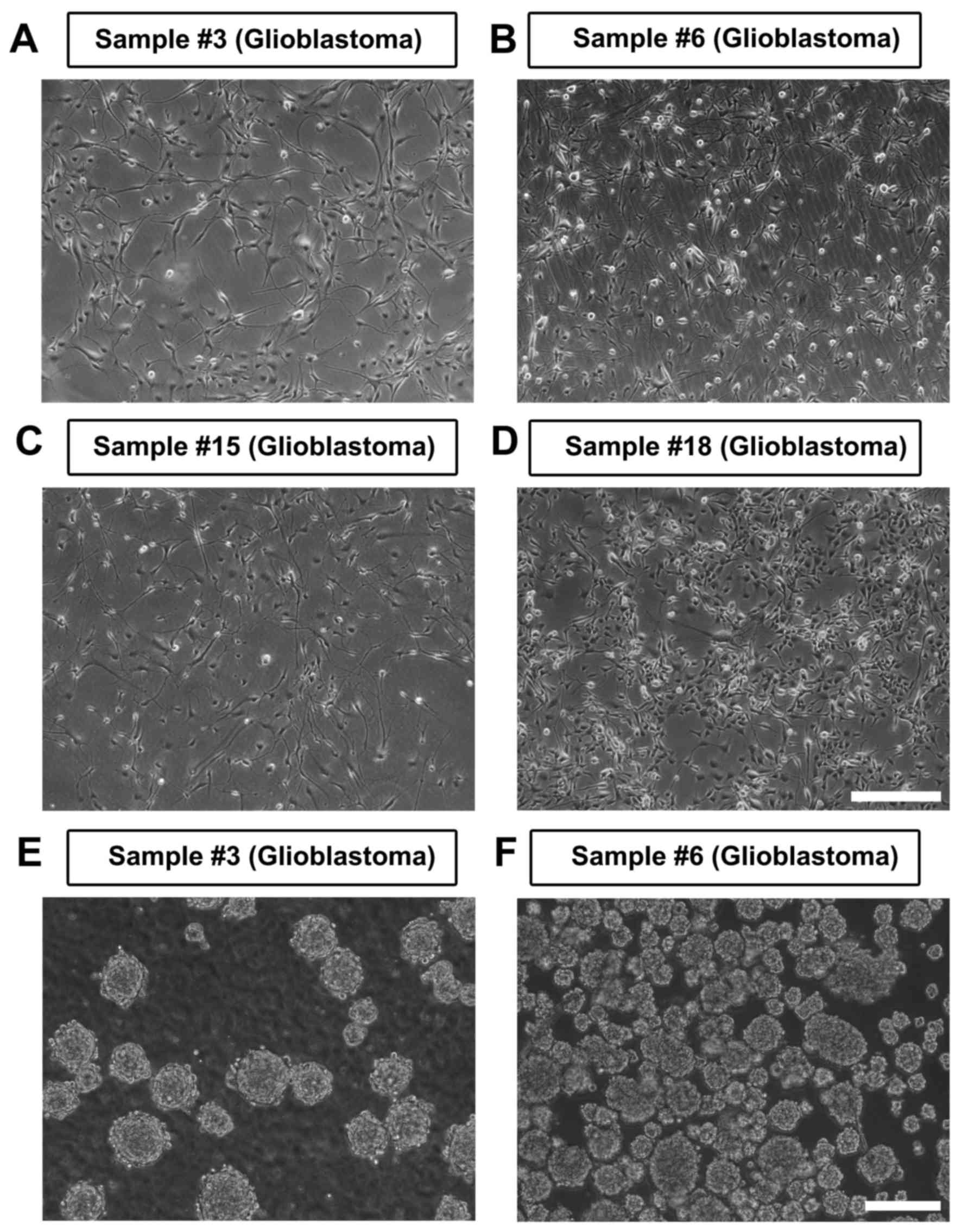
derivation efficiency was ~50% (Fig.
