Introduction
The American Cancer Society projections for the
incidence of invasive breast cancer and mortality estimate 246,660
newly diagnosed invasive breast cancer cases and 40,450 invasive
breast cancer related deaths in women in 2018 (1). Therapy resistant invasive breast cancer
continues to remain the leading cause of mortality in women in the
USA. The triple negative breast cancer (TNBC) represents an
aggressive molecular subtype that lacks the expressions of estrogen
receptor-α (ER-α), progesterone receptor (PR) and human epidermal
growth factor receptor-2 (HER-2). TNBC is notable for its
resistance to conventional endocrine or HER-2 targeted therapy
(2–4). Current treatment options for the TNBC
subtype are restricted to anthracyclin, taxol and platinum based
conventional chemotherapy or to poly(ADP-ribose) polymerase (PARP),
phosphadityl-inositol-3 kinase (PI3K) and mammalian target of
rapamycin (m-TOR) selective inhibitor based targeted therapy
(4,5). These conventional and targeted
treatment options are frequently associated with long-term dose
limiting systemic toxicity, de novo or acquired tumor
resistance, and emergence of drug resistant cancer stem cells that
impact therapeutic efficacy, and thereby, promote drug resistant
disease progression (6). These
limitations emphasize a need to identify novel, non- toxic
treatment options as testable alternatives to existing treatment
strategies.
Complementary and alternative approaches utilizing
herbal medicines are being extensively used for general health
issues and for management of breast cancer. Nutritional herbs
represent widely used non-toxic natural substances in traditional
Chinese herbal medicine (7–9). Non-toxic natural substances offer a
testable alternative as potentially novel approach for secondary
prevention/therapy of breast cancer, reduction of therapy
associated toxicity and/or for enhanced therapeutic efficacy
(8,9). Extracts from mechanistically distinct
Chinese nutritional herbs have displayed growth inhibitory effects
on cell culture models for Luminal A and the triple-negative
molecular subtypes for clinical breast cancer (10–17).
Cornus officinalis (CO) is a nutritional herb
in the form of a cherry fruit. It is used as a major ingredient in
some Chinese herbal formulations for general health management
purposes. Several Cornus species have documented
anti-proliferative, anti-oxidant and anti-inflammatory properties.
Anthocyanins are among the major bio-active agents (18,19).
Additionally, CO extract exhibits growth inhibitory effects on the
estrogen receptor positive Luminal A model predominantly via
inhibiting estrogen stimulated growth and altering the cellular
metabolism of estradiol to favor the generation of
anti-proliferative metabolites (12). In an effort to evaluate the utility
of CO in the TNBC model, the experiments in the present study were
designed to examine the growth inhibitory effects of CO in a cell
culture model for TNBC, and to identify potential mechanistic leads
and molecular targets for its efficacy.
Materials and methods
Experimental model
The human mammary carcinoma derived
ER-α−, PR− and HER-2− MDA-MB-231
cells (20,21) represented the model for TNBC. The
MDA-MB-231 cells were obtained from American Type Culture
Collection (ATCC; Manassas, VA, USA) and were maintained in RPMI
medium with L-glutamine and 5% fetal bovine serum (Life
Technologies, Grand Island, NY, USA) following the protocol
recommended by the vendor.
Dose response
For the dose response experiments the
non-fractionated aqueous extract of CO was prepared following
previously published protocol (12).
Briefly, the herb was sequentially boiled in water and centrifuged
to concentrate their water soluble components in the final volume
of 20 ml. This 100% stock solution of CO was serially diluted in
the culture medium to obtain the final concentration range of 1.0,
0.5, 0.1 and 0.05% CO. The cells were treated with the CO extract
within this concentration range. The dose response of CO extract
was determined by the cell viability assay using the trypan blue
dye exclusion test kit (Sigma-Adrich; Merck KGaA, Darmstadt,
Germany). The cell viability measurements were conducted for CO
treated and control cultures at day 7 post-seeding of
1.0×105 cells. The cell viability was calculated as %
viable cells=[1.00-(Number of blue cells ÷ Number of total cells)]
×100. These data were used to identify minimum effective, half
maximum, maximum cytostatic and toxic concentrations. The maximum
cytostatic concentration was defined by the cell viability equal to
or higher than the initial seeding density. The toxic concentration
was defined as the cell viability lower than the initial seeding
density. The data were expressed as viable cell number
(×105), relative to the initial seeding density.
Anchorage independent (AI) growth
assay
This assay was performed following the optimized
protocol (13,16). The stock solution of agar was
prepared by mixing DNA grade agar (Sigma-Adrich; Merck KGaA) with
an appropriate volume of 2X RPMI medium (Sigma-Adrich; Merck KGaA)
to obtain a 6% agar stock solution. To prepare the basement layer,
this stock solution was diluted to 0.6% using the culture medium,
dispersed in a 6-well plate and allowed to solidify overnight at
37°C. MDA-MB-231 cell suspension, at a density of 5×105
cells per ml, was prepared in RPMI medium containing 0.33% agar,
and this cell suspension was overlaid on the basement layer in the
presence or absence of CO. The cultures were incubated at 37°C in a
CO2 incubator for 21 days. The AI colonies were stained
with 0.005% crystal violet and colony counts were determined at 10X
magnification. The data were expressed as AI colony number.
