Introduction
Thyroid cancer is one of the most common endocrine
cancers and includes anaplastic carcinoma, medullary carcinoma,
follicular carcinoma and papillary carcinoma (1). Among these, the most common type of
thyroid carcinoma is papillary thyroid carcinoma (PTC), which
accounts for about 85% of all thyroid cancers (2). Thyroid cancer incidence has been
reported to have increased rapidly worldwide, with the PTC
incidence increasing the most (3,4). Between
1975 and 2009, the incidence of thyroid cancer has nearly tripled
from 4.9 to 14.3 per 100,000 individuals, and the increase was
attributable to PTC, the incidence of which increased from 3.4 to
12.5 per 100,000 individuals (5).
Although thyroid cancer typically has low mortality and high
survival rates, disease recurrence is high and a subgroup of
patients with tumours exhibits aggressive characteristics (6). Therefore, it is important to examine
the factors and underlying molecular mechanisms of thyroid cancer
development to further assist in guiding the lifestyle choices of
patients with thyroid cancer.
Numerous studies have investigated potential
prognostic factors of thyroid cancer (7–9); a high
prevalence of BRAF gene mutations (10) and significantly increased expression
levels of the sodium/iodide symporter in thyroid cancer have been
reported (11). Although the exact
mechanism remains unknown, it is unlikely that genetic factors
alone can explain the increase in thyroid cancer incidence in
recent years therefore, an association with other factors has been
proposed (12,13). Previous epidemiological surveys have
indicated that high iodine intake may play an important role in the
promotion of thyroid cancer (14–16).
Iodine is an essential trace element and affects the
function of the thyroid gland (17).
Both deficient and excessive levels of iodine may lead to thyroid
diseases (18). However, the effects
of iodine in thyroid cancer are controversial and whether iodine
promotes or prevents the progression of thyroid cancer remains
unknown. A previous study demonstrated that iodine can induce
apoptosis and prevent the progress of thyroid cancer development
(19). It was further reported that
iodine prevents the transformation from PTC to anaplastic thyroid
cancer (20,21). Gerard et al (22) suggested that thyroid cancer benefits
from iodine deficiency through an angiogenic reaction via vascular
endothelial growth factor (VEGF) induction. However, it was further
reported that iodine may promotes the thyroid cancer incidence rate
(14). An increase in the prevalence
of thyroid cancer has been observed following the introduction of
universal salt iodization (14). The
papillary-to-follicular incidence rate ratios increased
significantly from 3.98 between 1980 and 1984 to 9.88 between 2005
and 2009 (23). Therefore, increased
attention has focused on the mechanism by which iodine affects
thyroid cancer development. For instance, a high iodine diet
promotes thyroid cancer development by upregulating p14ARF and
p16INK4a expression (24). An
association between high iodine intake and the T1799A BRAF mutation
in PTC has been described (25).
Therefore, high iodine intake may be a risk factor in thyroid
cancer development; however, the underlying mechanisms have not
been clearly elucidated and the key genes and pathways involved
remain unknown.
In the present study, the effects of varying extra
amounts of iodine on thyroid cancer cells were examined. In
addition, high throughput RNA-sequencing and RT-qPCR were used to
identify the key genes through which iodine affected thyroid cancer
cells. The phosphatidylinositol 3-kinase/protein kinase B
(PI3K/AKT) signalling pathway in a series of in vitro assays
was examined. The results revealed that SPANXA1 may serve an
important oncogenic role in thyroid cancer cells pre-treated with
extra-low doses of iodine via the PI3K/AKT signalling pathway.
