Introduction
Radiation-induced lung injury (RILI) is the most
common dose-limiting side effect in patients with thoracic tumors
following radiotherapy (1,2). Various strategies and drugs have been
applied for the prevention and treatment of RILI, with poor
therapeutic effect (3–6). Glucocorticoids have been administrated
in numerous clinical studies for the treatment of acute RILI
(1,7–9).
However, prolonged usage of high doses of glucocorticoids can lead
to a number of side effects, including osteoporosis, abnormal
glucose metabolism, digestive tract ulcers and infection (10). Averting the side effects of
radiotherapy without affecting the clinical benefits is an
important goal.
The pathogenesis of RILI is complicated. Alveolar
type II epithelial cells and lung capillary endothelial cells are
the main targets of RILI (11,12),
producing pro-inflammatory cytokines, including interleukin-6
(IL-6) (7) and transforming growth
factor-β1 (TGF-β1) (13).
Macrophages are the most important secretory cells of IL-6 and
TGF-β1. In addition, macrophages also produce tumor necrosis
factor-α (TNF-α), insulin-like growth factor 1, platelet-derived
growth factor-β, macrophage-derived chemokines and other cytokines,
These cytokines enhance the permeability of lung capillary
endothelium and the chemotactic recruitment of monocytes,
macrophages and neutrophils to the injured lung tissue in the
target area. The inflammatory signal is amplified due to increased
production of IL-6 and the generation of myeloperoxidase (MPO) by
neutrophils, aggravating local inflammation and leading to damage
of the alveolar structure (14–16). In
addition, the secretion of TGF-β by macrophages can also lead to
the progression of RILI into chronic pulmonary fibrosis. As a
result, macrophages and neutrophils are influential in this
process.
Annexin A1 (ANXA1) is a protein regulated by
glucocorticoids. ANXA1 mimics the anti-inflammatory effect of
glucocorticoids (17,18). ANXA1 is expressed in neutrophils,
eosinophils and subcellular granules of monocytes and macrophages
in the peripheral blood (19), and
it inhibits the migration of neutrophils (20). Thus, ANXA1 has clinical potential for
the treatment of RILI.
The role of ANXA1 in the pathogenesis of acute RILI
and the mechanism of its clinical benefit on RILI are unclear. The
present study evaluated the therapeutic potential of ANXA1 against
RILI by assessing ANXA1 expression in the plasma of patients with
RILI prior to and following 4 weeks of glucocorticoid treatment.
The anti-inflammatory mechanism of ANXA1 was investigated by
upregulating and downregulating ANXA1 expression in a macrophage
cell line to assess the impact on IL-6 and MPO expression. Further
understanding of the anti-inflammatory effect of ANXA1 on RILI may
provide pre-clinical evidence to support targeting ANXA1 in the
prevention and treatment of RILI.
Materials and methods
Subject information
A total of 50 patients with thoracic tumors
suffering with RILI following radiotherapy between January 2014 and
October 2016 at Taizhou People's Hospital (Taizhou, China) were
included in the present study. The patients had experienced RILI
with a severity of grade 2 or more after therapy, according to the
Common Terminology Criteria for Adverse Events from the National
Cancer Institute (version 3.0; National Institutes of Health,
Rockville, MD, USA) (21). A total
of 45 patients with complete clinical data were involved in the
result analysis, including 25 males and 20 females, and the age
ranging between 47 and 75 years old (median, 63.5 years). The
research was approved by the Ethics Committee of Taizhou People's
Hospital, and written informed consent was provided by each patient
prior to the study.
Treatment of acute RILI and evaluation
of treatment outcome
Glucocorticoid (methylprednisolone or dexamethasone)
was administered to patients with RILI as the basic comprehensive
treatment immediately after diagnosis. Methylprednisolone was
administrated intravenously (iv), with a starting dose 1–2
mg/kg/day, and the dose would reduce by half every 4 days.
Dexamethasone was administrated with a starting dose of 10 mg/day
iv, and reduced as described previously. Routine blood examination
was performed weekly during treatment, and a thoracic computed
tomography scan was performed after 4 weeks of treatment. Response
to treatment was assessed by alleviation of clinical symptoms (e.g.
cough, fever, chest distress, dyspnea) and imaging (X-ray, CT scan)
and laboratory findings (white blood cell count). The treatment
outcome was evaluated comprehensively based on the patients'
clinical symptoms and the aforementioned indicators, including CT
scan and white blood cell count and neutrophils count.
Neutrophil count and ELISA
Peripheral venous blood was drawn from each patient
weekly from the time of diagnosis until 1 month post-treatment.
Then, the neutrophil count was measured through routine blood
tests, and the expression of ANXA1, IL-6 and MPO was assessed using
ELISA in the plasma extracted by centrifugation from the blood
samples. The ANXA1 (cat. no. ab222868; antibody name: AF3770) and
IL-6 (cat. no. PD6050; antibody name: MAB206) kits, and the primary
antibody were purchased from R&D Systems China Co., Ltd. The
MPO kit was purchased from Abcam Shanghai Co., Ltd. (cat. no.
ab119605; antibody name: ab45977). Measurements were performed in
triplicate for each sample. The absorbance was read at 450 nm and
the protein levels were calculated based on the standard curve.
Blood samples from 20 healthy donors were also assessed as the
negative control.
Plasmid construction for ANXA1
expression
Eca109 human esophageal cancer cells, purchased from
Shanghai Institute of Biochemistry and Cell Biology Chinese Academy
of Sciences were cultured in DMEM with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.) and antibiotics at 37°C with
5% CO2. RNA from Eca109 human esophageal cancer cells
was extracted using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc., Shanghai, China). The AMV reverse
transcription kit (Promega Corporation) was subsequently used for
reverse transcription to obtain the cDNA which was used as a
template, together with ANXA1 primers (synthesized by Invitrogen;
Thermo Fisher Scientific, Inc. Shanghai, China) for PCR
amplification: The primer sequences were as follows: Sense,
5′-ATGGCAATGGTATCAGAATTCCTC-3′ and antisense,
5′-TTAGTTTCCTCCACAAAGAGCCACC-3′. PrimerSTAR HS DNA polymerase
(Takara Biotechnology Co., Ltd., Dalian, China) was used for
amplification. The reaction parameters were as follows: 95°C ×3
min, 94°C ×30 sec, 58°C ×30 sec, 72°C ×5 min, 72°C ×5 min, for 27
cycles. The PCR product was purified using the QIAquick PCR
Purification kit (Qiagen China Co., Ltd., Shanghai, China). ANXA1
cDNA was cleaved with the restriction endonucleases XhoI and
BamHI (Fermentas; Thermo Fisher Scientific, Inc.,
Pittsburgh, PA, USA) and ligated with the pIRES2-EGFP vector
(Takara Biotechnology Co., Ltd.) to form a pIRES2-EGFP-ANXA1
(pIRES2-ANXA1) expression plasmid. The plasmid was expressed in
competent Escherichia coli DH5α cells (Shanghai Institute of
Biochemistry and Cell Biology Chinese Academy of Sciences) and
purified using a mini-prep kit from Axygen Biosciences (Union City,
CA, USA). The construct was examined by enzyme digestion and
sequenced as previously described (22).
