Introduction
Colorectal cancer (CRC) is one of the most common
cancers, affecting more than 1.2 million patients and resulting in
600,000 deaths annually (1,2). According to the National Cancer
Information Center of Korea, CRC has the third highest incidence
rate in both males and females, and the second highest mortality
rate in females (3). Established CRC
risk factors include exposure to processed meats and alcohol,
smoking, and gamma radiation; however, recent studies have likewise
investigated the effects of the microRNA (miR/miRNA)-34 miRNA
precursor family (1,2).
MicroRNAs (miRNAs, miRs) are small, 22–24 nucleotide
non-coding RNAs that negatively regulate the translation of
messenger RNA to protein via base pairing to a partially
complementary sequence in the open reading frames and
3′-untranslated regions (3′-UTR) (4,5).
Further, miRNAs are encoded across the entire genome, including
exonic, intronic, and intergenic regions. However, almost all
miRNAs are found in intronic regions (6). The RNA polymerases RNase II or III
transcribe miRNAs as long primary transcripts (pri-miRNAs), which
are then processed into stem-loop structure miRNA precursor
molecules (pre-miRNAs) in the nucleus by a nuclear complex
consisting of Drosha, a member of the ribonuclease III family
(RNase III), and its cofactors (DGCR8) (7,8). The
pre-miRNAs are exported to the cytoplasm by exportin-5 (XPO5) and
processed into mature, 18–25 nucleotide (nt) miRNAs following
cleavage of the double-stranded portion of the hairpin by the RNase
III enzyme Dicer (9,10). Altered miR expression resulting from
deregulation occurs in several human diseases including cancer. As
well, miRs act as either tumor suppressor genes or oncogenes,
resulting in the dissemination of cancer (11–13). The
deregulation of miR via the epigenetic silencing of miR expression
is likewise associated with CpG island methylation and repressive
histone modifications (11).
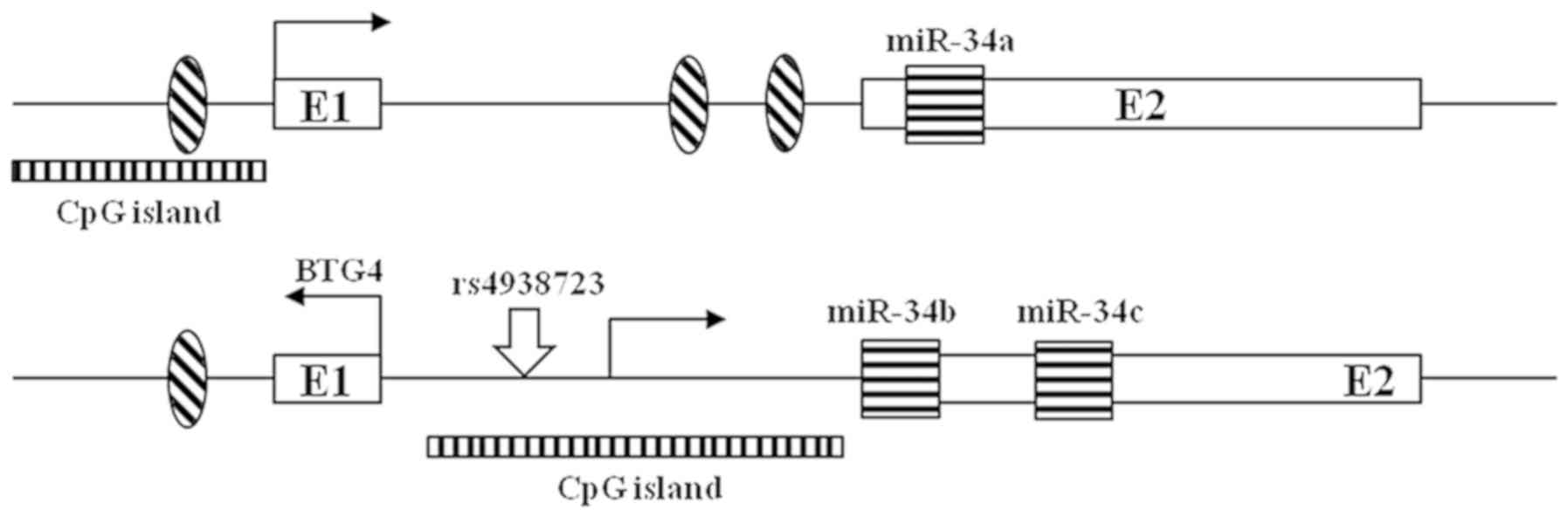
The miRNA-34 family, comprised of miR-34a, miR-34b,
and miR-34c, is part of the p53 network, which induces the
expression of miR-34 miRNAs in response to DNA damage or oncogenic
stress (14–16). Located on chromosome 1p36, miR-34a is
associated with glioma, neuroblastoma, pancreatic cancer, and
chronic myelogenous leukemia (17–20), and
is the key regulator of cell cycle progression (E2F3) and apoptosis
(BCL2) (14,15,20).
Reduced miR-34a expression is frequently observed in pancreatic
tumors and neuroblastomas (18,21).
Located on chromosome 11q23, miR-34b and miR-34c are co-transcribed
from one transcription unit (22).
Reduced miR-34b/c expression is seen in non-small cell lung cancer
(20). The methylation of miR-34b/c
CpG sites has been found in CRC (23) and oral squamous cell carcinoma
(24) as well as malignant melanoma,
in which miR-34b/c CpG site methylation is likewise correlated with
metastatic potential (25). The
cancer-related downregulation of members of the miR-34 family
indicates that these miRs function as tumor-suppressor genes,
suggesting a potential prognostic marker role (26).
Several mechanisms, including gene amplification,
deletion, epigenetic alterations, and single-nucleotide
substitutions, have been implicated as potential regulators of miR
expression; however, to date, no studies have actually demonstrated
a precise regulatory role for miR expression (26,27).
Although single nucleotide polymorphisms (SNPs) in miRs are not
considered functionally important, nucleotide variations in primary
(pri)- or precursor (pre)-miRs may affect miR processing and result
in modified miR expression (28).
Recently, studies reported that rs4938723, a potentially functional
SNP in the promoter region of pri-miR-34b/c, may contribute to
susceptibility to hepatocellular carcinoma (29), CRC (30), endometrial cancer (31), and decreased breast cancer survival
(32). However, reports on the
relationship between SNPs in the miR-34b/c promoter and the
subsequent risk and prognostic significance in CRC patients are
limited.
Mutations in p53 reportedly initiate or participate
in early events in several diverse cancers types. Prior reports
likewise indicate that the tumorigenicity is related to the
deregulation of p53-mediated transcription (28). Of the variations in the TP53
gene, Arg72Pro is the most widely investigated. The 72Arg allele
induces apoptosis more efficiently than the 72Pro allele (32). Prior reports have indicated that Pro
homozygosity in TP53Arg72Pro is a potential risk factor for
lung, esophageal, stomach, breast, nasopharynx, urothelium, and
prostate cancers (33,34). However, the results of a
meta-analysis of CRC, performed to estimate the effect of the
TP53Arg72Pro polymorphism on CRC risk, failed to identify a
significant association (35).
