Introduction
Pancreatic cancer (PC) is one of the most common
causes of cancer-associated mortality in humans worldwide (1). At present, the therapeutic efficacy of
PC treatment is far from satisfactory (2). Only 20% of patients with PC are eligible
for surgical resection (3). The
overall 5-year survival rate for patients with PC is <5%
(4). To improve the cure rate for PC,
it is necessary to develop improved therapeutic strategies that
target the molecules associated with pancreatic tumor growth.
Chemotherapy is an effective strategy for
controlling various cancer types (5,6). However,
natural plant products may provide promising novel options for the
development of effective chemotherapeutic strategies for various
cancer types (7). Proanthocyanidins
are oligomeric and polymeric end-products of the flavonoid
biosynthetic pathway and are widespread in natural products,
including fruits, vegetables, seeds, nuts, flowers and bark
(8,9).
Grape seeds are a particularly rich source of proanthocyanidins
(10). Previous studies demonstrated
the potent free radical scavenger ability of grape seed
proanthocyanidins (GSPs) and GSP extract in vitro and in
vivo (11,12). Additionally, GSPs possess antioxidant
properties and their promising anticancer effect has attracted
attention in numerous studies (13,14).
GSPs can reduce the viability of human PC cells and
induce G2/M phase cell cycle (CC) arrest in vitro, which
results in apoptosis in a dose- and time-dependent manner (15). It has been demonstrated that treatment
with GSPs inhibits the migration capacity of PC cells, which is
associated with a decrease in nuclear factor-κB (NF-κB) expression
and reversal of the epithelial-mesenchymal transition process
(16). In addition to PC cells, GSPs
have been identified to inhibit the migration and invasive
potential of melanoma (17),
non-small cell lung cancer (18) and
head and neck squamous cell carcinoma cells in in vitro cell
culture models (19).
Although numerous studies have investigated the
anticancer effect of proanthocyanidins (20), to the best of our knowledge, the
global gene expression in response to treatment and the pathway
network regulated by proanthocyanidins in cancer cells have not
been thoroughly investigated. Next-generation sequencing has
revealed a substantial amount of information regarding gene
expression at the transcriptome level and the underlying molecular
events in response to drug exposure or radiation (21). The present study used RNA-sequencing
(RNA-seq) to identify the genes associated with the effects of
treatment with GSPs in PC cells. Numerous differentially-expressed
genes (DEGs) were revealed at different treatment exposure times
and bioinformatic analysis identified that a number of DEGs were
associated with the CC, regulation of transcription, DNA
replication and DNA repair, which may explain the cytotoxicity of
GSPs in PC cells.
Materials and methods
Cell culture
The human PC cell line PANC-1 was obtained from
Procell (http://www.procell.com.cn; Wuhan,
China) and cultured in monolayers in Dulbecco's modified Eagle's
medium (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) in a humidified incubator
at 37°C and a 5% CO2 atmosphere. The GSP extract,
obtained from JF-NATURAL (Tianjin, China; cat. no. J011003),
contained monomeric (9.5%), dimeric (12.8%), trimeric (76.7%) and
oligomeric (1%) procyanidins. The 100 µg GSPs extract was dissolved
in 100 µl dimethylsulfoxide (DMSO; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) for 10 min at room temperature prior to
addition to the cell culture media. The maximum concentration of
DMSO in the media did not exceed 0.1%. PANC-1 cells were treated
with 20 µg/ml GSP for 3, 12 and 24 h at 37°C. Additionally, cells
were treated with DMSO for 3, 12 and 24 h at 37°C served as
controls.
Cell viability assay
GSP-treated PANC-1 cells were plated in 96-well cell
culture plates at 5×103 cells/well and incubated for 24
h at 37°C. Subsequently, 50 µl MTT solution (5 mg/ml;
Sigma-Aldrich; Merck KGaA) was added to each well and the cells
were incubated for a further 4 h at 37°C. Following 3 min
centrifugation at 5,500 × g at 4°C, the supernatant was removed
from each well. The colored formazan crystals produced by MTT in
each well were dissolved in 150 µl DMSO and the optical density
values were measured at 490 nm.
Flow cytometry
GSP-induced apoptosis in PC cells was determined by
flow cytometry using an Annexin V-fluorescein isothiocyanate (FITC)
Apoptosis detection kit (BD Biosciences, Franklin Lakes, NJ, USA).
Following treatment with GSPs for 48 h at 37°C, cells
(2×105) were harvested, washed twice with PBS and
incubated with Annexin V-FITC and propidium iodide for 10 min in
the dark at room temperature. The stained cells were then detected
and analyzed by a MoFLO XDP flow cytometer (Beckman Coulter, Inc.,
Brea, CA, USA) and the Cell Quest 3.3 software (BD
Biosciences).
RNA extraction and sequencing
Total RNA was extracted from the GSP- or
DMSO-treated PANC-1 cells using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). The RNA was further
purified by two 15 min phenol-chloroform (1:1) (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) at 4°C
treatments and then treated with RQ1DNase (Promega Corporation,
Madison, WI, USA) for 30 min at 37°C to remove DNA. The quality and
quantity of the purified RNA were monitored by measuring the
absorbance at 260 and 280 nm, and the A260:A280 ratio using a
Smartspec Plus Spectrophotometer (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The integrity of RNA was verified by 1.5%
agarose gel electrophoresis.
For each sample, 10 µg total RNA was used for
RNA-seq library preparation. Polyadenylated mRNAs were purified and
concentrated with oligo(dT)-conjugated magnetic beads (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol prior to directional RNA-seq library preparation. The
libraries were prepared using the purified mRNAs by the TruSeq
Stranded Total RNA LT Sample Prep Kit (Illumina, Inc., San Diego,
CA, USA), according to the the manufacturer's protocol. The Nextseq
500 system (Illumina, Inc., San Diego, CA, USA) was used to collect
data from 151-base pair pair-end sequencing from ABlife Inc.
(https://www.ablife.cc; Wuhan, China).
Bioinformatic analysis
Using TopHat 2.0 software (22), reads were successfully mapped against
the current human reference genome (GRCH38). Reads per kilobase per
million mapped reads (RPKM) was used to calculate the expression
level of genes. To measure the RPKM value and screen out the DEGs,
the edgeR 3.22 software package of Bioconductor software (23) was used to specifically analyze the
differential expression of genes using RNA-seq data. The genes in
every sample with RPKM values <0.1 were removed prior to
analysis. A fold change (FC)≥2 or ≤0.5 and P≤0.01 indicated a
statistically significant DEG.