Cell cycle progression
For the analysis of cell cycle progression,
5×104 cells were seeded in T-25 flasks and treated at 24
h. post-seeding with different concentrations of CO for 48 h. The
cells were harvested by trypsinization, pelleted at 500 × g, and
washed twice with cold phosphate buffered saline pH 7.4 (PBS;
Sigma-Adrich; Merck KGaA). The cells were then fixed with cold 70%
ethanol, washed with cold PBS, and stained with 50 µg/ml propidium
iodide (PI; Sigma-Adrich; Merck KGaA) in PBS, followed by the
addition of 10 µg/ml ribonuclease (Sigma-Adrich; Merck KGaA) and
incubation for 4 h. in the dark. DNA content was analyzed by flow
cytometry using Becton Dickinson FACSCAN Flow Cytometer (BD
Biosciences, Research Triangle Park, NC, USA) and analyzed with
FACS Express software (De Novo Software, Glendale, CA, USA). The
data were expressed as G1:S+G2/M ratio.
Western blot analysis
For the western blot assay, cells were seeded in
10-cm dishes at 70% confluence 1 day before the treatment.
Different concentrations of CO were added and incubated for 48 h.
in a CO2 incubator at 37°C. Cells were harvested and
immediately lysed with radio-immunoprecipitation assay (RIPA)
buffer containing protease inhibitors (Sigma-Adrich; Merck KGaA),
and centrifuged for 15 min at 10,000 × g. An equal quantity of
protein was separated by 10% SDS-PAGE and transferred onto a
nitrocellulose membrane (Bio-Rad Laboratories, Hercules, CA., USA)
and was blocked for 1 h with 5% nonfat dry milk followed by
incubation with primary and secondary antibodies. The antibodies
against the following proteins were used: β-actin and cyclin D1
(Santa Cruz Biotechnologies, Santa Cruz, CA, USA) and phospho-RB
(Ser 780, Cell Signaling Technology, Inc. Danvers, CA, USA). The
antibodies were used following the protocol provided by the
vendors. The chemo-luminescent signal was developed with ECL-plus
reagent (Bio-Rad Laboratories), and detected by autoradiography.
The data were expressed as arbitrary scanning unit (ASU).
Reverse transcription-polymerase chain
reaction assay for gene expression
The effect of CO on the status of select apoptosis
specific gene expression was evaluated using the semi-quantitative
Reverse Transcription PCR assay. Gene amplification was monitored
for apoptosis specific BAX and BCL-2 genes using gene specific
promoters (Cepheid, Inc. Sunnyvale, CA, USA). Briefly, a total of
25 µl of reaction mix was prepared that contained MgCl2
(2 mmol/l), 12.5 µl of 2X Taq PCR Master Mix (Qiagen, Inc.,
Valencia, CA, USA), 0.25X SYBR dye (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), and gene specific primer sets (0.3 µmol/l,
provided by the core facility, University of Texas Health Sciences
Center, San Antonio, TX, USA). The PCR reaction was set for 40
cycles and the data were compared after normalization with β-actin
RNA levels. The data were expressed as ∆∆Ct values for relative
gene expression.
Caspase assay
Caspase-3/7 activity in the MDA-MB-231 cells was
measured using Caspase-Glo assay kit (Promega Life Sciences, Inc.,
Madison, WI, USA). Briefly, the cells treated with CO were
homogenized by sonication in homogenization buffer (25 mmol/l
HEPES, pH 7.5, 5 mmol/l MgCl2, and 1 mmol/l EGTA) and
protease inhibitors (all from Sigma-Adrich; Merck KGaA). The
homogenate was centrifuged at 6,500 × g at 4°C for 15 min. To 10 µl
of the supernatant containing the cellular protein, equal volume of
the assay reagent was added and incubated at room temperature for 2
h. The luminescence was measured using Luminometer (Thermo Fisher
Scientific, Inc.). The data were expressed as relative luminescence
units (RLU).
Statistical analysis
The experiments for dose response, anchorage
independent growth, caspase-3/7 activity and relative gene
expression profile were conducted in triplicate. The data are
expressed as the mean ± standard deviation. Statistically
significant differences between the control and multiple treatment
groups were assessed by one-way analysis of variance and Dunnett's
test as a post hoc test with a threshold of α=0.05, using Microsoft
Excel 2013 XLSTAT-Base software.
Results
Growth characteristics of MDA-MB-231
cells
The data presented in Table I summarizes the growth pattern of the
experimental model. The MDA-MB-231 cells exhibited a short
population doubling time of 15 h and high saturation density,
exhibiting a ~33-fold increase relative to the initial seeding
density. Additionally, these cells exhibited accelerated cell cycle
progression as evidenced by a substantially low
G1:S+G2/M ratio of 0.6, relative to a ratio
of 2.3 exhibited by non-tumorigenic triple-negative 184-B5 cells
(data not shown). Furthermore, unlike the non-tumorigenic 184-B5
cells, these breast carcinoma derived MDA-MB-231 cells exhibited a
high number of AI colonies.
 | Table I.Status of proliferation end points in
a cellular model for triple-negative breast cancer. |
Table I.