Conversely, we detected a negative relationship between the
TP53Arg72Pro polymorphism and CRC risk in the Korean
population. Transcriptional silencing of CpG methylation represents
an important mechanism for the inactivation of key tumor suppressor
genes (25,36), and approximately 60% of CpG islands
are located in promoter regions. However, only CpG islands located
in promoter regions exhibit methylation of cytosine at position 5
and the inactivation of surrounding chromatin via the recruitment
of histone deacetylases following proteins binding to methylated
CpG residues (37). Therefore, the
aim of this study has investigated the differences in the degree of
methylation according to SNPs in the DNA promoter region (Fig. 1) and confirmed the changes in the
expression pattern of the miR-34 family according to the gene
mutation of p53.
Materials and methods
Tissue samples and genomic DNA
isolation
Paired tumor and normal tissues were retrospectively
selected from 104 CRC patients treated at the CHA Bundang Medical
Center (Seongnam, Korea) between March 2010 and March 2012. Genomic
DNA (gDNA) isolation was performed according to an established
protocol (38). Briefly, 50 µl of
solution 1 (420 µl STE buffer and 10% SDS) was added to the tissues
followed by the addition of 30 µl proteinase K and overnight
incubation at 50°C. After incubation, 500 µl of solution 2
(phenol:chloroform:isoamyl alcohol=25:24:1) was added to the tissue
samples and the supernatants were collected following
centrifugation. Next, 500 µl of solution 3 (chloroform:isoamyl
alcohol=24:1) was added, the supernatants were collected after
centrifugation, and 25 µl 3 M sodium acetate and 900 µl 100% EtOH
were added. The samples were centrifuged again, and the resulting
supernatants were quenched on ice for 1 h. The resulting gDNA
samples were dissolved in the rehydration buffer. The present study
was approved by the Institutional Review Board of CHA Bundang
Medical Center; IRB no. 2009-08-077-010) and written informed
consent was provided by all patients.
Genetic analysis
Three TP53 single nucleotide polymorphisms (SNPs),
TP53 codon 72G>C (rs1042522, Arg>Pro, exon 4), TP53
MSPI A>G (rs1625895, intron 6), and TP53 PIN (rs17878362,
intron 6), and one miR-34b/c SNP, miR-34bc T>C (rs4938723,
promoter) were selected from the human genome SNP database (dbSNP,
www.ncbi.nlm.nih.gov/snp). The genotypes
were determined using a polymerase chain reaction (PCR)-restriction
fragment length polymorphism assay. The PCR primers for the TP53
codon 72G>C polymorphism were forward
5′-TTGCCGTCCCAAGCAATGGATGA-3′ and reverse
5′-TCTGGGAAGGGACAGAAGATGAC-3′. The PCR conditions included an
initial 5 min of denaturation at 94°C followed by 35 cycles of
denaturing at 94°C for 30 sec, annealing at 56°C for 30 sec, and
extension at 72°C for 30 sec followed by a final extension at 72°C
for 5 min. The PCR products were cut using the BstUI
restriction enzyme (New England Biolabs, Beverly, MA, USA) at 60°C
for 16 h.
The PCR primers for the TP53 MSPI A>G
polymorphism were forward 5′-ATAGTGTGGTGGTGCCCTAT-3′ and reverse
5′-CCTTAGCCTCTGTAAGCTTCA-3′, and the primers for the TP53 PIN
polymorphism were forward: 5′-GACTGACTTTCTGCTCTTGTCTT-3′ and
reverse 5′-ATCGTCCGG-3′. The PCR conditions included an initial 5
min of denaturation at 94°C followed by 35 cycles of denaturing at
94°C for 30 sec, annealing at 60°C for 30 sec, and extension at
72°C for 30 sec followed by a final extension at 72°C for 5 min.
The PCR products were cut using the MSPI restriction enzyme
(New England Biolabs) at 37°C for 16 h.
The PCR primers for the miR-34b/c T>C
polymorphism were forward 5′-CCTCTGGGAACCTTCTTTGACCAAT-3′ and
reverse 5′-TGAGATCAAGGCCATACCATTCAAGA-3′. The PCR conditions
included an initial 5 min of denaturation at 94°C was followed by
35 cycles of denaturing at 94°C for 30 sec, annealing at 55°C for
30 sec, and extension at 72°C for 30 sec followed by a final
extension at 72°C for 5 min. The PCR products were cut using the
Tsp509I restriction enzyme (New England Biolabs) at 65°C for
16 h.
All of the resulting PCR products were analyzed by
gel electrophoresis. In this study, the designated A1 and A2
alleles for the TP53 PIN polymorphism indicated a 16-base pair (bp)
deletion and insertion in intron 6, respectively.
gDNA bisulfite modification and
methylation analysis
The DNA was treated with bisulfite using an EZ DNA
methylation kit (Zymo Research, Irvine, CA, USA). The modified DNA
was then eluted in a final volume of 10 µl and 1 µl was used for
the real-time methylation PCR (MethyLight). Typically,
methylation-specific PCR (MSP) is used to measure DNA methylation,
but we opted to use the semi-Quantitative MethyLight technology
(39). In the MethyLight technology,
the discrimination is made during the PCR amplification step using
primers and probes that specifically anneal to either the converted
methylated or converted unmethylated sequence. Real-time PCR was
performed in triplicate to confirm the results, and the average
value was used. The EpiTect® PCR control DNA set was
also used (Qiagen, Valencia, CA, USA). The resulting data were
analyzed using the Cmeth method (EpiTect® MethyLight PCR
Handbook; Qiagen) (40).
RNA extraction and reverse
transcriptase reaction
Total RNA was extracted from each tumor and normal
tissue sample using the TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) and treated with
RNase-free DNase (Promega, Madison, WI, USA). The concentration of
the RNA was measured using a spectrophotometer and the samples were
stored at −80°C until analysis. Reverse transcription (RT) of the
total RNA was performed using an Invitrogen RT kit according to the
manufacturer's instructions.
