Introduction
Esophageal cancer (EsC) is one of the least studied
and deadliest cancers worldwide because of its extremely aggressive
nature and poor survival rate (1).
Due to its aggressive nature, it ranks sixth in all
cancer-associated mortalities in China and other developing
countries in 2014 (2). While many
other types of cancer are expected to decrease in incidence over
the next 10 years by 2025, the prevalence of esophageal cancer is
expected to increase by 140% (2).
There are two primary histological types of esophageal cancer.
Adenocarcinoma is prevalent in the United States of America and
certain other developed countries, while squamous cell carcinoma is
the most common esophageal cancer worldwide, including Japan, China
and other developing counties (1,3). Treatment
primarily includes surgery, chemotherapy and radiotherapy,
depending on the stages of the disease. Despite intensive
investigation, patient prognosis has not significantly improved
over the past 20 years, with a 5-year survival rate of <20%
(1,4).
Combined preoperative chemotherapy and radiotherapy
(chemo-radiotherapy) has demonstrated certain benefits (5–8). However,
a significant proportion of patients respond poorly to chemotherapy
and/or radiotherapy (9), which have
severe side effects. Furthermore, tumor resistance and relapse
occur despite chemo-radiotherapy (10,11).
Understanding the molecular mechanisms conveying tumor resistance
is important.
Survivin is a member of the inhibitor of apoptosis
protein (IAP) family (12). Members
of IAPs bind directly with caspase 3, 7 and 9, resulting in
function inhibition and/or ubiquitination, and consequent
degradation of these caspase proteins (13,14).
Survivin was revealed to be overexpressed in multiple types of
cancer, but was absent or expressed at low levels in normal
terminally-differentiated tissues (15). High levels of survivin expression have
been detected in cisplatin-resistant thyroid cancer cell lines,
flutamide-resistant prostate cancer cells and radiation-resistant
pancreatic cancer cells (16).
Survivin is also expressed in esophageal carcinomas
(17), and a high expression level
was associated with poor prognosis in esophageal cancer patients
(18,19). Targeted down-regulation of survivin by
RNA interference (RNAi) repressed the growth of KYSE510 cells, an
esophageal squamous carcinoma (ESC) cell line (20). Survivin may bind to the Inhibitor of
nuclear factor β promoter, and enhance nuclear factor-κB
expression, maintaining the oncogenic characteristics of esophageal
cancer cells (21). Despite these
data, the role of survivin in the mechanisms of
chemo-radioresistance in different types of esophageal cancer has
been poorly defined. The present study demonstrated that
lentivirus-mediated knockdown of survivin in ESC cell lines
suppressed the tumorigenic capacity of these cells, and enhanced
the sensitivity to conventional chemotherapy and radiotherapy in
vitro.
Materials and methods
Cell lines and cell culture
Human esophagus epithelial cells (HEEC) were
purchased from the American Type Culture Collection (Manassas, VA,
USA) and cultured in Dulbecco's modified Eagle's medium (DMEM)
(Lonza Group, Ltd., Basel, Switzerland). KYSE-150, TE-1 and ECA-109
and TE-13 were obtained from the Cell Bank of Type Culture
Collection of Chinese Academy of Sciences (Shanghai, China). The 4
ESC cell lines were cultured in RPMI-1640 (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.). All cells were
incubated at 37°C with 5% CO2. The native survivin
expression in these four ESC cell lines and HEEC was evaluated
using western blotting. KYSE-150, ECA-109 and TE-1 were selected to
evacuate the efficiency of survivin short hairpin RNA (shRNA)
transfection. KYSE-150 and ECA-109 were further used in colony
formation, cell invasion, wound healing assays and their
sensitivity to chemotherapeutic drugs or radiotherapy was
assessed.
Plasmids and viruses
Forward oligo
(5′-CCGGCTGGACAGAGAAAGAGCCAAGCTCGAGCTTGGCTCTTTCTCTGTCCAGTTTTTG-3′)
and reverse oligo
(5′-AATTCAAAAACTGGACAGAGAAAGAGCCAAGCTCGAGCTTGGCTCTTTCTCTGTCCAG-3′)
DNA were annealed and ligated to EcoR I and Age I digested pLKO.1
vector (22) (a gift from Bob
Weinberg, Addgene plasmid #8453). The resulting recombinant pLKO.1
(or empty pLKO.1 as control) was co-transfected with pMD2.G and
psPAX2 (supplied as a gift from Didier Trono, Addgene plasmid
#12259 and #12260) into 293T cells (ATCC Inc., Manassas, VA, USA).
Lentivirus particles in culture supernatant were collected 24 and
48 h after transfection and filtered through a 0.45 um membrane,
which were used immediately or aliquoted and frozen at −80°C until
use. For virus transfection, half of the culture medium 2 ml
DMEM+10% FBS + Penicillin (100 U/ml)-Streptomycin (100 µg/ml) (all
Gibco; Thermo Fisher Scientific) was immediately changed to 1,000
µl Lentivirus and (1,000 µl DMEM+10% FBS+ Penicillin-Streptomycin+
2 µl Polybrene) in the 6-well plate. Prior to continuing culture in
the incubator (37°C, 5% CO2) for 4–5 h, the medium was
changed to fresh DMEM+10% FBS + Penicillin (100 U/ml)-Streptomycin
(100 µg/ml) and cultured overnight. Transfection was performed
using Entranster-R4000 regent (Engreen Biosystem New Zealand, Ltd.,
Auckland, New Zealand) according to the manufacturer's protocol.
