Introduction
Curcumin is the primary active component of turmeric
and is part of the ginger family (Zingiberaceae). Due to it having
no deleterious effects and it having a broad-spectrum anticancer
activity, curcumin has become one of the most promising anticancer
phytochemicals that has been frequently studied for its molecular
mechanism (1–4). Accumulating evidence has indicated that
curcumin may significantly inhibit the proliferation of tumor cells
and induce apoptosis (5–8). Additionally, a number of studies have
determined that curcumin may trigger the intrinsic apoptotic
pathway (9,10).
Multiple myeloma (MM) is a B-cell malignancy
characterized by overabundance of monoclonal plasma cells in the
bone marrow, which can cause osteolytic lesions (11). Previous studies demonstrated that
direct contact between osteocytes and MM cells activates the Notch
signaling pathway, increases Notch receptor expression in MM cells,
particularly Notch3 and 4, and stimulates MM proliferation
(12–14). The Notch signaling pathway is a highly
conserved system that dictates cell fate and critically influences
cell proliferation, differentiation and apoptosis (15). Notch3 is one of the four Notch
receptors (Notch1-4) that are overexpressed in numerous cancer
cells, including ovarian high-grade serous carcinoma (16), pancreatic cancer (17) and urothelial carcinoma (18). It has been reported that
overexpression of Notch3 may be responsible for promoting tumor
cell growth in human lung cancer types, including hepatocellular
carcinoma and non-small cell lung cancers (19,20). A
similar study determined that enforced expression of Notch3 could
cause dysregulated hyperplasia of T cells (21). All these data indicated that Notch3
serves a crucial role in the anti-apoptosis of cancer cells.
Components of the two families of basic helix-loop-helix
transcription factors, Hes family BHLH transcription factor 1
(Hes1) and Hes-related family transcription factor with YRPW motif
1 (Hey1), are two of the main effectors of the Notch signaling
pathway (22).
Tumor suppressor p53 is an important transcription
factor that modulates cell death and survival (23). A previous study determined that Notch3
regulates p53 at the post-transcriptional level by controlling
cyclin G1 expression and the cell signal transduction circuit in
hepatocellular carcinoma (24). There
are a number of potential mediators of p53-induced apoptosis,
including murine double minute 2 (MDM2), first apoptosis signal
receptor (Fas) and leucine repeat death domain containing protein
(LRDD) (25). p53 has been
demonstrated to function by direct interactions with, or activation
of, B-cell lymphoma-2 (Bcl-2) family proteins in the mitochondria
(26). The members of the Bcl-2
family regulate apoptosis by the permeabilization of the outer
mitochondrial membrane (27).
Notch3 and p53 signaling pathways are important in
cell fate (15,23). Curcumin is known to activate cell
death signals and induce apoptosis in cancer cells by regulating
multiple important cellular signaling pathways, including Notch
(28); however, the effects of
curcumin on Notch3 are less well studied. Therefore, the present
study focused on the effect of curcumin on mouse myeloma cells and
the underlying mechanisms mediated through the Notch3-p53 signaling
pathway.
Materials and methods
Cell culture
Mouse myeloma P3X63Ag8 cells (Stem Cell Bank,
Chinese Academy of Sciences, Shanghai, China) were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum (FBS, Hyclone; GE Healthcare Life Sciences,
Logan, UT, USA) and 1% penicillin-streptomycin (cat. no., 15140122,
Gibico, Thermo Fisher Scientific, Inc., USA) and incubated in a
humidified incubator containing 5% CO2 at 37°C. P3X63Ag8
cells were stimulated with curcumin (0, 30, 40 and 50 µM) for 24 h
at 37°C with 5% CO2.
MTT assay
The cells were placed into 96-well plates
(Eppendorf, Hamburg, Germany) at a density of 4×104/200
µl/well in DMEM (Hyclone; GE Healthcare Life Sciences) supplemented
with 10% FBS and 1% penicillin-streptomycin, and cultured overnight
at 37°C. Medium was replaced with fresh DMEM containing different
concentrations (0, 10, 20, 30, 40, 50, 60, 70 and 80 µM) of
curcumin (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Following
a further incubation for 24 h at 37°C, 20 µl MTT (2 mg/ml) was
added to each well followed by a 4 h incubation at 37°C. Viability
was determined with formazan crystal pellets dissolved in dimethyl
sulfoxide (Sigma-Aldrich; Merck KGaA), and the absorbance of the
plate was determined with a microplate reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) at 570 nm. The results are
expressed as percentage viability, calculated using the following
formula:
%Viability=100(O.D Text Item/O.D of
control),%Activity=100-%Viability.
Isolation of RNA and DNase
treatment
Total RNA was extracted using TRIzol
Reagent® (Sigma-Aldrich; Merck KGaA). All RNA extracts
were treated with DNase using a RQ1 RNase-Free DNase kit (Promega
Corporation, Madison, WI, USA), according to the manufacturer's
protocols.
Reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) assay. Synthesis of complementary DNA (cDNA) was
performed using the GoScript™ Reverse Transcription system (Promega
Corporation), according to the manufacturer's protocols.