RT-quantitative polymerase chain
reaction (RT-qPCR) analysis
The expression of miR was measured by qPCR using the
EvaGreen® Master Mix, primers for each gene, and the
Rotor-Gene RG-3000 thermal cycler (Corbett Research). The primer
sequences were as follows (forward and reverse): ACTB (213 bp)
5′TGACATTAAGGAGAAGCTGTGCTAC3′ and 5′GAGTTGAAGGTAGTTTCGTGGATG3′;
miR-34a (128 bp) 5′CGTCACCTCTTAGGCTTGGA3′ and
5′CATTGGTGTCGTTGTGCTCT3′; and miR-34b/c (84 bp)
5′-GTGCTCGGTTTGTAGGCAGT3′ and 5′-GTGCCTTGTTTTGATGGCAG3′ (7,41). The
expression results were calculated using the -∆∆Cq method and
expressed graphically using ACTB (β-actin) as the reference gene
(42). The experiments were
performed in triplicate and the average values were used.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 4.0 (GraphPad Software, Inc., La Jolla, CA, USA) and MedCalc
version 12.1.4 (MedCalc Software bvba, Ostend, Belgium). The
genotype and allele frequencies were calculated to investigate
deviations from Hardy-Weinberg equilibrium. The expression pattern
of the miRNA identified in CRC tissues was analyzed using Student's
t-test and one-way analysis of variance with Student-Newman-Keuls
post hoc test. The allele combinations were estimated with SNPAlyze
version 5.1 (Dynacom Co., Ltd., Nakase, Japan) and HapStat version
3.0 (dlin.web.unc.edu/). The results are
presented as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
Of the 104 CRC patients, 54 were colon cancer
patients and 50 were rectal cancer patients. The mean age was 64
years. Other clinical characteristics including sex, TNM stage,
tumor location, and tumor size are summarized in Table I. The relationship between miR-34
methylation level and clinical characteristics was further
evaluated. A significant association was found between miR-34a
methylation level and age in tumor tissues (>64 vs. ≤64 years;
P=0.035). However, no significant association was detected between
miR-34 methylation level and sex, location of the primary tumor,
TNM stage, tumor size, or BMI (Table
I).
 | Table I.Associations between miR-34
methylation level and clinical characteristics of colorectal cancer
patients. |
Table I.
Associations between miR-34
methylation level and clinical characteristics of colorectal cancer
patients.
|
|
| Normal tissue | Tumor tissue |
|---|
|
|
|
|
|
|---|
| Characteristic | Total n (n=104), n
(%) | miR-34a |
P-valuea | miR-34b/c |
P-valuea | miR-34a |
P-valuea | miR-34b/c |
P-valuea |
|---|
| Sex |
|
|
|
|
|
|
|
|
|
|
Male | 61 (58.7) | 37.07±37.42 | 0.851 | 12.40±19.17 | 0.628 | 46.46±36.30 | 0.554 | 57.25±31.54 | 0.407 |
|
Female | 43 (41.3) | 35.71±35.18 |
| 14.31±20.11 |
| 50.77±36.70 |
| 51.71±34.73 |
|
| Age, years |
|
|
|
|
|
|
|
|
|
|
>64 | 55 (52.9) | 36.76±34.93 | 0.897 | 11.73±18.55 | 0.381 | 56.93±35.08 | 0.035c | 54.60±35.96 | 0.864 |
|
≤64 | 49 (47.1) | 35.84±37.15 |
| 15.12±20.63 |
| 41.92±36.45 |
| 53.47±31.27 |
|
| TNM stage |
|
|
|
|
|
|
|
|
|
| 1 | 5
(4.8) | 34.82±41.51 | 0.967b | 12.05±15.42 | 0.178b | 63.02±42.83 | 0.728b | 64.40±34.64 | 0.073b |
| 2 | 23 (22.1) | 33.17±34.05 |
| 21.17±25.92 |
| 46.07±37.16 |
| 59.42±36.58 |
|
| 3 | 66 (63.5) | 37.06±36.15 |
| 11.89±18.19 |
| 47.88±36.92 |
| 55.16±31.65 |
|
| 4 | 10 (9.6) | 38.95±41.42 |
| 7.43±8.54 |
| 56.05±30.79 |
| 28.70±29.55 |
|
| Tumor site |
|
|
|
|
|
|
|
|
|
|
Colon | 54 (51.9) | 35.08±34.57 | 0.726 | 16.60±22.83 | 0.097 | 51.15±37.46 | 0.532 | 53.57±33.07 | 0.892 |