Status of proliferation end points in
a cellular model for triple-negative breast cancer.
| Proliferation end
point | Model
MDA-MB-231 |
|---|
| Doubling time
(h)a | 15.0±2.2 |
| Saturation density
(×105)b | 32.9±2.3 |
|
G1:G2/M
ratioc | 0.6±0.3 |
| AI colony
numberd | 257±57 |
Dose response of CO
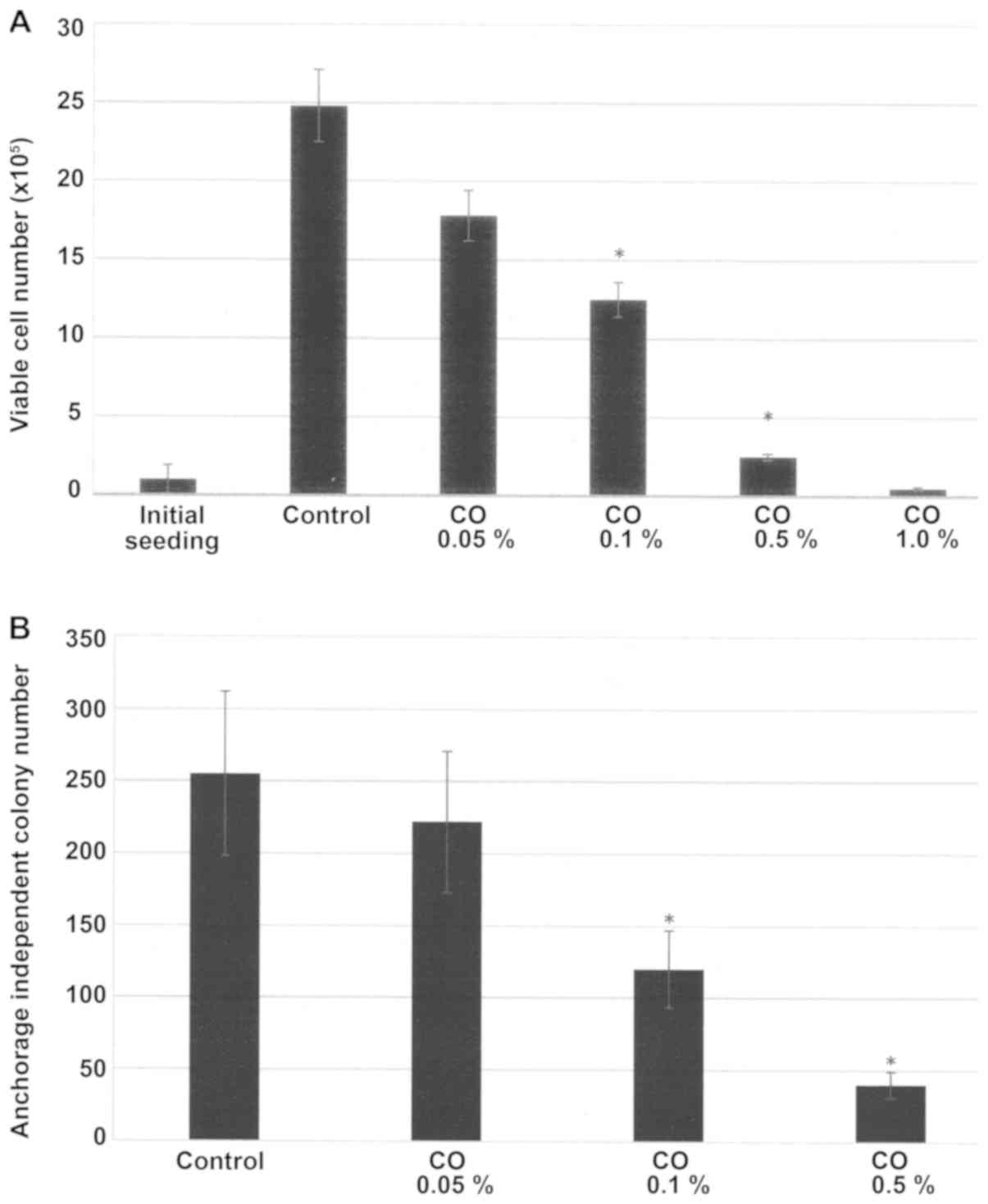
The data presented in Fig. 1A demonstrate a dose-dependent growth
inhibition of MDA-MB-231 cells in response to a 7-day treatment
with CO. In response to treatment with CO concentrations of 0.05,
0.1 and 1.0%, the viable cell number was reduced to
17.8±1.6×105, 12.5±1.1×105 and
2.5±0.2×105, respectively, relative to 24.8±2.3
×105 in the control. This dose response experiment
identified IC25 as 0.05%, IC50 as 0.1% and
IC90 as 0.5% for CO respectively, relative to control.
Statistical analysis using ANOVA and Dunnett's test confirmed that
control >IC50 and control >IC90
(α=0.05). The concentration of 1% CO exhibited >98% inhibition
in viable cell number. In this treatment group the viable cell
number was lower than the initial seeding density of 1.0
×105. The concentration of 1% was therefore considered
toxic.