Briefly, cells were plated in 24-well plates in 500 µl DMEM and 10%
FBS without antibiotics and allowed to grow to 90% confluency. Both
plasmid DNA (2.5 µg) and Entranster-R4000 reagent were diluted in
50 µl of serum-free Opti-MEM (Invitrogen; Thermo Fisher Scientific,
Inc.) medium separately and incubated for 5 min. Following
incubation, plasmid DNA and Entranster-R4000 reagent were mixed
gently and added to each well containing cells and medium. 6 h
later, medium was changed to (DMEM+10% FBS + PS). Cells were
incubated at 37°C for 24 h in an incubator containing 5%
CO2 at full humidity. The plate was subjected to
centrifugation (Eppendorf, Hamburg, Germany) at 1,200 × g for 30
min at room temperature. For quantitative polymerase chain reaction
(qPCR) analysis of survivin expression, another round of infection
was performed at day 5 after the first infection.
The full length of the coding region of human
survivin (accession number: NM_001168.2) was cloned to the pcDNA3.1
plasmid (Invitrogen, Thermo Fisher Scientific, Inc.). A total of 4
µg recombinant plasmid was transfected into the cultured cell lines
with 6 µl Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) for each well of 6-well plates. A blank vector
was used as the negative control.
Reverse transcription-qPCR
(RT-qPCR)
Cells subjected to survivin shRNA lentivirus or
control virus treatment for 24 h were lysed with TRIzol®
reagent (Ambion; Thermo Fisher Scientific, Inc.) and frozen at
−80°C. RNA was extracted according to the protocol of the
manufacturer. cDNA was synthesized using random six primers
according to the protocol of the reverse transcription kit (cat.
no. 240589; Tiangen Biotech, Beijing, China). qPCR was performed
using a SYBR green-based protocol (Takara Bio, Inc., Otsu, Japan;
cat. no. RR820L) (23). Primers were
designed and synthesized by Sangon Biotech. (Shanghai, China).
Relative gene expression was determined using the 2−ΔΔCq
method (23).
Western blot
Cultured cells were lysed with
radioimmunoprecipitation buffer (Solarbio, Beijing, China; cat. no.
R0030). Protein concentration was determined with a bicinchoninic
acid assay according to kit protocols provided by the manufacturer
(cat. no. 23250; Pierce; Thermo Fisher Scientific, Inc.). A total
of 50 µg total protein from each sample was separated using
SDS-PAGE (10% gel), transferred to a polyvinylidene fluoride
membrane, blocked by 5% non-fat milk at room temperature for 1 h.
The membranes were then incubated with survivin (1:3,000), β-actin
(1:5,000) or cleaved Poly (adenosine 5′-diphosphate-ribose)
polymerase (PARP1; 1:1,000) antibodies (cat. nos. ab76424, ab227387
and ab32561, respectively; all from Abcam, Cambridge, UK) overnight
at 4°C. Subsequent to washing 3 times with TBST and incubated with
peroxidase conjugated anti-rabbit secondary antibody (1:10,000;
cat. no. SAB3700928; Sigma-Aldrich; Thermo Fisher Scientific, Inc.)
at room temperature for 1 h, the bands were detected with a
chemiluminescence kit (cat. no. WBKLS0100; EMD Millipore,
Billerica, MA, USA) and exposed to X-ray film (Kodak, Rochester,
NY, USA).
Colony formation, cell invasion and
wound healing assays
A total of ~1,000 cells in the logarithmic growth
period were seeded in 6-well dishes. Giemsa staining (5% solution,
30 min at room temperature) was performed when colonies became
visible under a light microscope with ×100 magnification. A colony
was defined as consisting of at least 50 cells. For the invasion
assay, a transwell insert with 8 µm pore size (Corning
Incorporated, Corning, NY, USA) was coated with
Matrigel®. ESC cells resuspended in serum-free RPMI-1640
medium were added to the Transwell insert, and then placed into
24-well dishes containing RPMI-1640 medium with 10% FBS. Cells were
further cultured for 24 h at 37°C with 5% CO2. The
invading cells adhering to the underside of the insert were stained
with 0.05% crystal violet at room temperature for 20 min and
counted under a light microscope with ×200 magnification. Wound
healing assay was performed according to a previously published
protocol (24) with minor adjustment.
Briefly, adherent ESC cells in 35 mm dishes were scratched with 10
µl pipette tips, washed twice with PBS, and cultured (37°C, 5%
CO2) in serum-free medium at 37°C overnight. The width
of the scratch was then calculated. A terminal deoxynucleotidyl
transferase dUTP nick end labeling (TUNEL) assay was performed
according to the protocol of the manufacturer (Promega Corporation,
Madison, WI, USA).
Chemotherapeutic drugs and
radiotherapy
Cells were treated with various concentrations of
cisplatin (0.5–64 µg/ml) or paclitaxel (0.25–32 µg/ml) for 24 h or
radiation for 3 or 6 Gy. MTT assays were then performed, as
described previously (21), to
evaluate the live cells. For the TUNEL assay, cells were treated
with 1 µg/ml paclitaxel, 2 µg/ml cisplatin or 3 Gy radiation.
Statistical analysis
All results presented were obtained from three
independent experiments, with triplicate wells for the RT-qPCR, MTT
and TUNEL assays. The data are presented as the mean ± SEM. One-way
analysis of variance (ANOVA) was used to compare between groups.
The post-hoc test used following the ANOVA to perform the pairwise
comparisons was the Least Significant Difference test. Analysis was
performed with SPSS software (version 13.0; SPSS Inc., Chicago, IL,
USA) P<0.05 was considered to indicate a statistically
significant difference.