Quantitative expression of the genes was conducted by qPCR using a
CFX96 Touch Deep Well Real-Time PCR Detection system (Bio-Rad
Laboratories, Inc.). Target genes were amplified using the primers
in Table I. β-actin was employed as
the endogenous control. The reaction mixture contained 6.25 µl 2X
GoTaq®qPCR Master mix (Promega Corporation), 1 µl cDNA,
0.25 µl upstream and 0.25 µl downstream PCR primers, and
nuclease-free water up to a final volume of 12.5 µl. Each reaction
was conducted in triplicate. The RT-qPCR reaction conditions were
subjected to an initial pre-degeneration step of 95°C for 3 min,
followed by 39 cycles of 95°C for 20 sec, followed by 60°C for 30
sec. mRNA levels were quantified using the 2−ΔΔCq method
(29) and normalized to those of
β-actin. Fluorescence data were collected and analyzed with the
Bio-Rad CFX Manager software 3.0 (Bio-Rad Laboratories, Inc.). The
melting curves were produced following qPCR.
 | Table I.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Gene symbol | NCBI Ref seq
no. | Sequence
(5′→3′) | Product length
(bp) |
|---|
| β-actin | NM_007393.4 |
| 149 |
|
Forward |
|
TGAGAGGGAAATCGTGCGTGAC |
|
|
Reverse |
|
GCTCGTTGCCAATAGTGATGACC |
|
| p53 | NM_001127233.1 |
| 179 |
|
Forward |
|
ACAGGCAGACTTTTCGCCACAG |
|
|
Reverse |
|
CCGTCCCAGAAGGTTCCCACT |
|
| Bax | NM_007527.3 |
| 118 |
|
Forward |
|
TGGAGATGAACTGGACAGCA |
|
|
Reverse |
|
GAAGTTGCCATCAGCAAACA |
|
| Bcl-2 | NM_009741.4 |
| 131 |
|
Forward |
|
CGATTGTGGCAGTCCCTTA |
|
|
Reverse |
|
CCAGGATGAAGTGCTCAGGT |
|
| Hes1 | NM_008235.2 |
| 157 |
|
Forward |
|
CAACACGACACCGGACAAAC |
|
|
Reverse |
|
GGAATGCCGGGAGCTATCTT |
|
| Hey1 | NM_010423.2 |
| 164 |
|
Forward |
|
TGCAGTTAACTCCTCCTTGCC |
|
|
Reverse |
|
CGCCGAACTCAAGTTTCCATT |
|
Western blot analysis
P3X63Ag8 cells were stimulated with curcumin (0, 30,
40 or 50 µM) for 24 h at 37°C. Subsequently, the cells treated with
indicated reagents were lysed in radioimmunoprecipitation assay
buffer (Thermo Fisher Scientific, Inc., Waltham, MA, USA) with
phenylmethylsulfonyl fluoride (Wuhan Boster Biological Technology,
Ltd., Wuhan, China) for 30 min on ice. Following centrifugation at
16,000 × g for 20 min at 4°C, the protein concentration was
determined using a Bradford Protein Assay kit (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China). Subsequently,
30 µg protein was subjected to 10% SDS-PAGE electrophoresis. The
separated proteins were transferred to a nitrocellulose filter
membrane using the Trans-Blot®SD Semi-Dry Transfer Cell
(Bio-Rad Laboratories, Inc.), according to the manufacturer's
protocols. Membranes were blocked with 5% skimmed milk powder at
4°C overnight and incubated with primary antibodies [anti-Notch3
rabbit polyclonal antibody (dilution, 1:1,000; cat. no., ab23426),
anti-Bcl-2 associated X (Bax) rabbit monoclonal antibody (dilution,
1:8,000; cat. no., ab32503) and anti-Bcl-2 mouse monoclonal
antibody (dilution, 1:500; cat. no., ab692) (all from Abcam,
Cambridge, UK); anti-P53 mouse monoclonal antibody (dilution,
1:5,000; cat. no., GTX70214; GeneTex, Inc., Irvine, CA, USA); and
anti-β-Actin mouse monoclonal antibody (dilution, 1:8,000; cat.
no., HC201; Beijing Transgen Biotech Co., Ltd., Beijing, China)] in
5% skimmed milk in TBS with Tween 20 (TBST) for 1 h at room
temperature. Following three washes with TBST for 5 min each, the
membrane was incubated with the appropriate secondary antibody
[goat anti-mouse IgG (H+L) horseradish peroxidase (HRP)-conjugated
(dilution, 1:3,000; cat. no., BA1050; Wuhan Boster Biological
Technology, Ltd., Wuhan, China) and goat anti-rabbit IgG (H+L)
HRP-conjugated (dilution, 1:3,000; cat. no., BA1054; Wuhan Boster
Biological Technology, Ltd.) secondary antibodies] at room
temperature for 1 h, followed by washing three times with TBST for
10 min each time. The membrane was developed with a Western Bright
enhanced chemiluminescent (ECL) kit (Advansta, Menlo Park, CA,
USA).
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) apoptosis assay
Cells were harvested by centrifugation at 1,000 × g
for 5 min, and washed twice with 500 µl of PBS at room temperature.
Annexin V-FITC/PI double staining kit (cat. no., KGA106; Jiangsu
KeyGen Biotech Co., Ltd., Nanjing, China), according to the
manufacturer's protocols, were employed to analyze apoptotic rate
in cells. In brief, liquots of 1×106 cells were
suspended in 500 µl binding buffer followed by the addition of 5 µl
Annexin V-FITC and 5 µl PI at room temperature for 5 min. Following
incubation in the dark at 37°C for 5 min, the cells were analyzed
using a flow cytometer BD Accuri C6 Plus (BD Accuri™; BD
Biosciences, Franklin Lakes, NJ, USA). A total of 10,000 events
were measured per sample.