Effect of CO on anchorage independent
(AI) growth
The data presented in Fig. 1B summarizes the growth inhibitory
effect of CO on AI colony formation of 21 day duration in
MDA-MB-231 cells. In response to treatment with CO at the
pre-determined IC25, IC50 and IC90
concentrations, the number of AI colonies was reduced to 222±49,
120±27 and 40±9, respectively, relative to 255±57 AI colonies in
the control. Statistical analysis using ANOVA and Dunnett's test
confirmed that control >IC50 and control
>IC90 (α=0.05).
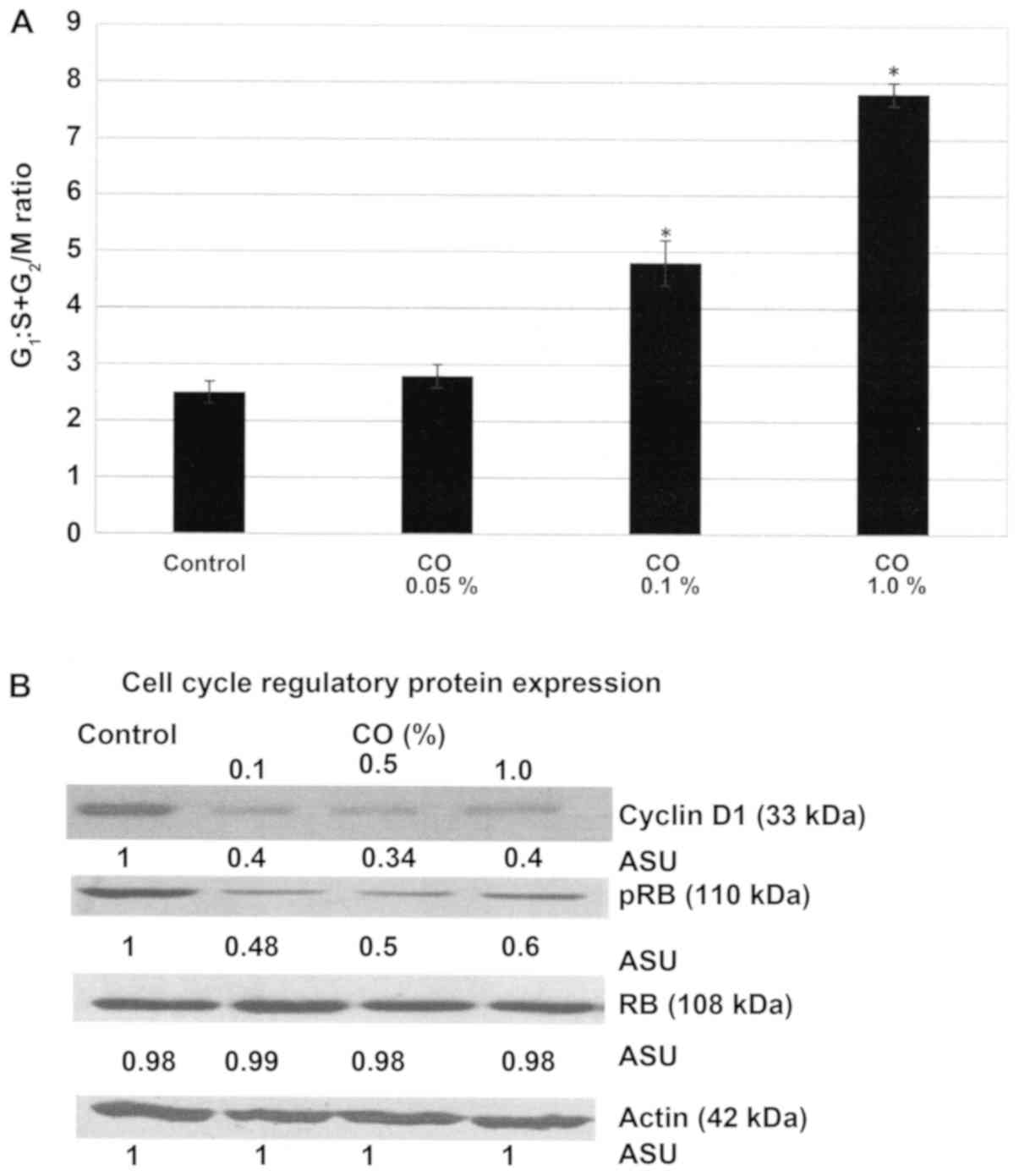
Effect of CO on cell cycle
progression
The experiment presented in Fig. 2A examined the effect of CO on the
cell cycle progression of MDA-MB-231 cells. The data presented as
the G1:S+G2/M ratio demonstrate that a 48 h.
treatment with CO was associated with a dose-dependent increase in
this ratio. Thus, treatment with 0.1% CO exhibited the
G1:S+G2/M ratio of 4.8±0.4, and treatment
with 1.0% CO exhibited a ratio of 7.8±0.2, relative to the ratio of
2.5±0.2 exhibited by control. Statistical analysis using ANOVA and
Dunnett's test confirmed that control <0.1% CO and control
<1.0% CO (α=0.05). These data indicate that CO treatment induced
a 92% and a 2.2-fold progressive increase due to G1
arrest and inhibition of the S and G2/M phases of the
cell cycle.