Results
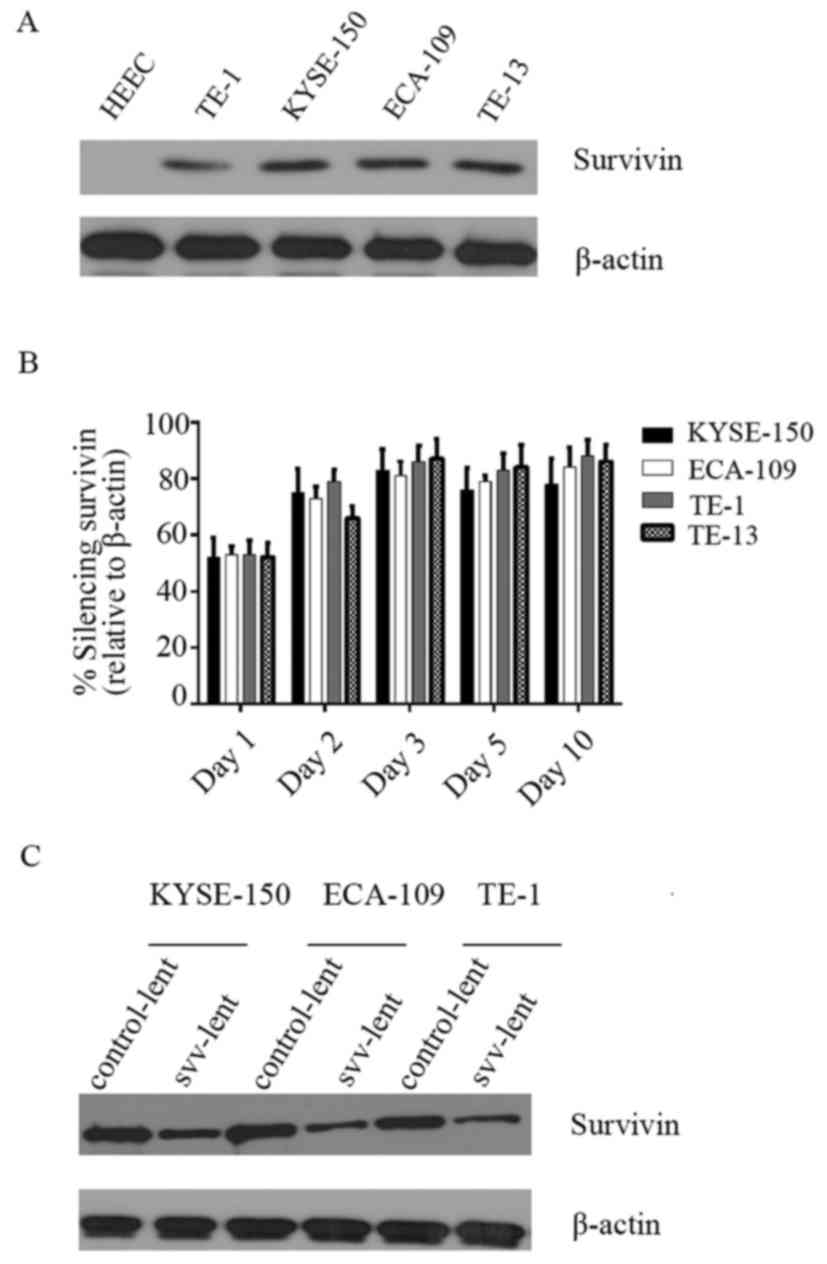
Survivin is overexpressed in ESC cell
lines and knocked down by short hairpin (sh)RNA
A panel of ESC cell lines was tested for survivin
expression. Western blot analysis indicated that survivin was not
expressed in the normal esophageal HEEC cell line, whilst highly
expressed in the 4 ESC cell lines, TE-1, TE-13, KYSE-150 and
ECA-109. No marked difference in survivin expression was observed
among these cancer cell lines (Fig.
1A). To clarify if survivin expression in these cancer cell
lines was knocked down by shRNA, oligonucleotides were cloned,
which had been previously verified for their efficiency in survivin
knockdown (25), into lentiviral
vectors, transfected it into 293T cells and the supernatant virus
(svv-lent) was harvested. The supernatant virus was used to
transduce these 4 cell lines for 1, 2, 3, 5 or 10 days. RT-qPCR was
performed to assess survivin expression. (Fig. 1B) indicates that survivin was knocked
down efficiently in all 4 cell lines, with ~50% at day 1 and up to
80% at day 3. Western blot analysis performed at day 2 following
transduction also demonstrated that svv-lent efficiently inhibited
survivin expression (Fig. 1C)
compared with the empty control lentiviral vector
(control-lent).