Differentiation and cultivation of M2
macrophages
Human peripheral-blood monocytes are used as an
established in vitro system for generating macrophages, and
monocytic cell lines, as THP-1 cells have been considered as a
possible alternative (23–25). In the present study, THP-1 cells were
acquired from the Shanghai Institute of Biochemistry and Cell
Biology, and were cultured in RPMI-1640 medium (Thermo Fisher
Scientific, Inc.) supplemented with non-heat-treated 10% FBS
(Invitrogen; Thermo Fisher Scientific, Inc.), 2 mM L-glutamine,
0.05 mM β-mercaptoethanol, 4,500 mg/l glucose, 100 U/ml penicillin
and 100 µg/ml streptomycin at 37°C with 5% CO2. THP-1
cells were kept at a minimum density of 5×105 cells/ml
and were passaged when reaching 5×106 cells/ml. The
cells were induced to differentiate into M2 macrophages by PMA
(Sigma-Aldrich; Merck KGaA) at a final concentration of 150 ng/ml
and IL-4 at a final concentration of 20 ng/ml incubated for 48 h.
The cells were imaged using an optical microscope. Differentiation
was verified by assessing expression of a marker of the M2
phenotype, macrophage mannose receptor 1 (CD206) (26–28).
Cells were collected at 48 h after induction for detection. The
induced cells were digested with trypsin and collected by
centrifugation at 200 × g for 10 min at 4°C (induction group).
Uninduced cells were centrifuged, as described previously, and
collected (control group). The pellet was resuspended in PBS and
the cells were washed with PBS solution twice. Then, the cells were
suspended at ~1×106 cells/ml in PBS for further use. The
Phycoerythrin (PE)-conjugated anti-CD206 monoclonal antibody
(dilution, 1:20; cat. no: 555954; BD Pharmingen) was added to the
cell suspension, which was subsequently incubated in the dark for
30 min at 4°C. After two washes in PBS, the cells were resuspended
with 2% paraformaldehyde (in cold PBS) and incubated for 20 min at
4°C. Suspension was centrifuged at 200 × g for 5 min at 4°C and the
pellet resuspended with glycine 0.1 M (in cold PBS) and incubated
for 10 min at 4°C. Then, the last centrifugation of 5 min at 200 ×
g at 4°C was performed before cell resuspension in 1 ml of PBS and
the expression of CD206 was measured using a flow cytometer
(FACScalibur; BD Biosciences), and analyzed by the FlowJo 7.6.4
software (TreeStar Inc.).
Overexpression and inhibition of ANXA1
in THP-1 cells differentiated into M2 macrophages
THP-1 cells were induced into macrophage 48 h after
transfection. The pIRES2-ANXA1 expression plasmid was transfected
into THP-1 cells using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). The pIRES2-EGFP empty plasmid was
transfected as the control. Prior to transfection, the plasmid (1
µg/ul) was mixed with liposomes and allowed to stand for 20 min.
The mixture was dispensed into culture wells (2 µg DNA and 5 ul
liposomes per well) following the manufacturer's protocol. The
transfection efficiency was assessed by fluorescence microscopy and
western blot analysis. The transfected THP-1 cells were then
induced to generate ANXA1-overexpressing macrophages. Macrophages
harboring the ANXA1 plasmid comprised the pIRES2-ANXA1 group and
those with the empty plasmid comprised the no-load group.
Non-transfected macrophages comprised the blank control. Each group
of cells was partitioned into 3 wells and the experiments were
repeated 3 times.
Small interfering RNAs (siRNAs) against ANXA1 were
transfected into THP-1 cells to inhibit the expression of ANXA1.
The siRNAs included 2 sequences targeting ANXA1 and a scrambled
control sequence as follows: siRNA (a), 5′-CAGCGUCAACAGAUCAAAG-3′;
siRNA (b), 5′-CCGAUCUGAGGACUUUGGU-3′; and control siRNA,
5′-CAGUCGCGUUUGCGACUGG-3.′ These siRNA sequences were designed as
previously described (29). The
siRNAs were synthesized by Shanghai Sangon Company (Shanghai,
China). After 48 h these transfected THP-1 cells were then induced
to differentiate into macrophages. Macrophages transfected with
siRNA (a) and siRNA (b) were designated as the siRNA (a) and siRNA
(b) groups, respectively. Macrophages transfected with control
siRNA or plasmid were designated as the siRNA (control) group or
the no-load group, respectively. Non-transfected macrophages
comprised the blank control group.
Western blot analysis
Macrophages in each well were trypsinized and total
cell protein was extracted using cell lysis buffer (cat. no. P0013;
Beyotime Institute of Biotechnology). Sample protein concentrations
were determined for each group using the DC Protein assay kit
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). SDS-PAGE (15%) was
performed for each group using 50 µg protein sample per lane, and
proteins were then transferred from the gel to polyvinylidene
difluoride membranes (EMD Millipore, Billerica, MA, USA). Following
a 1-h incubation in blocking buffer (cat. no. C-0042; 5% bovine
serum albumin; Xiamen, BOYAO Biotechnology Co., Ltd.) at 4°C,
membranes were incubated with the following primary antibodies at
4°C overnight with agitation: Anti-IL-6 (1:1,000; cat. no. RAB0307;
Sigma-Aldrich; Merck KGaA), anti-ANXA1 (1:1,000; cat. no.
SAB1405457; Sigma-Aldrich; Merck KGaA), anti-MPO (1:1,000; cat.no.
76923; Cell Signaling Technology, Inc., Danvers, MA, USA) and
anti-GAPDH (1:3,000; cat.no. sc-47724; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA). Following washing with stripping buffer
[β-mercaptoethanol 342 µl; 20% SDS 5 ml; Tris-Cl (pH 6.7) 3.125 ml;
ddH2O 41.533 ml to total volume 50 ml], the membranes were
incubated with agitation at room temperature for 1 h with a
secondary antibody (m-IgGκ BP-HRP: sc-516102;Santa Cruz
Biotechnology, Inc.) labeled with horseradish peroxidase in the
presence of 2% bovine serum albumin (Invitrogen; Thermo Fisher
Scientific, Inc), then HRP was visualized by Chemiluminescent
Western Blot Detection kit (SuperSignal West Femto; Thermo Fisher
Scientific, Inc.). The membranes were washed and imaged, and the
bands were analyzed semi-quantitatively with ImageJ 1.8 software
(https://imagej.nih.gov/ij/download.html).
Effect of altered expression of ANXA1
on expression of IL-6 and MPO
M2 macrophages in which ANXA1 was overexpressed or
inhibited were resuspended for cell counting following transfection
to ensure that the cell density in each well for further
cultivation was the same. Following 72 h in culture, ELISA was
performed on the medium to evaluate the level of IL-6 and MPO
secreted by the macrophages, and western blot analysis was
performed to evaluate the levels of IL-6 and MPO protein in the
macrophages. ELISA was performed as aforementioned for peripheral
venous blood samples.
Statistical analysis
Measurement data are presented as the mean ±
standard deviation. SPSS (version 19.0; IBM Corp., Armonk, NY, USA)
was utilized to create the database. Differences between means of
two groups were compared using independent-samples t-test and those
among multiple groups were compared using one-way analysis of
variance (ANOVA). ANOVA was used to compare the means among
baseline group and 1, 2, 3 and 4 weeks after treatment groups.
Then, the Least Significant Difference post hoc test was used to
compare between every two groups. Associations between treatment
outcome and ANXA1, IL-6 and MPO levels were compared using a
χ2 test. Correlation analysis was performed using
Spearman's rank correlation method. P<0.05 was considered to
indicate that a difference was statistically significant.