The experiment presented in Fig. 2B examines the status of select
regulatory proteins in the RB signaling pathway in response to
treatment with CO. These data demonstrate that a 48 h. CO treatment
resulted in ~60–66% decrease in cyclin D1 expression, and ~40–52%
decrease in p-RB expression depending on CO concentration, relative
to that in the untreated control cells.
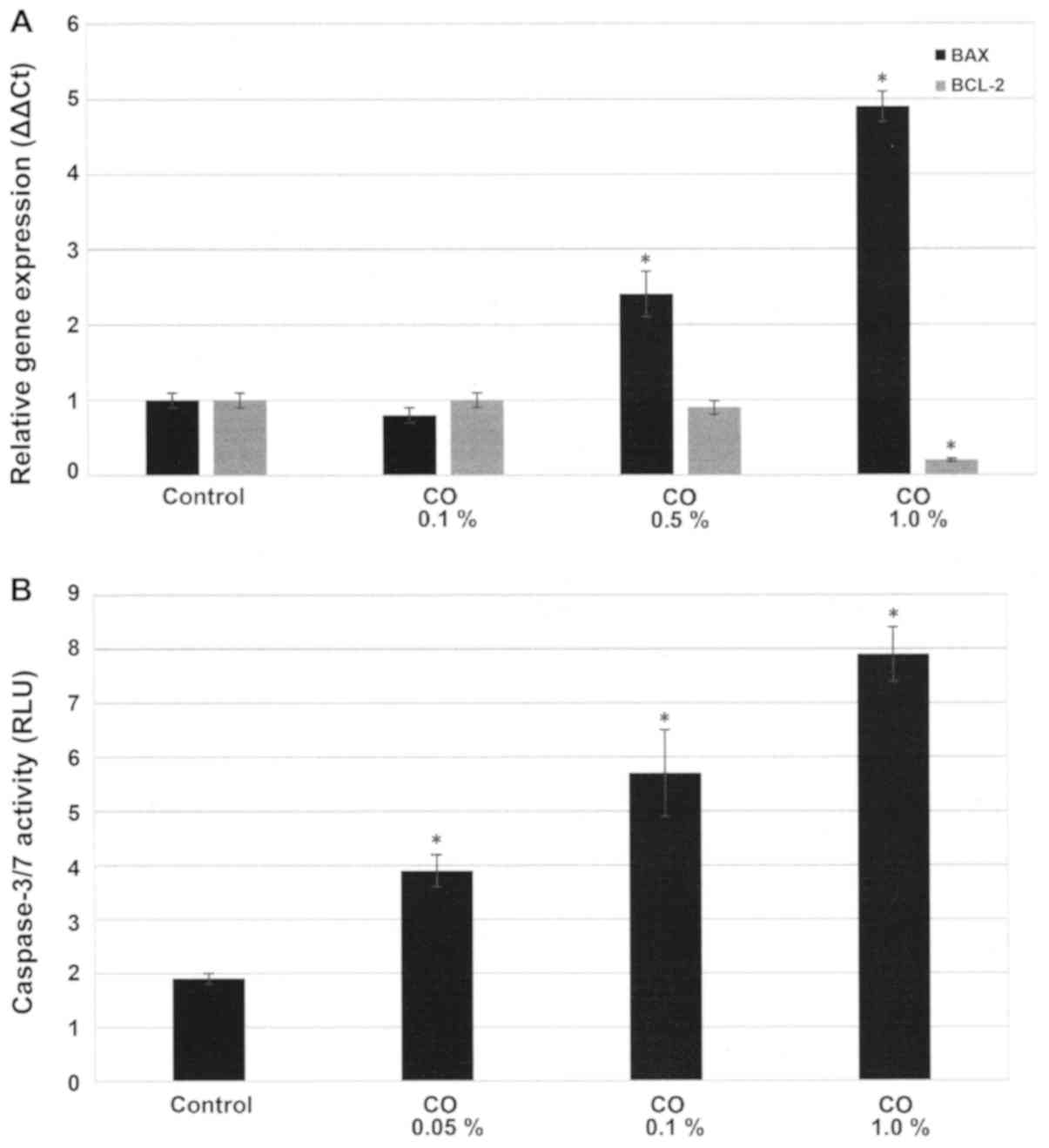
Effect of CO on cellular
apoptosis
The experiment presented in Fig. 3A examines the status of apoptosis
specific BAX and BCL-2 gene expressions. These data presented as
∆∆Ct values demonstrate that in response to a 48 h. treatment with
0.5% CO and 1.0% CO the expression of pro-apoptotic BAX gene was
2.5±0.2 and 4.9±0.5, while that of anti-apoptotic BCL-2 gene was
0.9±0.1 and 0.2±0.1, relative to untreated control. Statistical
analysis using ANOVA and Dunnett's test confirmed that control
<0.5% CO and control <1.0% CO for BAX expression (α=0.05),
and control >1.0% CO for BCL-2 expression (α=0.05). Thus, these
data demonstrate that in response to treatment with CO the
expression of pro-apoptotic BAX gene exhibited a ~2.5- and
~2.9-fold increase, while that of anti-apoptotic BCL-2 gene
exhibited a ~10 and ~80% decrease, relative to the untreated
control.