To predict the gene function and calculate the
functional category distribution frequency, Gene Ontology (GO) and
Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis was
employed using the DAVID bioinformatics database (https://david.ncifcrf.gov) (24). Networks were constructed by
calculating the Pearson's correlation coefficient (PCC) for the
expression levels of the DEGs. Cytoscape 3.5.1 software was used to
display the co-expression network (25).
Validation of DEGs by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
To validate the RNA-seq data, RT-qPCR was performed
for selected DEGs and normalization was achieved with the human
reference gene GAPDH. The primers used are presented in Table I. The same RNA samples for RNA-seq
were used for RT-qPCR, and RNA extraction was performed in
accordance with the aforementioned protocol and materials. In each
pooled sample, l µg total RNA was reverse transcribed using a
PrimeScript™ RT Reagent kit (Takara Bio, Inc., Otsu, Japan),
according to the manufacturer's protocol. qPCR was performed with a
S1000 Thermal Cycler (Bio-Rad Laboratories, Inc.) and Bestar
SYBR® Green RT-PCR Master mix (DBI Bioscience, Shanghai,
China), according to the manufacturer's protocols. The following
thermocycling conditions were used: 95°C for 10 min, followed by 38
cycles of 95°C for 15 sec and 60°C for 1 min. PCR amplifications
were performed in triplicate for each sample, and the results were
quantified by 2−∆∆Cq method (26).
 | Table I.Genes and primers used for reverse
transcription-polymerase chain reaction. |
Table I.
Genes and primers used for reverse
transcription-polymerase chain reaction.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| ATR |
ACAGTTGGTGACAGTGCTT |
CCTTGAGTTTCAGTTGTTGAGA |
| RAD50 |
ACATCAGAAGTTGGAAGAGAAC |
AGCATCCCGAAATTGTGGTT |
| NBN |
TCTATCTGAGAATGAGCCTGTG |
GTGTCCTTGAATAACTGTTCCA |
| BLM |
CAAGAGATGTGACTGACGATGT |
ATCTTCCAGAAGGACCTACATG |
| CDK1 |
CCGCAACAGGGAAGAACAG |
AAGCCAAGATAAGCAACTCCTT |
| MDM2 |
AACGCCACAAATCTGATAGT |
GATTCACTGCTACTGCTTCTT |
| HIF1A |
CGTTCCTTCGATCAGTTGTCA |
GTAGTGGTGGCATTAGCAGTAG |
| E2F3 |
GTTCTTTGTTGTGGCTCTTGTT |
TCCTTCATATCCCTCCTTCTCA |
| GAPDH |
GGTCGGAGTCAACGGATTTG |
GGAAGATGGTGATGGGATTTC |
Statistical analysis
The data were analyzed using Microsoft Excel 2007
software (https://www.microsoft.com; Microsoft
Corporation, Redmond, WA, USA). All data are presented as the mean
± standard deviation. For comparisons between two groups,
statistically significant differences between means were identified
by paired Student's t-test. For multiple comparisons, the
significance was determined by simple one-way analysis of variance
followed by a Tukey's post-hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Online data deposition
The datasets (GSE85610) generated in the present
study are available from the National Center for Biotechnology
Information Gene Expression Omnibus database (http://www.ncbi.nlm.nih.gov/geo/).
Results
GSPs inhibited the viability of PC
cells in vitro
Using flow cytometry, the present study identified
that treatment with 0, 20, 40 or 60 µg/ml GSP significantly induced
apoptosis of the PC cell line PANC-1 in a dose-dependent manner
(Fig. 1A and B). Additionally, the
effect of treatment with GSPs on the viability of PC cells was
determined with an MTT assay. A significant dose-dependent decrease
in the viability of PC cells was revealed 24 h after treatment
(Fig. 1C) Furthermore, it was
identified that treatment with GSPs was associated with necrosis
(upper left quadrants) only in a small number of PC cells in a
dose-dependent manner (Fig. 1A).
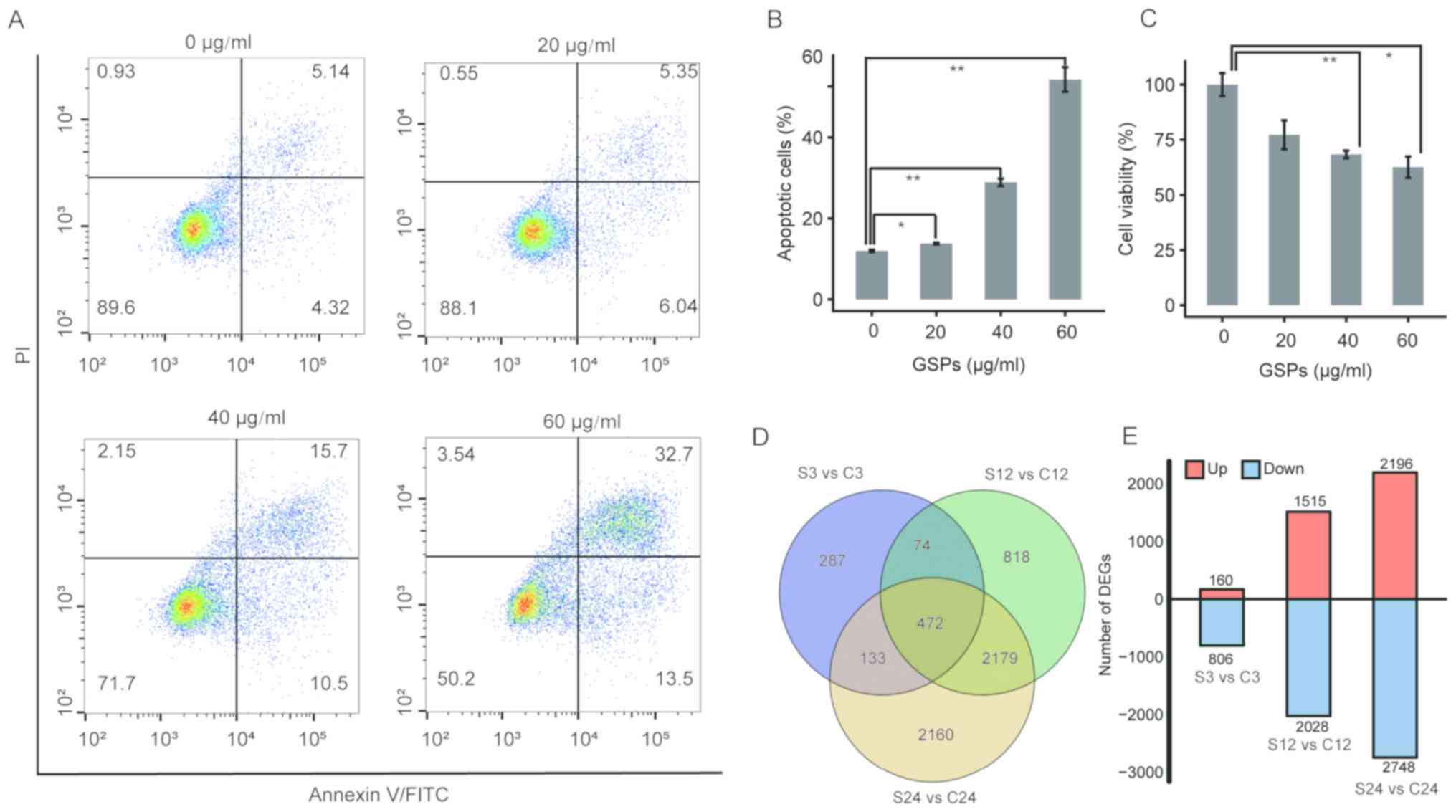 | Figure 1.Determination of DEGs in PC cells
following treatment with GSPs. (A) Treatment with GSPs induces
apoptosis of PANC-1 cells in a dose-dependent manner. Cells were
treated with various concentrations of GSPs and harvested at 12 h
post-treatment for assessment of cellular apoptosis using Annexin
V-fluorescein isothiocyanate staining coupled with flow cytometry.