Results
ANXA1, IL-6 and MPO levels in the
plasma of patients with RILI
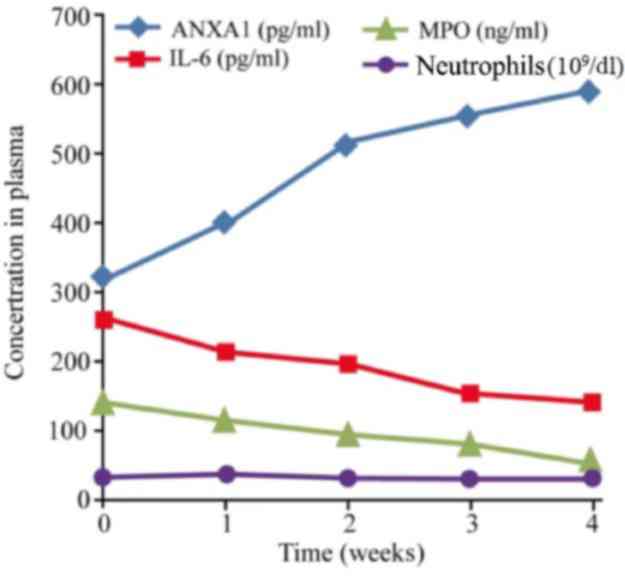
Among the 45 cases with RILI, the baseline level of
ANXA1, IL-6 and MPO in the plasma prior to glucocorticoid treatment
was 317.35±191.06, 258.45±172.32 pg/ml and 139.24±57.94 ng/ml,
respectively. The corresponding level of healthy control group was
22.75±9.56, 41.05±12.33 pg/ml and 62.24±27.39 ng/ml, respectively.
There was significant difference of ANXA1, IL-6 and MPO between the
baseline of RILI patients and the healthy control (P<0.001)
Following glucocorticoid administration, the expression of ANXA1
increased gradually (ANOVA test, F=31.503; P<0.001). The post
hoc test showed that ANXA1 of 1, 2, 3 and 4 weeks after treatment
was significantly higher compared with that of baseline, (P=0.003,
<0.001, <0.001, <0.001, respectively), while that of IL-6
and MPO decreased, and the number of neutrophils remained unchanged
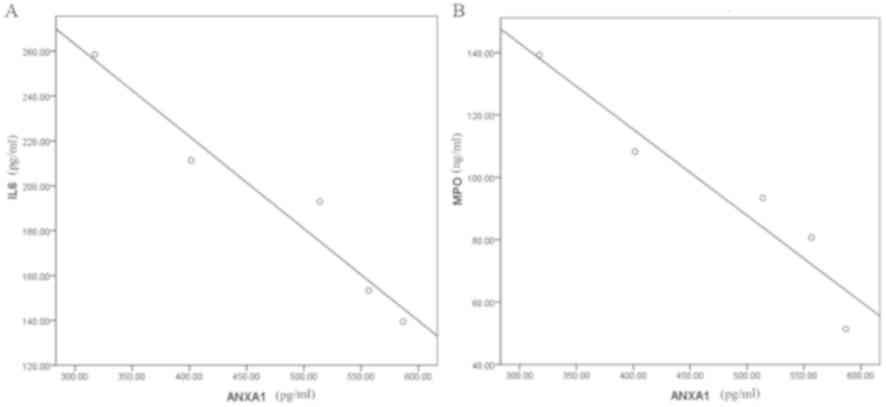
(Fig. 1). Spearman's correlation
analysis indicated that the ANXA1 level in the plasma following
glucocorticoid treatment was significantly negatively correlated
with the levels of IL-6 and MPO, with a correlation coefficient of
−0.974 and −0.956, respectively (P<0.001; Fig. 2).
Association between treatment outcome
and expression levels of ANXA1, IL-6 and MPO in patients with
RILI
Among the 45 cases, recovery occurred in 20 patients
2 weeks after glucocorticoid treatment, and a further 11 patients
with repeat symptoms were cured following 4 weeks of treatment.
These 31 cases were considered to have had an efficient treatment
outcome. Interstitial lung diseases combined with chronic lung
infection occurred in the remaining 14 cases, which were considered
to have progressive disease. Cases were grouped by these two
outcomes-efficient or progressive group. According to the increase
of ANXA1 after hormone treatment, patients were divided into two
groups: Patients with more than twice the baseline level and
patients with less than two times the baseline level. Similarly,
patients were divided into two groups according to the decrease of
IL-6 and MPO, which were less than half of the baseline level and
more than half of the baseline level group. Then, χ2
testing was performed to compare treatment outcome to expression
levels of ANXA1, IL-6 and MPO in plasma following glucocorticoid
treatment. The results indicated that the expression levels of
ANXA1, IL-6 and MPO were associated with the treatment outcome for
RILI. The increased expression of ANXA1 following 4 weeks of
treatment was associated with improved treatment outcome for RILI
(P=0.007), while decreased expression of IL-6 and MPO expression
following 4 weeks of treatment was associated with improved
treatment outcome (P=0.042 and P=0.003, respectively) (Table I).
 | Table I.Association between the clinical
effect against RILI and the plasma levels of ANXA1, IL-6 and MPO at
4 weeks of glucocorticoid treatment. |
Table I.
Association between the clinical
effect against RILI and the plasma levels of ANXA1, IL-6 and MPO at
4 weeks of glucocorticoid treatment.
