Introduction
Lung cancer is one of the leading causes of
cancer-associated mortality worldwide (1). Lung cancer can be classified into the
following two subgroups: Small-cell lung cancer and non-small-cell
lung cancer (NSCLC) (2). In 2017,
NSCLC was reported to account for almost 80% of all cases of lung
cancer worldwide (3). The overall
survival rate for patients with NSCLC is poor, as diagnosis often
occurs at an advanced stage due to a lack of efficient diagnosis
methods (4). Investigations
regarding the mechanisms underlying the development and progression
of NSCLC may improve the precise diagnosis of the disease.
MicroRNAs (miRNAs or miRs) are a family of
endogenous RNAs with a length of ~8–25 nucleotides (5). miRNAs have been reported to serve
crucial roles in the development and progression of human cancer as
they participate in almost all cell malignancy behaviors (6–8). No
miRNA has been used as diagnostic or prognostic marker in the
clinic; however, numerous studies have highlighted the significance
of these molecules in human cancer (9,10).
miR-20b-5p is located at human chromosome Xq26.2, a site that has
previously been reported to be associated with the initiation or
progression of a number of types of cancer (11). Recently, it has been reported that
miR-20b-5p is abnormally expressed in human cancer and functions as
a tumor suppressor or oncogene in a context-dependent manner
(12–14). Li et al (12) demonstrated that miR-20b-5p expression
was downregulated in renal cell carcinoma, in the first study to
reveal a tumor suppressive role of miR-20b-5p. A recent study
(13) demonstrated that miR-20b-5p
was upregulated in breast cancer and could be used as a biomarker
to diagnose breast cancer. A study using RT-qPCR revealed that an
increased expression level miR-20b-5p could effectively distinguish
NSCLC from a control (14). However,
the biological functions of miR-20b-5p in NSCLC remain unclear.
The present study revealed that miR-20b-5p is
overexpressed in NSCLC tissues and cell lines. Cell Counting Kit-8
(CCK-8) and wound-healing assays were performed to investigate the
effects of miR-20b-5p expression on NSCLC cell proliferation and
migration. In addition, B-cell translocation gene 3 (BTG3) was
identified as a direct target of miR-20b-5p by using
bioinformatics, a luciferase activity reporter assay and western
blot analysis.
Materials and methods
Clinical specimens
A total of 113 pairs of tumor tissue samples and
adjacent non-cancerous tissue samples (≥2 cm away from the tumor)
were collected from patients with NSCLC (57 male and 56 female;
56.7±4.2 years) who underwent treatment at Guangzhou General
Hospital of The People's Liberation Army (PLA) (Guangzhou, China)
between March 2011 and November 2012. The tissue samples were
snap-frozen in liquid nitrogen following surgery and stored at
−80°C prior to further use. Tumor stage was classified according to
the American Joint Committee on Cancer staging system (15). The present study was approved by the
Ethics Committee of Guangzhou General Hospital of PLA (Guangzhou,
China). Written informed consent was obtained from all recruited
patients.
Cell culture and transfection
A549, H1299, and 16HBE cell lines were purchased
from the Cell Bank of the Chinese Academy of Sciences (Shanghai,
China). All cells were cultured in RPMI-1640 medium (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (Invitrogen; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin in a
humidified incubator at 37°C containing 5% CO2. The
miR-20b-5p mimic (5′-CAAAGUGCUCAUAGUGCAGGUAG-3′), inhibitor
(5′-CUACCUGCACUAUGAGCACUUUG-3′) and negative control (NC;
5′-GCUAGAUGCACUCAUCUCUACGU-3′) were obtained from Shanghai
GenePharma Co., Ltd. (Shanghai, China). The BTG3 expression
construct and the pcDNA3.3 NC were purchased from GenScript
(Nanjing, China). Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) was used for cell transfections,
according to the manufacturer's protocol. Subsequent experiments
were performed 48 h after transfection.
RT-qPCR
Total RNA from tissues and cell lines was extracted
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). cDNA was synthesized using a miScript reverse
transcription kit (Qiagen GmbH, Hilden, Germany). RT-qPCR was
performed with a SYBR-Green PCR master mix (Thermo Fisher
Scientific, Inc.) on a 7500 Real-time PCR system (Thermo Fisher
Scientific, Inc.). Primers were synthesized by GenScript (Nanjing,
China) with the following sequences: miR-20b-5p forward,
5′-TGTCAACGATACGCTACGA-3′ and reverse, 5′-GCTCATAGTGCAGGTAGA-3′;
and U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. Relative expression levels were
calculated using the 2−ΔΔCq method with U6 as the
internal control (16). The
thermocycling conditions were as follows: 95°C for 2 min, 95°C for
10 sec, 55°C for 30 sec and 72°C for 30 sec, for 40 cycles. Each
experiment was performed in triplicate.