1A-D). The derivation efficiency may be more closely associated
with the molecular signatures of CSCs rather than the proliferation
rate of the tumor cells, since there is no difference in the
percentage of Ki67+ cells in the tumor mass between the
groups that succeeded and failed (Table
I). A total of 3 out of 4 adherently cultured CSCs were able to
be for >50 passages, while cell line 15 gradually stopped
growing and died within 3 passages. All derived cell lines
exhibited typical progenitor morphology, and tumorspheres were
easily formed from adherent cells when re-suspended in a petri dish
(Fig. 1E and F). In addition, the
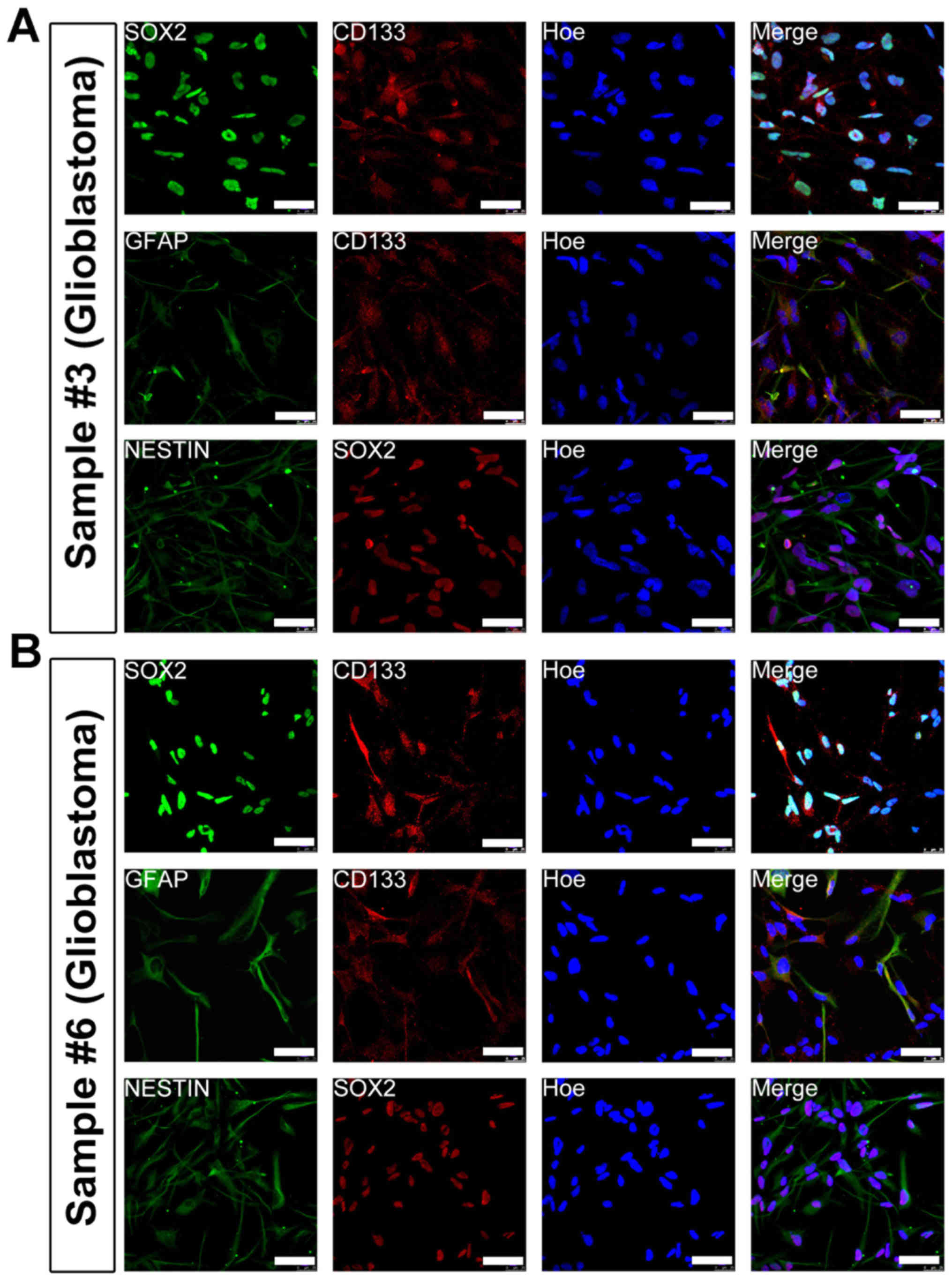
cultured cells uniformly expressed CSC markers-Nestin, Sox2, CD133
and GFAP (Fig. 2). Together, these
data indicated that the adherent culture paradigm is suitable for
enriching homogenous CSCs. In addition, this adherent culture
method is more suitable for enriching CSCs from glioblastomas and
is less efficient for other brain tumor subtypes.
Secondary brain tumor does not contain
Sox2+/CD133+ cells and fails to generate
adherent CSCs
Next, whether different types of brain tumors might
harbor various molecular signatures were examined, which may
account for the differences in the efficiency of CSC enrichment.