Materials and methods
Cell culture and experimental
groups
Human thyroid cancer cell line BCPAP was purchased
from Jennio Biotech Co., Ltd. The genetic alteration most often
found in thyroid cancer is the mutant BRAF gene; BCPAP cells
harbour the BRAF mutation (26). Therefore, this cell line was selected
to examine the impacts of iodine on the development of thyroid
cancer. Cells cultured in Roswell Park Memorial Institute
(RPMI)-1640 (Corning, Inc.), containing 10% foetal bovine serum
(FBS; Corning, Inc.) was referred to as the control group. A
significant increase in the prevalence of thyroid cancer had been
observed after universal salt iodization (15), with potassium iodate
(KIO3) being the major additive material in the process
of salt iodization (27). Therefore,
KIO3 was dissolved in RPMI-1640 to adjust the
concentration of iodine. LY294002 was used to inhibit the PI3K/AKT
signalling pathway, which may serve an important role in the
effects of iodine on thyroid cancer cell growth. According to the
amount of KIO3 added in to RPMI-1640 medium, the iodine
concentrations in the media were 1.0×10−3,
1.0×10−4, 1.0×10−5, 1.0×10−6,
1.0×10−7 and 1.0×10−8 mol/l. The cells
(3×105 cells/well) were cultured in iodine-enriched
environment with 5% CO2 at 37°C to examine the impact of
iodine. pH values at 37°C of all culture mediums supplemented with
iodine at 1.0×10−3, 1.0×10−4,
1.0×10−5, 1.0×10−6, 1.0×10−7 and
1.0×10−8 mol/l for 24 h were measured; KIO3
did not change the pH value of the culture medium (data not shown).
Iodine supplementation at 1.0×10−6 mol/l promoted cell
proliferation and inhibited cell apoptosis the most previously
compared with control group, therefore, this concentration was used
for subsequent experiments. To detect the expression of PI3K and
p-AKT affected by downregulating SPANXA1, the cells treated with
siRNA-SPANXA1 were compared with control group treated with
siRNA-NC and blank group treated without siRNAs.
Cell proliferation assay
BCPAP cells (3×103) were seeded in
96-well plates in triplicates, and were cultured in 100 µl
RPMI-1640 containing 10% FBS. MTT (Sigma-Aldrich; Merck KGaA) was
added into each well at a final concentration of 5 mg/ml. After 4 h
of MTT incubation with 5% CO2 at 37°C on days 0, 1, 2
and 3, dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) was added to
dissolve the insoluble formazan reduced from MTT, and was
subsequently injected into each well. The absorbance was measured
using the Infinite® 200 pro NanoQuant spectrophotometer
(Tecan Group, Ltd.) at a wavelength of 490 nm to determine cell
proliferation.
Apoptosis analysis
BCPAP cells (3×105) were seeded into
6-well plates with RPMI-1640 complete culture medium. After 48 h of
treatment, the cells were washed with PBS, suspended in binding
buffer (Beyotime Institute of Biotechnology) and serum-deprived for
24 h prior to FACS. The cells were stained with 5 µl Annexin
V-fluorescein isothiocyanate (FITC) and 5 µl propidium iodide
(Vazyme Biotech Co., Ltd.) and incubated at room temperature for 15
min, according to the manufacturer's protocol. After staining, the
proportion of apoptotic cells was analysed using a flow cytometer
(BD Biosciences).
High-throughput RNA-sequencing
High-throughput RNA-sequencing was conducted
(Seqhealth Technology Co., Ltd.) to screen differentially expressed
genes between BCPAP cells incubated in ordinary medium and the ones
treated with extra-lower iodine. Total RNA was isolated using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and
purified using an RNeasy Plus Mini kit (Qiagen). Each group was
sequenced on one sequencing lane of an Illumina Genome Analyzer II
system (Illumina). The paired-end reads were trimmed with the
Illumina standard adapter (‘AGATCGGAAGAGC’) low quality sequence
3end by using Trim Galore (version 3, http://www.bioinformatics.babraham.ac.uk/projects/trim_galore)
and mapped to the human genome (version h19; ftp://ftp.ensembl.org/pub/release-75/gtf/homo_sapiens/Homo_sapiens.GRCh37.75.gtf.gz)
using Tophat (version 2.1.0, http://ccb.jhu.edu/software/tophat/index.shtml). The
expression levels of each gene were analysed using Cufflinks
(version 2.21, http://cole-trapnell-lab.github.io/cufflinks). The
mapped reads were used to quantify the transcripts from the RefSeq
reference database. For the functional annotation analysis of
genes, the Database for Annotation, Visualization and Integrated
Discovery (DAVID, http://david.ncifcrf.gov/home.jsp) online tool was
used.