Statistical analysis
Differences between groups were analyzed using
one-way analysis of variance and multiple comparison between the
groups was performed using Tukey's post hoc test. Experimental data
are expressed as the mean ± standard error of the mean of three
independent experiments. P<0.05 was considered to indicate a
statistically significant difference. Statistical analyses were
performed with SPSS version 10.0 (SPSS, Inc., Chicago, IL,
USA).
Results
The effect of curcumin on the
proliferation of P3X63Ag8 cells
To determine the effect of curcumin on cell
viability, cells were treated with different concentrations of
curcumin for 24 h, and viable cells were measured using an MTT
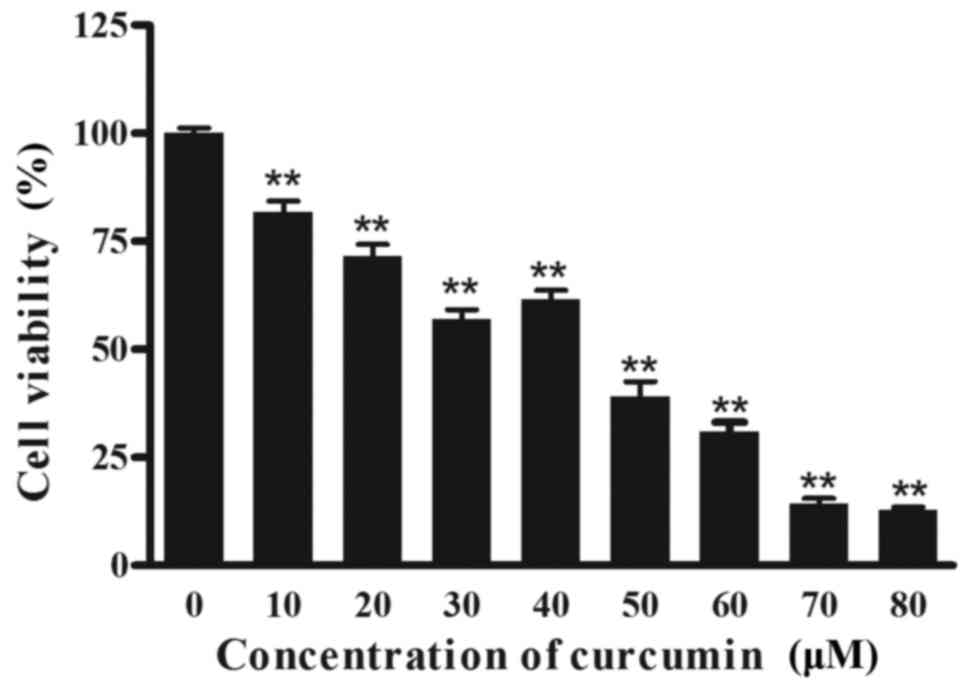
assay. As depicted in Fig. 1,
increasing concentrations of curcumin resulted in a significant
decrease (P<0.01) in cell viability compared with the 0 µM
group. Notably, curcumin treatment resulted in a reduced inhibition
at 40 µM, compared with 30 µM. A total of 3 effective
concentrations (30, 40 and 50 µM) were selected for all further
mechanistic studies.
The effect of curcumin on the mRNA and
protein levels of P3X63Ag8 cells
To determine the underlying molecular mechanisms of
apoptosis of cells by curcumin, the mRNA levels of Notch3-p53
signaling axis genes were examined. Compared with the negative
control group, the expression of Hes1 transcripts was significantly
increased in curcumin-treated cells (Fig.
2A; P<0.01). Similarly, the expression of Hey1 mRNA copies
was significantly increased in high-dose curcumin-treated cells
compared with the negative control group (Fig. 2B; P<0.01). Furthermore, no
significant differences were determined in the expression levels of
p53 in 40 and 50 µM curcumin-treated cells, compared with the
control, whereas levels increased significantly in 30 µM
curcumin-treated cells (Fig. 2C;
P<0.05). As depicted in Fig. 2D,
Bcl-2 mRNA was expressed at significantly reduced levels in 40 and
50 µM curcumin-treated cells (P<0.01). By contrast, Bax
transcripts were significantly overexpressed in curcumin-treated
cells (Fig. 2E; P<0.01), compared
with the control.