|
Rectum | 50 (48.1) | 37.57±37.69 |
| 10.20±15.06 |
| 46.66±35.49 |
| 54.46±34.09 |
|
| Tumor size, cm |
|
|
|
|
|
|
|
|
|
|
<5 | 38 (36.5) | 42.41±37.19 | 0.188 | 13.56±19.01 | 0.989 | 54.69±36.71 | 0.227 | 51.61±35.91 | 0.582 |
| ≥5 | 66 (63.5) | 32.74±35.02 |
| 13.50±20.17 |
| 45.70±36.12 |
| 55.38±32.07 |
|
| BMI,
kg/m2 |
|
|
|
|
|
|
|
|
|
|
≤25 | 80 (76.9) | 38.77±36.25 | 0.198 | 13.34±19.94 | 0.863 | 49.24±35.92 | 0.899 | 53.15±33.62 | 0.638 |
|
>25 | 24 (23.1) | 27.96±34.35 |
| 14.14±19.08 |
| 48.16±38.82 |
| 56.84±33.22 |
|
The data analysis was performed using the Cmeth
method and the EpiTect® PCR control DNA set (Qiagen).
The correlation of methylation status and miR-34 expression levels
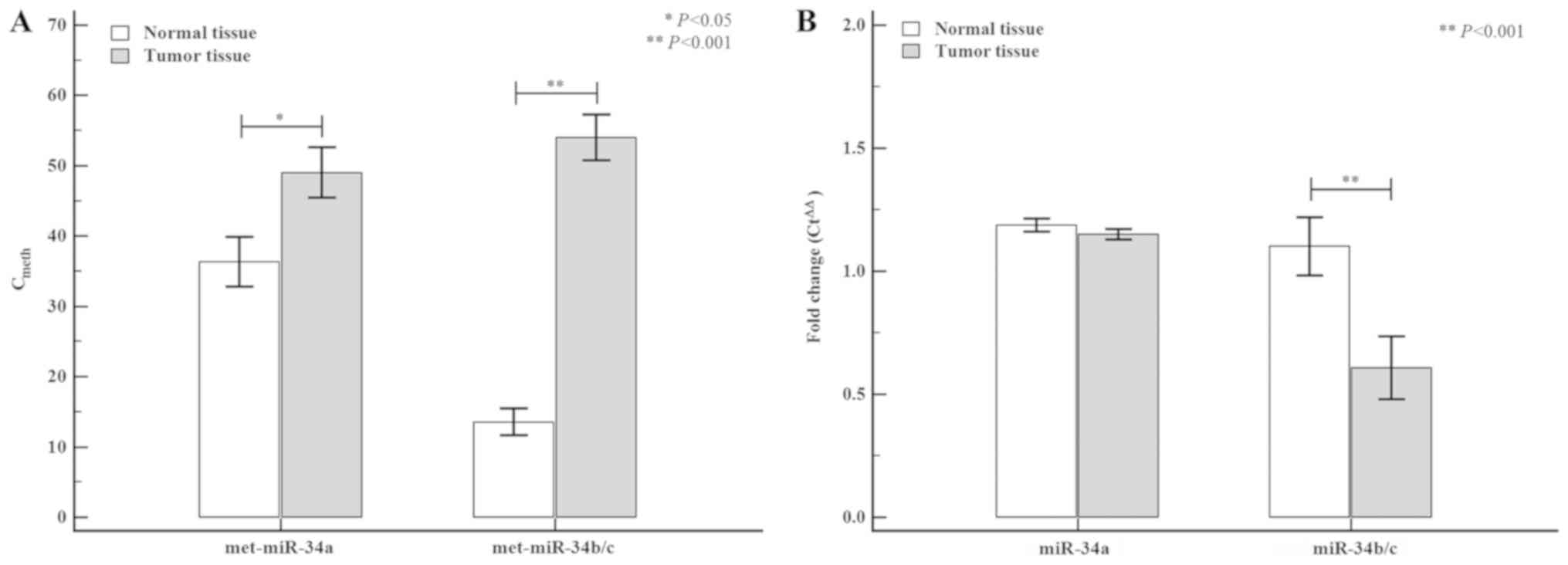
between normal and tumor tissue are shown in Fig. 2. The tumor tissue exhibited higher
miR-34a (normal tissue: 35.93±3.53 vs. tumor tissue: 48.99±3.57;
P=0.012) and miR-34b/c (normal tissue: 13.08±1.94 vs. tumor tissue:
55.63±3.35; P<0.0001) methylation levels than the normal tissue,
whereas differences in miR-34 miRNA expression levels between
normal and tumor tissues were only detected for miR-34b/c (normal
tissue: 1.10±1.19 vs. tumor tissue: 0.60±1.56; P=0.005). The
correlation of the methylation status of miR-34 miRNAs between
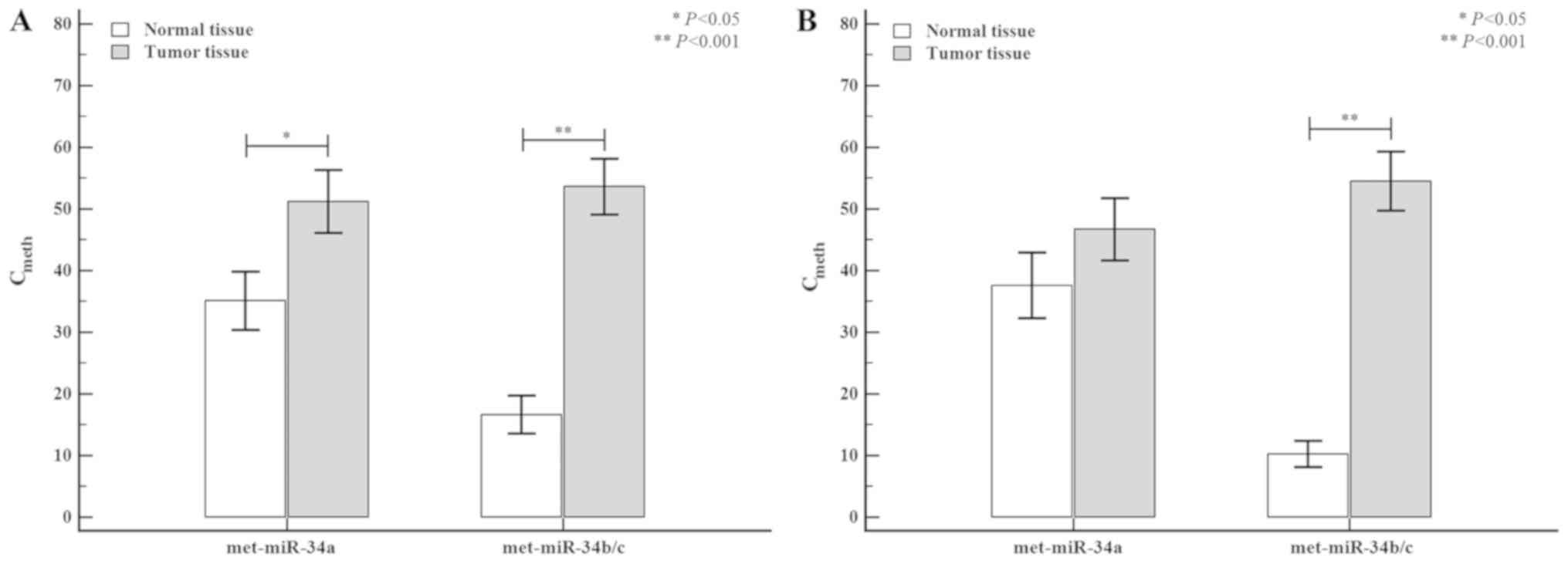
colon and rectal tumors is shown in Fig.
3. When the tumor tissue was stratified by tumor location
(colon or rectum), the methylation level of miR-34a was higher in
colon tumors (normal tissue: 35.66±4.78 vs. tumor tissue:
51.30±5.12; P=0.041) than in rectal tumors (normal tissue:
35.39±5.32 vs. tumor tissue: 45.24±5.10; P=0.141), but the
differences were not statistically significant. However, the
methylation level of miR-34b/c in colon tumors was significantly
higher (normal tissue: 16.48±3.12 vs. tumor tissue: 55.34±4.63;
P<0.0001) than the corresponding methylation levels observed in
rectal tumors (normal tissue: 10.07±2.23 vs. tumor tissue:
54.46±4.99; P<0.0001).
The gene polymorphism of TP53 showed a significant
correlation with the methylation of the miR-34a and −34b/c promoter
region in the analysis of the correlation between the polymorphisms
and DNA methylation levels (data not shown). Therefore, we were
performed that analysis of variance for the methylation amount of
miR-34a and miR-34b/c, the results of the TP53 and mir-34bc
polymorphism frequency analysis are shown in Table II. No statistically significant
differences in genotype frequencies were found when the normal and
tumor tissues were compared. However, the methylation levels of the
miR-34 miRNAs were significantly increased in the tumor tissues
compared with normal tissue. The miR-34a methylation levels in the
TP53 PIN A1A1 (48.56±36.49), TP53 MSP GG (49.00±36.44), and
miR-34b/c TC (51.61±35.64) genotypes were increased in the tumor
tissue compared with normal tissue; however, the methylation levels
of the miR-34 miRNAs were not significantly different between the
genotypes in each polymorphism. Interestingly, the miR-34b/c
methylation levels were significantly elevated in tumor tissue
compared with normal tissue across all of the polymorphisms, with
the miR-34b/c GG genotype (59.64±31.19; P<0.001) exhibiting the
highest degree of methylation (Table
II).
| Table II.TP53 and miR-34b/c genotype
frequencies and miR-34 methylation status in normal and tumor
tissues. |
Table II.