Protein sample extraction and western
blot analysis
Total protein from tissues and cell lines was
extracted using RIPA lysis buffer (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Protein concentration was measured using a BCA kit (Beyotime
Institute of Biotechnology, Haimen, China). The extracted protein
samples (50 µg) were separated by 10% SDS-PAGE and then transferred
to polyvinylidene fluoride membranes (Invitrogen; Thermo Fisher
Scientific, Inc.). Subsequently, the membranes were blocked with 5%
non-fat milk for 2 h at room temperature prior to incubation with
primary antibodies targeting BTG3 (1:1,000; catalog no. ab112938;
Abcam, Cambridge, MA, USA) or GAPDH (1:1,000; catalog no. ab181602;
Abcam) overnight at 4°C. Subsequently, the membranes were incubated
with horseradish peroxidase-conjugated goat anti-rabbit secondary
antibody (1:5,000; catalog no. ab6721; Abcam) for 1 h at room
temperature. Protein signals were visualized using an enhanced
chemiluminescence detection system (Beyotime Institute of
Biotechnology) and analyzed with ImageJ 1.42 software (National
Institutes of Health, Bethesda, MD, USA).
CCK-8 assay
Cell proliferation ability was measured using a
CCK-8 assay (Beyotime Institute of Biotechnology), according to the
manufacturer's protocol. Briefly, the A549 and H1299 cells were
seeded at a density of 5,000 cells/well in a 96-well plate and
incubated in RPMI-1640 medium supplemented with 10% fetal bovine
serum, 100 U/ml penicillin and 100 µg/ml streptomycin. CCK-8
reagent was added to each well at 0, 24, 48 and 72 h, and the cells
were further incubated for 2 h. The absorbance was measured at 450
nm using an ELISA reader (BioTek Instruments, Inc., Winooski, VT,
USA).
Wound-healing assay
Cell migration ability was measured using a
wound-healing assay, as described previously (17). Briefly, 5×105 cells (A549
and H1299) were seeded in a 12-well plate and cultured until ~80%
confluence. A wound was then created using a sterile 200-µl pipette
tip at the surface of the cells in each well. Subsequently, the
cells were washed with PBS to remove cell debris. Images were
acquired using a Leica DMI 6000B inverted light microscope (Leica
Microsystems, Inc., Buffalo Grove, IL, USA) at 0 or 24 h after the
wounds were made to measure the wound width.
Bioinformatic analysis
miR-20b-5p targets were predicted and analyzed using
the online miRNA targets prediction algorithm TargetScan
(www.targetscan.org). Targets to be
investigated were selected based on the reported gene functions.
These results revealed that BTG3 may be a target of miR-20b-5p as
it contains a miR-20b-5p binding site in its 3′-untranslated region
(3′-UTR).
Dual-luciferase reporter assay
The putative wild-type (wt) and mutant (mut)
miR-20b-5p binding sequences were cloned into a pGL3 vector
(Promega Corporation, Madison, WI, USA). Cells were co-transfected
with the built constructs along with miR-20b-5p mimic or NC using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
Luciferase activity was measured using a Dual-Luciferase assay kit
(Promega Corporation) 48 h after transfection. Data were normalized
to the activity of the Renilla luciferase reference
plasmid.
Statistical analysis
Data were analyzed with GraphPad Prism 6 software
(GraphPad Software, Inc., La Jolla, CA, USA) and presented as the
mean ± standard deviation. Analysis between two groups was
performed with a paired Student's t-test. One-way analysis of
variance with Tukey's post hoc test was used for the comparison of
multiple groups. A χ2 test was performed to analyze the
associations of miR-20b-5p expression and clinicopathological
features. The correlation between miR-20b and BTG3 was analyzed
with Pearson's correlation. Survival analysis was performed with
Kaplan-Meier curve and log-rank test. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-20b-5p is upregulated in NSCLC
tissues and cell lines
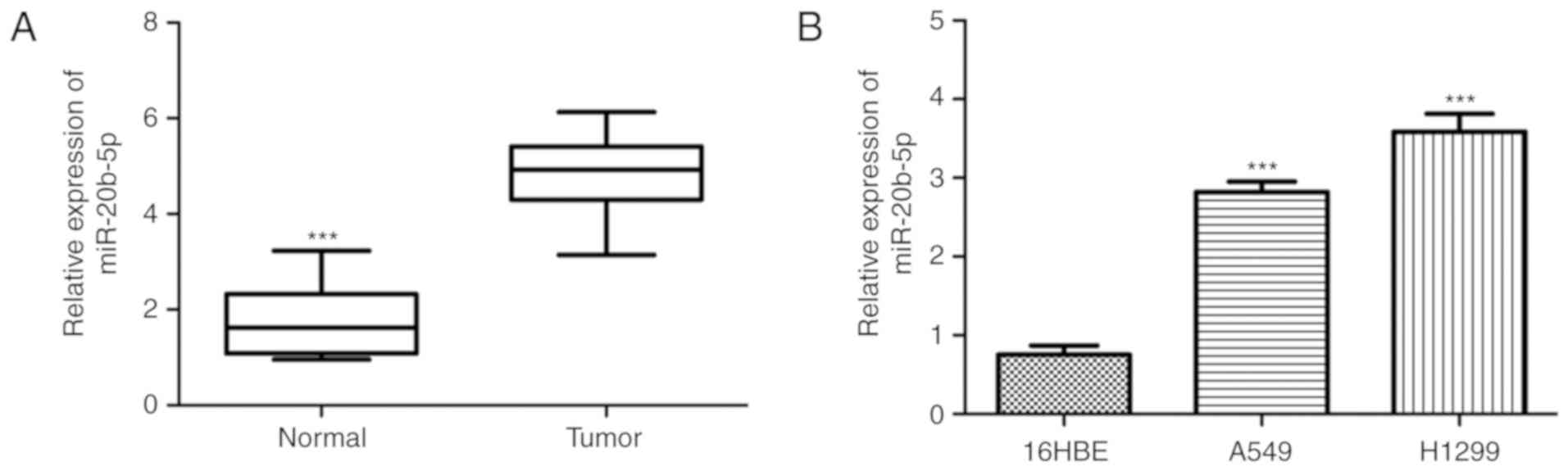
It was identified that the miR-20b-5p expression
level was significantly higher in NSCLC tissues compared with
normal adjacent tissues (Fig. 1A).