The upper left, upper right and lower right quadrants represent
necrotic, late apoptotic and early apoptotic cells, respectively;
while lower left quadrant indicates normal cells. (B) Total
percentage of apoptotic PC cells in each treatment group are
summarized with data presented as the mean ± SD of two independent
experiments. (C) Treatment with GSPs reduces the viability of PC
cells. The viability of PC cells was determined with an MTT assay.
The data are reported as the percentage relative to the percentage
of control cells and presented as the mean ± SD of three
replicates. (D) Venn diagrams of the DEGs identified in the S3 vs.
C3, S12 vs. C12 and S24 vs. C24 comparisons. (E) The number of
upregulated and downregulated DEGs in the three comparisons. Data
are presented as the mean ± SD. Statistical analysis was performed
using one-way analysis of variance followed by Tukey's post hoc
test. *P<0.05 and **P<0.01. DEG, differentially expressed
gene; PC, pancreatic cancer; GSP, grape seed proanthocyanidin; SD,
standard deviation; PI, propidium iodide; PE, phycoerythrin. |
Analysis of RNA-seq data and
determination of DEGs
To investigate how GSPs regulate gene expression and
avoid high levels of necrotic cells, PANC-1 cells were treated with
20 µg/ml GSP for 3, 12 and 24 h, and the RNA samples termed S3, S12
and S24 were prepared, respectively. Cells treated with DMSO for 3,
12 and 24 h served as controls and the control RNA samples termed
C3, C12 and C24 were prepared. Two biological replicates were
generated at each time point, therefore, a total of 12 cDNA
libraries (S3-A, S3-B, C3-A, C3-B, S12-A, S12-B, C12-A, C12-B,
S24-A, S24-B, C24-A and C24-B) were constructed for RNA-seq.
Using Illumina NextSeq 500, >0.65 billion
pair-end reads were generated; corresponding to ~54.6 million
sequence reads per sample (Table
II). Using TopHat 2.0 software (22), 68.4% of all reads were successfully
mapped against the current human reference genome (GRCH38).
| Table II.Reads obtained from sequencing. |
Table II.
Reads obtained from sequencing.
| Sample | Raw reads | Clean reads | Total mapped
reads | Unique mapped
reads |
|---|
| C3-A | 52,943,434 | 44,154,519 | 39,447,474 | 36,610,491 |
| S3-A | 51,800,450 | 40,793,788 | 35,894,276 | 33,010,744 |
| C12-A | 47,942,930 | 36,813,374 | 31,477,445 | 28,953,208 |
| S12-A | 51,594,804 | 4,812,991 | 35,626,805 | 32,558,389 |
| C24-A | 52,301,536 | 41,559,257 | 36,042,733 | 33,192,336 |
| S24-A | 68130,442 | 48,397,199 | 40,704,353 | 36,919,068 |
| C3-B | 55,301,368 | 45,043,351 | 3,981,052 | 36,977,763 |
| S3-B | 59,484,624 | 46,200,745 | 40,600,169 | 37,426,268 |
| C12-B | 54,587,246 | 42,500,995 | 36,816,554 | 34,184,032 |
| S12-B | 47,518,592 | 37,483,400 | 32,481,709 | 29,620,467 |
| C24-B | 51,110,770 | 40,435,967 | 35,338,949 | 32,542,109 |
| S24-B | 62,127,244 | 4,6691,863 | 40,212,872 | 36,466,759 |
Using edgeR 22.0 software (23), a total of 966, 3,543 and 4,944
statistically significant DEGs were identified in S3 vs. C3, S12
vs. S12 and S24 vs. C24, respectively (Fig. 1D and E), which indicates marked
changes to the transcriptome in PANC-1 cells following treatment
with GSPs. Additionally, 456 common DEGs were identified between
all three comparisons.