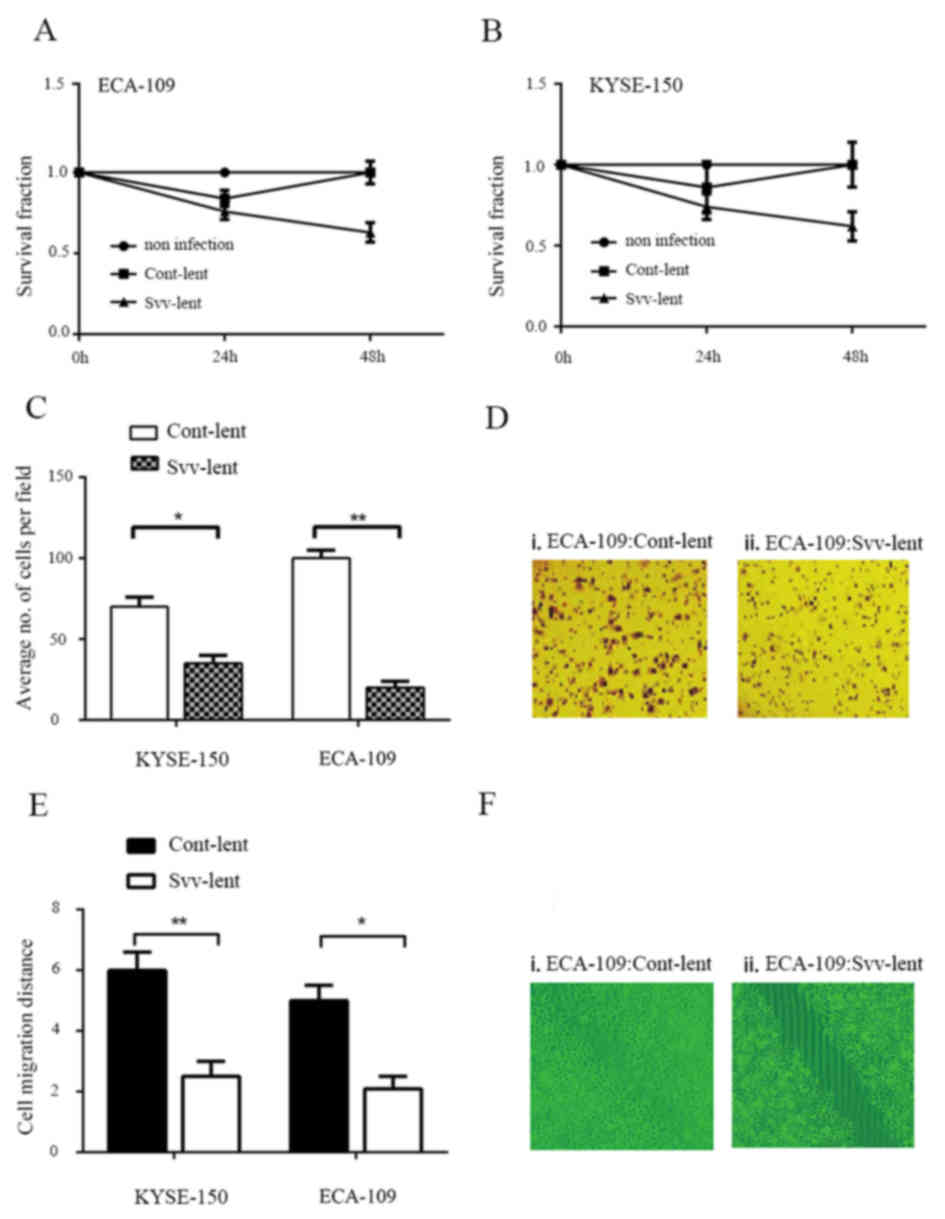
Survivin knockdown impairs the
colony-forming, migratory and invasive capabilities of ESC
cells
A clonogenic assay was performed in ECA-109 and
KYSE-150 cell lines to evaluate the role of survivin in
tumorigenicity. Svv-lent infection for 24 h significantly reduced
colony formation of the 2 cell lines, while cont-lent transduction
had no effect, compared with no infection. A long exposure time to
the RNAi virus additionally reduced the number of colonies
(Fig. 2A and B). A
Matrigel® invasion assay was also performed in KYSE-150
and ECA-109 cells. Svv-lent infection inhibited the invasive
ability of these 2 cell lines. The number of invading cells was
reduced up to 50% in KYSE-150 cells and ~75% in ECA-109 cells
(Fig. 2C and D). Consistent with the
Matrigel® invasion assay, the wound healing assay
additionally confirmed that survivin knockdown affected ESC cell
migration (Fig. 2E and F).
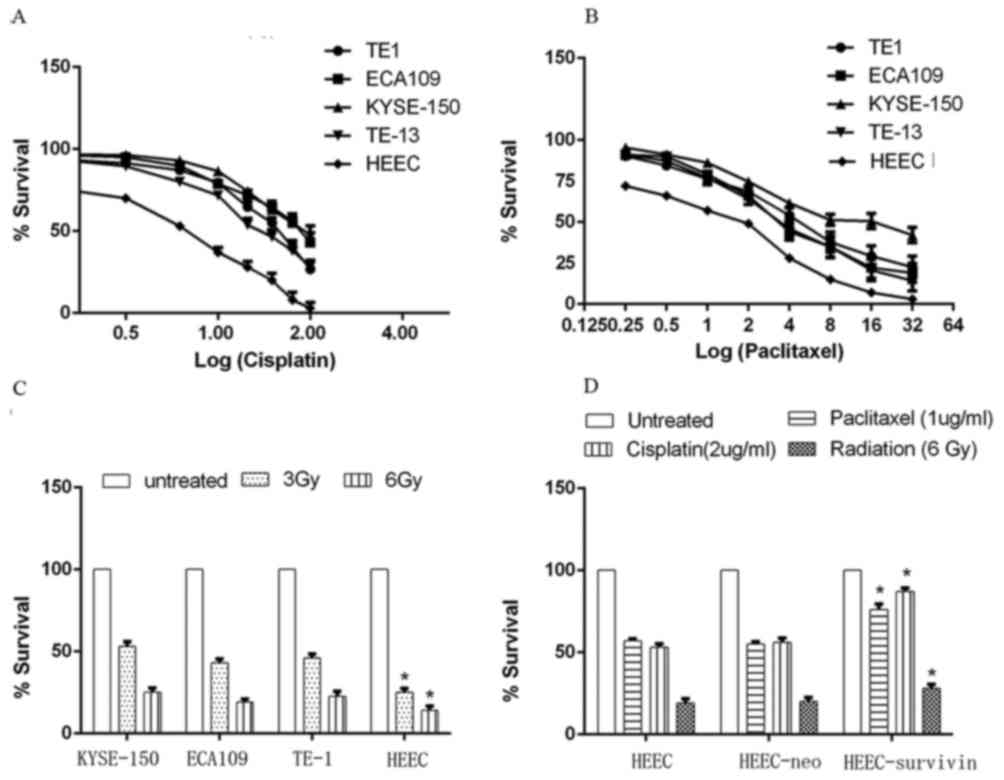
Survivin overexpression facilitates
chemo-radiotherapy resistance
The cytotoxic effects of cisplatin and paclitaxel, 2
commonly used drugs for chemotherapy in esophageal cancer, were
initially measured in ESC and HEEC cells. Cells seeded in 96-well
plates were treated with various concentrations of the drugs for 24
h; the resulting live cells were measured by an MTT assay. HEEC
cells were fairly sensitive to cisplatin and paclitaxel, with half
maximal inhibitory concentrations (IC50) of 2.6
(cisplatin) and 1.3 (paclitaxel) µg/ml. All of the 4 ESC cell
lines, however, were resistant to the 2 drugs. The IC50
of cisplatin in the TE1, ECA109, KYSE-150 and TE-13 cell lines were
18, 47.5, 54.5 and 13.7 µg/ml, respectively. The IC50 of
paclitaxel in these 4 cell lines were 4.7, 3.9, 13.5 and 3.6 µg/ml,
respectively (Fig. 3A and B).