The experiment presented in Fig. 3B examined the effect of a 48 h.
treatment with CO on pro-apoptotic caspase-3/7 activity. The data
presented as RLU demonstrate that treatment with 0.1% CO resulted
in RLU value of 5.7±0.3, and treatment with 1.0% CO resulted in RLU
value of 7.9±0.5, relative to RLU value of 1.9±0.1 for the control.
Statistical analysis using ANOVA and Dunnett's test confirmed that
control <0.1% CO and control <1.0% CO (α=0.05). Thus,
treatment with CO induced a 2.0- and 3.1-fold increase in
caspase-3/7 activity.
Discussion
Approximately 10–20% of clinical breast cancers lack
the expressions of ER-α, PR and HER-2 and are classified as TNBC
(2,4,22,23). The
treatment option for the TNBC molecular subtype is predominantly
restricted to conventional chemotherapy or to pathway selective
small molecule based targeted therapy (3–5).
Long-term therapeutic efficacy of these treatment options is
compromised predominantly due to acquired tumor resistance and
emergence of drug resistant cancer stem cell population (6,22–24).
These limitations emphasize a need for identification of non-toxic
effective alternatives as testable therapeutic options.
Human tissue derived cell culture models offer
valuable mechanistic approaches to identify clinically translatable
leads to the inhibitory efficacy of a number of nutritional herbs
against the Luminal A and TNBC models (10–17).
Compared to the non-tumorigenic triple-negative 184-B5 cells, the
present TNBC model exhibits loss of homeostatic growth control and
persistent cancer risk as evidenced by hyper-proliferation,
accelerated cell cycle progression and increased AI colony
formation. In the present TNBC model several nutritional herbs
induce cytostatic growth arrest and/or cellular apoptosis and
inhibit anchorage independent colony formation, thereby
re-establishing cellular homeostasis and reducing cancer risk. At
the mechanistic levels, growth inhibitory efficacy of these herbs
involve RB, BAX/BCL-2, caspase, and RAS-RAF-MEK-ERK mediated
signaling pathways (15–17).
In the present study, MDA-MB-231 cells in response
to treatment with CO exhibited dose-dependent growth inhibitory
effects as evidenced by cytostatic growth arrest, and reduction in
AI colony formation. The data generated from dose response
experiments identified the effective range of 0.05–0.5% for CO in
the present model, suggesting potent growth inhibitory efficacy at
relatively low doses of CO. Additionally, the cytostatic growth
arrest induced by CO was associated with a progressive increase in
the G1:S+G2/M ratio, indicating an effect of
CO on cell cycle progression.
The data from the experiment designed to examine the
effect of CO on the RB pathway clearly demonstrate that CO
inhibited the expression of cyclin D1 and of phosphorylated RB
(p-RB) in a dose-dependent manner. The tumor suppressor
retinoblastoma (RB) gene has a well-documented function in the
regulation of cell cycle progression. The tumor suppressive
function of RB via the cyclin D-CDK4/6-pRB pathway has been
documented to be frequently compromised in therapy resistant
basal-like and triple-negative molecular subtypes of clinical
breast cancer and therefore, may represent a therapeutic target
(25–27). Additionally, small molecule
inhibitors of CDK4/CDK6 in combination with aromatase inhibitors or
with selective estrogen receptor modulators have documented
clinical efficacy in hormone receptor positive metastatic breast
cancer (28,29). Specific small molecule inhibitors of
CDK4/CDK6 may also represent viable treatment options for TNBC
(30,31). In the present study, the
dose-dependent decrease in cyclin D1 and pRB expression suggests
involvement of the RB pathway in the efficacy of CO. In this
context it is noteworthy that kinase-dependent site specific
phosphorylation of RB represents one of the major
post-translational modification that is critical for its tumor
suppressive function via inactivation of cell cycle progression and
promotion of cellular apoptosis (25,26). It
is also noteworthy that another nutritional herb Dipsacus
asperoides exhibits inhibition of CDK4 and CDK6 expression in
the MDA-MB-231 model for TNBC (17).
Additionally, recent evidence on human TNBC samples and on patient
derived TNBC xenograft models suggests that expression of EGFR,
it's recently identified partner MT4-MMP and RB is strongly
associated with the efficacy of a combination of EGFR and CDK4/6
based targeted therapy (32). Thus,
the present data on inhibition of cyclin D1 and pRB provide
mechanistic leads that the RB pathway might represent a molecular
target for the efficacy of CO in the present model system.