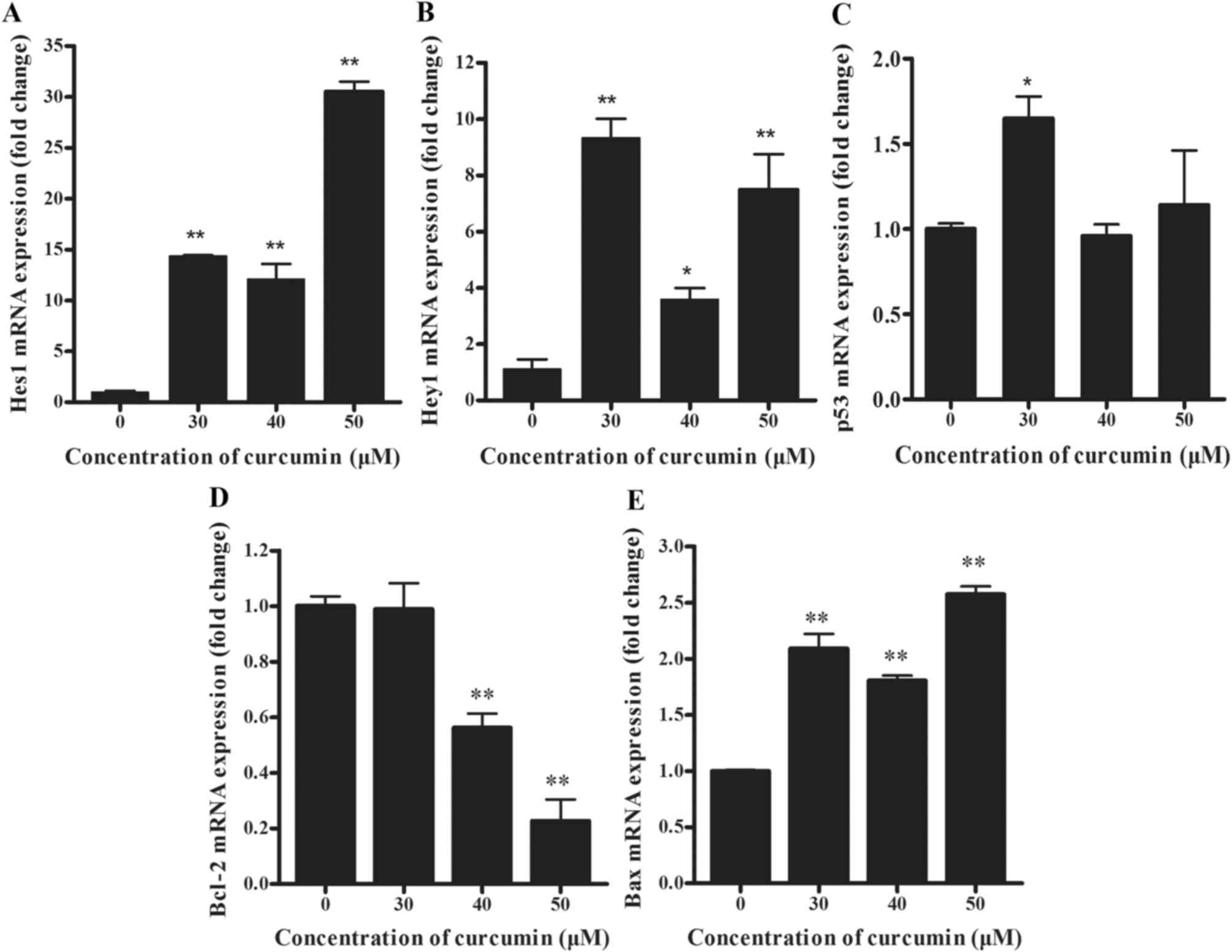 | Figure 2.(A) Hes1, (B) Hey1, (C) p53, (D)
Bcl-2 and (E) Bax mRNA levels of curcumin-treated (0, 30, 40 and 50
µM) P3X63Ag8 cells for 24 h by reverse transcription-quantitative
polymerase chain reaction. The results are expressed as the mean ±
standard error of the mean of eight independent experiments.
*P<0.05 and **P<0.01, compared with the curcumin-untreated
group. Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X; Hes1, Hes
family BHLH transcription factor 1; Hey1, Hes-related family
transcription factor with YRPW motif 1. |
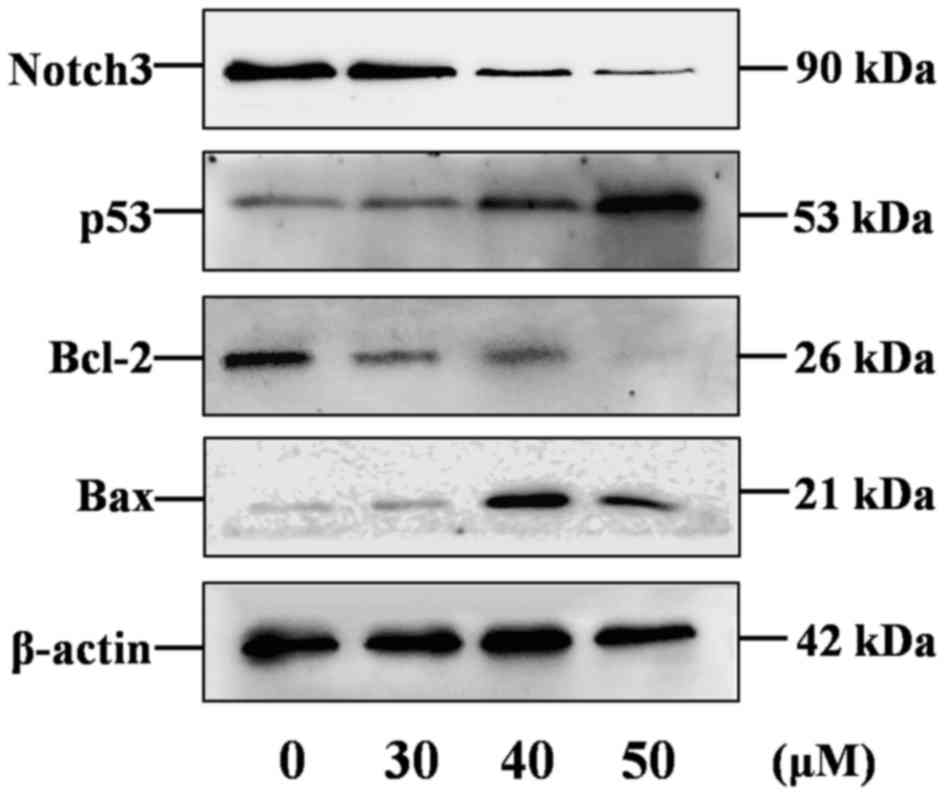
The activation status of the Notch3-p53 signaling
pathways in curcumin-stimulated P3X63Ag8 cells was investigated
using western blotting. The 90 kDa band for Notch3 was detectable
in samples (Fig. 3). As predicted,
Notch3 was overexpressed in P3X63Ag8 cells, compared with β-actin,
and significantly decreased in curcumin-treated cells. The results
demonstrated that Bcl-2 (26 kDa) and Bax (21 kDa) protein
expression levels (Fig. 3) were
similar to their mRNA levels, while the p53 (53 kDa) expression
level was not similar to its mRNA levels.