Furthermore, the level of miR-20b-5p in NSCLC cell lines was
investigated, which revealed that the expression level of
miR-20b-5p was significantly higher in the NSCLC cell lines A549
and H1299 compared with the normal 16HBE cell line (Fig. 1B).
Clinical significance of miR-20b-5p
expression in NSCLC
Patients with NSCLC were classified into high or low
miR-20b-5p expression groups based on the median miR-20b-5p
expression level. Patients with an miR-20b-5p expression level
equal to or higher than the median value (4.12) were classified
into the high expression group. Otherwise, the patients were
classified into the low expression group. It was identified that
miR-20b-5p expression was not significantly associated with age,
sex, hepatitis B surface antigen and smoking status (Table I). However, significant differences
were revealed between miR-20b-5p expression level and tumor size
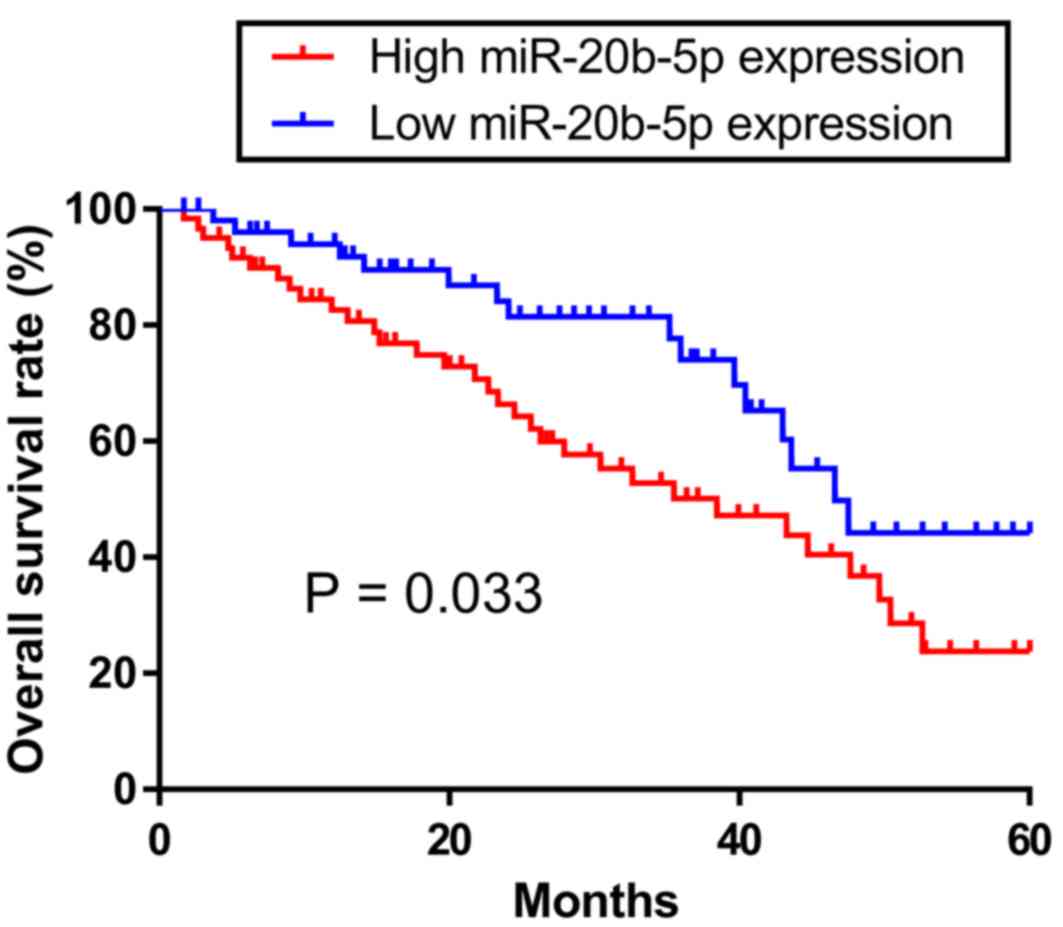
and tumor stage. Kaplan-Meier analysis and a log-rank test were
performed to analyze the association of miR-20b-5p expression with
the overall survival rate. The overall survival rate of patients
with a high miR-20b-5p expression level was significantly lower
compared with those with a low miR-20b-5p expression level
(P=0.033; Fig. 2).
 | Table I.Association between miR-20b-5p
expression and clinicopathological features of patients with
non-small cell lung cancer. |
Table I.
