Introduction
According to the statistics of the American Cancer
Society, in 2015, there were more than 230,000 new cases of breast
cancer diagnosed in the United States, and approximately 40,000
patients died of breast cancer (1).
It is estimated that one in eight women will develop breast cancer
in their lifetime, and it is also estimated that approximately one
in 37 women with breast cancer will die of the disease, most of
whom are aged over 45 years. The survival rate of patients with
non-metastatic breast cancer is relatively high (98.5%), but that
of patients with metastatic breast cancer is notably decreased
(24%) (2,3). Among patients with metastatic breast
cancer, approximately 20–30% of them have metastases of cancer
cells to lung tissue, liver and other sites, which then develop
into lung cancer and liver cancer, ultimately leading to death of
the patient (4–6). Breast cancer metastases to the brain
are observed in approximately 10–20% of women, resulting in a
decrease in life expectancy by 8–30 months (7–11). There
are many causes of breast cancer, among which one of the more
important risk factors is individual gene mutations including p53
and phosphatase and tensin homolog deleted on chromosome ten
protein (PTEN) genes. The study was approved by the Ethics
Committee of Dongying People's Hospital (Dongying, China).
As a member of G protein coupled receptor family,
ovarian cancer G-protein-coupled receptor 1 (OGR1) is named for its
first discovery in ovarian cancer cell line HEY. The expression of
OGR1 messenger ribonucleic acid (mRNA) is detected in many normal
tissues. The function and ligand of OGR1 have not been unified.
Some studies have proved that OGR1 has proton sensing function,
with proton as its ligand (12).
Other studies have demonstrated that OGR1 is able to sense changes
in proton concentration and maintain the balance of cell potential
of hydrogen (pH) microenvironment and bone metabolism. It has been
claimed that OGR1 can inhibit the growth and proliferation of
cancer cells. Moreover, it has been reported that OGR1 can inhibit
the metastasis of tumor cells in mice.
To investigate the function of OGR1, the
physiological function of breast cancer cells was studied through
constructing highly expressed OGR1 via transient transfection of
eukaryotic vector using breast cancer cells as the material in this
study.
Materials and methods
Cell culture
MCF-7 cells were cultured according to conventional
cell culture methods. Well-grown cells in logarithmic growth phase
were used for experiments. After the cells were transfected with
pIRESpuro3 vector and pIRESpuro3-OGR1 using Lipofectamine 2000, the
culture medium was replaced and the cells continued to be cultured
for a certain period of time for detection. The two groups of cells
were named W-MCF-7 and O-MCF-7, respectively.
Detection of cell proliferation
activity
According to the operation manual, single cell
suspension was prepared and inoculated into the culture plate with
100 µl cell suspension (approximately 5×103 cells) per
well. Cell Counting Kit-8 (CCK8) was adopted for determination of
cell proliferation activity at a specific time after
transfection.
RT-qPCR detection
According to the instructions, total RNA was
extracted using TRIzol (Thermo Fisher Scientific, Inc., Waltham,
MA, USA) from the cultured Michigan Cancer Foundation-7 (MCF-7)
cells, purified and reversely transcribed to obtain complementary
deoxyribonucleic acid (cDNA) with TaqMan microRNA reverse
transcription kit (Thermo Fisher Scientific, Inc.). The expression
levels of OGR1, serine-threonine kinase (AKT), p53 and other genes
were analyzed via qPCR. All reactions were repeated for at least
three times. The expression of mRNA was standardized by β-actin.
Calculation of all expression levels was conducted by
2−∆∆Cq method (13). All
primer pair sequences are summarized in Table I.
 | Table I.Primers used in fluorescence
quantitative qPCR assay. |
Table I.
Primers used in fluorescence
quantitative qPCR assay.
| Name | Primer pair |
|---|
| OGR1 | F:
AAAGCCAATCAGTGTGGGTATGG |
|
| R:
AGGATCTAGGCATCACTGGTGGT |
| AKT | F:
GTGCTGGAGGACAATGACTA |
|
| R:
AGCAGCCCTGAAAGCAAGGA |
| p53 | F:
CTGAGGTTGGCTCTGACTGTACCACCATCC |
|
| R:
CTCATTCAGCTCTCGGAACATCTCGAAGCG |
| β-actin | F:
CTTCCTTCCTGGGCATG |
|
| R:
GTCTTTGCGGATGTCCAC |
Detection of protein expression level
in cells via western blotting
The cells that have been cultured for a certain
period of time were collected for extraction of total protein in
accordance with the use requirements of the kit (total protein
extraction kit; Invent Biotechnologies Inc., Plymouth, MN, USA).