Similarly, HEEC cells and the 4 ESC cell lines were
assessed for their sensitivity to radiotherapy. The majority of the
cells (3 Gy, ≤50%; 6 Gy, ≤75%) were killed by radiation (Fig. 3C). However, HEEC cells were more
sensitive to radiation compared with ESC cells; for example, 3 Gy
irradiation killed ~75% HEEC cells, while ~50% KYSE-150, ECA109 and
TE-1 cells survived (Fig. 3C). To
identify if the resistance exhibited by these cancer cell lines was
due to survivin expression, a survivin-pcDNA3.1 plasmid was
transfected into normal HEEC cells, and the empty pcDNA3.1 plasmid
was used as a control. A cytotoxicity test was repeated. Survivin
overexpression enhanced HEEC resistance to cisplatin and
paclitaxel, while the control vector (HEEC-neo) had no effect
(Fig. 3D).
HEEC with survivin overexpression also exhibited
elevated resistance to radiation (Fig.
3D), indicating the role of survivin in chemo- and radiotherapy
resistance of esophageal cells.
Survivin knockdown promotes ESC cells
apoptosis and chemo-radiotherapy sensitivity
KYSE-150 and ECA-109 cells infected with svv-lent or
cont-lent, or the non-infected negative controls, were treated with
low-dose paclitaxel or cisplatin. TUNEL assays were performed to
assess apoptosis in these cells. Compared to negative controls or
cont-lent virus infection, drug treatment or svv-lent transfection
significantly increased the proportion of apoptotic cells,
indicating the role of survivin in ESC cell survival. Cells
pretreated with svv-lent to knock down survivin, then exposed to
paclitaxel (Fig. 4A and B) or
cisplatin (Fig. 4C and D), exhibited
an additionally increased ratio of apoptotic cells of ~50%,
compared to drug treatment alone. Western blot analysis of cleaved
PARP1, which was cleaved by caspase 3 or other caspases into an 89
kD fragment during apoptosis, confirmed the results obtained by the
TUNEL assay (Fig. 4A and C). Drug
treatment and svv-lent transfection induced cell apoptosis, while
the combination of transfection with treatment additionally
increased the proportion of apoptotic cells. Similarly, survivin
knockdown sensitized the KYSE-150 and ECA-109 cells to radiation.
Svv-lent transfection followed by irradiation increased the
proportion of apoptotic cells (Fig. 4E
and F).
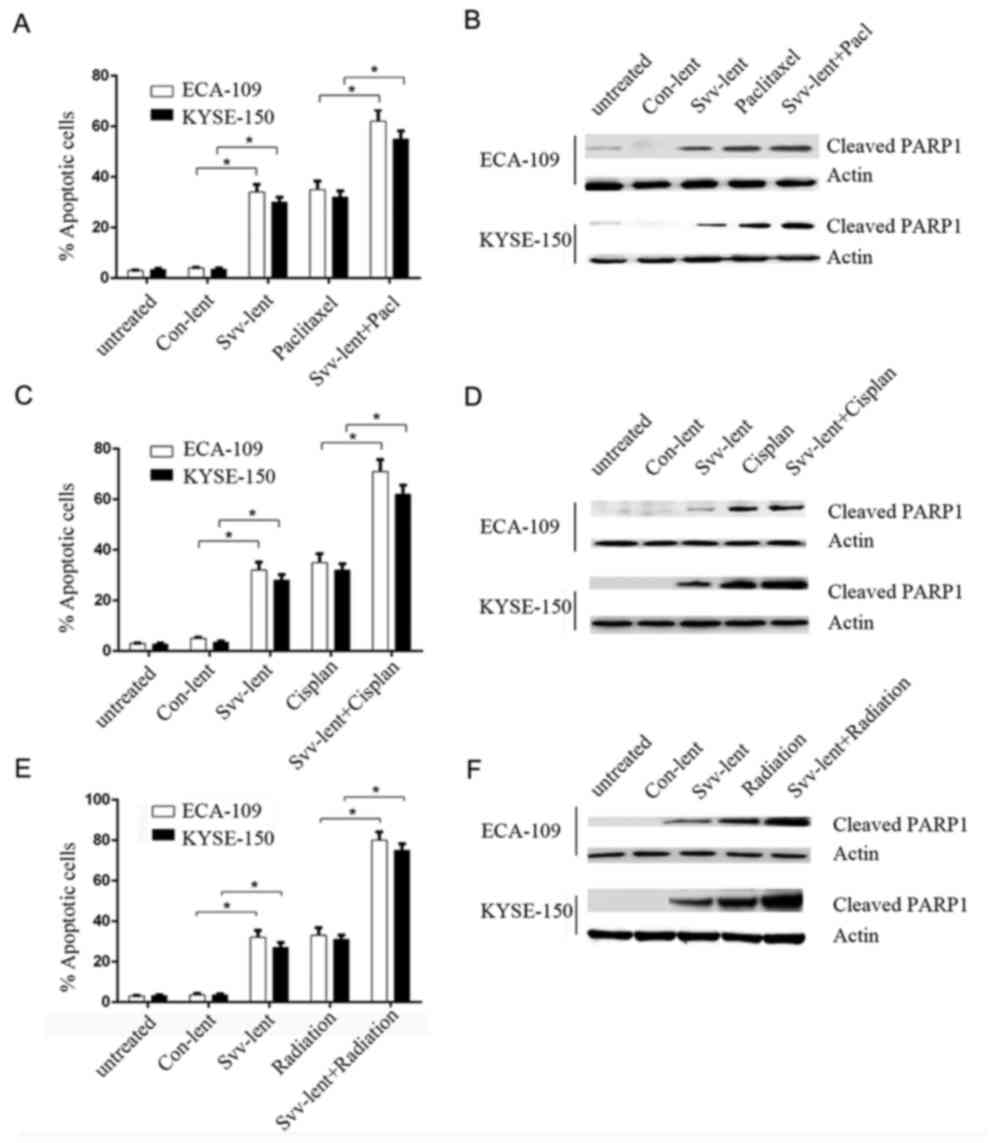 | Figure 4.Survivin knockdown promotes ESC cells
apoptosis and chemo-radiotherapy sensitivity. KYSE150 and ECA-109
cells were pretreated with survivin shRNA lentivirus or negative
control, and then a TUNEL assay was performed to evaluate the
apoptosis rate of these cells exposure to paclitaxel, cisplatin and
radiation. (A) TUNEL assays were performed to assess apoptotic
cells infected with svv-lent, cont-lent or untreated, and then
svv-lent- and cont-lent-infected cells treated with 1 µg/ml
paclitaxel. (B) Western blotting was performed to detect cleaved
PARP1 levels following paclitaxel exposure. (C) TUNEL assays were
performed to assess apoptotic cells infected with svv-lent,
cont-lent or untreated, and then svv-lent- and cont-lent-infected
cells treated with 2 µg/ml cisplatin. (D) Western blotting was
performed to detect cleaved PARP1 levels following Cisplan
exposure. (E) TUNEL assays were performed to assess apoptotic cells
infected with svv-lent, cont-lent or untreated, and then svv-lent-
and cont-lent-infected cells treated with 3 Gy radiation. (F)
Western blotting was performed to detect cleaved PARP1 levels
following radiation exposure. All experiments were performed in
triplicate and data presented as mean ± standard error of the mean.
*P<0.05 with comparisons shown by lines. svv-lent, survivin
shRNA lentivirus infected; cont-lent, infected with empty
lentivirus; Cisplan, cisplatin; svv-lent+pacl, cells pretreated