Nestin, Sox2, CD133 and GFAP are key progenitor markers used for
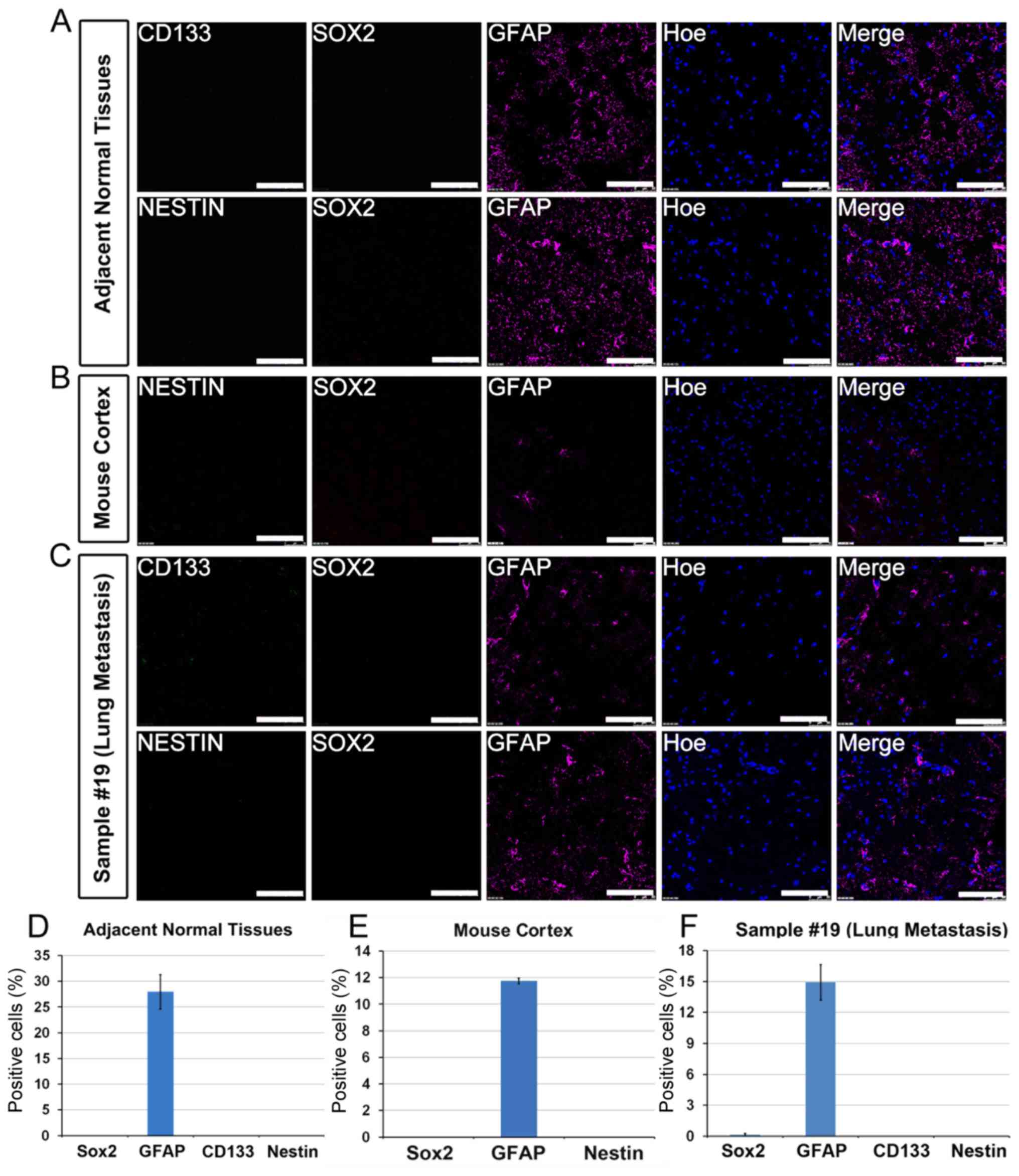
labeling brain CSCs (17). To test
the specificity of the antibodies, normal brain tissues that were
adjacent to tumors and adult mouse brain tissues were used for
immunostaining. As expected, only GFAP-positive astrocytes were
detected in normal brain tissues that were adjacent to tumors and
adult mouse brain sections. By contrast, no immunostaining were
detected for Nestin, Sox2 or CD133 (Fig.
3A, B, D and E). The tumor from patient no. 19 was a secondary
brain tumor that was metastasized from lung adenocarcinoma. The
results indicated that the metastasized tumor only had infiltrated
GFAP+ astrocytes, whereas it was negative for Nestin,
Sox2 or CD133, confirming that the metastasized tumor cell origin
was not of a neural lineage (Fig. 3C and
F). Furthermore, enriching CSCs from this tumor by using the
adherent culture paradigm failed, suggesting that the adherent
culture method may favor tumors of a neural lineage.