|
| ANXA1 in
plasma | IL-6 in plasma | MPO in plasma |
|---|
|
|
|
|
|
|---|
| Clinical effect
against RILI | >2× baseline
level | <2× baseline
level | <50% of baseline
level | >50% of baseline
level | <50% of baseline
level | >50% of baseline
level |
|---|
| Efficient, n | 20 | 11 | 19 | 12 | 19 | 12 |
| Progressive, n | 3 | 11 | 4 | 10 | 2 | 12 |
| χ2 | 7.166 |
| 4.132 |
| 8.562 |
|
| P-value | 0.007 |
| 0.042 |
| 0.003 |
|
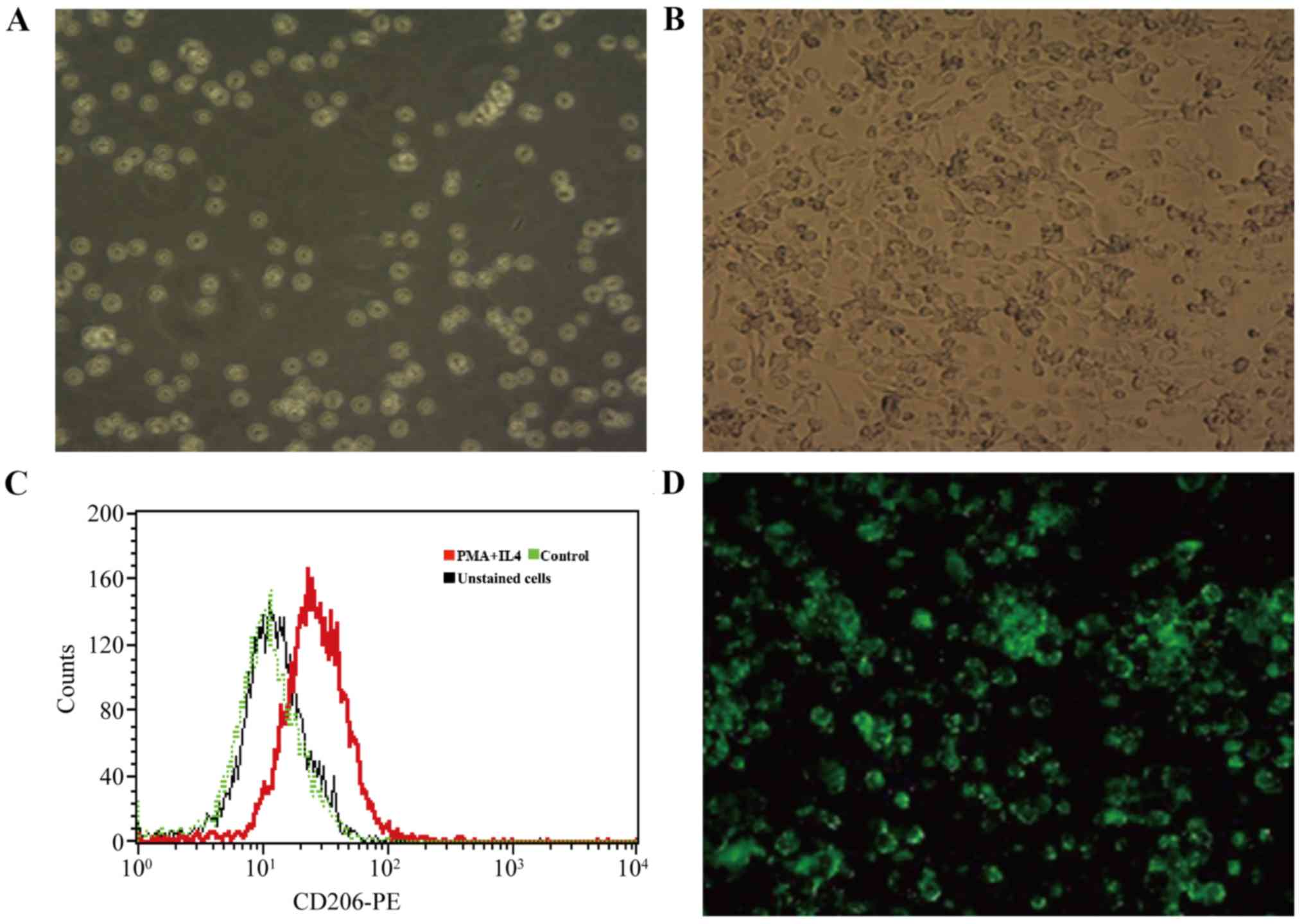
Differentiation of THP-1 cells and
transfection of p-IRES2-ANXA1
PMA and IL-4 were added to the culture medium of
THP-1 cells to induce differentiation to an M2 macrophage
phenotype. THP-1 cells were round or quasi-circular, uniform in
size and suspended in culture medium (Fig. 3A) and became adherent, spindle-shaped
or polygonal cells (Fig. 3B) after
48 h in the culture medium with PMA and IL-4, which expressed CD206
(Fig. 3C) in flow cytometry. These
results indicated that THP-1 cells were successfully induced to
differentiate into M2 macrophages. Successful ANXA1 plasmid
transfection indicated by green fluorescence was observed by
fluorescence microscopy in the pIRES2-ANXA1 group of cells
(Fig. 3D). A similar appearance was
noted in the no-load group. Green fluorescence was absent in the
blank control group. The results of the present study demonstrated
that THP-1 cells transfected with p-IRES2-ANXA1 plasmid could
differentiate into M2 macrophages, which enabled stable expression
of ANXA1.
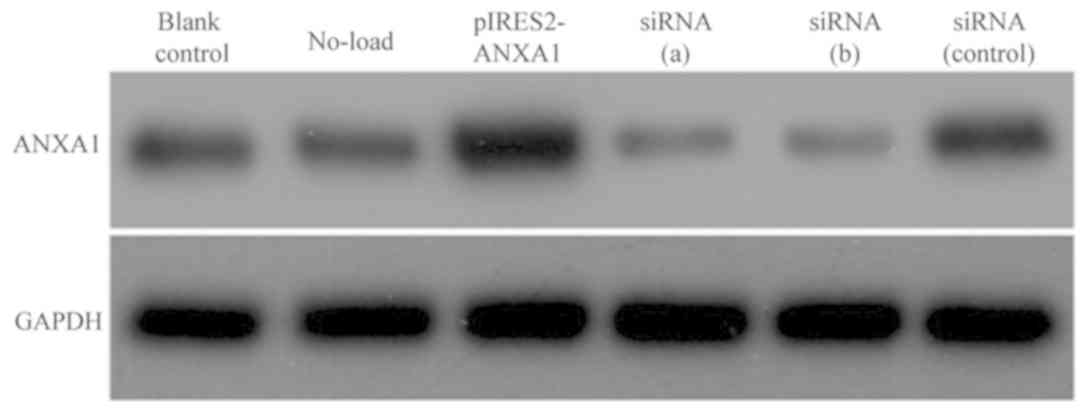
Expression of ANXA1 in M2 macrophages
following transfection
Following transfection of M2 macrophages with the
p-IRES2-ANXA1 plasmid or siRNA against ANXA1, western blot analysis
was used to determine ANXA1 protein expression. The cells
transfected with pIRES2-ANXA1 exhibited a stronger ANXA1 signal
compared with the cells in the blank control and no-load group. In
macrophages transfected with siRNA (a) and siRNA (b), the ANXA1
band was weaker compared with that of the blank control and siRNA
(control) groups (Fig. 4).
Effect of ANXA1 expression on IL-6
protein expression and secretion in macrophages
Western blot results revealed decreased IL-6 protein
expression in macrophages transfected with p-IRES2-ANXA1, while
ANXA1 inhibition caused by siRNA led to enhanced IL-6 expression
(Fig. 5A). The concentration of IL-6
in the culture medium exhibited a similar changing pattern, with a
mean concentration of 215.23±42.11 pg/ml in the blank control
group, 103.62±28.26 pg/ml in the ANXA1 overexpression group,
271.73±67.83 pg/ml in the siRNA (a) group and 264.24±55.97 pg/ml in
the siRNA (b) group (Fig. 5B). The
concentration of IL-6 in the culture medium of the siRNA (control)
and no-load group was 217.71±52.34 and 209.65±42.15 pg/ml,
respectively, which was similar to that in the blank control group,
indicating that ANXA1 inhibited the synthesis of IL-6 in
macrophages and subsequent secretion.