Treatment with GSPs disrupts the CC of
PC cells
To identify the primary functions in which the DEGs
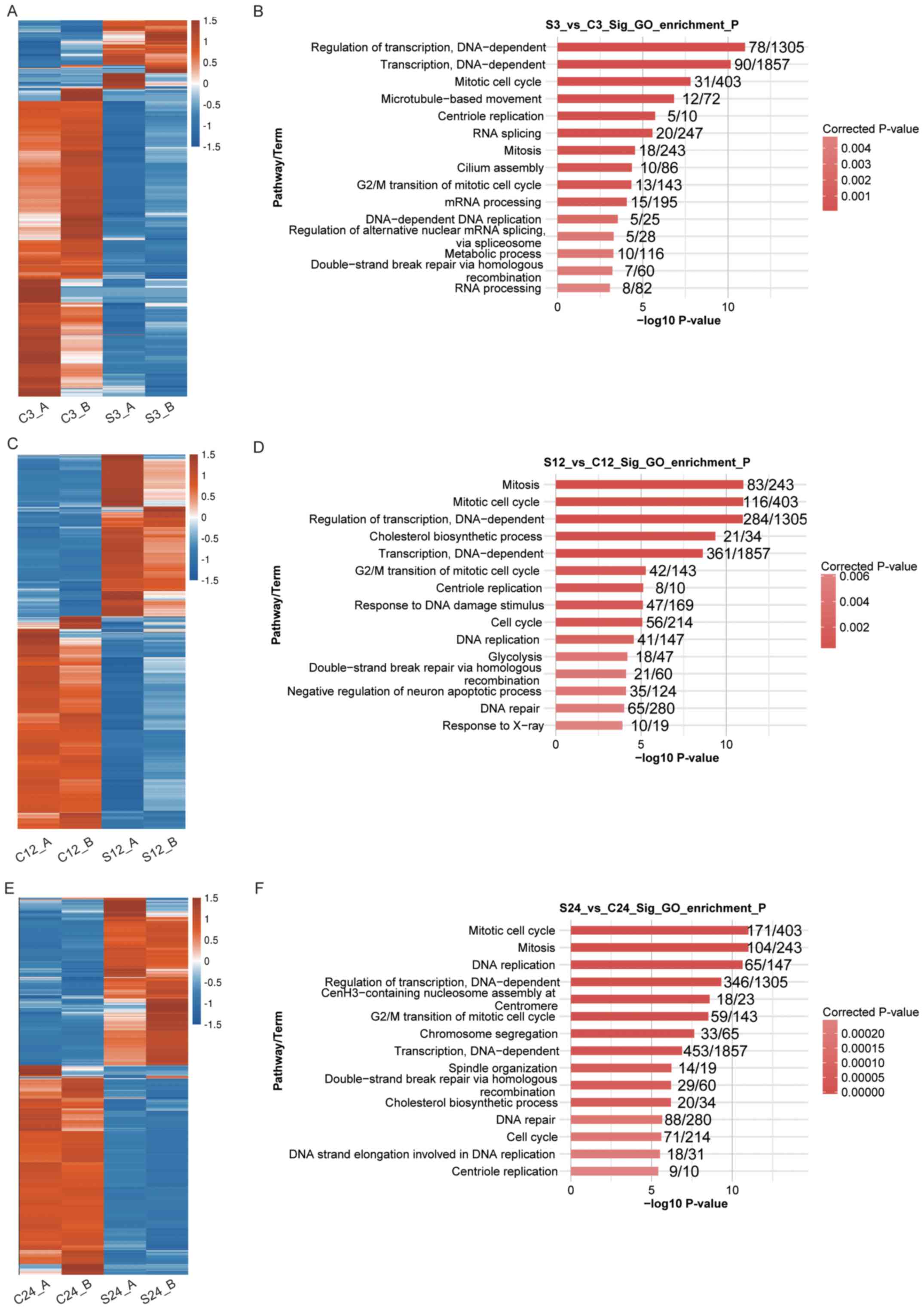
are involved, GO enrichment analysis was performed (Fig. 2). A total of 24, 52 and 64 GO terms
were identified in S3 vs. C3, S12 vs. C12 and S24 vs. C24,
respectively (P<0.05; data not shown).
For all three comparisons, numerous DEGs were
enriched in the following terms: ‘Mitotic cell cycle’ (GO:0000278),
‘G2/M transition of mitotic cell cycle’ (GO:0000086), ‘mitosis’
(GO:0007067) and ‘cell cycle’ (GO:0007049) (Fig. 2B, D and F). This indicates that
treatment with GSPs exhibits an effect on the CC of PANC-1 cells.
To determine how GSPs affect the CC of PANC-1 cells, the expression
levels of DEGs enriched in the aforementioned GO terms were
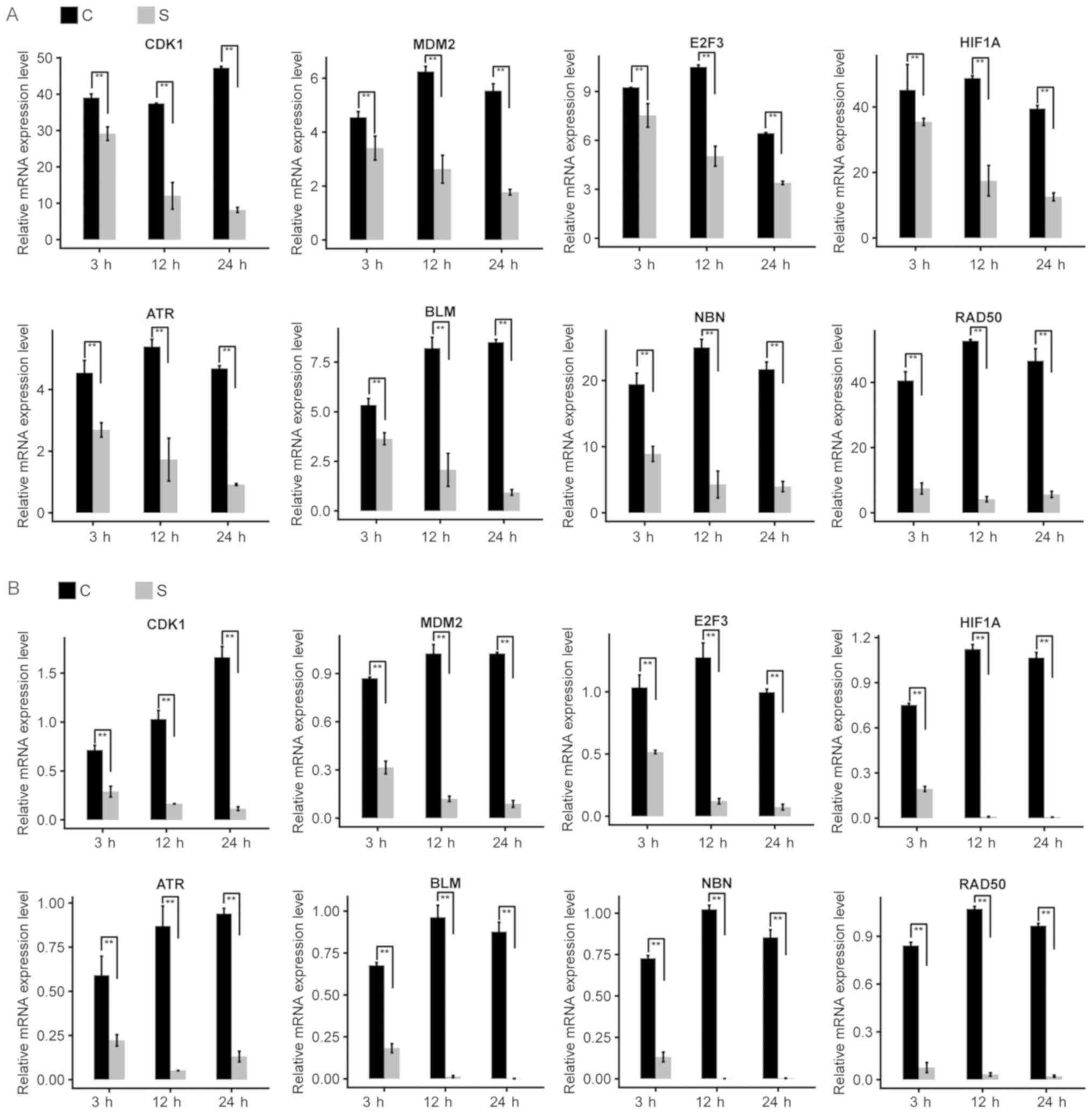
analyzed by RNA-seq. This demonstrated that the expression levels
of a number of DEGs associated with the CC, including cyclin
dependent kinase 1 (CDK1) and mouse double minute 2 homolog (MDM2),
decreased in GSP-treated cells, compared with the control cells
(Fig. 3). CDK1 serves a key role in
the eukaryotic CC by regulating the centrosome cycle and the onset
of mitosis, and modulating the G2-M transition, G1 progression and
G1-S transition via an association with multiple interphase
cyclins, including CDC7, CDC20 and CDC25A (27). MDM2 can inhibit CC arrest and
apoptosis by binding to transcriptional activation domain of TP53
(28).