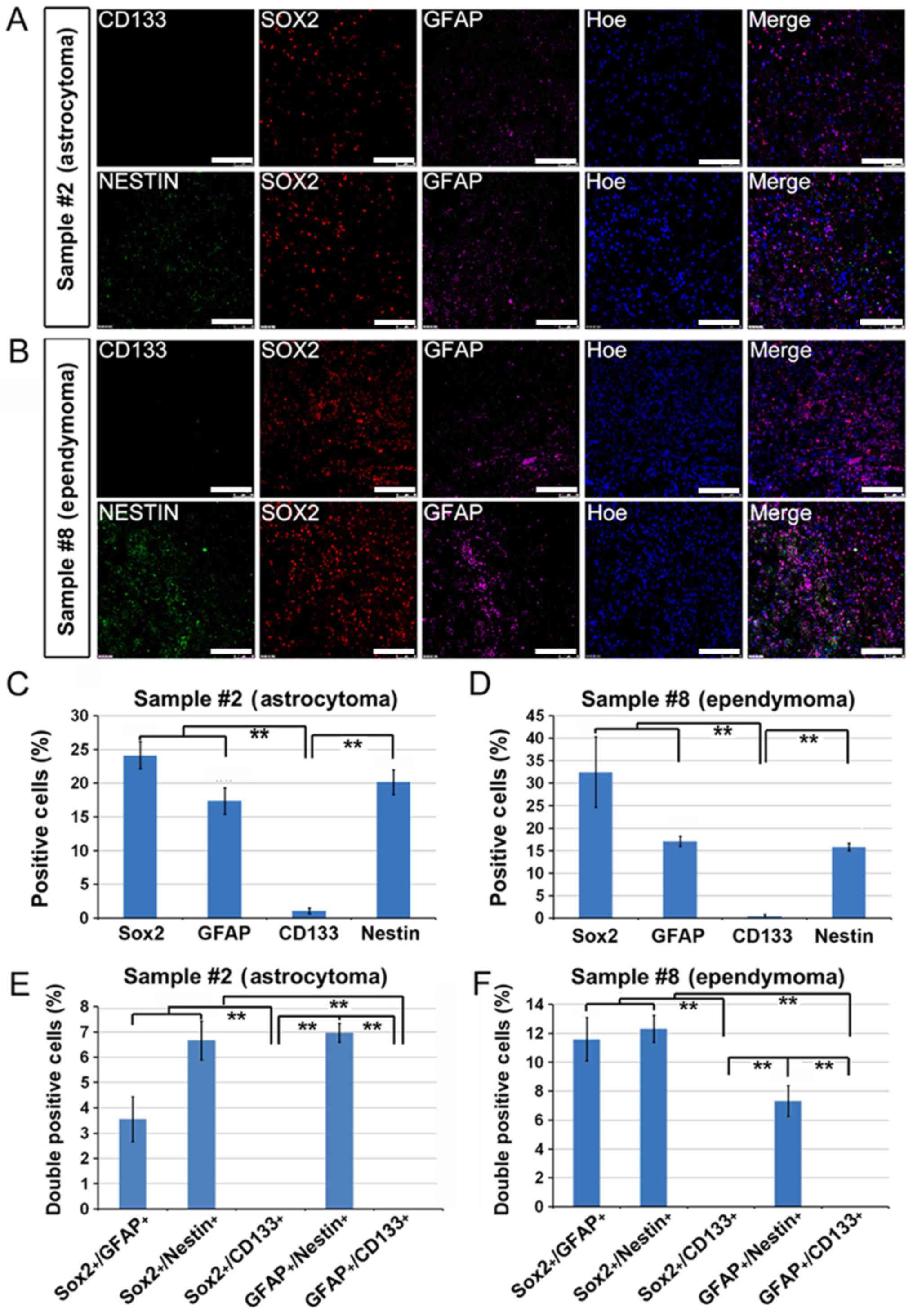
Sox2-negative brain tumors fail to
generate adherent CSCs
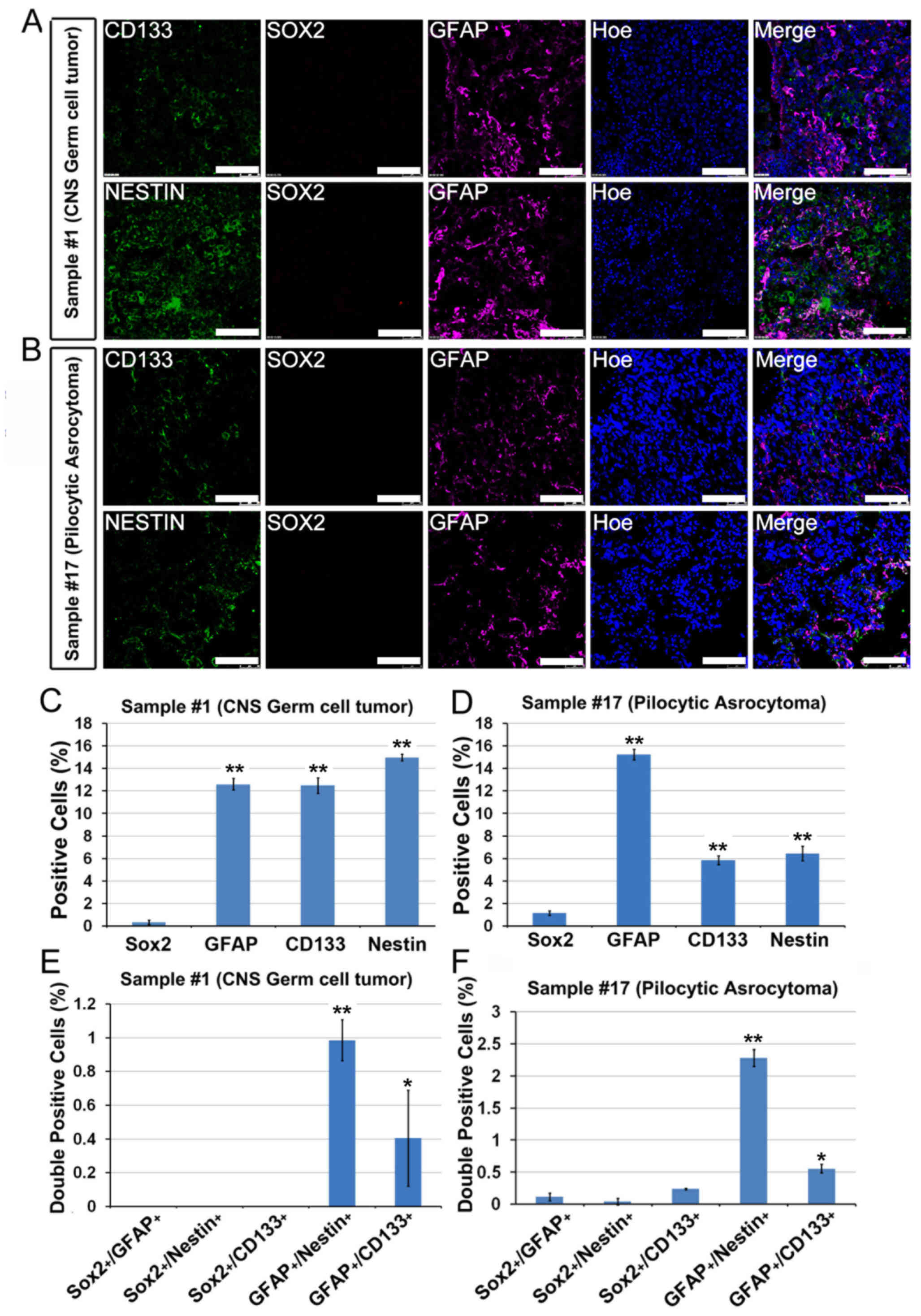
The tumors from patients with pilocytic astrocytoma
(WHO grade I, patient no. 17) and CNS germ cell tumor (patient no.
1) failed to yield adherent CSCs. The immunostaining results showed
that there was a significant number of GFAP+,
Nestin+, CD133+,
GFAP+/Nestin+ and
GFAP+/CD133+ cells within the original biopsy
compared with Sox2+ cells, which were rarely identified
(Fig. 4A-F). These data indicated
that Sox2+ cells may be from cells that were enriched on
laminin-coated culture surface and neither
GFAP+/Nestin+/Sox2− nor
GFAP+/CD133+/Sox2− cells were able
to be cultured under the adherent culture conditions.
CD133-negative brain tumors fail to
generate adherent CSCs
The tumors from patients with astrocytoma (WHO grade
II, patient no. 2) and ependymoma (WHO grade II, patient no. 8)
also failed to generate adherent CSCs. The immunostaining studies
indicated that these tumors did contain large populations of
Sox2+ cells (24-33%). Moreover, there were also a large
amount of GFAP+, Netstin+,
Sox2+/GFAP+,
Sox2+/Nestin+ and
GFAP+/Nestin+ cells within the tumor mass.
However, these tumors almost completely lacked CD133 expression
(Fig. 5A-F). It was therefore
hypothesized that CD133 expression may be another prerequisite for
enrichment of adherent CSCs.
Sox2+/CD133+
glioblastomas were able to generate adherent CSCs
Adherent CSCs were successfully enriched from
glioblastomas (WHO grade IV, patient nos. 3, 6, 15 and 18). The
immunostaining of these tumor samples indicated a high level of
Sox2 expression (30%) as well as a high level of CD133 expression
(from 10 to 14%) (Fig. 6D-F). The
percentage of GFAP+ and Nestin+ cells were
similarly detected within the tumor samples (Fig. 6D-F). The glioblastomas comprised 4-5%
Sox2+/CD133+ cells as quantified in Fig. 6G-I. Combined with aforementioned data
that Sox2− or CD133− tumors failed to enrich
CSCs, it was concluded that Sox2+/CD133+
cells may represent CSCs within the tumor biopsy, and these double
positive cells may be adherently enriched.