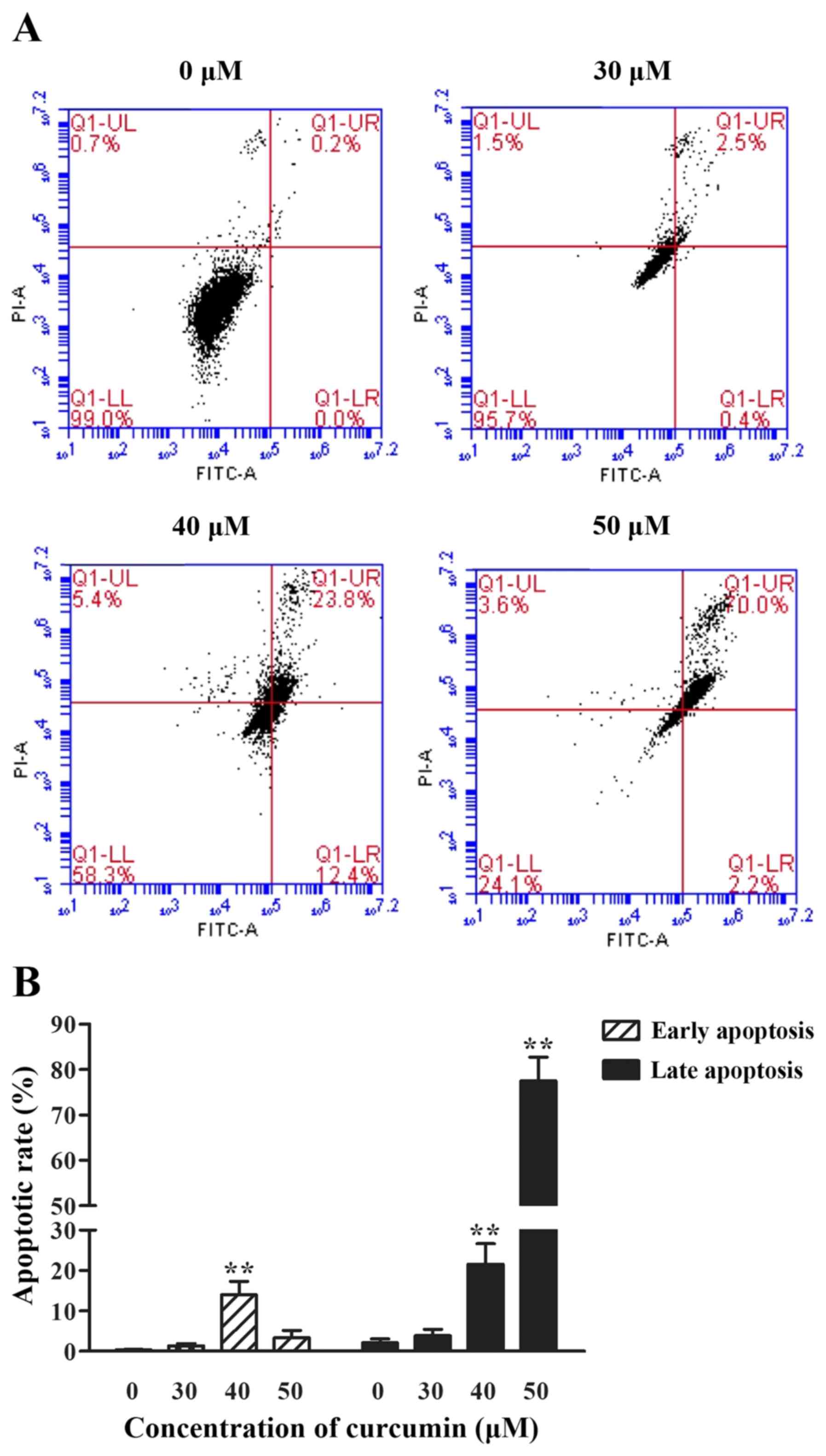
The effect of curcumin on the
apoptosis of P3X63Ag8 cells
To examine whether curcumin could cause apoptosis,
cells treated with different concentrations of curcumin (30, 40 and
50 µM) were stained with PI and FITC, and analyzed by
fluorescence-activated cell sorting analysis. Curcumin induced
apoptosis in a dose-dependent manner, with the exception in a
decrease of early apoptotic rate at 50 µM. The percentage of late
apoptosis cells was notably increased (P<0.01) compared with the
0 µM group. As depicted in Fig. 4,
the percentage of early apoptosis cells reached the highest level
when stimulated with 40 µM curcumin (P<0.01).
Discussion
Apoptosis is a multi-step, multi-pathway cell-death
program that is inherent in every cell of the body. A
characteristic of cancer cells is anti-apoptosis; therefore, cancer
treatment is primarily dependent on inducing apoptosis (30). Curcumin has been frequently used as a
spice, food additive and herbal medicine in Asia, and has not been
demonstrated to cause any toxicity (31). A number of studies indicated
anticancer effects for curcumin, and the mechanisms underlying
these effects may be the modulation of multiple oncogenic signaling
transduction elements (32–34). In the present study, it was determined
that curcumin-induced mouse myeloma P3X63Ag8 cell apoptosis is
mediated by modulation of the molecular targets p53, Bcl-2 and Bax.