Protein extracted per lane (10 μl) was subjected to 10 % gel
electrophoresis and then transferred onto the polyvinylidene
fluoride (PVDF) membrane, followed by blocking with 5% milk at 20°C
for 1.5 h. According to the instructions, the mouse anti-human p53,
AKT, phosphorylated-AKT (p-AKT) and β-actin monoclonal antibodies
(1:400; cat. nos. sc-47698, sc-81434, sc-81433, 58673 all from
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) were added,
respectively, for reaction. After the membrane was washed,
horseradish peroxidase (HRP)-labeled second goat anti-mouse
polyclonal antibody (1:1,200; cat. no. sc-2005; Santa Cruz
Biotechnology, Inc.) was added for color development, and the gel
imaging analysis system was used for scanning.
Detection of cell apoptosis
For detection of apoptosis, cells cultured for
different time were collected and washed with phosphate buffered
saline (PBS) three times. The concentration was adjusted to
5×105/ml. Staining was conducted according to the
instructions of apoptosis detection kit. BD FACSCalibur (BD
Biosciences, Franklin Lakes, NJ, USA) was applied for detection of
apoptotic cells.
Statistical analysis
t-test was used for intergroup analysis, and
P<0.05 was considered to indicate a statistically significant
difference.
Results
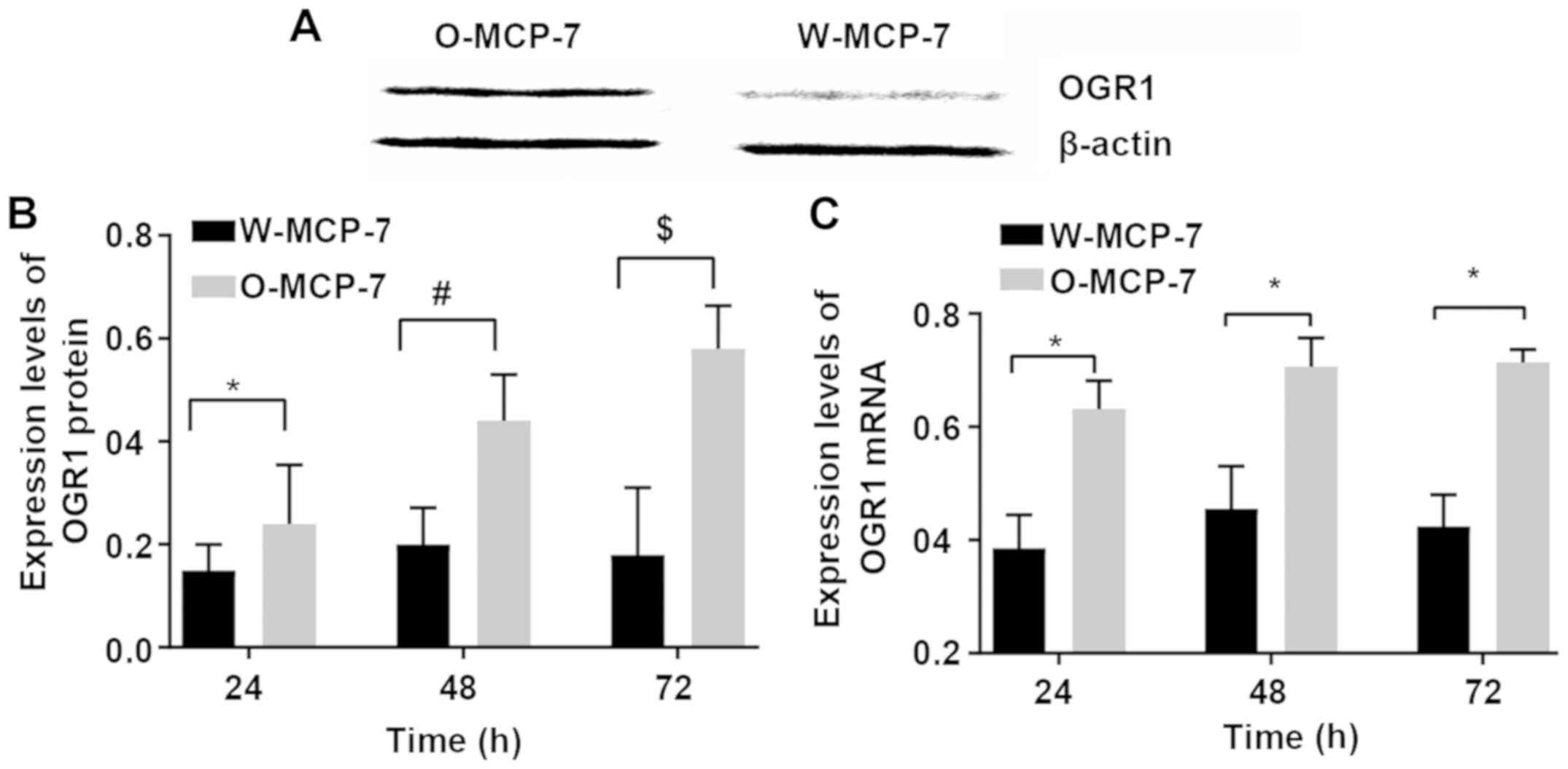
Expression levels of OGR 1 in the two
groups of cells
The relative expression levels of OGR1 mRNA at 24,
48 and 72 h in W-MCF-7 cells were 0.38±0.06, 0.46±0.08 and
0.42±0.06, respectively, while those in O-MCF-7 cells were
0.63±0.05, 0.70±0.03 and 0.70±0.02, respectively. The relative
expression levels of OGR1 protein at 24, 48 and 72 h were
0.15±0.04, 0.20±0.07 and 0.18±0.13, respectively, in W-MCF-7 cells,
and 0.24±0.11, 0.44±0.09 and 0.58±0.08, respectively, in O-MCF-7
cells. The above results indicated successful transfection of
O-MCF-7 cells and successfully high expression of OGR1 protein
(Fig. 1).
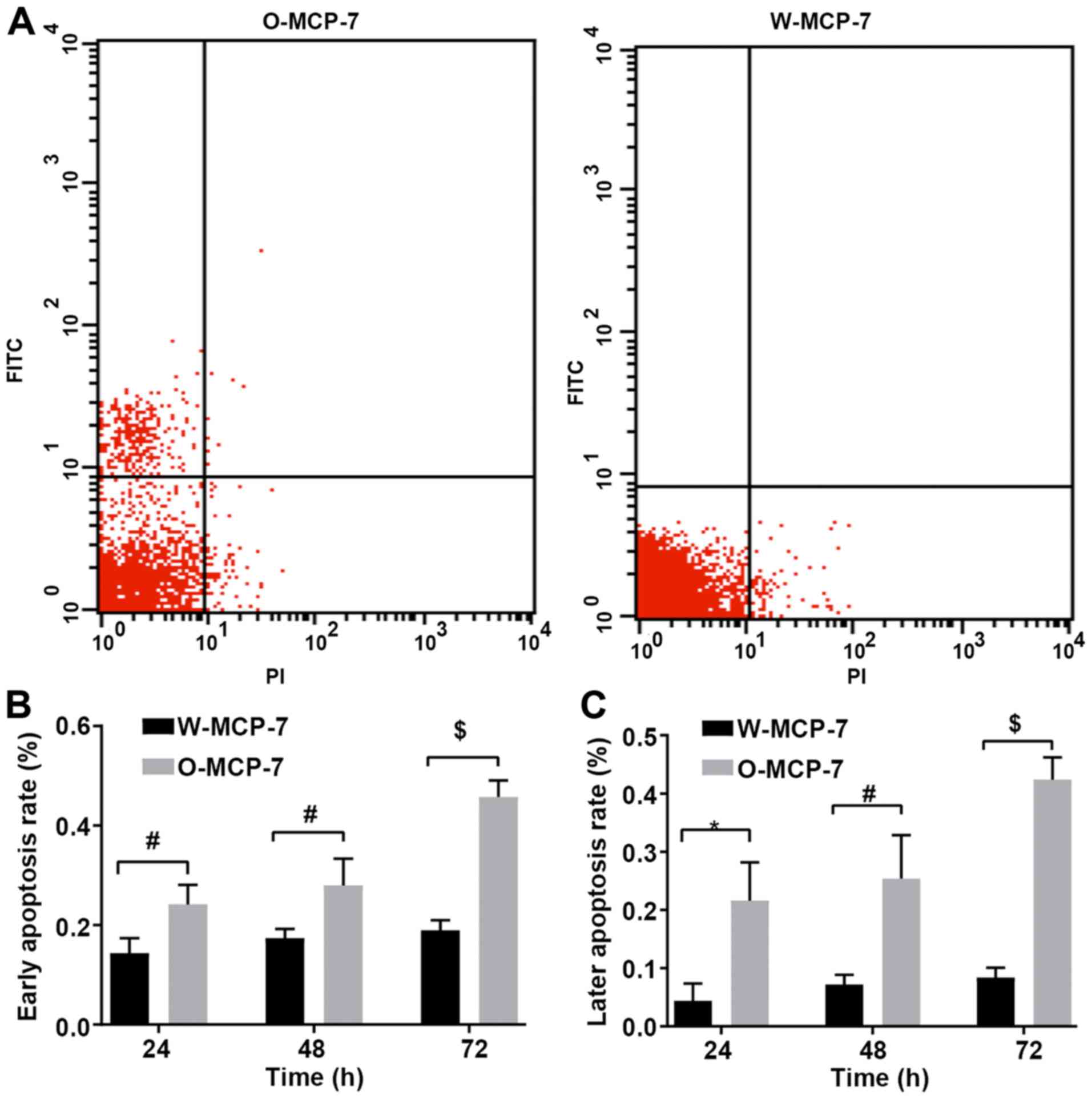
Detection of apoptosis by flow
cytometry
According to the results of flow cytometry, at 24,