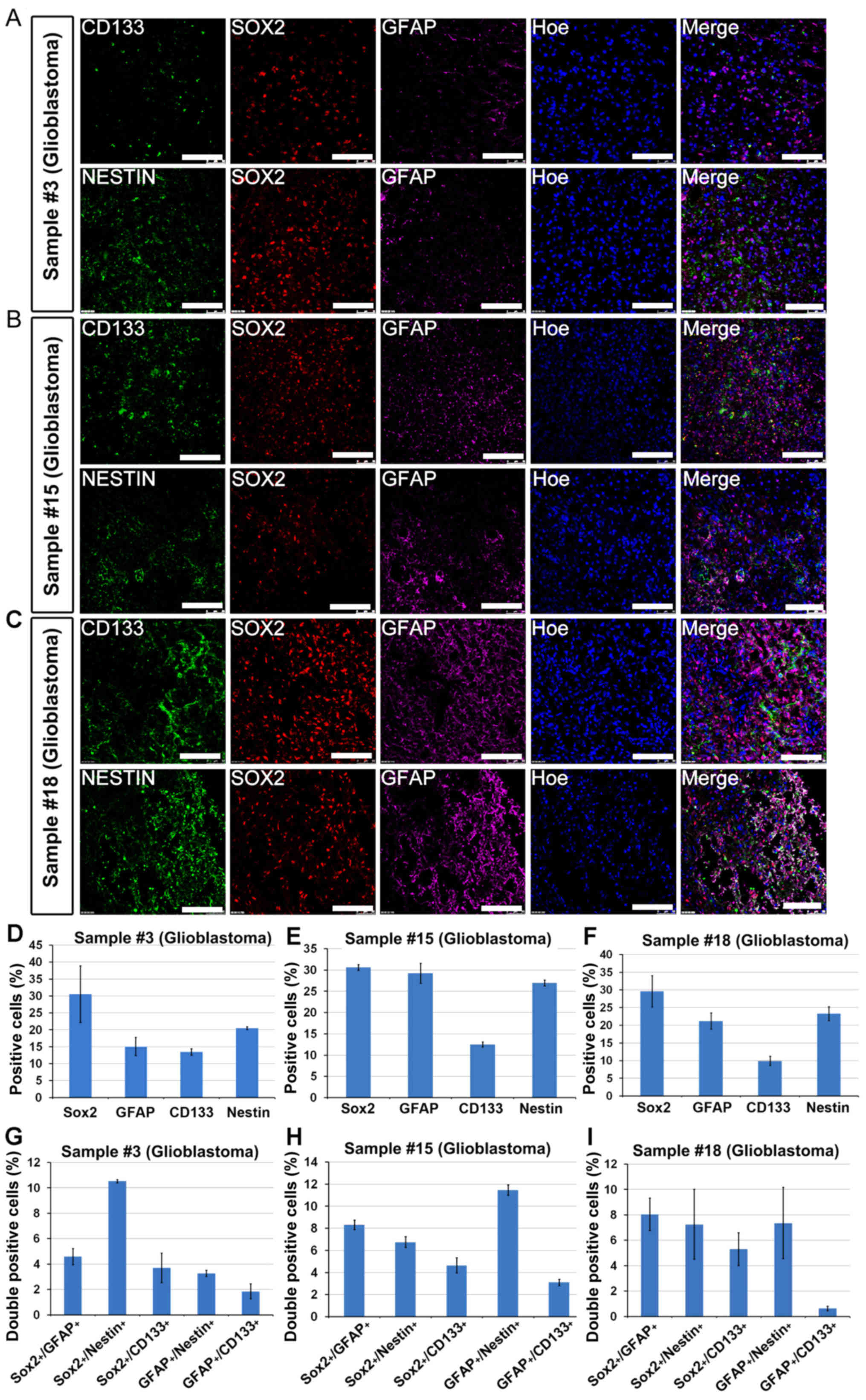 | Figure 6.Sox2+/CD133+
glioblastomas were able to generate adherent CSCs. A total of 3 WHO
grade IV glioblastomas, sample nos. (A) 3; (B) 15 and (C) 18,
successfully yielded adherent CSCs. Immunostaining indicated the
presence of Sox2+, CD133+,
Nestin+, GFAP+ and most importantly
Sox2+/CD133+ cells within the original
biopsy. Positive cells (%) in sample (D) nos. 3, (E) 15 and (F) 18.
Double positive cells (%) in sample (G) nos. 3, (H) 15 and (I) 18.
Data are presented as the mean ± standard error of the mean. Scale
bar, 100 µm. CSC, cancer stem cell; GFAP, glial fibrillary acidic
protein. |
Discussion
In the present study, the feasibility of the
adherent culture paradigm in enriching brain CSCs was analyzed. A
total of 4/9 glioblastomas were able to successfully yield CSCs
with a high purity, which was positive for GFAP, Nestin, Sox2 and
CD133. It was also revealed that Sox2+/CD133+
cells might be originated from cells that were enriched under the
adherent culture conditions.
CD133, Nestin, GFAP and Sox2 are frequently used as
key markers to characterize CSCs in human glioblastomas (17). Among these key markers, CD133 is the
most widely used for identification and enrichment of CSCs. CD133
may also be used to predict prognosis of individual glioblastoma of
patient and allow specific therapies (18–20). At
the cellular level, the knockdown of CD133 in CSCs significantly
compromised their self-renewal ability and tumorigenic potential
(21).
Sox2 is a transcription factor involved in
pluripotent stem cell or neural stem cell maintenance and lineage
reprogramming (22–24). Sox2 is also another CSC marker for
glioblastomas due in part to its association with initiating
gliomas as well as its role in multipotency and cell cycle
progression of progenitor cells (9,25,26). Silencing of Sox2 in glioblastomas
results in cell cycle arrest (9,25–29). Moreover, high level of Sox2 expression
could also serve as a reliable prognostic indication for
accelerated disease progression and poor clinical outcomes
(26,29,30).
In the present study,
Sox2+/CD133+ cells may be easily enriched
through adherent culture. There is not a single marker, which has
been shown to be sufficient to confer stem cell-like properties.
Therefore, a combination of different markers is used to identify
and isolate CSCs in glioma, including Nestin, Sox2 and GFAP
(4,30–35).