TP53 and miR-34b/c genotype
frequencies and miR-34 methylation status in normal and tumor
tissues.
|
|
| miR-34a | miR-34b/c |
|---|
|
|
|
|
|
|---|
| Genotypes | CRC (n=104), n
(%) | Normal tissue | Tumor tissue |
P-valueb | Normal tissue | Tumor tissue |
P-valueb |
|---|
| TP53 72
codon |
|
|
|
|
|
|
|
| GG | 46 (44.2) | 35.40±34.22 | 46.96±34.69 | 0.111 | 14.44±20.76 | 55.14±35.65 |
<0.001e |
| GC | 42 (40.4) | 31.58±36.90 | 43.82±38.06 | 0.138 | 14.55±21.09 | 54.60±32.81 |
<0.001e |
| CC | 16 (15.4) | 51.11±36.62 | 68.38±32.43 | 0.168 | 8.19±10.44 | 49.15±29.60 |
<0.001e |
|
P-valuea |
| 0.177 | 0.062 |
| 0.502 | 0.820 |
|
| TP53
PIN |
|
|
|
|
|
|
|
|
A1A1 | 96 (92.3) | 35.93±36.00 | 48.56±36.49 | 0.017c | 13.39±20.01 | 53.60±33.89 |
<0.001e |
|
A1A2 | 7
(6.7) | 39.83±40.48 | 51.91±40.01 | 0.585 | 16.42±16.60 | 55.58±29.02 |
0.009d |
|
A2A2 | 1
(1.0) | 44.14 | 69.38 |
| 6.00 | 81.30 |
|
|
P-valuea |
| 0.941 | 0.833 |
| 0.862 | 0.710 |
|
| TP53
MSP |
|
|
|
|
|
|
|
| GG | 95 (91.3) | 35.60±36.05 | 49.00±36.44 | 0.012c | 13.32±20.10 | 53.51±34.05 |
<0.001e |
| AG | 8
(7.7) | 43.24±38.70 | 46.32±40.27 | 0.878 | 16.86±15.42 | 56.41±26.97 |
0.003d |
| AA | 1
(1.0) | 44.14 | 69.38 |
| 6.00±0.00 | 81.30 |
|
|
P-valuea |
| 0.829 | 0.839 |
| 0.827 | 0.698 |
|
|
miR-34b/c |
|
|
|
|
|
|
|
| TT | 55 (52.9) | 41.55±38.35 | 46.73±36.61 | 0.471 | 14.50±19.90 | 59.64±31.19 |
<0.001e |
| TC | 41 (39.4) | 32.15±34.12 | 51.61±35.64 | 0.014c | 13.44±20.57 | 45.51±35.27 |
<0.001e |
| CC | 8
(7.7) | 21.11±20.91 | 51.14±42.82 | 0.096 | 7.26±12.72 | 58.76±33.27 |
0.001e |
|
P-valuea |
| 0.208 | 0.801 |
| 0.627 | 0.112 |
|
We confirmed the expression of miRNAs by genetic
polymorphism, unfortunately, we did not find any significant effect
on polymorphisms (Table III).
However, when the miRNA expression levels analyzed according to
haplotype of TP53 polymorphisms, we could confirm difference for
the expression of miR-34a and miR-34b/c (Table IV). In particular, the expression
pattern of miR-34b/c was increased in TP53 PIN/MSP haplotypes,
which were confirmed in both normal and tumor tissues. In addition,
this haplotype also confirmed the increased expression of miR-34a
in tumor tissues.
| Table III.TP53 and miR-34b/c genotype
frequencies and miR-34 family expression levels between normal and
tumor tissue. |
Table III.
TP53 and miR-34b/c genotype
frequencies and miR-34 family expression levels between normal and
tumor tissue.
|
|
| miR-34a | miR-34b/c |
|---|
|
|
|
|
|
|---|
| Genotypes | CRC (n=104) | Normal tissue | Tumor tissue |
P-valueb | Normal tissue | Tumor tissue |
P-valueb |
|---|
| TP53 72 codon |
|
|
|
|
|
|
|
| GG | 46 (44.2) | 1.16±0.24 | 1.14±0.19 | 0.801 | 0.92±1.13 | 0.68±1.50 | 0.386 |
| GC | 42 (40.4) | 1.22±0.30 | 1.14±0.22 | 0.201 | 1.09±1.18 | 0.73±1.03 | 0.150 |
| CC | 16 (15.4) | 1.19±0.18 | 1.19±0.25 | 0.992 | 1.64±1.35 | 0.14±0.99 | 0.001d |
|
P-valuea |
| 0.546 | 0.749 |
| 0.117 | 0.259 |
|
| TP53 PIN |
|
|
|
|
|
|
|
|
A1A1 | 96 (92.3) | 1.19±0.27 | 1.16±0.22 | 0.336 | 1.08±1.22 | 0.63±1.29 | 0.016c |
|
A1A2 | 7
(6.7) | 1.05±0.04 | 1.06±0.08 | 0.747 | 1.38±1.05 | 0.46±0.75 | 0.080 |
|
A2A2 | 1
(1.0) | 1.39 | 1.01 |
| 0.94 | −0.62 |
|
|
P-valuea |
| 0.258 | 0.394 |
| 0.807 | 0.586 |
|
| TP53 MSP |
|
|
|
|
|
|
|
| GG | 95 (91.3) | 1.19±0.26 | 1.16±0.22 | 0.436 | 1.09±1.22 | 0.62±1.29 | 0.011c |
| AG | 8
(7.7) | 1.13±0.23 | 1.06±0.07 | 0.430 | 1.21±1.08 | 0.66±0.89 | 0.280 |
| AA | 1
(1.0) | 1.39 | 1.01 |
| 0.94 | −0.62 |
|
|
P-valuea |
| 0.594 | 0.345 |
| 0.956 | 0.622 |
|
| miR-34b/c |
|
|
|
|
|
|
|
| TT | 55 (52.9) | 1.19±0.26 | 1.18±0.25 | 0.883 | 1.24±1.36 | 0.74±0.95 | 0.032c |
| TC | 41 (39.4) | 1.18±0.26 | 1.10±0.15 | 0.125 | 0.96±0.95 | 0.44±1.66 | 0.089 |
| CC | 8
(7.7) | 1.22±0.24 | 1.19±0.21 | 0.836 | 0.84±1.12 | 0.51±0.75 | 0.499 |
|
P-valuea |
| 0.948 | 0.241 |
| 0.435 | 0.532 |
|
| Table IV.Associations between miR-34 family
expression levels and the TP53 haplotype in normal and tumor
tissues. |
Table IV.
Associations between miR-34 family
expression levels and the TP53 haplotype in normal and tumor
tissues.