It was also demonstrated, with RT-qPCR and western blotting, that
curcumin regulates the Notch3 signaling pathway. Additionally, it
was observed that curcumin dose-dependently induces apoptosis in
P3X63Ag8 cells, as indicated by Annexin V-FITC/PI staining.
Notch3 overexpression has been observed in a number
of tumor types, including T-cell acute lymphoblastic leukemia
(35), triple-negative breast cancer
(36) and prostate cancer (37). Notch3 activation favors cell
proliferation, resistance to apoptotic stimuli, metastatic
capability and maintenance of stem cell-like features (38). The results of the present study
demonstrated that the Notch3 protein was overexpressed in mouse
myeloma P3X63Ag8 cells, as predicted. Furthermore, curcumin
treatment significantly decreased Notch3 protein expression, which
is associated with the induction of apoptosis. p53 is a tumor
suppressor and transcription factor (39). When activated, p53 induces cell cycle
arrest in G1 or G2, p53 also induces cell
death by apoptosis (39). The
association between Notch3 and p53 has been studied in
hepatocellular carcinoma (24,40). The
results of the present study demonstrated that curcumin
significantly decreased Notch3 expression, promoting cell
apoptosis, which is consistent with the anticancer effects of
sorafenib and valproic acid (41). In
the present study, the mRNA and protein expression level of p53
demonstrated a comparative trend, which indicated that p53 may
depend on the post-transcriptional regulation of curcumin. The
explanation of the post-transcriptional regulation of curcumin on
p53 has been reported. Curcumin could upregulate scaffold/matrix
attachment region 1, decreasing histone acetylation at H3K9, a
lysine residue 9 on histone H3, and H3K18, a lysine residue 18 on
histone H3, resulting in the inhibition of E6 promoter, which
interrupts degradation of E6-mediated p53 in HPV18-infected
cervical carcinoma (42). In the
present study, the protein expression level of p53 is significantly
increased in the middle- and high-dose groups, while the low-dose
group demonstrated no significant difference. We hypothesized that
the apoptosis caused by the low-dose group may not depend on p53
activation, which is different to the middle- and high-dose groups.
This may be the reason for reduced inhibition at 40 µM, compared
with 30 µM. From the flow cytometry results (Fig. 4), it was also speculated that the
middle-dose group causes an early appearance of apoptotic cells,
which is comparative to the other groups. The mechanism underlying
this remains to be further investigated.
The present study indicated that Hes1 lies
downstream of Notch3 and mediates Notch3 signaling-induced
proliferation (43). Notably,
Notch-mediated cell death has been associated with upregulation of
Hes1 expression (44). Hey1, a
well-known transcriptional target of Notch signaling, may be a
pathway of Notch3 in sustaining mature vessel structure and
vascular integrity (45).
Furthermore, the expression of Hes1 and Hey1 could induce cell
death through the Notch signaling pathway (15). The Bcl-2 family is able to promote or
inhibit apoptosis by direct action on permeabilization of the
mitochondrial outer membrane (27).
Additionally, it is well known that p53 can directly regulate the
expression of Bcl-2 and Bax (26,46,47). To
define the role of curcumin on the impact of the Notch3-p53
signaling pathway on apoptosis, the p53-responsive genes Bcl-2 and
Bax were examined at the mRNA and protein expression levels
(Figs. 2 and 3). Additionally, the Notch3-responsive gene
Hes1 and Hey1 mRNA expression levels were measured. Curcumin
inhibited the expression of Bcl-2 while promoting the expression of
Bax, Hes1 and Hey1, indicating that the apoptosis effects of
curcumin on P3X63Ag8 cells depend on the Notch3-p53 signaling
pathway.
In conclusion, the results of the present study
indicated that curcumin exhibited antitumor effects in mouse
myeloma cells by inducing apoptosis through effects on the
Notch3-p53 signaling pathway. Further studies are warranted to
investigate the involvement of the Notch3-p53 signaling pathway in
the mechanism underlying curcumin suppressing MM. Furthermore, the
effect of inducing cell apoptosis by curcumin with inhibition of
the Notch3-p53 signaling pathway will be the focus of our future
investigations.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant No. 31572484; Beijing,
China).
Availability of data and materials
All data generated or analyzed during the present
study are included in this article.
Authors' contributions
XH conceived and designed the experiments. Data
collection and experiments were performed by YZ and XL. JZ, ZX, HD
and YH analyzed the data. YZ, XL and YH contributed to the writing
of the manuscript. All authors revised and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent to participate
Not applicable.
Competing interests
The authors declare no conflict of interest.
References
|
1
|
Zhang X, Wang R, Chen G, Dejean L and Chen
QH: The Effects of curcumin-based compounds on proliferation and
cell death in cervical cancer cells. Anticancer Res. 35:5293–5298.
2015.PubMed/NCBI
|
|
2
|
Hu A, Huang JJ, Li RL, Lu ZY, Duan JL, Xu
WH, Chen XP and Fan JP: Curcumin as therapeutics for the treatment
of head and neck squamous cell carcinoma by activating SIRT1. Sci
Rep. 5:134292015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Balakrishna A and Kumar MH: Evaluation of
synergetic anticancer activity of berberine and curcumin on
different models of A549, Hep-G2, MCF-7, jurkat, and K562 cell
lines. Biomed Res Int. 2015:3546142015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ting CY, Wang HE, Yu CC, Liu HC, Liu YC
and Chiang IT: Curcumin triggers DNA damage and inhibits expression
of DNA repair proteins in human lung cancer cells. Anticancer Res.