Despite CD133-positive glioma cells exhibiting CSC properties,
there is a subset of CD133-negative cells with apparent stem
cell-like characteristics (17–20). On
the other hand, Sox2 is also expressed in more differentiated
neoplastic cells within the glioblastoma (9,25–27,29). These
findings are in line with our conclusion that
Sox2+/CD133+ cells rather than those single
positive cells represent real CSCs and the cells were efficiently
enriched under adherent culture conditions in vitro.
Acknowledgements
Not applicable
Funding
The present study was supported by the National Key
Research and Development Program of China (grant no.
2018YFA0108000), the National Natural Science Foundation of China
(grant no. 31271588) and partly by the Fundamental Research Funds
for the Central Universities.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
All authors discussed the experiments and
contributed to the text of the manuscript. KL and ZC conducted most
of the experiments and wrote the draft of the manuscript. LL, QZ
and XZ conceived, discussed the project and modified the
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of the Shanghai Tenth People's Hospital (Shanghai,
China).
Patient consent for publication
All patients or parents of patients under 16
provided written informed consent for publication according to the
ethical principles of Shanghai Tenth People's Hospital.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Thurnher MM: 2007 World Health
Organization classification of tumours of the central nervous
system. Cancer Imaging. 9(Special Issue A): S1–S3. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen J, Li Y, Yu TS, McKay RM, Burns DK,
Kernie SG and Parada LF: A restricted cell population propagates
glioblastoma growth following chemotherapy. Nature. 488:522–526.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen R, Nishimura MC, Bumbaca SM,
Kharbanda S, Forrest WF, Kasman IM, Greve JM, Soriano RH, Gilmour
LL, Rivers CS, et al: A hierarchy of self-renewing tumor-initiating
cell types in glioblastoma. Cancer Cell. 17:362–375. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gillet JP, Calcagno AM, Varma S, Marino M,
Green LJ, Vora MI, Patel C, Orina JN, Eliseeva TA, Singal V, et al:
Redefining the relevance of established cancer cell lines to the
study of mechanisms of clinical anti-cancer drug resistance. Proc
Natl Acad Sci USA. 108:18708–18713. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee J, Kotliarova S, Kotliarov Y, Li A, Su
Q, Donin NM, Pastorino S, Purow BW, Christopher N, Zhang W, et al:
Tumor stem cells derived from glioblastomas cultured in bFGF and
EGF more closely mirror the phenotype and genotype of primary
tumors than do serum-cultured cell lines. Cancer Cell. 9:391–403.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Singh SK, Hawkins C, Clarke ID, Squire JA,
Bayani J, Hide T, Henkelman RM, Cusimano MD and Dirks PB:
Identification of human brain tumour initiating cells. Nature.
432:396–401. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang X, Ketova T, Litingtung Y and Chiang
C: Isolation, enrichment, and maintenance of medulloblastoma stem
cells. J Vis Exp. 2086:2010.
|
|
9
|
Favaro R, Appolloni I, Pellegatta S, Sanga
AB, Pagella P, Gambini E, Pisati F, Ottolenghi S, Foti M,
Finocchiaro G, et al: Sox2 is required to maintain cancer stem
cells in a mouse model of high-grade oligodendroglioma. Cancer Res.
74:1833–1844. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Reynolds BA and Rietze RL: Neural stem
cells and neurospheres-re-evaluating the relationship. Nat Methods.
2:333–336. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Singec I, Knoth R, Meyer RP, Maciaczyk J,
Volk B, Nikkhah G, Frotscher M and Snyder EY: Defining the actual
sensitivity and specificity of the neurosphere assay in stem cell
biology. Nat Methods. 3:801–806. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Suslov ON, Kukekov VG, Ignatova TN and
Steindler DA: Neural stem cell heterogeneity demonstrated by
molecular phenotyping of clonal neurospheres. Proc Natl Acad Sci
USA. 99:14506–14511. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Balvers RK, Kleijn A, Kloezeman JJ, French
PJ, Kremer A, van den Bent MJ, Dirven CM, Leenstra S and Lamfers
ML: Serum-free culture success of glial tumors is related to
specific molecular profiles and expression of extracellular
matrix-associated gene modules. Neuro Oncol. 15:1684–1695. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fael A, l-Mayhani TM, Ball SL, Zhao JW,
Fawcett J, Ichimura K, Collins PV and Watts C: An efficient method
for derivation and propagation of glioblastoma cell lines that
conserves the molecular profile of their original tumours. J
Neurosci Methods. 176:192–199. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pollard SM, Yoshikawa K, Clarke ID, Danovi
D, Stricker S, Russell R, Bayani J, Head R, Lee M, Bernstein M, et
al: Glioma stem cell lines expanded in adherent culture have
tumor-specific phenotypes and are suitable for chemical and genetic
screens. Cell Stem Cell. 4:568–580. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 World Health Organization
Classification of Tumors of the Central Nervous System: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ma YH, Mentlein R, Knerlich F, Kruse ML,
Mehdorn HM and Held-Feindt J: Expression of stem cell markers in
human astrocytomas of different WHO grades. J Neurooncol. 86:31–45.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pallini R, Ricci-Vitiani L, Montano N,
Mollinari C, Biffoni M, Cenci T, Pierconti F, Martini M, De Maria R
and Larocca LM: Expression of the stem cell marker CD133 in
recurrent glioblastoma and its value for prognosis. Cancer.