Association between miR-20b-5p
expression and clinicopathological features of patients with
non-small cell lung cancer.
|
|
| miR-20b-5p
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | No. of cases | High | Low | P-valuea |
|---|
| Sex |
|
|
| 0.484 |
| Male | 57 | 29 | 28 |
|
|
Female | 56 | 31 | 25 |
|
| Age, years |
|
|
| 0.100 |
|
≥50 | 62 | 32 | 30 |
|
|
<50 | 51 | 28 | 23 |
|
| HBsAg |
|
|
| 0.106 |
|
Negative | 52 | 30 | 22 |
|
|
Positive | 61 | 30 | 31 |
|
| Smoking status |
|
|
| 0.166 |
|
Yes | 52 | 26 | 26 |
|
| No | 61 | 34 | 27 |
|
| Tumor size, cm |
|
|
| 0.025 |
| ≥5 | 64 | 35 | 29 |
|
|
<5 | 49 | 25 | 24 |
|
| Tumor stage |
|
|
| 0.014 |
|
I–II | 63 | 36 | 27 |
|
|
III | 50 | 24 | 26 |
|
Knockdown of miR-20b-5p inhibits NSCLC
cell proliferation and migration
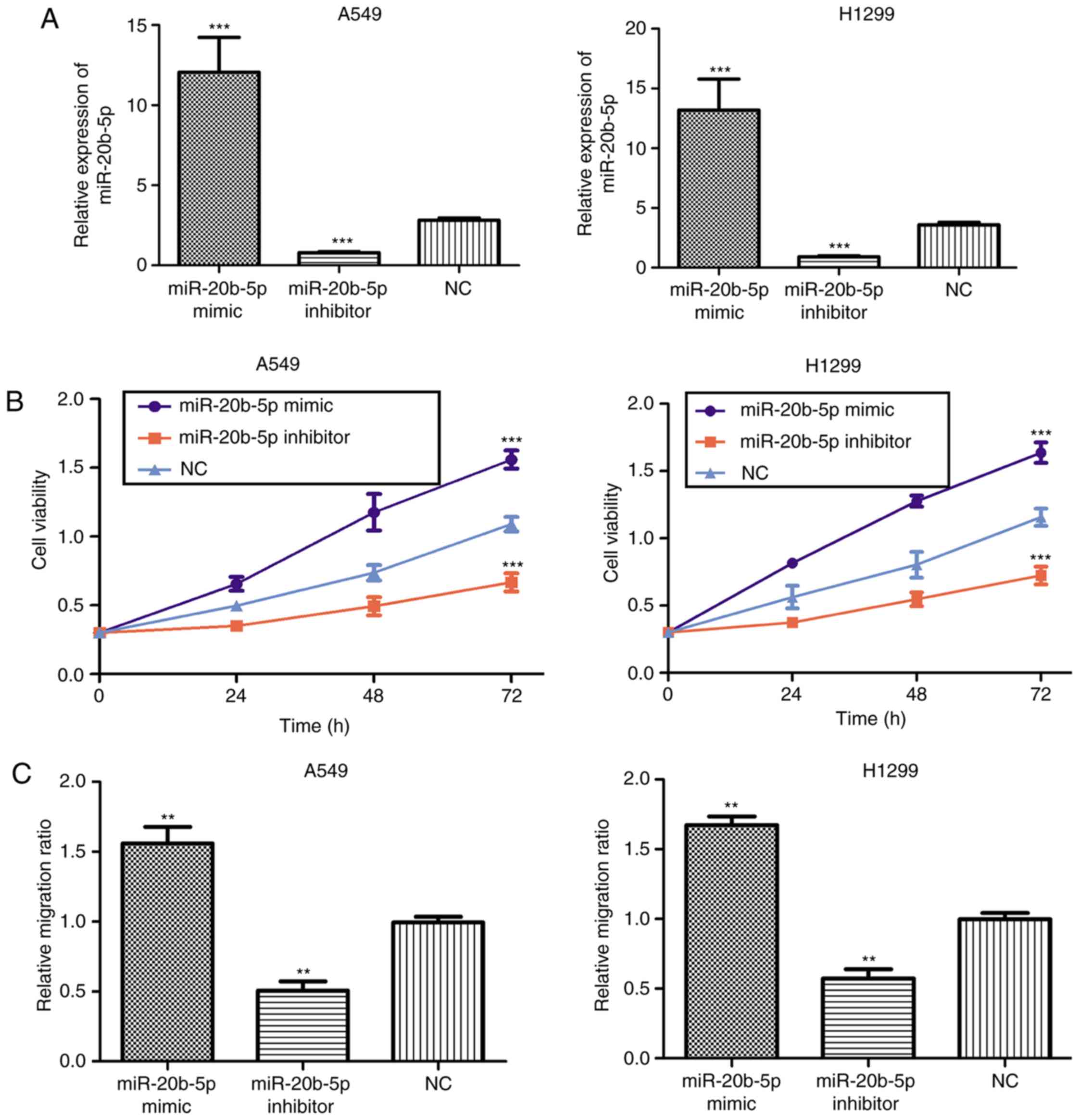
The in vitro biological function of
miR-20b-5p was assessed by CCK-8 assay and a wound-healing assay.