The results of RT-qPCR were consistent with the
results of RNA-seq (Fig. 3B). PCC
values were >0.9, which indicates that treatment with GSPs may
disrupt the CC of PANC-1 cells. Additionally, certain genes
associated with the CC were identified to be upregulated in cells
treated with GSPs, compared with control cells. For example, the
mitogen-activated protein kinase 3 (MAPK3) gene was revealed as an
upregulated DEG in S12 vs. C12 and S24 vs. C24 (data not shown).
MAPK3 serves a role in a signaling cascade that mediates various
cellular processes, including proliferation, differentiation and CC
progression, in response to a variety of extracellular signals
(29).
Treatment with GSPs disrupts the
transcription of PC cells
There are intricate connections between CC
regulation and transcriptional control, and progression of the CC
is partially controlled at the transcriptional level in eukaryotes
(30). Therefore, disruption of the
transcription machinery may induce disruption of the CC.
GO analysis revealed that the DEGs identified in S3
vs. C3, S12 vs. C12 and S24 vs. C24 were enriched in the
‘regulation of transcription, DNA-dependent’ (GO:0006355) and
‘transcription, DNA-dependent’ (GO:0006351) terms (Fig. 2B, D and F). This indicates that normal
regulation of transcription may be disrupted in PANC-1 cells
following treatment with GSPs. Furthermore, the expression levels
of multiple DEGs associated with transcriptional regulation,
including E2F transcription factor 3 (E2F3) and hypoxia inducible
factor 1 α subunit (HIF1A), decreased in GSP-treated cells,
compared with control cells (Fig. 3).
E2F3 is a transcription factor that recognizes a specific sequence
motif in DNA and interacts directly with the retinoblastoma protein
to modulate the expression of genes involved in the CC (31). HIF1A functions as a master
transcriptional regulator of the adaptive response to hypoxia,
which serves as a stimulus for CC arrest (32). A number of genes associated with
regulation of transcription were also identified to be upregulated
in GSP-treated cells, compared with control cells. For example,
JunB proto-oncogene, a family member of the activator protein 1
transcription factors, was identified as an upregulated DEG in S3
vs. C3, S12 vs. C12 and S24 vs. C24 (data not shown). These
observations indicate that the transcriptional machinery in PANC-1
cells may be disrupted following treatment with GSPs, which may
result in disruption of the CC.
Treatment with GSPs disrupts DNA
replication and repair
Accurate DNA replication and repair are crucial in
the normal CC. The DEGs identified in S3 vs. C3, S12 vs. C12 and
S24 vs. C24 were enriched in the ‘DNA-dependent DNA replication’
(GO: 0006261), ‘DNA replication’ (GO: 0006260), ‘DNA repair’ (GO:
0006281) and ‘double-strand break repair via homologous
recombination’ (GO: 0000724) GO terms (Fig. 2B, D and F). The expression levels of
numerous DEGs enriched in the aforementioned terms, including
ataxia telangiectasia and Rad3-related (ATR), bloom syndrome RecQ
like helicase (BLM), nibrin (NBN) and RAD50 double strand break
repair protein (RAD50), were decreased in the PANC-1 cells treated
with GSPs, compared with the control cells (Fig. 3). ATR serves a critical role in the
DNA damage response (33). NBN serves
a role in the control of the intra-S-phase checkpoint (34). Additionally, evidence indicates that
NBN is involved in the G1 and G2 checkpoints, and is involved in
DNA double-strand break repair and DNA damage-induced checkpoint
activation (35). BLM serves a role
in DNA replication and repair (36).
RAD50, in cooperation with other molecules, is important in DNA
double-strand break repair, CC checkpoint activation, telomere
maintenance and meiotic recombination (37). A number of genes associated with DNA
replication and repair were also identified to be upregulated in
GSP-treated cells, compared with control cells. For example, the
ubiquitin C gene, which serves an important role in DNA repair
(38), was revealed to be an
upregulated DEG in S12 vs. C12 and S24 vs. C24 (data not shown).
These observations indicate that the regulation of DNA replication
and repair may be disrupted in PANC-1 cells following treatment
with GSPs, which may result in disruption of the CC.
Network analysis of DEGs reveals that
treatment with GSPs induces apoptosis
The aforementioned results revealed that disruption
of transcription and DNA replication may affect the CC of PC cells,
and induce apoptosis of cancer cells. To determine the
co-expression associations of the DEGs, a network was constructed
using the PCCs of the common DEGs (PPC≥0.95; P<0.01), which were
enriched in the ‘mitotic cell cycle’ (GO:0000278), ‘G2/M transition
of mitotic cell cycle’ (GO:0000086), ‘mitosis’ (GO:0007067), ‘cell
cycle’ (GO:0007049), ‘regulation of transcription, DNA-dependent’
(GO:0006355), ‘transcription, DNA-dependent’ (GO:0006351),
‘DNA-dependent DNA replication’ (GO:0006261), ‘DNA replication’
(GO:0006260), ‘DNA repair’ (GO:0006281) and ‘double-strand break
repair via homologous recombination’ (GO:0000724) pathways
(Fig. 4). The co-expression
associations between these DEGs indicated that disorders of
transcription, DNA replication, DNA repair and CC worked together
to induce apoptosis, resulting in the anticancer effects of
GSPs.