117:162–174. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He J, Shan Z, Li L, Liu F, Liu Z, Song M
and Zhu H: Expression of glioma stem cell marker CD133 and
O6-methylguanine-DNA methyltransferase is associated with
resistance to radiotherapy in gliomas. Oncol Rep. 26:1305–1313.
2011.PubMed/NCBI
|
|
20
|
Shin JH, Lee YS, Hong YK and Kang CS:
Correlation between the prognostic value and the expression of the
stem cell marker CD133 and isocitrate dehydrogenase1 in
glioblastomas. J Neurooncol. 115:333–341. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brescia P, Ortensi B, Fornasari L, Levi D,
Broggi G and Pelicci G: CD133 is essential for glioblastoma stem
cell maintenance. Stem Cells. 31:857–869. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maucksch C, Jones KS and Connor B: Concise
review: The involvement of SOX2 in direct reprogramming of induced
neural stem/precursor cells. Stem Cells Transl Med. 2:579–583.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu Z, Chi L, Fang Y, Liu L and Zhang X:
Specific expression pattern of a novel Otx2 splicing variant during
neural differentiation. Gene. 523:33–38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang X, Huang CT, Chen J, Pankratz MT, Xi
J, Li J, Yang Y, Lavaute TM, Li XJ, Ayala M, et al: Pax6 is a human
neuroectoderm cell fate determinant. Cell Stem Cell. 7:90–100.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Berezovsky AD, Poisson LM, Cherba D, Webb
CP, Transou AD, Lemke NW, Hong X, Hasselbach LA, Irtenkauf SM,
Mikkelsen T and deCarvalho AC: Sox2 promotes malignancy in
glioblastoma by regulating plasticity and astrocytic
differentiation. Neoplasia. 16:193–206.e25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ellis P, Fagan BM, Magness ST, Hutton S,
Taranova O, Hayashi S, McMahon A, Rao M and Pevny L: SOX2, a
persistent marker for multipotential neural stem cells derived from
embryonic stem cells, the embryo or the adult. Dev Neurosci.
26:148–165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Oppel F, Müller N, Schackert G, Hendruschk
S, Martin D, Geiger KD and Temme A: SOX2-RNAi attenuates S-phase
entry and induces RhoA-dependent switch to protease-independent
amoeboid migration in human glioma cells. Mol Cancer. 10:1372011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Alonso MM, Diez-Valle R, Manterola L,
Rubio A, Liu D, Cortes-Santiago N, Urquiza L, Jauregi P, Lopez de
Munain A, Sampron N, et al: Genetic and epigenetic modifications of
Sox2 contribute to the invasive phenotype of malignant gliomas.
PLoS One. 6:e267402011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fang X, Yoon JG, Li L, Yu W, Shao J, Hua
D, Zheng S, Hood L, Goodlett DR, Foltz G and Lin B: The SOX2
response program in glioblastoma multiforme: An integrated
ChIP-seq, expression microarray, and microRNA analysis. BMC
Genomics. 12:112011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cox JL, Wilder PJ, Desler M and Rizzino A:
Elevating SOX2 levels deleteriously affects the growth of
medulloblastoma and glioblastoma cells. PLoS One. 7:e440872012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Skalli O, Wilhelmsson U, Orndahl C, Fekete
B, Malmgren K, Rydenhag B and Pekny M: Astrocytoma grade IV
(glioblastoma multiforme) displays 3 subtypes with unique
expression profiles of intermediate filament proteins. Hum Pathol.
44:2081–2088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Irshad K, Mohapatra SK, Srivastava C, Garg
H, Mishra S, Dikshit B, Sarkar C, Gupta D, Chandra PS,
Chattopadhyay P, et al: A combined gene signature of hypoxia and
notch pathway in human glioblastoma and its prognostic relevance.
PLoS One. 10:e01182012015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kang TW, Choi SW, Yang SR, Shin TH, Kim
HS, Yu KR, Hong IS, Ro S, Cho JM and Kang KS: Growth arrest and
forced differentiation of human primary glioblastoma multiforme by
a novel small molecule. Sci Rep. 4:55462014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang XQ and Zhang SC: Differentiation of
neural precursors and dopaminergic neurons from human embryonic
stem cells. Methods Mol Biol. 584:355–366. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hu BY, Weick JP, Yu J, Ma LX, Zhang XQ,
Thomson JA and Zhang SC: Neural differentiation of human induced
pluripotent stem cells follows developmental principles but with
variable potency. Proc Natl Acad Sci USA. 107:4335–4340. 2010.
View Article : Google Scholar : PubMed/NCBI
|