The expression level of miR-20b-5p in NSCLC cells was altered by
transfection with miR-20b-5p mimic or inhibitor. It was
demonstrated that transfection with miR-20b-5p mimic significantly
increased the level of miR-20b-5p, while miR-20b-5p inhibitor
significantly downregulated the level of miR-20b-5p (Fig. 3A). CCK-8 assay results demonstrated
that the proliferation ability of NSCLC cells was significantly
decreased following transfection with miR-20b-5p inhibitor but
significantly increased by miR-20b-5p mimic (Fig. 3B). Furthermore, a wound-healing assay
revealed that the migratory distance of NSCLC cells transfected
with miR-20b-5p mimic was significantly larger compared with the NC
group. In addition, the migratory distance was significantly
smaller for the NSCLC cells transfected with inhibitor compared
with the NC group (Fig. 3C). These
data indicate that miR-20b-5p overexpression significantly
increases cell proliferation and migration.
BTG3 is a target of miR-20b-5p in
NSCLC
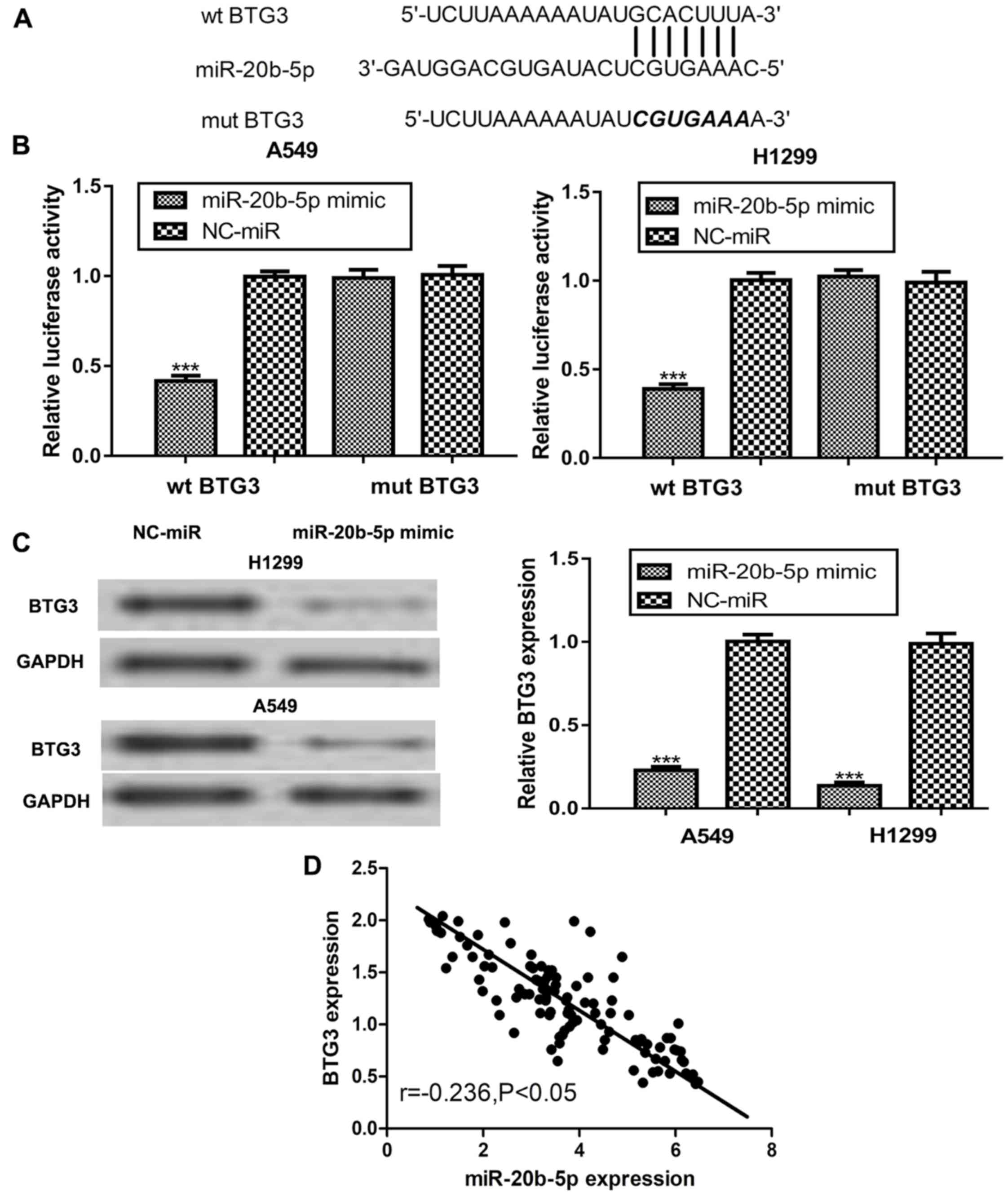
Online targets analysis revealed that BTG3 contains
a putative binding site for miR-20b-5p in its 3′-UTR (Fig. 4A). Furthermore, a dual-luciferase
reporter assay demonstrated that transfection with miR-20b-5p mimic
significantly decreased the luciferase activity in NSCLC cells
transfected with wt BTG3 but not in those transfected with mut BTG3
(Fig. 4B). Subsequently, BTG3
protein expression was examined in the miR-20b-5p mimic or
NC-transfected NSCLC cells. It was identified that transfection
with miR-20b-5p mimic significantly decreased BTG3 protein
expression in NSCLC cells compared with that in the NC group
(Fig. 4C). Furthermore, an inverse
correlation was revealed between miR-20b-5p and BTG3 expression
levels in NSCLC tissues (Fig. 4D).