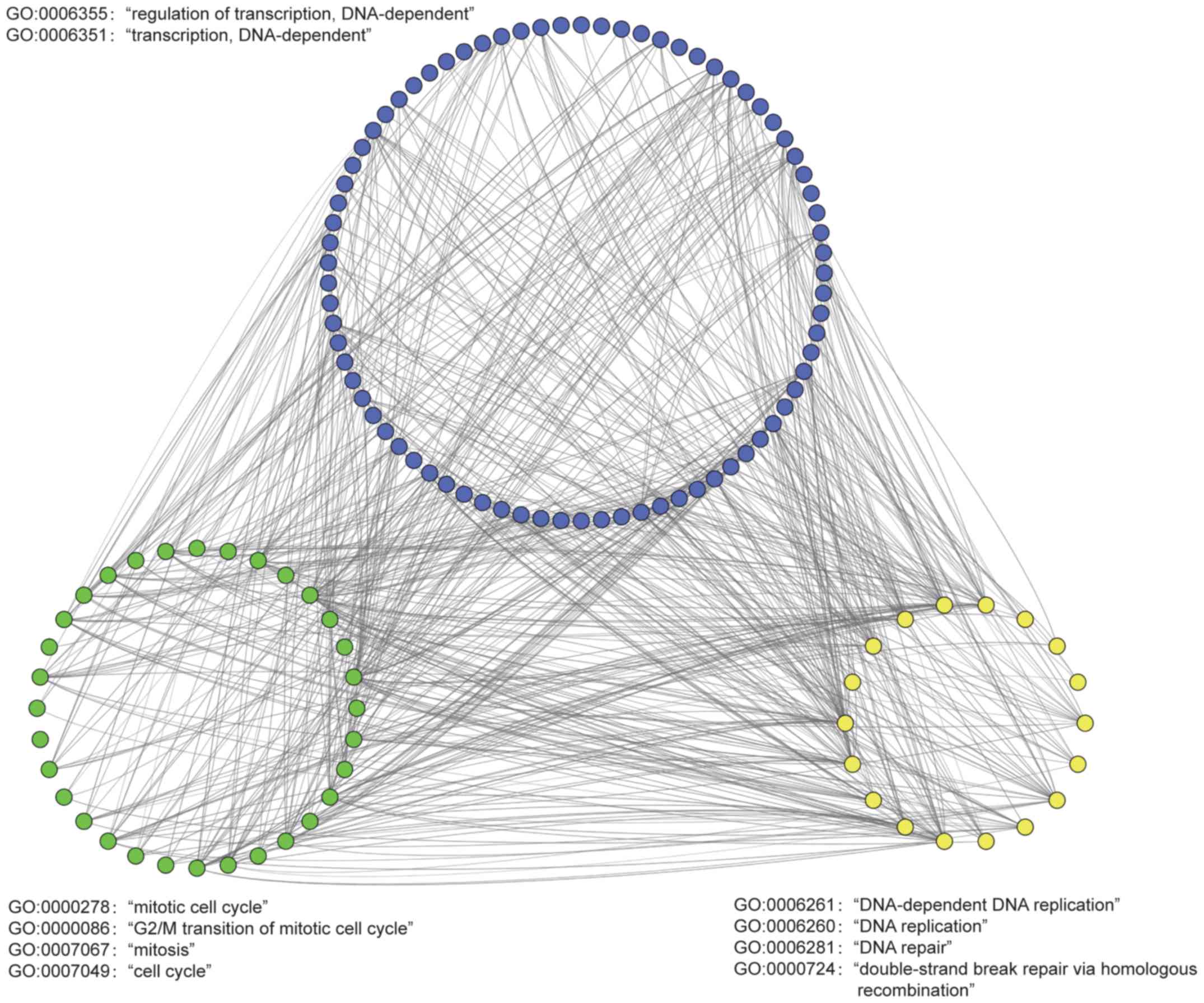 | Figure 4.Co-expression analysis of DEGs
associated with regulation of the cell cycle. Network analysis was
performed using the Pearson correlation coefficients of the common
DEGs enriched in ‘mitotic cell cycle’ (GO:0000278), ‘G2/M
transition of mitotic cell cycle’ (GO:0000086), ‘mitosis’
(GO:0007067), ‘cell cycle’ (GO:0007049), ‘regulation of
transcription, DNA-dependent’ (GO:0006355), ‘transcription,
DNA-dependent’ (GO:0006351), ‘DNA-dependent DNA replication’
(GO:0006261), ‘DNA replication’ (GO:0006260), ‘DNA repair’
(GO:0006281) and ‘double-strand break repair via homologous
recombination’ (GO:0000724). The nodes represent DEGs and the lines
signify significant correlations. PCC≥0.95 and P<0.01 indicated
a statistically significant correlation. DEG, differentially
expressed gene. |
Discussion
GSPs are biologically active components of grape
seeds and have been demonstrated to possess a wide array of
pharmacological and biochemical properties (39). Previous studies revealed that GSPs
exhibit inhibitory effects on various cancer types, including PC
(15,16,40), and
certain molecular mechanisms of the anticancer effects have been
identified (41). It has been
revealed that treatment with GSPs induces apoptosis mediated by the
mitochondrial-pathway in human colorectal carcinoma cells (42). It has also been demonstrated that GSPs
can inhibit the proliferation, migration and invasion of tongue
squamous cell carcinoma cells by suppressing the protein kinase
B/NF-κB signaling pathway (43).
In the present study, RNA-seq analysis indicated
that treatment with GSPs did not affect the mitochondrial-pathway
and the protein kinase B/NF-κB signaling pathway; however, the
expression levels of numerous genes involved in the regulation of
the CC were markedly affected in PC cells. This observation was
consistent with previous studies that have indicated that treatment
with GSPs can upregulate and downregulate gene expression in
certain functional pathways, including MAPK and cytokinesis
pathways (44). Furthermore, the
results of functional analysis of DEGs were consistent with those
of previous studies (15,44) and more targeted pathways were
identified, including specific pathways associated with cell
metabolism. For example, ‘metabolic process’ (GO:0008152) was
identified in S3 vs. C3, while ‘glycolysis’ (GO:0006096) and
‘cholesterol biosynthetic process’ (GO:0006695) were revealed in
S12 vs. C12 and S24 vs. C24. These observations indicate that GSPs
have the capacity to affect numerous aspects of PC cells and GSPs
are promising bioactive molecules for blocking the proliferation of
cancer cells.