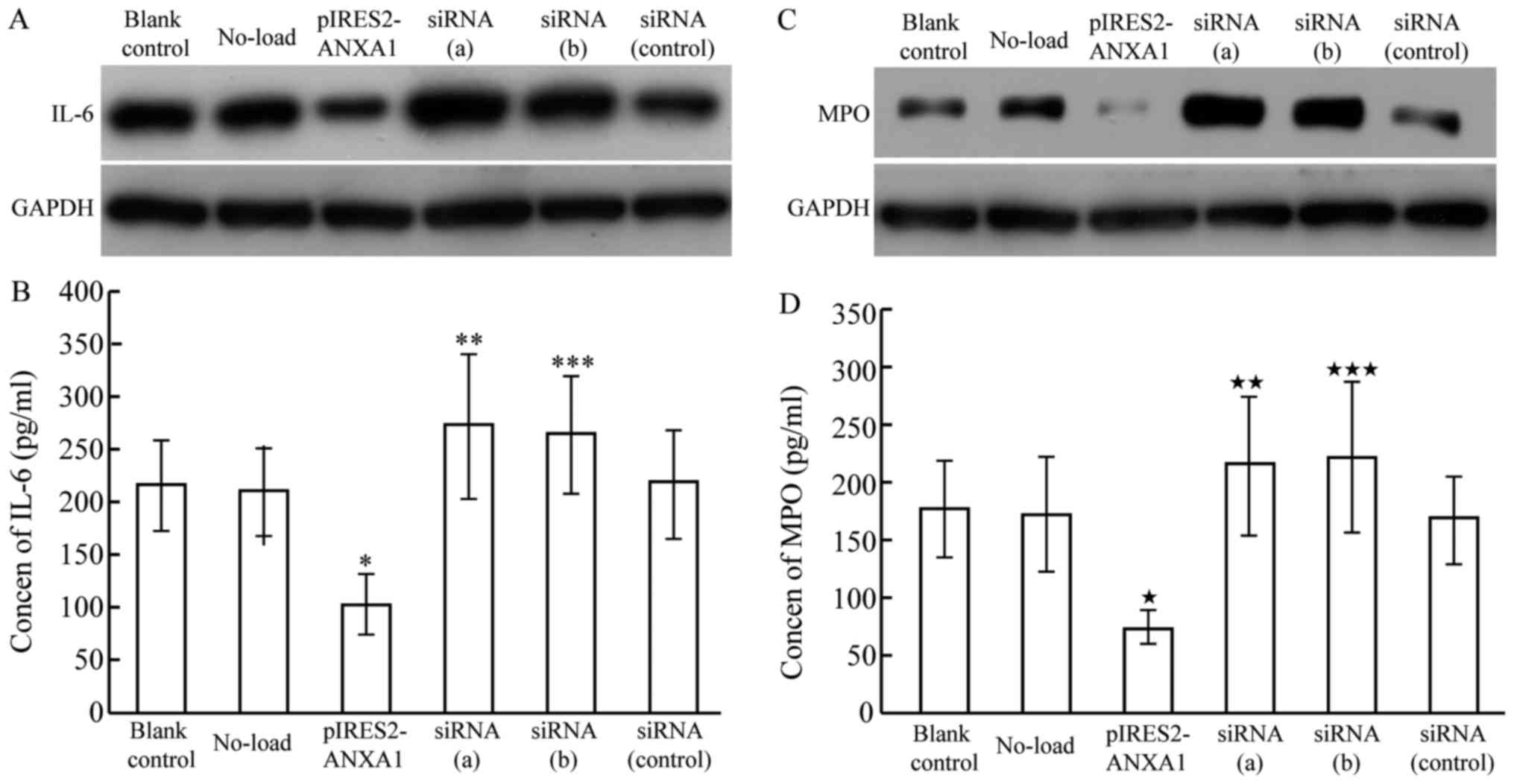 | Figure 5.IL-6 and MPO expression in M2
macrophages transfected with pIRES2-ANXA1 or siRNA against ANXA1,
siRNA (a) and siRNA (b). (A) IL-6 protein expression was decreased
in macrophages with overexpression of ANXA1, while ANXA1 inhibition
induced by siRNA led to increased IL-6 expression. (B) IL-6 in
culture medium was decreased with overexpression of ANXA1
(*P<0.05 compared with blank control) and increased with
siRNA-mediated ANXA1 inhibition (**P<0.05 and ***P<0.05,
compared with blank control). (C) MPO protein expression was
decreased in macrophages with overexpression of ANXA1, while ANXA1
inhibition induced by siRNA led to increased IL-6 expression. (D)
MPO in cell culture medium was decreased with overexpression of
ANXA1 («P<0.01, compared with blank control), and
increased with siRNA-mediated ANXA1 inhibition
(««P<0.05 and «««P<0.05 compared with
blank control). ANXA1, annexin A1; IL-6, interleukin 6; MPO,
myeloperoxidase; siRNA, short interfering RNA. |
Effect of ANXA1 expression on protein
expression and secretion of MPO
MPO protein expression decreased in macrophages
transfected with p-IRES2-ANXA1, while ANXA1 inhibition enhanced MPO
protein expression (Fig. 5C). ELISA
results demonstrated a similar pattern of MPO expression in the
culture medium: 175.32±42.35 pg/ml in the blank control group,
73.04±15.06 pg/ml in the ANXA1 overexpression group, 214.15±59.37
pg/ml in the siRNA (a) group and 221.21±64.87 pg/ml in the siRNA
(b) group (Fig. 5D). The respective
levels in the siRNA (control) and no-load group (167.51±38.26 and
171.92±50.07 pg/ml) were similar to that in the blank control
group, indicating that ANXA1 inhibited the synthesis of MPO in
macrophages and subsequent secretion.
Discussion
To date, efforts to treat acute RILI have focused on
neutralizing pro-inflammatory cytokines or attenuating the
infiltration of inflammatory cells (5). These efforts have been hampered by a
lack of knowledge of the mechanisms of the cytokine or signaling
pathways involved in the pathogenesis of RILI. Thus, no standard
countermeasure or specific drug with a definite target has been
developed. ANXA1 is a member of the calcium-dependent, phospholipid
binding protein superfamily, which was first discovered to be a
secondary messenger downstream of the glucocorticoid receptor
signaling pathway (30). ANXA1 is
critical in the induction and regulation of anti-inflammatory
treatment using glucocorticoids. ANXA1 also takes part in
regulating phagocytosis, differentiation, proliferation, apoptosis
and signaling transduction (31).
Additionally, several animal model studies demonstrated the
anti-inflammatory effect of ANXA1 (32,33).
Treatment of acute RILI using ANXA1 has not been described and its
roles in RILI pathogenesis remain unclear.
The present study observed that glucocorticoid
treatment is associated with increased ANXA1 in plasma. An
increased concentration of ANXA1 in plasma following glucocorticoid
treatment was associated with a more positive treatment outcome for
RILI, indicating that ANXA1 may serve a beneficial role in the
treatment of acute RILI.
ANXA1 in the plasma of patients with RILI was
negatively correlated with the plasma levels of IL-6 and MPO,
implying that ANXA1 may impact RILI treatment outcome by
influencing the expression of these pro-inflammatory cytokines.
IL-6 is an important pro-inflammatory factor, which is synthesized
by activated alveolar macrophages, T helper 2 cells, lung
fibroblasts and alveolar type II epithelial cells. IL-6 is involved
in the responses to lung injury and early-stage inflammation
(34), and promotes the transition
from CD4+ cells to T helper 17 cells, facilitating the
accumulation and activation of neutrophils within the inflammatory
area (35). MPO is mainly expressed
by neutrophils and macrophages. During inflammation, neutrophils
are activated and degranulated, and MPO is secreted into
extracellular matrix.