with survivin shRNA exposed to paclitaxel; PARP1, Poly (adenosine
5′-diphosphate-ribose) polymerase 1; shRNA, short hairpin RNA;
TUNEL, terminal deoxynucleotidyl transferase dUTP nick end
labeling. |
Discussion
Esophageal cancer is one of the most common types of
cancer; it is characterized by poor prognosis, high mortality and
relapse rate (2). Worldwide, the
predominant histological type is squamous cell carcinoma (1). Incidence of esophageal cancer is
considered to increase by 0.5% each year (1). The 1-year survival rate is <45%, and
the 5-year overall survival is 16.9% (1). No specific solution to this situation is
available at present (2). Survivin is
specifically expressed in cancer cells, and is not expressed in
normal differentiated tissues, with the exception of a small number
of cells including T cells and hematopoietic progenitor cells
(16). This expression pattern makes
survivin a promising target of cancer therapy. Shepherdin, which
targets heat shock protein 90, the partner and stabilizer of
survivin, inhibited breast and prostate cancer cells proliferation
in vitro and decreased the growth of tumor xenografts
(26). YM155, which inhibits survivin
transcription, was demonstrated to be effective in diminishing
non-small cell lung cancer cells tumors in xenograft models and
other cancer cells (27,28), and now is in phase II clinical studies
(www.clinicaltrials.gov; no. NCT01100931)
(29,30). In the present study, the role of
survivin in chemo-radiotherapy resistance of esophageal cancer
cells was evaluated.
The present study identified that survivin is
overexpressed in 4 ESC cell lines, but not normal esophageal HEEC
cells (Fig. 1A). This is consistent
with previous studies that revealed that survivin is overexpressed
in squamous cell esophageal cancer, but not adenocarcinoma or
Barrett's esophagus (15), and in
KYSE-150 cells (20), 1 of the 4 cell
lines investigated in the present study. Using lentivirus-mediated
RNAi, survivin was successfully knocked down (Fig. 1B and C). Survivin knockdown
significantly inhibited ESC cells colony formation, migration and
invasion (Fig. 2). Similar effects
have been identified in other tumors, for example, breast cancer
cells (31,32) and prostate and cervical cancer
(33,34). Survivin overexpression in the normal
HEEC cell line induced chemo-radioresistance (Fig. 3A-D). As determined by MTT experiments,
transient transfection of a survivin expression plasmid resulted in
more viable cells compared with the control plasmid in HEEC cells
treated with paclitaxel, cisplatin or radiation (Fig. 3D). Forced expression of survivin
conveying chemo-radioresistance has also been described previously,
and overexpression of survivin in HeLa cells resulted in resistance
to paclitaxel (16), and resistance
to temsirolimus in the renal cancer 786-O cell line (35). Overexpression of Multidrug resistance
gene and survivin in RPMI8226/VCR multiple myeloma cells conferred
multidrug resistance (36).
The results of the present study also indicated that
survivin knockdown in ESC cell lines increased their sensitivity to
chemo-radiotherapy (Fig. 4A-F). When
ESC cells were exposed to paclitaxel, cisplatin or radiation,
survivin knockdown increased the proportion of apoptotic cells, as
measured by a TUNEL assay and verified by an elevated cleaved PARP1
level. A similar role of survivin in other types of cancer has been
identified in vitro and in animal models previously
(37–40). Knockdown of survivin in head and neck
squamous cell carcinoma enhanced sensitivity to chemotherapy and
radiation (25), and overexpression
of an alternative splicing form of survivin in breast cancer cells
preserved cell viability to doxorubicin, while YM155 treatment
attenuated it (41).