35:3867–3873. 2015.PubMed/NCBI
|
|
5
|
Mukhopadhyay A, Banerjee S, Stafford LJ,
Xia C, Liu M and Aggarwal BB: Curcumin-induced suppression of cell
proliferation correlates with down-regulation of cyclin D1
expression and CDK4-mediated retinoblastoma protein
phosphorylation. Oncogene. 21:8852–8861. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li ZC, Zhang LM, Wang HB, Ma JX and Sun
JZ: Curcumin inhibits lung cancer progression and metastasis
through induction of FOXO1. Tumour Biol. 35:111–116. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Choudhuri T, Pal S, Agwarwal ML, Das T and
Sa G: Curcumin induces apoptosis in human breast cancer cells
through p53-dependent Bax induction. FEBS Lett. 512:334–340. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang L, Wang L, Song R, Shen Y, Sun Y, Gu
Y, Shu Y and Xu Q: Targeting sarcoplasmic/endoplasmic reticulum
Ca2+-ATPase 2 by curcumin induces ER stress-associated
apoptosis for treating human liposarcoma. Mol Cancer Ther.
10:461–471. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chang PY, Peng SF, Lee CY, Lu CC, Tsai SC,
Shieh TM, Wu TS, Tu MG, Chen MY and Yang JS: Curcumin-loaded
nanoparticles induce apoptotic cell death through regulation of the
function of MDR1 and reactive oxygen species in cisplatin-resistant
CAR human oral cancer cells. Int J Oncol. 43:1141–1150. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rashmi R, Santhosh Kumar TR and
Karunagaran D: Human colon cancer cells differ in their sensitivity
to curcumin-induced apoptosis and heat shock protects them by
inhibiting the release of apoptosis-inducing factor and caspases.
FEBS Lett. 538:19–24. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Raab MS, Podar K, Breitkreutz I,
Richardson PG and Anderson KC: Multiple myeloma. Lancet.
374:324–339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Delgado-Calle J, Anderson J, Cregor MD,
Hiasa M, Chirgwin JM, Carlesso N, Yoneda T, Mohammad KS, Plotkin
LI, Roodman GD and Bellido T: Bidirectional notch signaling and
osteocyte-derived factors in the bone marrow microenvironment
promote tumor cell proliferation and bone destruction in multiple
myeloma. Cancer Res. 76:1089–1100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Terpos E, Ntanasis-Stathopoulos I,
Gavriatopoulou M and Dimopoulos MA: Pathogenesis of bone disease in
multiple myeloma: From bench to bedside. Blood Cancer J. 8:72018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jundt F, Pröbsting KS, Anagnostopoulos I,
Muehlinghaus G, Chatterjee M, Mathas S, Bargou RC, Manz R, Stein H
and Dörken B: Jagged1-induced notch signaling drives proliferation
of multiple myeloma cells. Blood. 103:3511–3515. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
development. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park JT, Chen X, Tropè CG, Davidson B,
Shih IeM and Wang TL: Notch3 overexpression is related to the
recurrence of ovarian cancer and confers resistance to carboplatin.
Am J Pathol. 177:1087–1094. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dang T, Vo K, Washington K and Berlin J:
The role of Notch3 signaling pathway in pancreatic cancer. J Clin
Oncol. 25:21049. 2007.
|
|
18
|
Zhang H, Liu L, Liu C, Pan J, Lu G, Zhou
Z, Chen Z and Qian C: Notch3 overexpression enhances progression
and chemoresistance of urothelial carcinoma. Oncotarget.
8:34362–34373. 2017.PubMed/NCBI
|
|
19
|
Gramantieri L, Giovannini C, Lanzi A,
Chieco P, Ravaioli M, Venturi A, Grazi GL and Bolondi L: Aberrant
Notch3 and Notch4 expression in human hepatocellular carcinoma.
Liver Int. 27:997–1007. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Konishi J, Kawaguchi KS, Vo H, Haruki N,
Gonzalez A, Carbone DP and Dang TP: Gamma-secretase inhibitor
prevents Notch3 activation and reduces proliferation in human lung
cancers. Cancer Res. 67:8051–8057. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bellavia D, Campese AF, Checquolo S,
Balestri A, Biondi A, Cazzaniga G, Lendahl U, Fehling HJ, Hayday
AC, Frati L, et al: Combined expression of pTalpha and Notch3 in T
cell leukemia identifies the requirement of preTCR for
leukemogenesis. Proc Natl Acad Sci USA. 99:3788–3793. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iso T, Kedes L and Hamamori Y: HES and
HERP families: Multiple effectors of the Notch signaling pathway. J
Cell Physiol. 194:237–255. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kruiswijk F, Labuschagne CF and Vousden
KH: p53 in survival, death and metabolic health: A lifeguard with a
licence to kill. Nat Rev Mol Cell Biol. 16:393–405. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Giovannini C, Minguzzi M, Baglioni M,
Fornari F, Giannone F, Ravaioli M, Cescon M, Chieco P, Bolondi L
and Gramantieri L: Suppression of p53 by Notch3 is mediated by
Cyclin G1 and sustained by MDM2 and miR-221 axis in hepatocellular
carcinoma. Oncotarget. 5:10607–10620. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hasty P, Campisi J and Sharp ZD: Do p53
stress responses impact organismal aging? Transl Cancer Res.