Extracellular MPO is an alkaline protein with
positive charge, which is easily adsorbed on the surface of
negatively charged cell membrane (36). Hydrogen dioxide (H2O2) in
extracellular matrix can aggregate to the site where the two bind,
thus causing tissue damage (37). In
addition, MPO is absorbed by vascular endothelial cells after being
release from neutrophils, leading to vasculitis and further
aggravating tissue damage (38). It
is well known that radiotherapy can lead to the ionization of water
molecules in the target area to produce H2O2, so reactions
catalyzed by MPO lead to oxidative injury and enhanced
radiation-induced injury to the target tissue (39). In the present study, the baseline
levels of IL-6 and MPO in RILI patients prior to glucocorticoid
treatment were higher compared with those in the healthy controls,
and the expression of the two factors was positively correlated.
These observations are consistent with the previous literature
(40,41). The present findings support a
previous report indicating that ANXA1 overexpression results in
downregulation of IL-6 (42). A
similar correlation between ANXA1 and IL-6 was also observed
following fibroblastoma irradiation treatment (43). ANXA1 can induce apoptosis in
neutrophils (44), and inhibit
neutrophil migration and accumulation in the inflammatory area
(45). In the present study, the
expression of ANXA1 was increased in patients with RILI without an
evident reduction of neutrophils in the circulation system, but
with a significant reduction in the level of MPO. A similar result
has been reported in a previous study (46). In a further study, more apoptotic
neutrophils were observed when the observation time frame was
elongated, possibly due to the impaired function of the releasing
granules of neutrophils following 4 weeks of increased ANXA1
expression, when no evident apoptosis was observed (47).
In the present study, ANXA1 was increased in the
plasma of patients with acute RILI following glucocorticoid
therapy. These results are consistent with several reports
(22,48). The present study also demonstrated
that higher ANXA1 expression was associated with a beneficial
treatment outcome for RILI following glucocorticoid administration,
indicating an increased anti-inflammatory effect. A prior study
reported that the anti-inflammatory effect of glucocorticoid was
associated with induced expression of ANXA1 (49). Further studies demonstrated that
ANXA1 gene knockdown in a mouse model of inflammation markedly
upregulated the pro-inflammatory cytokine IL-6 (50), and an anti-inflammatory effect could
not be achieved with glucocorticoid administration (51). Similar results were acquired when an
antibody against ANXA1 was applied to inhibit its function
(52). These results collectively
demonstrate that the anti-inflammatory effect of glucocorticoid
depends on the induction of ANXA1 expression, highlighting the
potential of this protein as an anti-inflammatory therapy without
the use of hormones.
In the present study, the overexpression of ANXA1 in
macrophages inhibited the synthesis and secretion of IL-6 and MPO,
while the inhibition of ANXA1 promoted these events, indicating
that ANXA1 could serve an anti-inflammatory role, as previously
reported (19,44). ANXA1 alone has been reported to exert
an anti-inflammatory effect. The ANXA1 N-terminal-derived peptide
Ac2-26 can inhibit local and systematic inflammatory responses
(53). Induction of compound 43, an
agonist of the ANXA1 receptor, inhibits the expression of IL-6 in
patients with rheumatoid arthritis and ameliorates bone damage by
reducing the infiltration of inflammatory cells in synovial tissues
(54). ANXA1 inhibits the activity
of calcium-activated, phospholipid-dependent membrane-bound
enzymes, particularly phospholipase A2 (55,56).
ANXA1 and its N-terminal fragment were also demonstrated to inhibit
the synthesis of arachidonic acid derivatives (57,58).
ANXA1 restrains the inflammatory response induced by tumor necrosis
factor-α by suppressing Rac-1-dependent NADPH oxidase in vascular
epithelial cells (59). A more
recent study demonstrated that ANXA1 could exert anti-inflammatory
effects by inhibiting the transcriptional activity of nuclear
factor-κB (NF-κB), as ANXA1 binds specifically to the p65 subunit
of NF-κB, blocking binding to its target genes (49). However, conflicting findings have
been reported. In one study, ANXA1 was found to promote
inflammation by facilitating the transendothelial migration of
neutrophils and monocytes (60).
ANXA1 has also been demonstrated to mediate the activation of the
extracellular signal-regulated kinase, c-Jun N-terminal kinase and
NF-κB inflammatory pathways (61).
Notably, one study indicated a role of endogenous ANXA1 in the
anti-inflammatory effect, while exogenous ANXA1 induced
inflammation (62). It is clear that
more studies are required to clarify the exact role of ANXA1 in
inflammation and RILI pathogenesis.
In conclusion, ANXA1 reduces the expression of IL-6
and inhibits the release of MPO from monocytes and macrophages.
This anti-inflammatory effect may underscore the mechanism by which
ANXA1 in the plasma of patients with RILI is associated with
treatment outcome. Although glucocorticoids are common drugs in
RILI treatment, the present study indicates the potential of ANXA1
to mimic the anti-inflammatory effect of glucocorticoids, which
would potentially avoid certain adverse effects of this
therapy.
Acknowledgements
Not applicable.
Funding
This study was supported by the Scientific Program
of the Health and Family Planning Commission of Jiangsu Province
(grant No. H2017076) and the Foundation of Jiangsu Provincial
Medical Innovation Team (grant No. CXTDA2017042).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
GH and JH conceived and were responsible for the
design of the present study. GS, WX and KL analyzed and interpreted
the patients' data. CD was responsible for the diagnosis of RILI
and assessment of the curative effect based on imaging
examinations. SZ and JY analyzed and interpreted the laboratory
data. GH and SZ were the major contributors in writing the
manuscript. All authors read and approved the present
manuscript.
Ethics approval and consent to
participate
This research was approved by the Ethics Committee
of Taizhou People's Hospital, and written informed consent was
provided by each patient prior to the study.
Patient consent for publication
All patients signed an informed consent for
publication approved by the institutional Review Board.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Graves PR, Siddiqui F, Anscher MS and
Movsas B: Radiation pulmonary toxicity: From mechanisms to
management. Semin Radiat Oncol. 20:201–207. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Han S, Gu F, Lin G, Sun X, Wang Y, Wang Z,
Lin Q, Weng D, Xu Y and Mao W: Analysis of clinical and dosimetric
factors influencing radiation-induced lung injury in patients with
lung cancer. J Cancer. 6:1172–1178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Giridhar P, Mallick S, Rath GK and Julka
PK: Radiation induced lung injury: Prediction, assessment and
management. Asian Pac J Cancer Prev. 16:2613–2617. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Flechsig P, Dadrich M, Bickelhaupt S,
Jenne J, Hauser K, Timke C, Peschke P, Hahn EW, Gröne HJ, Yingling
J, et al: LY2109761 Attenuates radiation-induced pulmonary murine
fibrosis via reversal of TGF-β and BMP-associated proinflammatory
and proangiogenic signals. Clin Cancer Res. 18:3616–3627. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang T, Mathew B, Wu X, Shimizu Y, Rizzo
AN, Dudek SM, Weichselbaum RR, Jacobson JR, Hecker L and Garcia JG:
Nonmuscle myosin light chain kinase activity modulates
radiation-induced lung injury. Pulm Circ. 6:234–239. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao DY, Qu HJ, Guo JM, Zhao HN, Yang YY,
Zhang P, Cao K, Lei X, Cui JG, Liu C, et al: Protective effects of
myrtol standardized against radiation-induced lung injury. Cell
Physiol Biochem. 38:619–634. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kainthola A, Haritwal T, Tiwari M, Gupta
N, Parvez S, Tiwari M, Prakash H and Agrawala PK: Immunological
aspect of radiation-induced pneumonitis, current treatment
strategies, and future prospects. Front Immunol. 8:5062017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ghafoori P, Marks LB, Vujaskovic Z and
Kelsey CR: Radiation-induced lung injury. Assessment, management,
and prevention. Oncology (Williston Park). 22:37–47.