The experiment designed to examine the effect of CO
on cellular apoptosis demonstrated that treatment with CO produced
a progressive increase in pro-apoptotic BAX expression, while that
of anti-apoptotic BCL-2 was decreased. Additionally, this
CO-mediated modulation in the BAX/BCL-2 pathway positively
correlated with a dose-dependent increase in the pro-apoptotic
caspase-3/7 activity. It is well established that the intrinsic
mitochondrial apoptotic pathway involves altered membrane
permeability, cytochrome-c release, and apoptosome mediated
activation of caspase-9 and subsequently of caspase-3/7 (33,34). In
the present TNBC model, treatment with CO has identified
pro-apoptotic mechanistic leads for the efficacy of CO (16). Thus collectively, these data on
cellular apoptosis support the evidence that induction of cellular
apoptosis by CO may be via a caspase-dependent mechanism. This
suggestive mechanistic lead indicates involvement of the intrinsic
apoptotic cascade where expression of caspase-3/7 activity
represents a late-occurring event. Investigating additional
molecular pathways relevant to intrinsic apoptosis cascade
represents future research directions that are focused to examine
the specific effect of CO on cellular apoptosis and to investigate
mechanisms and molecular targets responsible for pro-apoptotic
efficacy of CO.
Traditional Chinese herbal medicine utilizes
combination of several herbs for herbal formulations that are
boiled in water to prepare herbal tea for consumption by the
patients. Thus, non-fractionated aqueous extract of CO used in the
present study simulates clinical administration of herbal
formulations. In this context it is conceivable that individual
constitutive herbs in the formulations may contain multiple
bio-active agents with distinct growth modulatory roles. Although
in the present study, non-fractionated aqueous extract from CO has
exhibited potent growth inhibitory effects via potential
mechanistic leads for its efficacy, little evidence is available
regarding the identity of active agent(s) responsible for these
effects. However, it is noteworthy that anthocyanins, representing
major bio-active agents, may in part be responsible for the
cellular and biochemical effects of CO (18,19).
In conclusion, the present study outcome validates a
relevant cell culture model for the triple-negative molecular
subtype of clinical breast cancer, and offers a facile experimental
approach to prioritize efficacious natural substances as testable
alternatives for endocrine therapy resistant breast cancer.
Acknowledgements
Part of the data included in the present study has
been previously presented at the San Antonio Breast Cancer
Symposium, December 2015. (https://cancerres.aacrjournals.org/content/76/4_Supplement/P3-09-04)
(16).
Funding
Principal funding support for this research was
provided by the philanthropic contributions to the American
Foundation for Chinese Medicine by the Randall and Barbara Smith
Foundation, and the Sophie Stenbeck Family Foundation.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NTT conceived the study design, formulated the
experimental protocols and prepared the manuscript. HBN performed
the experiments, and organized and analyzed the data. GYCW selected
the nutritional herb for the present study, and contributed to the
data interpretation and preparation of the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
American Cancer Society, . Facts &
Figures. American Cancer Society Inc. Atlanta, GA: 2017.
|
|
2
|
Sørlie T, Perou CM, Tibshirany R, Aas T,
Geisler S, Johnsen H, Hastie T, Eisen MB, van de Rijn M, Jeffrey
SS, et al: Gene expression patterns of breast carcinomas
distinguish tumor subclasses with clinical implications. Proc Natl
Acad Sci USA. 98:10869–10874. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baselga J and Swain SM: Novel anticancer
targets: Revisiting ERBB2 and discovering ERBB3. Nat Rev Cancer.
9:463–475. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dinh P, Satirou C and Piccart MJ: The
evaluation of treatment strategies: Aiming at the target. Breast.
16 (Suppl 2):S10–S16. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Anders CK, Winer EP, Ford JM, Dent R,
Silver DP, Siedge GW and Carey LA: Poly(ADP-ribose) polymerase
inhibition: ‘Targeted’ therapy for triple-negative breast cancer.
Clin Cancer Res. 16:4702–4710. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dean M, Fojo T and Bates S: Tumor stem
cells and drug resistance. Nat Rev Cancer. 5:275–284. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tindle HA, Davis RB, Phillips RS and
Eisenburg DM: Trends in the use of complementary and alternative
medicine by US adults: 1997–2002. Altern Ther Health Med. 11:42–49.
2005.PubMed/NCBI
|
|
8
|
Molassiotis A, Scott JA, Kearney N, Pud D,
Magri M, Selvekerova S, Bruyns I, Fernadez-Ortega P, Panteli V,
Margulies A, et al: Complementary and alternative medicine use in
breast cancer patients in Europe. Support Care Cancer. 14:260–267.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Helyer LK, Chin S, Chui BK, Fitzgerald B,
Verma S, Rakovitch E, Dranitsaris G and Clemons M: The use of
complementary and alternative medicines among patients with locally
advanced breast cancer - a descriptive study. BMC Cancer. 6:39–46.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mukherjee B, Telang N and Wong GY: Growth
inhibition of estrogen receptor positive human breast cancer cells
by Taheebo from the inner bark of Tabebuia avellanedae tree.
Int J Mol Med. 24:253–260. 2009.PubMed/NCBI
|
|
11
|
Li G, Sepkovic DW, Bradlow HL, Telang NT
and Wong GY: Lycium barbarum inhibits growth of estrogen
receptor positive human breast cancer cells by favorably altering
estradiol metabolism. Nutr Cancer. 61:408–414. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Telang NT, Li G, Sepkovic DW, Bradlow HL
and Wong GY: Anti-proliferative effects of Chinese herb Cornus
officinalis in a cell culture model for estrogen
receptor-positive clinical breast cancer. Mol Med Rep. 5:22–28.