48 and 72 h, the early apoptosis rates of W-MCF-7 cells were
0.144±0.03, 0.17±0.02 and 0.19±0.05%, respectively, while those of
cells transfected with O-MCF-7 were 0.24±0.04, 0.28±0.05 and
0.46±0.03%, respectively, and the late apoptosis rates of W-MCF-7
cells were 0.04±0.03, 0.07±0.02 and 0.08±0.02%, respectively, while
those of O-MCF-7 cells were 0.22±0.07, 0.25±0.07 and 0.42±0.04%,
respectively. The apoptosis rate was markedly decreased in W-MCF-7
group, compared with that in O-MCF-7 group, with a significant
difference. Thus it can be seen that OGR 1 can regulate apoptosis
through the signal pathway (Fig.
2).
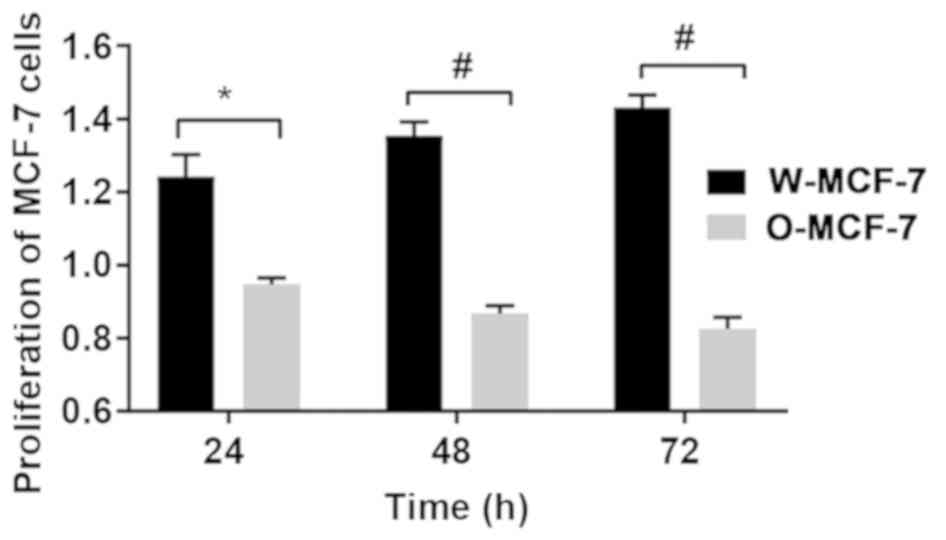
Detection of cell proliferation
At 24, 48 and 72 h, the relative proliferative
levels of W-MCF-7 cells were 1.24±0.06, 1.35±0.04 and 1.43±0.03,
respectively, while those of O-MCF-7 cells were 0.95±0.02,
0.87±0.02 and 0.83±0.03, respectively, suggesting that the
upregulation of OGR1 expression can regulate cell proliferation
through signal transduction (Fig.
3).