These results suggest that BTG3 is a direct target of
miR-20b-5p.
miR-20b-5p promotes NSCLC cell
proliferation and migration by targeting BTG3
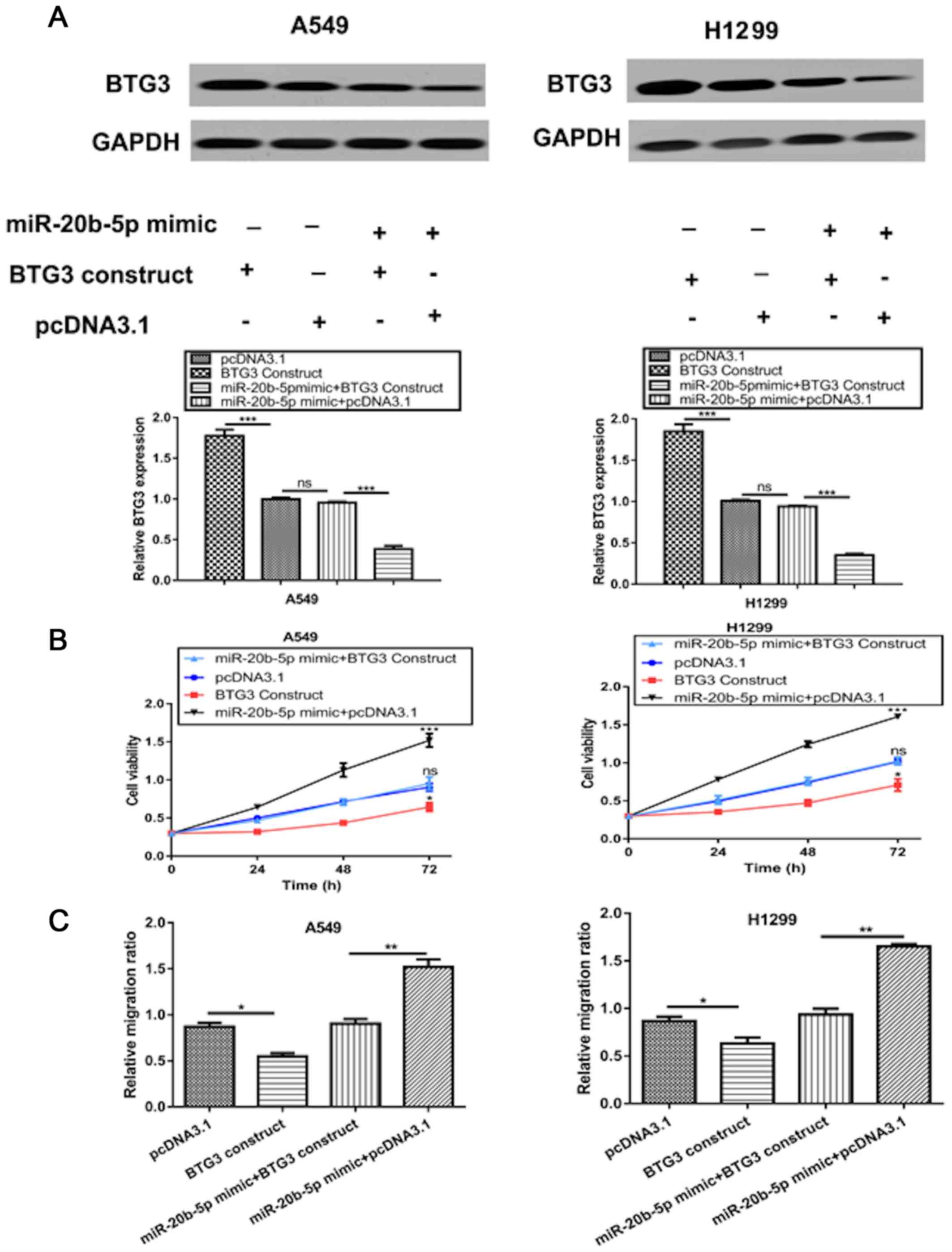
To validate BTG3 as a functional target of
miR-20b-5p, NSCLC cells were co-transfected with a BTG3 expression
construct and miR-20b-5p mimic. Western blot analysis demonstrated
that transfection with the BTG3 expression construct markedly
increased the protein level of BTG3 in NSCLC cells (Fig. 5A). Furthermore, transfection with
miR-20b-5p mimic reduced the stimulation effect of BTG3 expression
construct on the expression level of BTG3 (Fig. 5A). As presented in Fig. 5B, overexpression of BTG3
significantly reversed the stimulation effect of miR-20b-5p mimic
on cell proliferation. Similarly, the promoting effect of
miR-20b-5p mimic on cell migration was significantly inhibited by
BTG3 overexpression (Fig. 5C).