In the normal CC, the control of gene expression
timing is of great importance and deregulation of the timing can be
detrimental (45). Numerous factors,
including transcription factors, kinases, DNA polymerase, kinesins
and cyclins, require precise timing of expression for their
function in CC regulation (46) and
dysregulation of the expression of these factors may inhibit cell
proliferation. For example, it has previously been identified that
oligomeric proanthocyanidins target cancer stem-like cells and
suppress tumor organoid formation in colorectal cancer by
inhibiting the Hippo pathway through downregulation of its key
transcriptional regulators, yes-associated protein (YAP) and
tafazzin (TAZ) (47). In the present
study, treatment with GSPs also affected the expression levels of
multiple genes involved in the regulation of transcription in PC
cells, indicating that the timing of normal transcription may be
disrupted during the CC. However, YAP and TAZ were not revealed as
DEGs in S3 vs. C3, S12 vs. C12 and S24 vs. C24, and results of KEGG
analysis indicated that ‘hippo signaling pathway’ (ID: hsa04390)
was not enriched in the three comparisons (data not shown). It has
been indicated that treatment with GSPs may disturb normal
transcription through different molecular mechanisms in different
types of cancer cell. Therefore, in the present study, apoptosis of
PC cells may be partially induced by the deregulation of
transcription through novel mechanisms that had not been reported
in previous studies.
It is understood that identical genetic information
must be precisely transmitted from the mother cell to the daughter
cells during successive cell proliferation cycles (48). If a cell enters mitosis with partially
replicated chromosomes or if a cell over-replicates its chromosomal
DNA, genetic alterations could occur, causing possible cell death
(49). Therefore, accurate DNA
replication and repair is crucial for the normal CC, and disrupting
the mechanism of DNA replication and repair is a practical strategy
to inhibit the proliferation of cancer cells (50,51). The
present study revealed that treatment with GSPs had the capacity to
interfere with the expression levels of multiple genes associated
with DNA replication and repair in PC cells, indicating that these
mechanisms may have been disrupted. This may be the reason why
treatment with GSPs induced CC arrest and apoptosis in PC
cells.
Acknowledgements
The authors would like to thank Dr Shengjie Liao and
Miss Hong Wu from ABLife Inc. for their scientific input and
editorial assistance.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the Gene Expression Omnibus
repository (accession no. GSE85610), https://www.ncbi.nlm.nih.gov/geo/; these data will be
made publicly available on August 19, 2019.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 31360409, 31471667
and 31660489), Program for Young and Middle-aged Leading Talents of
China Xinjiang Production and Construction Corps (grant no.
2017CB009) and the Selection and Cultivation Project of ‘Talents of
Xinjiang Production and Construction Corps’ (grant no.
00608017).
Authors' contributions
LZ, WW and ZH designed and managed the project. WW,
LZ and ZH drafted the manuscript. DG, YZ, YX and MT participated in
sample collection and manuscript revision. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kamisawa T, Wood LD, Itoi T and Takaori K:
Pancreatic cancer. Lancet. 388:73–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang X, Zhi X, Gao Y, Ta N, Jiang H and
Zheng J: LncRNAs in pancreatic cancer. Oncotarget. 7:57379–57390.
2016.PubMed/NCBI
|
|
3
|
Yeo TP, Hruban RH, Leach SD, Wilentz RE,
Sohn TA, Kern SE, Iacobuzio-Donahue CA, Maitra A, Goggins M, Canto
MI, et al: Pancreatic cancer. Curr Probl Cancer. 26:176–275. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li D, Xie K, Wolff R and Abbruzzese JL:
Pancreatic cancer. Lancet. 363:1049–1057. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rajkumar SV and Moreau P: Decade in
review-haematological cancer: Advances in biology and therapy. Nat
Rev Clin Oncol. 11:628–630. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gisselbrecht C and Sibon D: New
perspectives in the therapeutic approach of peripheral T-cell
lymphoma. Curr Opin Oncol. 30:285–291. 2018.PubMed/NCBI
|
|
7
|
Lee Y: Cancer chemopreventive potential of
procyanidin. Toxicol Res. 33:273–282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bagchi D, Bagchi M, Stohs S, Ray SD, Sen
CK and Preuss HG: Cellular protection with proanthocyanidins
derived from grape seeds. Ann N Y Acad Sci. 957:260–270. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dixon RA, Xie DY and Sharma SB:
Proanthocyanidins-a final frontier in flavonoid research? New
Phytol. 165:9–28. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Katiyar SK: Grape seed proanthocyanidines
and skin cancer prevention: Inhibition of oxidative stress and
protection of immune system. Mol Nutr Food Res. 52 Suppl 1:S71–S76.
2008.PubMed/NCBI
|
|
11
|
Bagchi D, Bagchi M, Stohs SJ, Das DK, Ray
SD, Kuszynski CA, Joshi SS and Pruess HG: Free radicals and grape
seed proanthocyanidin extract: Importance in human health and
disease prevention. Toxicology. 148:187–197. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Du Y and Lou H: Catechin and
proanthocyanidin B4 from grape seeds prevent doxorubicin-induced
toxicity in cardiomyocytes. Eur J Pharmacol. 591:96–101. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rossi M, Negri E, Parpinel M, Lagiou P,
Bosetti C, Talamini R, Montella M, Giacosa A, Franceschi S and La
Vecchia C: Proanthocyanidins and the risk of colorectal cancer in
Italy. Cancer Causes Control. 21:243–250. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Singh T, Sharma SD and Katiyar SK: Grape
proanthocyanidins induce apoptosis by loss of mitochondrial
membrane potential of human non-small cell lung cancer cells in
vitro and in vivo. PLoS One. 6:e274442011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Prasad R, Vaid M and Katiyar SK: Grape
proanthocyanidin inhibit pancreatic cancer cell growth in vitro and
in vivo through induction of apoptosis and by targeting the
PI3K/Akt pathway. PLoS One. 7:e430642012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Prasad R and Katiyar SK: Grape seed
proanthocyanidins inhibit migration potential of pancreatic cancer
cells by promoting mesenchymal-to-epithelial transition and
targeting NF-κB. Cancer Lett. 334:118–126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vaid M, Singh T and Katiyar SK: Grape seed
proanthocyanidins inhibit melanoma cell invasiveness by reduction
of PGE2 synthesis and reversal of epithelial-to-mesenchymal
transition. PLoS One. 6:e215392011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Punathil T and Katiyar SK: Inhibition of
non-small cell lung cancer cell migration by grape seed
proanthocyanidins is mediated through the inhibition of nitric
oxide, guanylate cyclase, and ERK1/2. Mol Carcinog. 48:232–242.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun Q, Prasad R, Rosenthal E and Katiyar
SK: Grape seed proanthocyanidins inhibit the invasive potential of
head and neck cutaneous squamous cell carcinoma cells by targeting
EGFR expression and epithelial-to-mesenchymal transition. BMC
Complement Altern Med. 11:1342011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Smeriglio A, Barreca D, Bellocco E and
Trombetta D: Proanthocyanidins and hydrolysable tannins:
Occurrence, dietary intake and pharmacological effects. Br J
Pharmacol. 174:1244–1262. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Regier N, Beauvais-Flück R, Slaveykova VI
and Cosio C: Elodea nuttallii exposure to mercury exposure under
enhanced ultraviolet radiation: Effects on bioaccumulation,
transcriptome, pigment content and oxidative stress. Aquat Toxicol.