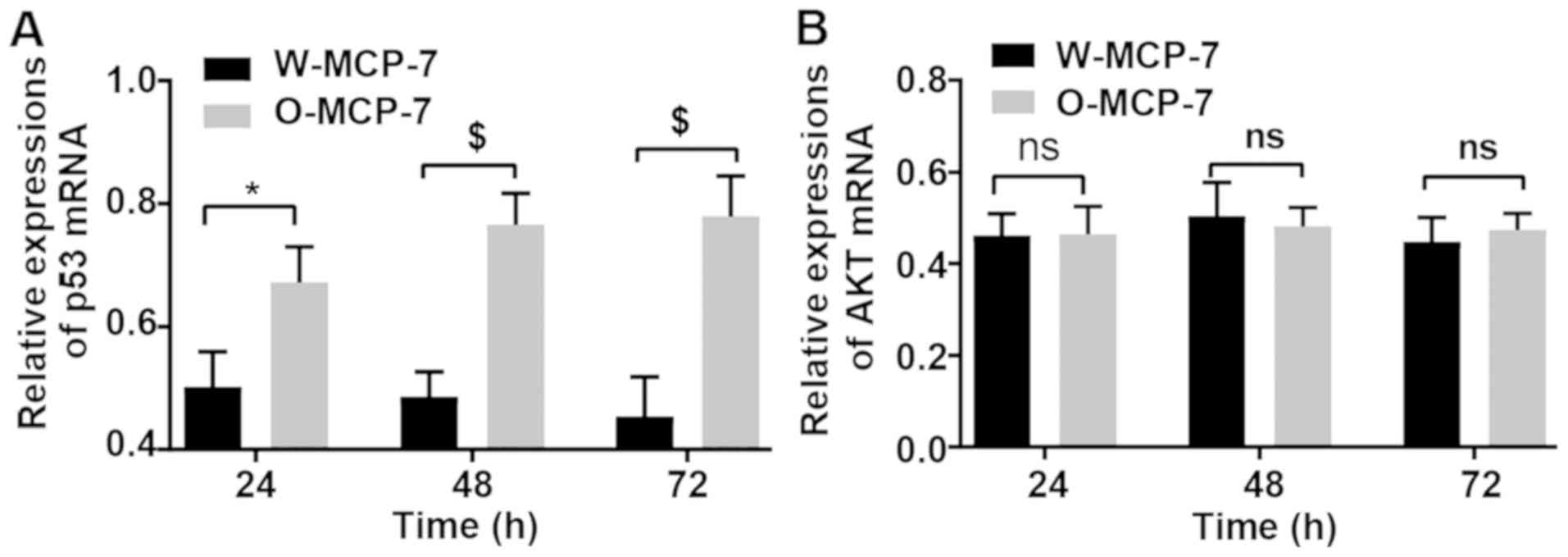
Detection of changes in gene
expression via RT-qPCR
At 24, 48 and 72 h, the relative expression levels
of p53 mRNA in W-MCF-7 cells were 0.50±0.07, 0.48±0.05 and
0.45±0.04, respectively, while those in O-MCF-7 cells were
0.67±0.06, 0.77±0.05 and 0.78±0.07, respectively. However, the mRNA
content of AKT displayed no remarkable difference between the two
groups (Fig. 4).