To conclude, the results of the present study
suggested that survivin serves a key role in ESC carcinogenesis,
proliferation, migration and invasion. Forced expression of
survivin confers resistance to chemo-radiotherapy in normal
esophageal cells. Down-regulation of survivin suppressed tumor
growth and migration, and increased tumor sensitivity to
conventional therapies.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PX conceived of the study. CZ and LZ performed the
experiments. PX analyzed the data and all authors contributed to
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Ji'nan Central Hospital Affiliated to Shandong
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang Y: Epidemiology of esophageal
cancer. World J Gastroenterol. 19:5598–5606. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Napier KJ, Scheerer M and Misra S:
Esophageal cancer: A Review of epidemiology, pathogenesis, staging
workup and treatment modalities. World J Gastrointest Oncol.
6:112–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lin Y, Totsuka Y, He Y, Kikuchi S, Qiao Y,
Ueda J, Wei W, Inoue M and Tanaka H: Epidemiology of esophageal
cancer in Japan and China. J Epidemiol. 23:233–242. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hiripi E, Jansen L, Gondos A, Hiripi E,
Jansen L, Gondos A, Emrich K, Holleczek B, Katalinic A, Luttmann S,
et al: Survival of stomach and esophagus cancer patients in Germany
in the early 21st century. Acta Oncol. 51:906–914. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
van Hagen P, Hulshof MC, van Lanschot JJ,
Steyerberg EW, van Berge Henegouwen MI, Wijnhoven BP, Richel DJ,
Nieuwenhuijzen GA, Hospers GA, Bonenkamp JJ, et al: Preoperative
chemoradiotherapy for esophageal or junctional cancer. N Engl J
Med. 366:2074–2084. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gebski V, Burmeister B, Smithers BM, Foo
K, Zalcberg J and Simes J: Survival benefits from neoadjuvant
chemoradiotherapy or chemotherapy in oesophageal carcinoma: A
meta-analysis. Lancet Oncol. 8:226–234. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ando N, Kato H, Igaki H, Shinoda M, Ozawa
S, Shimizu H, Nakamura T, Yabusaki H, Aoyama N, Kurita A, et al: A
randomized trial comparing postoperative adjuvant chemotherapy with
cisplatin and 5-fluorouracil versus preoperative chemotherapy for
localized advanced squamous cell carcinoma of the thoracic
esophagus (JCOG9907). Ann Surg Oncol. 19:68–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sgourakis G, Gockel I, Karaliotas C,
Moehler M, Schimanski CC, Schmidberger H and Junginger T: Survival
after chemotherapy and/or radiotherapy versus self-expanding metal
stent insertion in the setting of inoperable esophageal cancer: A
case-control study. BMC Cancer. 12:702012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liao Z, Cox JD and Komaki R:
Radiochemotherapy of esophageal cancer. J Thorac Oncol. 2:553–568.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Smit JK, Faber H, Niemantsverdriet M,
Baanstra M, Bussink J, Hollema H, van Os RP, Plukker JT and Coppes
RP: Prediction of response to radiotherapy in the treatment of
esophageal cancer using stem cell markers. Radiother Oncol.
107:434–441. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hummel R, Sie C, Watson DI, Wang T, Ansar
A, Michael MZ, van der Hoek M, Haier J and Hussey DJ: MicroRNA
signatures in chemotherapy resistant esophageal cancer cell lines.
World J Gastroenterol. 20:14904–14912. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ferrario A, Rucker N, Wong S, Luna M and
Gomer CJ: Survivin, a member of the inhibitor of apoptosis family,
is induced by photodynamic therapy and is a target for improving
treatment response. Cancer Res. 67:4989–4995. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Eckelman BP, Salvesen GS and Scott FL:
Human inhibitor of apoptosis proteins: Why XIAP is the black sheep
of the family. EMBO Rep. 7:988–994. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Suzuki Y, Nakabayashi Y and Takahashi R:
Ubiquitin-protein ligase activity of X-linked inhibitor of
apoptosis protein promotes proteasomal degradation of caspase-3 and
enhances its anti-apoptotic effect in Fas-induced cell death. Proc
Natl Acad Sci USA. 98:8662–8667. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rodel F, Sprenger T, Kaina B, Liersch T,
Rödel C, Fulda S and Hehlgans S: Survivin as a
prognostic/predictive marker and molecular target in cancer
therapy. Curr Med Chem. 19:3679–3688. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mobahat M, Narendran A and Riabowol K:
Survivin as a preferential target for cancer therapy. Int J Mol
Sci. 15:2494–2516. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kato J, Kuwabara Y, Mitani M, Shinoda N,
Sato A, Toyama T, Mitsui A, Nishiwaki T, Moriyama S, Kudo J and
Fujii Y: Expression of survivin in esophageal cancer: Correlation
with the prognosis and response to chemotherapy. Int J Cancer.
95:92–95. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xia H, Chen S, Huang H and Ma H: Survivin
over-expression is correlated with a poor prognosis in esophageal
cancer patients. Clin Chim Acta. 446:82–85. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rosato A, Pivetta M, Parenti A, Iaderosa
GA, Zoso A, Milan G, Mandruzzato S, Del Bianco P, Ruol A, Zaninotto
G and Zanovello P: Survivin in esophageal cancer: An accurate
prognostic marker for squamous cell carcinoma but not
adenocarcinoma. Int J Cancer. 119:1717–1722. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Zhu H, Quan L, Zhou C, Bai J,
Zhang G, Zhan Q and Xu N: Downregulation of survivin by RNAi
inhibits the growth of esophageal carcinoma cells. Cancer Biol
Ther. 4:974–978. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zeng W, Li H, Chen Y, Lv H, Liu L, Ran J,
Sun X, Bieerkehazhi S, Liu Y, Li X, et al: Survivin activates NF-κB
p65 via the IKKβ promoter in esophageal squamous cell carcinoma.