Discussion
miR-20b-5p, along with miR-17-92 and miR-106b-25
clusters, form a large and highly conserved miRNA family, termed
the miR-17 family (18). Members of
the miR-17 family have been reported to be highly expressed in
human cancer types and have been suggested to function as oncogenes
(19,20). In addition, previous studies have
demonstrated that miR-20b-5p can regulate a number of cell
behaviors, including cell proliferation, cell migration and cell
apoptosis (12,13). A previous study has examined the
expression level of miR-20b-5p in NSCLC (14); however, to the best of our knowledge,
no previous study has investigated its clinical significance in
NSCLC.
The present study identified that miR-20b-5p
expression was upregulated in NSCLC tissues, which is the same
result that was observed in a high throughput RT-qPCR experiment
(14). The investigation of
miR-20b-5p expression level in NSCLC cell lines and a normal cell
line also revealed that miR-20b-5p expression was upregulated in
NSCLC. Subsequently, the associations between miR-20b-5p expression
and clinicopathological parameters were evaluated, which revealed
that miR-20b-5p expression was significantly associated with two
classical tumor malignancy indicators, tumor size and tumor stage.
Furthermore, it was identified that a high expression level of
miR-20b-5p predicts a worse 5-year overall survival rate.
It has been established that miRNAs exert their
biological roles via regulation of cancer-associated genes
(7–10). In addition, miR-20b-5p has been
reported to regulate a number of human genes in cancer (12). BTG3 is a target of p53 and a negative
regulator of cell cycle progression and cell proliferation
(21,22). BTG3 expression has been identified to
be downregulated in colorectal cancer and reported to regulate cell
proliferation, migration, invasion, the cell cycle and apoptosis
(23). Furthermore, BTG3
overexpression can inhibit epithelial ovarian cancer cell
proliferation and invasion but promote cell apoptosis via
regulation of the AKT/glycogen synthase kinase-3β/β-catenin pathway
(24). It has been reported that
BTG3 expression is reduced in NSCLC and functions as a tumor
suppressor (25).
Notably, the present study identified that the
3′-UTR of BTG3 contains a binding site for miR-20b-5p. Therefore,
it was then investigated whether miR-20b-5p and BTG3 are associated
in NSCLC. Western blot analysis revealed that BTG3 expression could
be regulated by miR-20b-5p, which indicates that BTG3 is a direct
target of miR-20b-5p. Further in vitro functional assays
revealed that miR-20b-5p overexpression could promote NSCLC cell
proliferation and migration. These results suggest that miR-20b-5p
functions as an oncogene in NSCLC, which is a role that has been
identified in other cancer types, including breast and prostate
cancer (13,26). Mechanistic studies revealed that BTG3
was a functional target of miR-20b-5p. Notably, overexpression of
BTG3 only partially reversed the effects of miR-20b-5p mimic on
cell proliferation and migration, which indicates that other
molecules may also be involved in this process. For example, it has
been reported that miR-20b-5p regulates myoblast differentiation
and proliferation by directly regulating the expression of E2F
transcription factor 1 (E2F1) (27).
Additionally, BTG3 binds to and inhibits E2F1 via an N-terminal
domain to regulate growth (28).
Therefore, it is reasonable to suggest that E2F1 may also
participate in the miR-20b-5p-mediated NSCLC cell behaviors and
this requires further investigation in the future.
In conclusion, the present study revealed that
miR-20b-5p expression is elevated in NSCLC and overexpression of
miR-20b-5p can promote cell proliferation and migration by
targeting BTG3. Notably, a high miR-20b-5p expression level was
identified to be associated with a worse 5-year overall survival
rate. Therefore, miR-20b-5p may serve as a novel target for NSCLC
early diagnosis or treatment.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LP, SL, DL, and YX conceived and deigned the study.
LP, SL, YL, MW, XF, YZ, WZ, DL and YX performed the experiments and
were major contributors in writing the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by Ethic Committee of
Guangzhou General Hospital of PLA (Guangzhou, China). Written
informed consent was obtained from all enrolled patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gompelmann D, Eberhardt R and Herth FJ:
Advanced malignant lung disease: What the specialist can offer.