|
| miR-34a
expression | miR-34b/c
expression |
|---|
|
|
|
|
|---|
| Allele combination
model | Normal tissue | Tumor tissue |
P-valueb | Normal tissue | Tumor tissue |
P-valueb |
|---|
| TP53 codon
72/PIN/MSP |
|
|
|
|
|
|
|
G-A1-G | 1.36±1.88 | 2.29±3.42 | 0.015c | 2.53±3.62 | 2.56±5.35 | 0.963 |
|
G-A1-A | 2.30±2.24 | 4.96±5.34 | 0.288 | 6.32±6.84 | 11.45±15.95 | 0.485 |
|
C-A1-G | 1.25±2.13 | 2.17±2.94 | 0.069 | 2.88±3.60 | 3.41±7.10 | 0.639 |
|
C-A2-G | 2.25±2.50 | 5.22±5.93 | 0.331 | 6.25±7.64 | 10.71±17.71 | 0.619 |
|
P-valuea | 0.479 | 0.075 | | 0.031c | 0.003d |
|
| TP53 codon
72/PIN |
|
|
|
|
|
|
|
G-A1 | 1.41±1.90 | 2.43±3.56 | 0.008d | 2.75±3.93 | 3.09±6.64 | 0.664 |
|
C-A1 | 1.25±2.13 | 2.17±2.94 | 0.069 | 2.88±3.60 | 3.41±7.10 | 0.639 |
|
C-A2 | 1.87±2.14 | 4.43±5.02 | 0.239 | 4.97±6.61 | 7.74±15.33 | 0.669 |
|
P-valuea | 0.711 | 0.270 |
| 0.362 | 0.269 |
|
| TP53 codon
72/MSP |
|
|
|
|
|
|
|
G-G | 1.36±1.88 | 2.29±3.42 | 0.015c | 2.53±3.62 | 2.56±5.35 | 0.963 |
|
G-A | 2.30±2.24 | 4.96±5.34 | 0.288 | 6.32±6.84 | 11.45±15.95 | 0.485 |
|
C-G | 1.33±2.16 | 2.44±3.33 | 0.038c | 3.19±4.13 | 4.08±8.57 | 0.491 |
|
P-valuea | 0.694 | 0.338 |
| 0.124 | 0.024c |
|
| TP53 PIN/MSP |
|
|
|
|
|
|
|
A1-G | 1.32±1.96 | 2.25±3.26 | 0.002d | 2.65±3.60 | 2.85±6.00 | 0.727 |
|
A1-A | 2.30±2.24 | 4.96±5.34 | 0.288 | 6.32±6.84 | 11.45±15.95 | 0.485 |
|
A2-G | 2.25±2.50 | 5.22±5.93 | 0.331 | 6.25±7.64 | 10.71±17.71 | 0.619 |
|
P-valuea | 0.304 | 0.032c |
| 0.013c | 0.001e |
|
Discussion
CRC is an age-related disease and a multistage
process involving both genetic and epigenetic changes (43). Recently, several miRNA profiling
studies demonstrated that miRNAs are distinctively and
differentially expressed in cancer tissues compared with normal
adjacent tissues (44). Hence,
epigenetic mechanisms appear to influence the deregulation of
miRNAs in cancer pathogenesis (4).
The miR-34 miRNA family is comprised of miR-34a,
miR-34b, and miR-34c, which are encoded by two different genes.
Although miR-34a is encoded by its own transcript in chr.1p36.23,
miR-34b and miR-34c share a common primary transcript in chr.11q23.
The members of the miR-34 family are reportedly direct targets of
TP53, which induces apoptosis, cell cycle arrest, and senescence,
indicating a potential tumor suppressor role of the miR-34 miRNAs
(6,13). In particular, miR-34 miRNAs enable
TP53 to regulate numerous proteins, despite the prior synthesis of
their transcripts (6). As well,
several studies have shown that miR-34 miRNAs are silenced by CpG
methylation (45,46).
According to prior in vitro studies, miR-34a
overexpression causes the decreased proliferation and activation of
apoptosis in multiple tumor cells, indicating that miR-34a could
play a role in tumor suppression. In addition, miR-34a is
downregulated in several tumor types (7,17,19,47–49).
To date, over 77 miR-34 targets have been validated, including
factors that control the cell cycle (CDK4, CDK6, c-Myc, and E2F3),
regulators of apoptosis (BCL2, survivin, and CREB), proteins
involved in invasion (c-Met, AXL receptor, and the RAS-oncogene
homolog RRAS), factors related to epithelial mesenchymal transition
(EMT-inducing transcription factor SNAIL or the zinc finger 281
protein), proteins involved in the formation of cancer stem cells
(Notch1-4, WNT1, WNT3, β-catenin, and CD44), and factors that
regulate metabolism (hexokinase 1 and 2, glucose-6-phosphate
isomerase, pyruvate dehydrogenase kinase 1, and lactate
dehydrogenase A) (50,51). Consequently, the diverse roles of
members of the miR-34 family may lead to functional abnormalities
such as Wnt signaling, EMT, G1-arrest, or cancer cell progression,
leading to carcinogenesis (50). To
this end, recent reports suggest that aberrant miR-34b/c promoter
methylation is significantly correlated with the metastasis of
tumor cells to the lymph nodes (25,49).
Finally, miR-34b and miR-34c are directly regulated by promoter
hypermethylation and p53 in response to DNA damage or oncogenic
stress (14–16,25,52). In
fact, p53 regulates the expression of miR-34 via p53 binding sites
within miR-34a and miR-34b/c promoter regions. Consequently, the
antioncogenic action of miR-34 is regulated by p53.
In a previous study (53), we analyzed the association of SNPs of
miR-34b/c and TP53 Arg72Pro with the risk of colon cancer. The
previous study presented the association with risk of colon cancer
to the TP53 Arg72Pro CC genotype (53), and we considered that this
association may be related to the regulation of the miR-34 family
expression or methylation in cancer tissues. Therefore, in this
study, we showed that the methylation or expression of miR-34
miRNAs and the affected TP53 polymorphisms differ between
colorectal tumors and normal tissues. Specifically, the methylation
status of miR-34a and miR-34b/c was increased in tumor tissues
compared with the normal tissue. As well, we found that miR-34
miRNAs are downregulated in tumor tissues compared with paired
normal tissues and that this apparent downregulation is associated
with increased methylation of miR-34 miRNAs in colorectal tissues.
Furthermore, the haplotypes of TP53 polymorphisms (codon 72-PIN)
were shown to influence miR-34b/c expression level in normal and
cancer tissues. Interestingly, this is the first report of
association with expression and methylation by the SNPs, and this
results may be applicated for base data that functional research of
CRC. Although it has been established that p53 functions as a tumor
suppressor, reports also indicate an emerging role of p53 as an
important regulator of metabolic homeostasis, a critical aspect of
most major cellular processes (54,55). The
roles of p53 are fundamental for cell homeostasis and include
metabolic homeostasis, which safeguards against latent cancer, as
well as its more classical roles that include genome protection,
DNA repair, and programmed cell death (56). Critical mutations in the TP53
gene are common in most cancers and are major contributors to
cancer progression. Previous studies have likewise indicated the
regulation of the TP53 gene by TP53 haplotypes
(50,51,57). In
particular, studies have shown that the TP53 haplotype
containing the codon 72-PIN polymorphism affected TP53
function through the formation of various haplotypes (50,51), and
that TP53 polymorphisms were presented to linkage
disequilibrium (LD) blocks, which were formed many haplotypes and
affect to TP53 abnormality (57). In our previous study, we reported an
association between CRC risk and TP53 polymorphisms
combination model (53).
Furthermore, the TP53 haplotypes containing polymorphisms
(e.g., codon 72-PIN) influenced miR-34b/c expression levels in
normal and cancer tissues, suggesting altered regulation of
TP53. Interestingly, this is the first report of an
association between SNP expression and methylation; thus, these
results may be applicable to and serve as the foundation for future
CRC research.