Mol Med Rep. 13:1869–1880. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moffat J, Grueneberg DA, Yang X, Kim SY,
Kloepfer AM, Hinkle G, Piqani B, Eisenhaure TM, Luo B, Grenier JK,
et al: A lentiviral RNAi library for human and mouse genes applied
to an arrayed viral high-content screen. Cell. 124:1283–1298. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liang CC, Park AY and Guan JL: In vitro
scratch assay: A convenient and inexpensive method for analysis of
cell migration in vitro. Nat Protoc. 2:329–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Khan Z, Khan AA, Prasad GB, Khan N, Tiwari
RP and Bisen PS: Growth inhibition and chemo-radiosensitization of
head and neck squamous cell carcinoma (HNSCC) by survivin-siRNA
lentivirus. Radiother Oncol. 118:359–368. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Plescia J, Salz W, Xia F, Pennati M,
Zaffaroni N, Daidone MG, Meli M, Dohi T, Fortugno P, Nefedova Y, et
al: Rational design of shepherdin, a novel anticancer agent. Cancer
Cell. 7:457–468. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Minoda M, Kawamoto T, Ueha T, Kamata E,
Morishita M, Harada R, Toda M, Onishi Y, Hara H, Kurosaka M and
Akisue T: Antitumor effect of YM155, a novel small-molecule
survivin suppressant, via mitochondrial apoptosis in human MFH/UPS.
Int J Oncol. 47:891–899. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nakahara T, Kita A, Yamanaka K, Mori M,
Amino N, Takeuchi M, Tominaga F, Hatakeyama S, Kinoyama I,
Matsuhisa A, et al: YM155, a novel small-molecule survivin
suppressant, induces regression of established human
hormone-refractory prostate tumor xenografts. Cancer Res.
67:8014–8021. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kelly RJ, Thomas A, Rajan A, Chun G,
Lopez-Chavez A, Szabo E, Spencer S, Carter CA, Guha U, Khozin S, et
al: A phase I/II study of sepantronium bromide (YM155, survivin
suppressor) with paclitaxel and carboplatin in patients with
advanced non-small-cell lung cancer. Ann Oncol. 24:2601–2606. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Clemens MR, Gladkov OA, Gartner E,
Vladimirov V, Crown J, Steinberg J, Jie F and Keating A: Phase II,
multicenter, open-label, randomized study of YM155 plus docetaxel
as first-line treatment in patients with HER2-negative metastatic
breast cancer. Breast Cancer Res Treat. 149:171–179. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ma WH, Liu YC, Xue ML, Zheng Z and Ge YL:
Downregulation of survivin expression exerts antitumoral effects on
mouse breast cancer cells in vitro and in vivo. Oncol Lett.
11:159–167. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang H and Ye YF: Effect of survivin siRNA
on biological behaviour of breast cancer MCF7 cells. Asian Pac J
Trop Med. 8:225–228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kogo R, How C, Chaudary N, Bruce J, Shi W,
Hill RP, Zahedi P, Yip KW and Liu FF: The microRNA-218~Survivin
axis regulates migration, invasion, and lymph node metastasis in
cervical cancer. Oncotarget. 6:1090–1100. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang J, Li Z, Lin Z, Zhao B, Wang Y, Peng
R, Wang M, Lu C, Shi G and Shen Y: 17-DMCHAG, a new geldanamycin
derivative, inhibits prostate cancer cells through Hsp90 inhibition
and survivin downregulation. Cancer Lett. 362:83–96. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Carew JS, Espitia CM, Zhao W, Mita MM,
Mita AC and Nawrocki ST: Targeting survivin inhibits renal cell
carcinoma progression and enhances the activity of temsirolimus.
Mol Cancer Ther. 14:1404–1413. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tsubaki M, Takeda T, Ogawa N, Sakamoto K,
Shimaoka H, Fujita A, Itoh T, Imano M, Ishizaka T, Satou T and
Nishida S: Overexpression of survivin via activation of ERK1/2,
Akt, and NF-κB plays a central role in vincristine resistance in
multiple myeloma cells. Leuk Res. 39:445–452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shen X, Zheng JY, Shi H, Zhang Z and Wang
WZ: Survivin knockdown enhances gastric cancer cell sensitivity to
radiation and chemotherapy in vitro and in nude mice. Am J Med Sci.
344:52–58. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Morrison DJ, Hogan LE, Condos G, Bhatla T,
Germino N, Moskowitz NP, Lee L, Bhojwani D, Horton TM,
Belitskaya-Levy I, et al: Endogenous knockdown of survivin improves
chemotherapeutic response in ALL models. Leukemia. 26:271–279.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kunze D, Erdmann K, Froehner M, Wirth MP
and Fuessel S: Enhanced inhibition of bladder cancer cell growth by
simultaneous knockdown of antiapoptotic Bcl-xL and survivin in
combination with chemotherapy. Int J Mol Sci. 14:12297–12312. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cui M, Au JL, Wientjes MG, O'Donnell MA,
Loughlin KR and Lu Z: Intravenous siRNA silencing of survivin
enhances activity of mitomycin C in human bladder RT4 ×enografts. J
Urol. 194:230–237. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Faversani A, Vaira V, Moro GP, Tosi D,
Lopergolo A, Schultz DC, Rivadeneira D, Altieri DC and Bosari S:
Survivin family proteins as novel molecular determinants of
doxorubicin resistance in organotypic human breast tumors. Breast
Cancer Res. 16:R552014. View Article : Google Scholar : PubMed/NCBI
|