2008.PubMed/NCBI
|
|
9
|
Sun Y, Du YJ, Zhao H, Zhang GX, Sun N and
Li XJ: Protective effects of ulinastatin and methylprednisolone
against radiation-induced lung injury in mice. J Radiat Res.
57:505–511. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oray M, Abu Samra K, Ebrahimiadib N, Meese
H and Foster CS: Long-term side effects of glucocorticoids. Expert
Opin Drug Saf. 15:457–465. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim JH, Jenrow KA and Brown SL: Mechanisms
of radiation-induced normal tissue toxicity and implications for
future clinical trials. Radiat Oncol J. 32:103–115. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu T, Zhang Y, Chang P, Gong S, Shao L and
Dong L: Mesenchymal stem cell-based therapy for radiation-induced
lung injury. Stem Cell Res Ther. 9:182018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ding NH, Li JJ and Sun LQ: Molecular
mechanisms and treatment of radiation-induced lung fibrosis. Curr
Drug Targets. 14:1347–1356. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Straub JM, New J, Hamilton CD, Lominska C,
Shnayder Y and Thomas SM: Radiation-induced fibrosis: Mechanisms
and implications for therapy. J Cancer Res Clin Oncol.
141:1985–1994. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Siva S, MacManus M, Kron T, Best N, Smith
J, Lobachevsky P, Ball D and Martin O: A pattern of early
radiation-induced inflammatory cytokine expression is associated
with lung toxicity in patients with non-small cell lung cancer.
PLoS One. 9:e1095602014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang XJ, Sun JG, Sun J, Ming H, Wang XX,
Wu L and Chen ZT: Prediction of radiation pneumonitis in lung
cancer patients: A systematic review. J Cancer Res Clin Oncol.
138:2103–2116. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brooks AC, Rickards KJ and Cunninggham FM:
Modulation of equine neutrophil adherence and migration by the
annexin-1 derived N-terminal peptide, Ac2-26. Vet Immunol
Immunopathol. 145:214–222. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bena S, Brancaleone V, Wang JM, Perretti M
and Flower RJ: Annexin A1 interaction with the FPR2/ALX receptor:
Identification of distinct domains and downstream associated
signaling. J Biol Chem. 287:24690–24697. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Spurr L, Nadkarni S, Pederzoli-Ribeil M,
Goulding NJ, Perretti M and D'Acquisto F: Comparative analysis of
Annexin A1-formyl peptide receptor 2/ALX expression in human
leukocyte subsets. Int Immunopharmacol. 11:55–66. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Perretti M and Dalli J: Exploiting the
annexin A 1 pathway for the development of novel anti-inflammatory
therapeutics. Br J Pharmacol. 158:936–946. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tucker SL, Jin H, Wei X, Wang S, Martel
MK, Komaki R, Liu HH, Mohan R, Chen Y, Cox JD and Liao Z: Impact of
toxicity grade and scoring system on the relationship between mean
lung dose and risk of radiation pneumonitis in a large cohort of
patients with non-small cell lung cancer. Int J Radiat Oncol Biol
Phys. 77:691–698. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Han G, Lu K, Huang J, Ye J, Dai S, Ye Y
and Zhang L: Effect of Annexin A1 gene on the proliferation and
invasion of esophageal squamous cell carcinoma cells and its
regulatory mechanisms. Int J Mol Med. 39:357–363. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dehai C, Bo P, Qiang T, Lihua S, Fang L,
Shi J, Jingyan C, Yan Y, Guangbin W and Zhenjun Y: Enhanced
invasion of lung adenocarcinoma cells after co-culture with
THP-1-derived macrophages via the induction of EMT by IL-6. Immunol
Lett. 160:1–10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu H, Wang X and Wang W: Functional
suppression of macrophages derived from THP-1 cells by
environmentally-relevant concentrations of arsenite. Comp Biochem
Physiol C Toxicol Pharmacol. 214:36–42. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tedesco S, De Majo F, Kim J, Trenti A,
Trevisi L, Fadini GP, Bolego C, Zandstra PW, Cignarella A and
Vitiello L: Convenience versus biological significance: Are
PMA-differentiated THP-1 cells a reliable substitute for
blood-derived macrophages when studying in vitro polarization?
Front Pharmacol. 9:712018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Draijer C, Boorsma CE, Robbe P, Timens W,
Hylkema MN, Ten Hacken NH, van den Berge M, Postma DS and Melgert
BN: Human asthma is characterized by more IRF5+ M1 and CD206+ M2
macrophages and less IL-10+ M2-like macrophages around airways
compared with healthy airways. J Allergy Clin Immunol.
140:280–283.e3. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Du Q, Tsuboi N, Shi Y, Ito S, Sugiyama Y,
Furuhashi K, Endo N, Kim H, Katsuno T, Akiyama S, et al:
Transfusion of CD206+ M2 macrophages ameliorates antibody-mediated
glomerulonephritis in mice. Am J Pathol. 186:3176–3188. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Udeabor SE, Adisa AO, Orlowska A, Sader RA
and Ghanaati S: Tumor-associated macrophages, angiogenesis, and
tumor cell migration in oral squamous cell carcinoma. Ann Afr Med.
16:181–185. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Petrella A, Festa M, Ercolino SF, Zerilli
M, Stassi G, Solito E and Parente L: Induction of annexin-1 during
TRAIL-induced apoptosis in thyroid carcinoma cells. Cell Death
Differ. 12:1358–1360. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ng FS, Wong KY, Guan SP, Mustafa FB,
Kajiji TS, Bist P, Biswas SK, Wong WS and Lim LH:
Annexin-1-deficient mice exhibit spontaneous airway
hyperresponsiveness and exacerbated allergen-specific antibody
responses in a mouse model of asthma. Clin Exp Allergy.
41:1793–1803. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bizzarro V, Petrella A and Parente L:
Annexin A1: Novel roles in skeletal muscle biology. J Cell Physiol.
227:3007–3315. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Galvão I, Vago JP, Barroso LC, Tavares LP,
Queiroz-Junior CM, Costa VV, Carneiro FS, Ferreira TP, Silva PM,
Amaral FA, et al: Annexin A1 promotes timely resolution of
inflammation in murine gout. Eur J Immunol. 47:585–596. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Locatelli I, Sutti S, Jindal A, Vacchiano
M, Bozzola C, Reutelingsperger C, Kusters D, Bena S, Parola M,
Paternostro C, et al: Endogenous annexin A1 is a novel protective
determinant in nonalcoholic steatohepatitis in mice. Hepatology.
60:531–544. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hong ZY, Song KH, Yoon JH, Cho J and Story
MD: An experimental model-based exploration of cytokines in
ablative radiation-induced lung injury in vivo and in vitro. Lung.
193:409–419. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lowes MA, Russell CB, Martin DA, Towne JE
and Krueger JG: The IL-23/T17 pathogenic axis in psoriasis is
amplified by keratinocyte responses. Trends Immunol. 34:174–181.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Klinke A, Nussbaum C, Kubala L, Friedrichs
K, Rudolph TK, Rudolph V, Paust HJ, Schröder C, Benten D, Lau D, et
al: Myeloperoxidase attracts neutrophils by physical forces. Blood.