In summary, we determined that the hypermethylation
of miR-34a and miR-34b/c is a relatively common event in CRC, and
that the methylation status affects miR-34 expression. We likewise
found that promoter methylation in miR-34 miRNAs and polymorphisms
in the TP53 codon 72 are a relatively common event associated with
CRC risk. Our study had several limitations. First, all patients
were selected from a single institution in Korea and the small
sample size could limit the statistical power. Second, further
studies are needed to define the relationship between DNA
hypermethylation and the pathogenesis of CRC.
Acknowledgements
Not applicable.
Funding
This study was partly supported by the National
Research Foundation of Korea (NRF) Grant funded by the Korean
Government (NRF-2018R1D1A1A09082764 and 2018R1D1A1B07047604) and
partly supported by the Korea Health Technology R&D Project
from the Ministry of Health and Welfare, Republic of Korea (grant
no. HI18C19990200).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JWK, KK and NKK conceived and designed the
experiments. KHL, JOK, HSP, CSR, JYL and DK performed the
experiments. HHJ, JOK, HSP, CSR and JYL analyzed the data and
performed statistical analyses. HHJ, KK, KHL, DK and NKK
contributed reagents/material/analysis tools. HHJ and KK wrote the
manuscript. JWK and NKK prepared the references and managed the
data. All authors reviewed the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of CHA Bundang Medical Center (Seongnam, Republic of
Korea; IRB no. 2009-08-077-010) and written informed consent was
provided by all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Roy S, Levi E, Majumdar AP and Sarkar FH:
Expression of miR-34 is lost in colon cancer which can be
re-expressed by a novel agent CDF. J Hematol Oncol. 5:582012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lodygin D, Tarasov V, Epanchintsev A,
Berking C, Knyazeva T, Körner H, Knyazev P, Diebold J and Hermeking
H: Inactivation of miR-34a by aberrant CpG methylation in multiple
types of cancer. Cell Cycle. 7:2591–2600. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
The Korea National Statistical Office
Report 2015, . Change in leading causes of death (2000–2015). Korea
National Statistical Office. http://kosis.kr/statHtml/statHtml.do?orgId=101&tblId=DT_1B34E13&vw_cd=MT_ZTITLE&list_id=D11&seqNo=&lang_mode=ko&language=kor&obj_var_id=&itm_id=&conn_path=MT_ZTITLEJanuary
26–2018
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Park Y, Lee J, Oh JH, Shin A and Kim J:
Dietary patterns and colorectal cancer risk in a Korean population:
A case-control study. Medicine (Baltimore). 95:e37592016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gallardo E, Navarro A, Viñolas N, Marrades
RM, Diaz T, Gel B, Quera A, Bandres E, Garcia-Foncillas J, Ramirez
J and Monzo M: miR-34a as a prognostic marker of relapse in
surgically resected non-small-cell lung cancer. Carcinogenesis.
30:1903–1909. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tarasov V, Jung P, Verdoodt B, Lodygin D,
Epanchintsev A, Menssen A, Meister G and Hermeking H: Differential
regulation of microRNAs by p53 revealed by massively parallel
sequencing: miR-34a is a p53 target that induces apoptosis and
G1-arrest. Cell Cycle. 6:1586–1593. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zamore PD and Haley B: Ribo-gnome: The big
world of small RNAs. Science. 309:1519–1524. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hutvágner G, McLachlan J, Pasquinelli AE,
Bálint E, Tuschl T and Zamore PD: A cellular function for the
RNA-interference enzyme Dicer in the maturation of the let-7 small
temporal RNA. Science. 293:834–838. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lund E, Güttinger S, Calado A, Dahlberg JE
and Kutay U: Nuclear export of microRNA precursors. Science.
303:95–98. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Davalos V and Esteller M: MicroRNAs and
cancer epigenetics: A macrorevolution. Curr Opin Oncol. 22:35–45.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ahmad A, Zhang W, Wu M, Tan S and Zhu T:
Tumor-suppressive miRNA-135a inhibits breast cancer cell
proliferation by targeting ELK1 and ELK3 oncogenes. Genes Genomics.
40:243–251. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bommer GT, Gerin I, Feng Y, Kaczorowski
AJ, Kuick R, Love RE, Zhai Y, Giordano TJ, Qin ZS, Moore BB, et al:
p53-mediated activation of miRNA34 candidate tumor-suppressor
genes. Curr Biol. 17:1298–1307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He L, He X, Lim LP, de Stanchina E, Xuan
Z, Liang Y, Xue W, Zender L, Magnus J, Ridzon D, et al: A microRNA
component of the p53 tumour suppressor network. Nature.
447:1130–1134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
He X, He L and Hannon GJ: The guardian's
little helper: microRNAs in the p53 tumor suppressor network.
Cancer Res. 67:11099–11101. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bagchi A, Papazoglu C, Wu Y, Capurso D,
Brodt M, Francis D, Bredel M, Vogel H and Mills AA: CHD5 is a tumor
suppressor at human 1p36. Cell. 128:459–475. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chang TC, Wentzel EA, Kent OA,
Ramachandran K, Mullendore M, Lee KH, Feldmann G, Yamakuchi M,
Ferlito M, Lowenstein CJ, et al: Transactivation of miR-34a by p53
broadly influences gene expression and promotes apoptosis. Mol
Cell. 26:745–752. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mori N, Morosetti R, Spira S, Lee S,
Ben-Yehuda D, Schiller G, Landolfi R, Mizoguchi H and Koeffler HP:
Chromosome band 1p36 contains a putative tumor suppressor gene
important in the evolution of chronic myelocytic leukemia. Blood.
92:3405–3409. 1998.PubMed/NCBI
|
|
20
|
Welch C, Chen Y and Stallings RL:
MicroRNA-34a functions as a potential tumor suppressor by inducing
apoptosis in neuroblastoma cells. Oncogene. 26:5017–5022. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hrašovec S and Glavač D: MicroRNAs as
novel biomarkers in colorectal cancer. Front Genet. 3:1802012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cho SH, Ko JJ, Kim JO, Jeon YJ, Yoo JK, Oh
J, Oh D, Kim JW and Kim NK: 3′-UTR polymorphisms in the miRNA
machinery genes DROSHA, DICER1, RAN, and XPO5 are associated with
colorectal cancer risk in a Korean population. PLoS One.
10:e01311252015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Toyota M, Suzuki H, Sasaki Y, Maruyama R,
Imai K, Shinomura Y and Tokino T: Epigenetic silencing of
microRNA-34b/c and B-cell translocation gene 4 is associated with
CpG island methylation in colorectal cancer. Cancer Res.
68:4123–4132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kozaki K, Imoto I, Mogi S, Omura K and
Inazawa J: Exploration of tumor-suppressive microRNAs silenced by
DNA hypermethylation in oral cancer. Cancer Res. 68:2094–2105.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lujambio A, Calin GA, Villanueva A, Ropero
S, Sánchez-Céspedes M, Blanco D, Montuenga LM, Rossi S, Nicoloso
MS, Faller WJ, et al: A microRNA DNA methylation signature for
human cancer metastasis. Proc Natl Acad Sci USA. 105:13556–13561.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kong YW, Ferland-McCollough D, Jackson TJ
and Bushell M: microRNAs in cancer management. Lancet Ooncol.