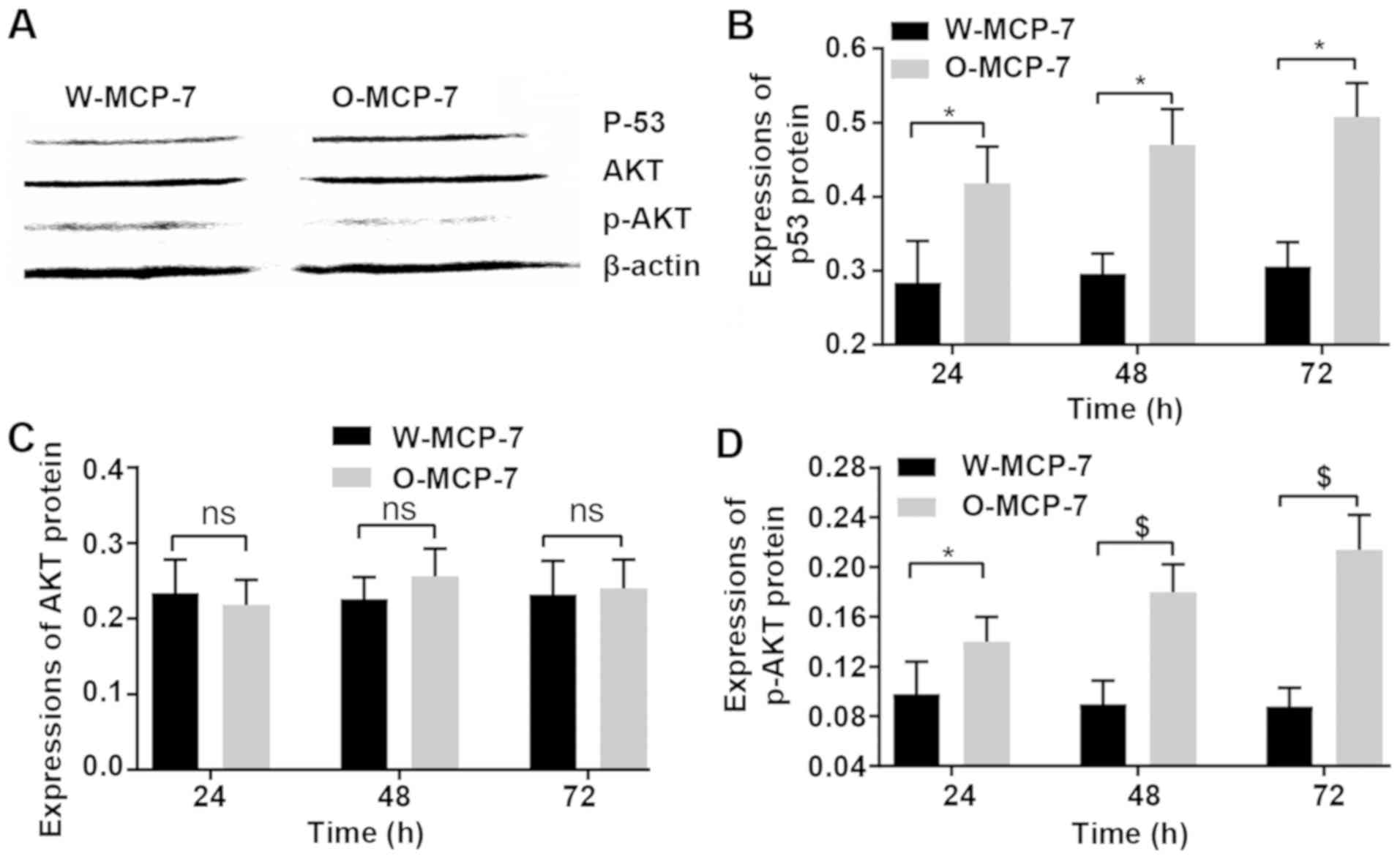
Effects of p53 and AKT protein
expression levels
At 24 and 48 and 72 h, the relative expression
levels of p53 in W-MCF-7 cells were 0.28±0.06, 0.30±0.03 and
0.31±0.03, respectively, while those in O-MCF-7 cells were
0.42±0.05, 0.47±0.04 and 0.51±0.05, respectively. However, the AKT
protein content showed no obvious difference between the two
groups. The relative expression levels of p-AKT at 24, 48 and 72 h
in W-MCF-7 cells were 10±0.03, 0.09±0.02 and 0.09±0.01,
respectively, and those in O-MCF-7 cells were 0.14±0.02, 0.18±0.040
and 0.21±0.03, respectively (Fig.
5).
Discussion
Breast cancer is a major cancer in women that
seriously threatens women's health. In particular, breast cancer
metastasis can cause serious consequences, leading to higher
recurrence and mortality rates. The occurrence of breast cancer is
associated with a variety of factors, such as environment, personal
life state and heredity, and it involves the intermolecular
information transmission within cells, and extracellular or
intercellular interactions and signal transmission. The gradual
transmission and interactions of these signals alter migration,
infiltration and growth processes of tumor cells. Tumor cell growth
is a dynamic process regulated and controlled by multiple factors.
In this study, the effects of OGR1 on the proliferation and
apoptosis of breast cancer cells were explored, and the possible
mechanism of its action is preliminarily discussed.