180:218–226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Trapnell C, Pachter L and Salzberg SL:
TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics.
25:1105–1111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Robinson MD, McCarthy DJ and Smyth GK:
edgeR: A Bioconductor package for differential expression analysis
of digital gene expression data. Bioinformatics. 26:139–140. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang P, Larouche M, Normandin K, Kachaner
D, Mehsen H, Emery G and Archambault V: Spatial regulation of
greatwall by Cdk1 and PP2A-Tws in the cell cycle. Cell Cycle.
15:528–539. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mendoza M, Mandani G and Momand J: The
MDM2 gene family. Biomol Concepts. 5:9–19. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liszka L: Ductal adenocarcinoma of the
pancreas usually retained SMAD4 and p53 protein status as well as
expression of epithelial-to-mesenchymal transition markers and cell
cycle regulators at the stage of liver metastasis. Pol J Pathol.
65:100–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dynlacht BD: Regulation of transcription
by proteins that control the cell cycle. Nature. 389:149–152. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ping Z, Lim R, Bashir T, Pagano M and
Guardavaccaro D: APC/C (Cdh1) controls the proteasome-mediated
degradation of E2F3 during cell cycle exit. Cell Cycle.
11:1999–2005. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hubbi ME and Semenza GL: An essential role
for chaperone-mediated autophagy in cell cycle progression.
Autophagy. 11:850–851. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zou L: DNA replication checkpoint: New ATR
activator identified. Curr Biol. 27:R33–R35. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pennisi R, Antoccia A, Leone S, Ascenzi P
and di Masi A: Hsp90α regulates ATM and NBN functions in sensing
and repair of DNA double-strand breaks. FEBS J. 284:2378–2395.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wei L, Nakajima S, Böhm S, Bernstein KA,
Shen Z, Tsang M, Levine AS and Lan L: DNA damage during the G0/G1
phase triggers RNA-templated, Cockayne syndrome B-dependent
homologous recombination. PNAS. 112:E3495–3504. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Manthei KA and Keck JL: The BLM
dissolvasome in DNA replication and repair. Cell Mol Life Sci.
70:4067–4084. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lafrance-Vanasse J, Williams GJ and Tainer
JA: Envisioning the dynamics and flexibility of Mre11-Rad50-Nbs1
complex to decipher its roles in DNA replication and repair. Prog
Biophys Mol Biol. 117:182–193. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Franko J, Ashley C and Xiao W: Molecular
cloning and functional characterization of two murine cDNAs which
encode Ubc variants involved in DNA repair and mutagenesis. Biochim
Biophys Acta. 1519:70–77. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Katiyar SK, Pal HC and Prasad R: Dietary
proanthocyanidins prevent ultraviolet radiation-induced
non-melanoma skin cancer through enhanced repair of damaged
DNA-dependent activation of immune sensitivity. Semin Cancer Biol.
46:138–145. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chung YC, Huang CC, Chen CH, Chiang HC,
Chen KB, Chen YJ, Liu CL, Chuang LT, Liu M and Hsu CP: Grape-seed
procyanidins inhibit the in vitro growth and invasion of pancreatic
carcinoma cells. Pancreas. 41:447–454. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Katiyar SK and Athar M: Grape seeds: Ripe
for cancer chemoprevention. Cancer Prev Res (Phila). 6:617–621.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang C, Chen W, Zhang X, Zheng Y, Yu F,
Liu Y and Wang Y: Grape seed proanthocyanidins induce mitochondrial
pathway-mediated apoptosis in human colorectal carcinoma cells.
Oncol Lett. 14:5853–5860. 2017.PubMed/NCBI
|
|
43
|
Yang N, Gao J, Cheng X, Hou C, Yang Y, Qiu
Y, Xu M, Zhang Y and Huang S: Grape seed proanthocyanidins inhibit
the proliferation, migration and invasion of tongue squamous cell
carcinoma cells through suppressing the protein kinase B/nuclear
factor-κB signaling pathway. Int J Mol Med. 40:1881–1888.
2017.PubMed/NCBI
|
|
44
|
Chu H, Tang Q, Huang H, Hao W and Wei X:
Grape-seed proanthocyanidins inhibit the lipopolysaccharide-induced
inflammatory mediator expression in RAW264.7 macrophages by
suppressing MAPK and NF-κb signal pathways. Environ Toxicol
Pharmacol. 41:159–166. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fischer M and Müller GA: Cell cycle
transcription control: DREAM/MuvB and RB-E2F complexes. Crit Rev
Biochem Mol Biol. 52:638–662. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sadasivam S and DeCaprio JA: The DREAM
complex: Master coordinator of cell cycle-dependent gene
expression. Nat Rev Cancer. 13:585–595. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Toden S, Ravindranathan P, Gu J, Cardenas
J, Yuchang M and Goel A: Oligomeric proanthocyanidins (OPCs) target
cancer stem-like cells and suppress tumor organoid formation in
colorectal cancer. Sci Rep. 8:33352018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vermeulen K, Van Bockstaele DR and
Berneman ZN: The cell cycle: A review of regulation, deregulation
and therapeutic targets in cancer. Cell Prolif. 36:131–149. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nishitani H and Lygerou Z: Control of DNA
replication licensing in a cell cycle. Genes Cells. 7:523–534.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Murai J: Targeting DNA repair and
replication stress in the treatment of ovarian cancer. Int J Clin
Oncol. 22:619–628. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Helleday T: Amplifying tumour-specific
replication lesions by DNA repair inhibitors-a new era in targeted
cancer therapy. Eur J Cancer. 44:921–927. 2008. View Article : Google Scholar : PubMed/NCBI
|