117:1350–1358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Klebanoff SJ: Myeloperoxidase: Friend and
foe. J Leukoc Biol. 77:598–625. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jerke U, Rolle S, Purfürst B, Luft FC,
Nauseef WM and Kettritz R: β2 integrin-mediated cell-cell contact
transfers active myeloperoxidase from neutrophils to endothelial
cells. J Biol Chem. 288:12910–12919. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
MacNaughton JI: Regional oxygenation and
radiotherapy: A study of the degradation of infused hydrogen
peroxide. Int J Radiat Biol Relat Stud Phys Chem Med. 19:405–413.
1971. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Barthelemy-Brichant N, Bosquee L, Cataldo
D, Corhay JL, Gustin M, Seidel L, Thiry A, Ghaye B, Nizet M, Albert
A, et al: Increased IL-6 and TGF-β1 concentrations in
bronchoalveolar lavage fluid associated with thoracic radiotherapy.
Int J Radial Oncol Biol Phys. 58:758–767. 2004. View Article : Google Scholar
|
|
41
|
Chen Y, Williams J, Ding I, Hernady E, Liu
W, Smudzin T, Finkelstein JN, Rubin P and Okunieff P: Radiation
pneumonitis and early circulatory cytokine markers. Semin Radiat
Oncol 12 (1 Suppl 1). S26–S33. 2002. View Article : Google Scholar
|
|
42
|
Girol AP, Mimura KK, Drewes CC, Bolonheis
SM, Solito E, Farsky SH, Gil CD and Oliani SM: Anti-inflammatory
mechanisms of the annexin A1 protein and its mimetic peptide Ac2-26
in models of ocular inflammation in vivo and in vitro. J Immunol.
190:5689–5701. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Desai S, Srambikkal N, Yadav HD, Shetake
N, Balla MM, Kumar A, Ray P, Ghosh A and Pandey BN: Molecular
understanding of growth inhibitory effect from irradiated to
bystander tumor cells in mouse fibrosarcoma tumor model. PLoS One.
11:e01616622016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Vago JP, Nogueira CR, Tavares LP, Soriani
FM, Lopes F, Russo RC, Pinho V, Teixeira MM and Sousa LP: Annexin
A1 modulates natural and glueocorticoid-induced resolution of
inflammation by enhancing neutrophil apoptosis. J Leukoc Biol.
92:249–258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Leoni G, Gripentrog J, Lord C, Riesselman
M, Sumagin R, Parkos CA, Nusrat A and Jesaitis AJ: Human neutrophil
formyl peptide receptor phosphorylation and the mucosal
inflammatory response. J Leukoc Biol. 97:87–101. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Dalli J, Norling LV, Renshaw D, Cooper D,
Leung KY and Perretti M: Annexin 1 mediates the rapid
anti-inflammatory effects of neutrophil-derived microparticles.
Blood. 112:2512–2519. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sugimoto MA, Vago JP, Teixeira MM and
Sousa LP: Annexin A1 and the resolution of inflammation: Modulation
of neutrophil recruitment, apoptosis, and clearance. J Immunol Res.
2016:82392582016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen Z, Yoshihara E, Son A, Matsuo Y,
Masutani H, Sugie K, Maeda M and Yodoi J: Differential roles of
Annexin A1 (ANXA1/lipocorton-1/lipomodulin) and thioredoxin binding
protein-2 (TBP-2/VDUP1/TXNIP) in glucocorticoid signaling of
HTLV–I-transformed T cells. Immunol Lett. 131:11–18. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang Z, Huang L, Zhao W and Rigas B:
Annexin 1 induced by anti-inflammatory drugs binds to NF-kappaB and
inhibits its activation: Anticancer effects in vitro and in vivo.
Cancer Res. 70:2379–2388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Yang YH, Aeberli D, Dacumos A, Xue JR and
Morand EF: Annexin-1 regulates macrophage IL-6 and TNF via
glucocorticoid-induced leucine zipper. J Immunol. 183:1435–1445.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Patel HB, Kornerup KN, Sampaio AL,
D'Acquisto F, Seed MP, Girol AP, Gray M, Pitzalis C, Oliani SM and
Perretti M: The impact of endogenous annexin A1 on glucocorticoid
control of inflammatory arthritis. Ann Rheum Dis. 71:1872–1880.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yang YH, Morand EF, Getting SJ, Paul-Clark
M, Liu DL, Yona S, Hannon R, Buckingham JC, Perretti M and Flower
RJ: Modulation of inflammation and response to dexamethasone by
Annexin 1 in antigen-induced arthritis. Arthritis Rheum.
50:976–984. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Stuqui B, de Paula-Silva M, Carlos CP,
Ullah A, Arni RK, Gil CD and Oliani SM: Ac2-26 mimetic peptide of
Annexin A1 inhibits local and systemic inflammatory processes
induced by bothropsmoojeni venom and the lys-49 phospholipase A2 in
a rat model. PLoS One. 10:e01308032015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kao W, Gu R, Jia Y, Wei X, Fan H, Harris
J, Zhang Z, Quinn J, Morand EF and Yang YH: A formyl peptide
receptor agonist suppresses inflammation and bone damage in
arthritis. Br J Pharmacol. 171:4087–4096. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
McArthur S, Gobbetti T, Kusters DH,
Reutelingsperger CP, Flower RJ and Perretti M: Definition of a
novel pathway centered on lysophosphatidic acid to recruit
monocytes during the resolution phase of tissue inflammation. J
Immunol. 195:1139–1151. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Headland SE and Norling LV: The resolution
of inflammation: Principles and challenges. Semin Immunol.
27:149–160. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yang Y, Liu Y, Yao X, Ping Y, Jiang T, Liu
Q, Xu S, Huang J, Mou H, Gong W, et al: Annexin 1 released by
necrotic human glioblastoma cells stimulates tumor cell growth
through the formyl peptide receptor 1. Am J Pathol. 179:1504–1512.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Protzel C, Richter M, Poetsch M, Kakies C,
Zimmermann U, Woenckhaus C, Klebingat KJ, Hakenberg OW and Giebel
J: The role of annexins I, II and IV in tumor development,
progression and metastasis of human penile squamous cell
carcinomas. World J Urol. 29:393–398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Peshavariya HM, Taylor CJ, Goh C, Liu GS,
Jiang F, Chan EC and Dusting GJ: Annexin peptide Ac2-26 suppresses
TNFα-induced inflammatory responses via inhibition of
Rac1-dependent NADPH oxidase in human endothelial cells. PLoS One.
8:e607902013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Williams SL, Milne IR, Bagley CJ, Gamble
JR, Vadas MA, Pitson SM and Khew-Goodall Y: A proinflammatory role
for proteolytically cleaved annexin A1 in neutrophil
transendothelial migration. J Immunol. 185:3057–3063. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Jia Y, Morand EF, Song W, Cheng Q, Stewart
A and Yang YH: Regulation of lung fibroblast activation by annexin
A1. J Cell Physiol. 228:476–484. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yang YH, Song W, Deane JA, Kao W, Ooi JD,
Ngo D, Kitching AR, Morand EF and Hickey MJ: Deficiency of annexin
A1 in CD4+ T cells exacerbates T cell-dependent inflammation. J
Immunol. 190:997–1007. 2013. View Article : Google Scholar : PubMed/NCBI
|