The results of apoptosis experiment demonstrated
that the highly expressed OGR1 in breast cancer cells could
effectively enhance apoptosis. Cell proliferation experiment
displayed that the growth and proliferation abilities of breast
cancer cells with highly expressed OGR1 were inhibited to some
extent, compared with those of breast cancer cells with low
expression of OGR1. According to some research, OGR1 deficiency in
prostate cancer model of transgenic mice reduces tumor formation
(14,15). The expression of OGR1 in metastatic
tumors was lower than that in primary prostate cancer tissues. Li
et al reported that OGR1 inhibits cell migration through
molecular signal transduction pathway in MCF-7 breast cells
(16). Recent research has also
indicated that OGR1 can activate calcium sensitive protease and its
downstream signaling molecules such as Bid, Bax and caspase-3, thus
inducing chondrocyte apoptosis in rats (17). These results manifested that OGR1 may
act as an inhibitory factor for tumor metastasis. However, the
anti-tumor effect of OGR1 needs to be further investigated.
Results of western blotting displayed that the gene
and protein expression levels of p53 in breast cancer cells with
highly expressed OGR1 were also increased. According to current
studies, p53 is a very important tumor suppressor gene in human
body, which can inhibit cell cycle progression, mediate cell cycle
arrest, inhibit cell proliferation, promote cell senescence and
regulate cell apoptosis, thus playing an important role in
inhibiting the development of tumors. At the same time, some
studies have revealed that regulation of p53 on cell metabolic
activity is also an important means of its anti-tumor effect. The
decreased p53 gene in human body will increase the occurrence and
development of tumors. The results suggest that OGR1 protein and
p53 protein interact with each other, transmit information, and
further regulate the expression and activity control of downstream
or other related proteins, thereby manipulating tumor cells.
Furthermore, the findings of this study indicated
that no obvious difference in protein expression of AKT between
breast cancer cells with low expressio of OGR1 and those with
highly expressed OGR1 was found, but the protein content of p-AKT
in the latter was reduced. The latest research findings have
revealed that the activated AKT protein can regulate the survival
state (18) and specific
physiological function of cells (19,20)
through multiple signaling pathways. Besides, p53 regulates nitric
oxide synthase in endothelial cells to regulate tumor angiogenesis
(21,22). Finally, AKT can interact with
phosphatidylinositol 3-kinase (PI3K) to participate in the invasion
and migration of tumor cells (23,24),
which is consistent with the fact that OGR1 can inhibit the
migration of breast cancer cells (16). Previous research on prostate cancer
has manifested that the migration ability of cancer cells with
highly expressed OGR1 is inhibited (25).
The results of this study indicated that the
expression level of OGR1, which has an effect on the proliferation
and apoptosis of breast cancer cells, may be related to the
expression levels of p-AKT and p53, and it regulates the expression
of related genes, thus influencing the function of breast cancer
cells. However, the risk factors of breast cancer are very
complicated. The growth of tumor cells is a physiological mechanism
regulated by multiple networks. Further studies on the pathogenesis
of breast cancer will contribute to understanding the pathogenesis
of tumors in a more detail way, providing support for better
diagnostic and therapeutic applications.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ wrote the manuscript. JZ and WS helped with cell
culture. LC, JS and MH were responsible for PCR and western
blotting. MT detected cell apoptosis. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Dongying People's Hospital (Dongying, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests
References
|
1
|
Sirkisoon SR, Carpenter RL, Rimkus T,
Miller L, Metheny-Barlow L and Lo HW: EGFR and HER2 signaling in
breast cancer brain metastasis. Front Biosci (Elite Ed). 8:245–263.
2016.PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fidler IJ: The pathogenesis of cancer
metastasis: The ‘seed and soil’ hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nola S, Sin S, Bonin F, Lidereau R and
Driouch K: A methodological approach to unravel organ-specific
breast cancer metastasis. J Mammary Gland Biol Neoplasia.
17:135–145. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
O'Shaughnessy J: Extending survival with
chemotherapy in metastatic breast cancer. Oncologist. 10 (Suppl
3):20–29. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shao MM, Liu J, Vong JS, Niu Y, Germin B,
Tang P, Chan AW, Lui PC, Law BK, Tan PH, et al: A subset of breast
cancer predisposes to brain metastasis. Med Mol Morphol. 44:15–20.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hohensee I, Lamszus K, Riethdorf S,
Meyer-Staeckling S, Glatzel M, Matschke J, Witzel I, Westphal M,
Brandt B, Müller V, et al: Frequent genetic alterations in EGFR-
and HER2-driven pathways in breast cancer brain metastases. Am J
Pathol. 183:83–95. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aragon-Ching JB and Zujewski JA: CNS
metastasis: An old problem in a new guise. Clin Cancer Res.