Respiration. 82:111–123. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kang M, Shi J, Peng N and He S:
MicroRNA-211 promotes non-small-cell lung cancer proliferation and
invasion by targeting MxA. Onco Targets Ther. 10:5667–5675. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mallory AC and Vaucheret H: MicroRNAs:
Something important between the genes. Curr Opin Plant Biol.
7:120–125. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
94:776–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Florczuk M, Szpechcinski A and
Chorostowska-Wynimko J: miRNAs as biomarkers and therapeutic
targets in non-small cell lung cancer: Current perspectives. Target
Oncol. 12:179–200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saleiban A, Faxälv L, Claesson K, Jönsson
JI and Osman A: miR-20b regulates expression of
proteinase-activated receptor-1 (PAR-1) thrombin receptor in
melanoma cells. Pigment Cell Melanoma Res. 27:431–441. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Y, Chen D, Jin L, Liu J, Su Z, Li Y,
Gui Y and Lai Y: MicroRNA-20b-5p functions as a tumor suppressor in
renal cell carcinoma by regulating cellular proliferation,
migration and apoptosis. Mol Med Rep. 13:1895–1901. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li M, Zhou Y, Xia T, Zhou X, Huang Z,
Zhang H, Zhu W, Ding Q and Wang S: Circulating microRNAs from the
miR-106a-363 cluster on chromosome X as novel diagnostic biomarkers
for breast cancer. Breast Cancer Res Treat. 170:257–270. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Leidinger P, Brefort T, Backes C, Krapp M,
Galata V, Beier M, Kohlhaas J, Huwer H, Meese E and Keller A:
High-throughput qRT-PCR validation of blood microRNAs in non-small
cell lung cancer. Oncotarget. 7:4611–4623. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Greene FL, Page DL, Fleming ID, Fritz A,
Balch CM and Haller DG: AJCC cancer staging handbook from the AJCC
cancer staging manual. (6th). Springer. (New York). 2002.
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao X, Zhao H, Diao C, Wang X, Xie Y, Liu
Y, Han J and Zhang M: miR-455-3p serves as prognostic factor and
regulates the proliferation and migration of non-small cell lung
cancer through targeting HOXB5. Biochem Biophys Res Commun.
495:1074–1080. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tanzer A and Stadler PF: Molecular
evolution of a microRNA cluster. J Mol Biol. 339:327–335. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hayashita Y, Osada H, Tatematsu Y, Yamada
H, Yanagisawa K, Tomida S, Yatabe Y, Kawahara K, Sekido Y and
Takahashi T: A polycistronic microRNA cluster, miR-17-92, is
overexpressed in human lung cancers and enhances cell
proliferation. Cancer Res. 65:9628–9632. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Landais S, Landry S, Legault P and Rassart
E: Oncogenic potential of the miR-106-363 cluster and its
implication in human T-cell leukemia. Cancer Res. 67:5699–5707.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng YC, Lin TY and Shieh SY: Candidate
tumor suppressor BTG3 maintains genomic stability by promoting
Lys63-linked ubiquitination and activation of the checkpoint kinase
CHK1. Proc Natl Acad Sci USA. 110:5993–5998. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Majid S, Dar AA, Ahmad AE, Hirata H,
Kawakami K, Shahryari V, Saini S, Tanaka Y, Dahiya AV, Khatri G and
Dahiya R: BTG3 tumor suppressor gene promoter demethylation,
histone modification and cell cycle arrest by genistein in renal
cancer. Carcinogenesis. 30:662–670. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lv C, Wang H, Tong Y, Yin H, Wang D, Yan
Z, Liang Y, Wu D and Su Q: The function of BTG3 in colorectal
cancer cells and its possible signaling pathway. J Cancer Res Clin
Oncol. 144:295–308. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
An Q, Zhou Y, Han C, Zhou Y, Li F and Li
D: BTG3 overexpression suppresses the proliferation and invasion in
epithelial ovarian cancer cell by regulating AKT/GSK3β/β-catenin
signaling. Reprod Sci. 24:1462–1468. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen X, Chen G, Cao X, Zhou Y, Yang T and
Wei S: Downregulation of BTG3 in non-small cell lung cancer.
Biochem Biophys Res Commun. 437:173–178. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo J, Xiao ZW, Yu XW and Cao RF: miR-20b
promotes cellular proliferation and migration by directly
regulating phosphatase and tensin homolog in prostate cancer. Oncol
Lett. 14:6895–6900. 2017.PubMed/NCBI
|
|
27
|
Luo W, Li G, Yi Z, Nie Q and Zhang X:
E2F1-miR-20a-5p/20b-5p auto-regulatory feedback loop involved in
myoblast proliferation and differentiation. Sci Rep. 6:279042016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ou YH, Chung PH, Hsu FF, Sun TP, Chang WY
and Shieh SY: The candidate tumor suppressor BTG3 is a
transcriptional target of p53 that inhibits E2F1. EMBO J.
26:3968–3980. 2007. View Article : Google Scholar : PubMed/NCBI
|