13:e249–e258. 2012. View Article : Google Scholar
|
|
27
|
Lujambio A, Ropero S, Ballestar E, Fraga
MF, Cerrato C, Setién F, Casado S, Suarez-Gauthier A,
Sanchez-Cespedes M, Git A, et al: Genetic unmasking of an
epigenetically silenced microRNA in human cancer cells. Cancer Res.
67:1424–1429. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Iwai N and Naraba H: Polymorphisms in
human pre-miRNAs. Biochem Biophys Res Commun. 331:1439–1444. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu Y, Liu L, Liu J, Zhang Y, Zhu J, Chen
J, Liu S, Liu Z, Shi H, Shen H and Hu Z: A potentially functional
polymorphism in the promoter region of miR-34b/c is associated with
an increased risk for primary hepatocellular carcinoma. Int J
Cancer. 128:412–417. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao LB, Li LJ, Pan XM, Li ZH, Liang WB,
Bai P, Zhu YH and Zhang L: A genetic variant in the promoter region
of miR-34b/c is associated with a reduced risk of colorectal
cancer. Biol Chem. 394:415–420. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hiroki E, Suzuki F, Akahira J, Nagase S,
Ito K, Sugawara J, Miki Y, Suzuki T, Sasano H and Yaegashi N:
MicroRNA-34b functions as a potential tumor suppressor in
endometrial serous adenocarcinoma. Int J Cancer. 131:E395–E404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bensen JT, Tse CK, Nyante SJ,
Barnholtz-Sloan JS, Cole SR and Millikan RC: Association of
germline microRNA SNPs in pre-miRNA flanking region and breast
cancer risk and survival: The carolina breast cancer study. Cancer
Causes Control. 24:1099–1109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dumont P, Leu JI, Della Pietra AC III,
George DL and Murphy M: The codon 72 polymorphic variants of p53
have markedly different apoptotic potential. Nat Genet. 33:357–365.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Irarrázabal CE, Rojas C, Aracena R,
Márquez C and Gil L: Chilean pilot study on the risk of lung cancer
associated with codon 72 polymorphism in the gene of protein p53.
Toxicol Lett. 144:69–76. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kuroda Y, Tsukino H, Nakao H, Imai H and
Katoh T: p53 codon 72 polymorphism and urothelial cancer risk.
Cancer Lett. 189:77–83. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rokni P, Shariatpanahi AM, Sakhinia E and
Kerachian MA: BMP3 promoter hypermethylation in plasma-derived
cell-free DNA in colorectal cancer patients. Genes Genomics.
40:423–428. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jones PA and Baylin SB: The epigenomics of
cancer. Cell. 128:683–692. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vogt M, Munding J, Grüner M, Liffers ST,
Verdoodt B, Hauk J, Steinstraesser L, Tannapfel A and Hermeking H:
Frequent concomitant inactivation of miR-34a and miR-34b/c by CpG
methylation in colorectal, pancreatic, mammary, ovarian,
urothelial, and renal cell carcinomas and soft tissue sarcomas.
Virchows Arch. 458:313–322. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Eads CA, Danenberg KD, Kawakami K, Saltz
LB, Blake C, Shibata D, Danenberg PV and Laird PW: MethyLight: A
high-throughput assay to measure DNA methylation. Nucleic Acids
Res. 28:E322000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cottrell S, Jung K, Kristiansen G, Eltze
E, Semjonow A, Ittmann M, Hartmann A, Stamey T, Haefliger C and
Weiss G: Discovery and validation of 3 novel DNA methylation
markers of prostate cancer prognosis. J Urol. 177:1753–1758. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Deng G, Kakar S and Kim YS: MicroRNA-124a
and microRNA-34b/c are frequently methylated in all histological
types of colorectal cancer and polyps, and in the adjacent normal
mucosa. Oncol Lett. 2:175–180. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fearon ER: Molecular genetics of
colorectal cancer. Annu Rev Pathol. 6:479–507. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Rinaldi A, Poretti G, Kwee I, Zucca E,
Catapano CV, Tibiletti MG and Bertoni F: Concomitant MYC and
microRNA cluster miR-17-92 (C13orf25) amplification in human mantle
cell lymphoma. Leuk Lymphoma. 48:410–412. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang Z, Chen Z, Gao Y, Li N, Li B, Tan F,
Tan X, Lu N, Sun Y, Sun J, et al: DNA hypermethylation of
microRNA-34b/c has prognostic value for stage I non-small cell lung
cancer. Cancer Biol Ther. 11:490–496. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li N, Fu H, Tie Y, Hu Z, Kong W, Wu Y and
Zheng X: miR-34a inhibits migration and invasion by down-regulation
of c-Met expression in human hepatocellular carcinoma cells. Cancer
Lett. 275:44–53. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tazawa H, Tsuchiya N, Izumiya M and
Nakagama H: Tumor-suppressive miR-34a induces senescence-like
growth arrest through modulation of the E2F pathway in human colon
cancer cells. Proc Natl Acad Sci USA. 104:15472–15477. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yan D, Zhou X, Chen X, Hu DN, Dong XD,
Wang J, Lu F, Tu L and Qu J: MicroRNA-34a inhibits uveal melanoma
cell proliferation and migration through downregulation of c-Met.
Invest Ophthalmol Vis Sci. 50:1559–1565. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Rokavec M, Li H, Jiang L and Hermeking H:
The p53/miR-34 axis in development and disease. J Mol Cell Biol.
6:214–230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hermeking H: The miR-34 family in cancer
and apoptosis. Cell Death Differ. 17:193–199. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Roman-Gomez J, Agirre X, Jiménez-Velasco
A, Arqueros V, Vilas-Zornoza A, Rodriguez-Otero P, Martin-Subero I,
Garate L, Cordeu L, San José-Eneriz E, et al: Epigenetic regulation
of microRNAs in acute lymphoblastic leukemia. J Clin Oncol.
27:1316–1322. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Oh J, Kim JW, Lee BE, Jang MJ, Chong SY,
Park PW, Hwang SG, Oh D and Kim NK: Polymorphisms of the
pri-miR-34b/c promoter and TP53 codon 72 are associated with risk
of colorectal cancer. Oncol Rep. 31:995–1002. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Itahana Y and Itahana K: Emerging roles of
p53 family members in glucose metabolism. Int J Mol Sci. 19(pii):
E7762018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gnanapradeepan K, Basu S, Barnoud T,
Budina-Kolomets A, Kung CP and Murphy ME: The p53 tumor suppressor
in the control of metabolism and ferroptosis. Front Endocrinol
(Lausanne). 9:1242018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Vousden KH: Functions of p53 in metabolism
and invasion. Biochem Soc Trans. 37:511–517. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Bellini I, Pitto L, Marini MG, Porcu L,
Moi P, Garritano S, Boldrini L, Rainaldi G, Fontanini G, Chiarugi
M, et al: DeltaN133p53 expression levels in relation to haplotypes
of the TP53 internal promoter region. Hum Mutat. 31:456–465. 2010.
View Article : Google Scholar : PubMed/NCBI
|