13:1644–1647. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brufsky AM, Mayer M, Rugo HS, Kaufman PA,
Tan-Chiu E, Tripathy D, Tudor IC, Wang LI, Brammer MG, Shing M, et
al: Central nervous system metastases in patients with
HER2-positive metastatic breast cancer: Incidence, treatment, and
survival in patients from registHER. Clin Cancer Res. 17:4834–4843.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Klein A, Olendrowitz C, Schmutzler R,
Hampl J, Schlag PM, Maass N, Arnold N, Wessel R, Ramser J, Meindl
A, et al: Identification of brain- and bone-specific breast cancer
metastasis genes. Cancer Lett. 276:212–220. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Niikura N, Hayashi N, Masuda N, Takashima
S, Nakamura R, Watanabe K, Kanbayashi C, Ishida M, Hozumi Y,
Tsuneizumi M, et al: Treatment outcomes and prognostic factors for
patients with brain metastases from breast cancer of each subtype:
A multicenter retrospective analysis. Breast Cancer Res Treat.
147:103–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ludwig MG, Vanek M, Guerini D, Gasser JA,
Jones CE, Junker U, Hofstetter H, Wolf RM and Seuwen K:
Proton-sensing G-protein-coupled receptors. Nature. 425:93–98.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yan L, Chang Z, Liu Y, He BR and Hao DJ:
Primary spinal melanoma: A case report and literature review. Chin
Med J (Engl). 125:4138–4141. 2012.PubMed/NCBI
|
|
15
|
Yan L, Singh LS, Zhang L and Xu Y: Role of
OGR1 in myeloid-derived cells in prostate cancer. Oncogene.
33:157–164. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Guo B, Wang J, Cheng X, Xu Y and
Sang J: Ovarian cancer G protein coupled receptor 1 suppresses cell
migration of MCF7 breast cancer cells via a Gα12/13-Rho-Rac1
pathway. J Mol Signal. 8:62013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yuan FL, Wang HR, Zhao MD, Yuan W, Cao L,
Duan PG, Jiang YQ, Li XL and Dong J: Ovarian cancer G
protein-coupled receptor 1 is involved in acid-induced apoptosis of
endplate chondrocytes in intervertebral discs. J Bone Miner Res.
29:67–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Datta SR, Brunet A and Greenberg ME:
Cellular survival: A play in three Akts. Genes Dev. 13:2905–2927.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Muise-Helmericks RC, Grimes HL, Bellacosa
A, Malstrom SE, Tsichlis PN and Rosen N: Cyclin D expression is
controlled post-transcriptionally via a phosphatidylinositol
3-kinase/Akt-dependent pathway. J Biol Chem. 273:29864–29872. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mayo LD and Donner DB: A
phosphatidylinositol 3-kinase/Akt pathway promotes translocation of
Mdm2 from the cytoplasm to the nucleus. Proc Natl Acad Sci USA.
98:11598–11603. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Michell BJ, Griffiths JE, Mitchelhill KI,
Rodriguez-Crespo I, Tiganis T, Bozinovski S, de Montellano PR, Kemp
BE and Pearson RB: The Akt kinase signals directly to endothelial
nitric oxide synthase. Curr Biol. 9:845–848. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hurt KJ, Musicki B, Palese MA, Crone JK,
Becker RE, Moriarity JL, Snyder SH and Burnett AL: Akt-dependent
phosphorylation of endothelial nitric-oxide synthase mediates
penile erection. Proc Natl Acad Sci USA. 99:4061–4066. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim D, Kim S, Koh H, Yoon SO, Chung AS,
Cho KS and Chung J: Akt/PKB promotes cancer cell invasion via
increased motility and metalloproteinase production. FASEB J.
15:1953–1962. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kubiatowski T, Jang T, Lachyankar MB,
Salmonsen R, Nabi RR, Quesenberry PJ, Litofsky NS, Ross AH and
Recht LD: Association of increased phosphatidylinositol 3-kinase
signaling with increased invasiveness and gelatinase activity in
malignant gliomas. J Neurosurg. 95:480–488. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Iremashvili V, Burdick-Will J and Soloway
MS: Improving risk stratification in patients with prostate cancer
managed by active surveillance: A nomogram predicting the risk of
biopsy progression. BJU Int. 112:39–44. 2013. View Article : Google Scholar : PubMed/NCBI
|