2012.PubMed/NCBI
|
|
13
|
Telang N, Li G, Sepkovic D, Bradlow HL and
Wong GY: Comparative efficacy of extracts from Lycium
barbarum bark and fruit on estrogen receptor positive human
mammary carcinoma MCF-7 cells. Nutr Cancer. 66:278–284. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Telang N, Li G, Katdare M, Sepkovic D,
Bradlow L and Wong G: Inhibitory effects of Chinese nutritional
herbs in isogenic breast carcinoma cells with modulated estrogen
receptor function. Oncol Lett. 12:3949–3957. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Telang NT, Nair HB and Wong GY: Efficacy
of Tabebuia avellanedae extract on a cell culture model for
triple negative breast cancer. Cancer Res. 74 Suppl:SABCS,
P5-14-02. 2014.
|
|
16
|
Telang N, Nair HB and Wong GY: Effect of
Cornus officinalis (CO) on a model for triple negative
breast cancer. Cancer Res. 75 Suppl:SABCS, P3-09-04. 2015.
|
|
17
|
Telang N, Nair HB and Wong GY: Efficacy of
Dipsacus asperoides (DA) in a model for triple negative
breast cancer. Cancer Res. 77 (Suppl 4):P4-13-04-P4-13-04. 2017.
View Article : Google Scholar
|
|
18
|
Seeram NP, Schutzki R, Chandra A and Nair
MG: Characterization, quantification, and bioactivities of
anthocyanins in Cornus species. J Agric Food Chem.
50:2519–2523. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chang JS, Chiang LC, Hsu FF and Lin CC:
Chemoprevention against hepatocellular carcinoma by Cornus
officinalis in vitro. Am J Chin Med. 32:717–725. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Neve RM, Chin K, Fridlyand J, Yeh J,
Baehner FL, Fevr T, Clark L, Bayani N, Coppe JP, Tong F, et al: A
collection of breast cancer cell lines for the study of
functionally distinct cancer subtypes. Cancer Cell. 10:515–527.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Subik K, Lee JF, Baxter L, Strzepak T,
Costello D, Crowley P, Xing L, Hung MC, Bonfiglio T, Hicks DG and
Tang P: The expression patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67
and AR by immunohistochemical analysis in breast cancer cell lines.
Breast Cancer (Auckl). 4:35–41. 2010.PubMed/NCBI
|
|
22
|
Hudis CA and Gianni L: Triple-negative
breast cancer: An unmet medical need. Oncologist. 16 (Suppl
1):S1–S11. 2011. View Article : Google Scholar
|
|
23
|
Lin NU, Vanderplas A, Hughes ME, Theriault
RL, Edge SB, Wong YN, Blayney DW, Niland JC, Winer EP and Weeks JC:
Clinicopathologic features, patterns of recurrence, and survival
among women with triple-negative breast cancer in the National
Comprehensive Cancer Network. Cancer. 118:5463–5472. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Telang N: Putative cancer-initiating stem
cells in cell culture models for molecular subtypes of clinical
breast cancer. Oncol Lett. 10:3840–3846. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cox LA, Chen G and Lee EY: Tumor
suppressor genes and their role in breast cancer. Breast Cancer Res
Treat. 32:19–38. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Burkhart DL and Sage J: Cellular
mechanisms of tumor suppression by the retinoblastoma gene. Nat Rev
Cancer. 8:671–682. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bosco EE and Knudson ES: RB in breast
cancer: At the crossroads of tumorigenesis and treatment. Cell
Cycle. 6:667–671. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sherr CJ and Roberts JM: CDK inhibitors:
Positive and negative regulators of G1-phase progression. Genes
Dev. 13:1501–1512. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Musgrove EA, Caldon CE, Barraclough J,
Stone A and Sutherland RL: Cyclin D as a therapeutic target in
cancer. Nat Rev Cancer. 11:558–572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Alves CL, Elias D, Lyng M, Bak M,
Kirkegaard T, Lykkesfeldt AE and Ditzel HJ: High CDK6 protects
cells from fulvestrant-mediated apoptosis and is a predictor of
resistance to fulvestrant in estrogen receptor-positive metastatic
breast cancer. Clin Cancer Res. 22:5514–5526. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
VanArsdale T, Boshoff C, Arndt KT and
Abraham RT: Molecular pathways: Targeting the cyclin D-CDK4/6 axis
for cancer treatment. Clin Cancer Res. 21:2905–2910. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Foidart P, Yip C, Radermacher J, Blacher
S, Lienard M, Montero-Ruiz L, Maquoi E, Montaudon E,
Château-Joubert S, Collignon J, et al: Expression of MT4-MMP, EGFR,
and RB in triple-negative breast cancer strongly sensitizes tumors
to erlotinib and palbociclib combination therapy. Clin Cancer Res.
Nov 30–2018.(Epub ahead of print). doi:
10.1158/1078–0432.CCR-18-1880.
|
|
33
|
Tait SW and Green DR: Mitochondrion and
cell death: Outer membrane permeabilization and beyond. Nat Rev Mol
Cell Biol. 11:621–632. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ichim G and Tait SW: A fate worse than
death: Apoptosis as an oncogenic process. Nat Rev Cancer.
16:539–548. 2016. View Article : Google Scholar : PubMed/NCBI
|