5:685–691. 2016. View Article : Google Scholar
|
|
26
|
Chipuk JE, Kuwana T, Bouchier-Hayes L,
Droin NM, Newmeyer DD, Schuler M and Green DR: Direct activation of
Bax by p53 mediates mitochondrial membrane permeabilization and
apoptosis. Science. 303:1010–1014. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Czabotar PE, Lessene G, Strasser A and
Adams JM: Control of apoptosis by the BCL-2 protein family:
Implications for physiology and therapy. Nat Rev Mol Cell Biol.
15:49–63. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Subramaniam D, Ponnurangam S, Ramamoorthy
P, Standing D, Battafarano RJ, Anant S and Sharma P: Curcumin
induces cell death in esophageal cancer cells through modulating
notch signaling. PLoS One. 7:e305902012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Igney FH and Krammer PH: Death and
anti-death: Tumour resistance to apoptosis. Nat Rev Cancer.
2:277–288. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ammon HP and Wahl MA: Pharmacology of
curcuma longa. Planta Med. 57:1–7. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang L, Cheng X, Gao Y, Bao J, Guan H, Lu
R, Yu H, Xu Q and Sun Y: Induction of ROS-independent DNA damage by
curcumin leads to G2/M cell cycle arrest and apoptosis in human
papillary thyroid carcinoma BCPAP cells. Food Funct. 7:315–325.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bhullar KS, Jha A and Rupasinghe HP: Novel
carbocyclic curcumin analog CUR3d modulates genes involved in
multiple apoptosis pathways in human hepatocellular carcinoma
cells. Chem Biol Interact. 242:107–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gali-Muhtasib H, Hmadi R, Kareh M, Tohme R
and Darwiche N: Cell death mechanisms of plant-derived anticancer
drugs: Beyond apoptosis. Apoptosis. 20:1531–1562. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bellavia D, Campese AF, Alesse E, Vacca A,
Felli MP, Balestri A, Stoppacciaro A, Tiveron C, Tatangelo L,
Giovarelli M, et al: Constitutive activation of NF-kappaB and
T-cell leukemia/lymphoma in Notch3 transgenic mice. EMBO J.
19:3337–3348. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Turner N, Lambros MB, Horlings HM, Pearson
A, Sharpe R, Natrajan R, Geyer FC, van Kouwenhove M, Kreike B,
Mackay A, et al: Integrative molecular profiling of triple negative
breast cancers identifies amplicon drivers and potential
therapeutic targets. Oncogene. 29:2013–2023. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pedrosa AR, Graça JL, Carvalho S,
Peleteiro MC, Duarte A and Trindade A: Notch signaling dynamics in
the adult healthy prostate and in prostatic tumor development.
Prostate. 76:80–96. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sansone P, Storci G, Giovannini C,
Pandolfi S, Pianetti S, Taffurelli M, Santini D, Ceccarelli C,
Chieco P and Bonafé M: p66Shc/Notch-3 interplay controls
self-renewal and hypoxia survival in human stem/progenitor cells of
the mammary gland expanded in vitro as mammospheres. Stem Cells.
25:807–815. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bieging KT, Mello SS and Attardi LD:
Unravelling mechanisms of p53-mediated tumour suppression. Nat Rev
Cancer. 14:359–370. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Giovannini C, Gramantieri L, Chieco P,
Minguzzi M, Lago F, Pianetti S, Ramazzotti E, Marcu KB and Bolondi
L: Selective ablation of Notch3 in HCC enhances doxorubicin's death
promoting effect by a p53 dependent mechanism. J Hepatol.
50:969–979. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu W, Liang Q, Yang X, Yu Y, Shen X and
Sun G: Combination of sorafenib and Valproic acid synergistically
induces cell apoptosis and inhibits hepatocellular carcinoma growth
via down-regulating Notch3 and pAkt. Am J Cancer Res. 7:2503–2514.
2017.PubMed/NCBI
|
|
42
|
Chakraborty S, Das K, Saha S, Mazumdar M,
Manna A, Chakraborty S, Mukherjee S, Khan P, Adhikary A, Mohanty S,
et al: Nuclear matrix protein SMAR1 represses c-Fos-mediated HPV18
E6 transcription through alteration of chromatin histone
deacetylation. J Biol Chem. 289:29074–29085. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Song Y, Zhang Y, Jiang H, Zhu Y, Liu L,
Feng W, Yang L, Wang Y and Li M: Activation of Notch3 promotes
pulmonary arterial smooth muscle cells proliferation via
Hes1/p27Kip1 signaling pathway. FEBS Open Biol. 5:656–660. 2015.
View Article : Google Scholar
|
|
44
|
Zweidler-McKay PA, He Y, Xu L, Rodriguez
CG, Karnell FG, Carpenter AC, Aster JC, Allman D and Pear WS: Notch
signaling is a potent inducer of growth arrest and apoptosis in a
wide range of B-cell malignancies. Blood. 106:3898–3906. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zaucker A, Mercurio S, Sternheim N, Talbot
WS and Marlow FL: Notch3 is essential for oligodendrocyte
development and vascular integrity in zebrafish. Dis Model Mech.
6:1246–1259. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Haldar S, Negrini M, Monne M, Sabbioni S
and Croce CM: Down-regulation of bcl-2 by p53 in breast cancer
cells. Cancer Res. 54:2095–2097. 1994.PubMed/NCBI
|
|
47
|
Harn HJ, Ho LI, Liu CA, Liu GC, Lin FG,
Lin JJ, Chang JY and Lee WH: Down regulation of bcl-2 by p53 in
nasopharyngeal carcinoma and lack of detection of its specific
t(14;18) chromosomal translocation in fixed tissues.
Histopathology. 28:317–323. 1996. View Article : Google Scholar : PubMed/NCBI
|