SPANXA1-knockdown by transient RNA
interference
BCPAP cells (3×105) were seeded into
6-well plates and incubated for 24 h. Cells were transiently
transfected with synthesized SPANXA1 small interfering (si)RNAs
(GCCTGCCACTGACATTGAA, 20 µM; Ribobio Co., Ltd.) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Red fluorescent protein (RFP) was used to detect
transfection efficiency. Media were changed at 4–6 h. Cells were
harvested 48 h after transfection, and reverse
transcription-quantitative PCR (RT-qPCR) was performed to confirm
transfection.
Wound healing assay
BCPAP cells in logarithmic growth phase were placed
in 6-well plates with a density of 3×105 cells/well.
Once the cells reached 80% confluence, a vertical scratch was made
on the cell layer using a 10-µl pipette tip. Thereafter, the cells
were incubated in serum-free RPMI-1640 with 5% CO2 at
37°C. Images of the plates were captured at 0, 24 and 48 h at ×20
magnification using an inverted microscope (Olympus Corporation,
Tokyo, Japan).
Cell migration assay
Transwell assays were performed to evaluate the
invasive capability of BCPAP cells. Transwell chambers with
polyvinylidene difluoride (PVDF) filters (pore size 8.0-µm;
Corning, Inc.) were pre-coated with 50 µl Matrigel (BD Biosciences)
diluted 1:3 in serum-free RPMI-1640. Cells (3×104
cells/well) were suspended in 200 µl serum-free RPMI-1640 and
placed into the upper chambers in triplicate. RPMI-1640 containing
600 µl FBS was added to the lower chamber as a chemotactic factor.
Following 48-h incubation, the non-invading cells on the upper
surface were carefully removed with cotton swabs. Cells that
migrated through the pores and adhered to the lower surface were
fixed with 4% paraformaldehyde for 30 min at room temperature and
stained with 0.1% crystal violet for 30 min at room temperature,
and images were captured under a fluorescent microscope (Olympus
Corporation).
RT-qPCR analysis
Total RNA extraction from the cells was performed
using TRIzol kit (Invitrogen; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The obtained RNA was
reversely transcribed into cDNA (10 µl) using a PrimeScript RT
Reagent kit (Takara, Bio, Inc.). cDNA was used as the template for
qPCR detection with primers and SYBR Master Mixture (Takara Bio,
Inc.). The PCR was performed using the VIIA7 Real-Time PCR System
(Thermo Fisher Scientific, Inc.), and the thermocycling conditions
were as follows: Pre-denaturation at 95°C for 2 min, 40 cycles of
denaturation at 95°C for 15 sec and annealing at 60°C for 15 sec,
followed by final extension at 95°C for 15 sec. The relative mRNA
expression levels in each sample were calculated using the
2−ΔΔCq method. The primers were synthesized by Generay
Biotech Co., Ltd., and the sequences are listed in Table I. mRNA levels were normalized to the
internal reference gene β-actin.
 | Table I.Primer sequences used in the present
study. |
Table I.
Primer sequences used in the present
study.
| Gene | Primers
(5′→3′) |
|---|
| BMS1P17 | F:
GCACAGGCTGCATCTCCACTA |
|
| R:
CCCACGGCATCAGACTAAAGG |
| CALD1 | F:
GTTTCCATCTGGGGTTTTAGTT |
|
| R:
TATGTGGGAGAAAGGGAATGT |
| H3F3BP1 | F:
CTGGAAGGGAAGTCTGCGAAT |
|
| R:
TACTGGAGGGGTGAAGAAACC |
| IGKV10R-2 | F:
GGAGTTTTCCTTGGTTTCTGC |
|
| R:
AGTCTCCATCCTCCCTGTCTG |
| MKI67 | F:
AAGCCCTCCAGCTCCTAGTCCTA |
|
| R:
GCCACTCTTTCTCCCTCCTCTCT |
| SACS | F:
CCAGGTGGTAAAGGAAGGAAA |
|
| R:
GTGGGCGAGGGATCAGTAGTA |
| VCAN | F:
GTATTTGTAGCACTGCCCTTG |
|
| R:
TGTCACTCTAATCCCTGTCGT |
| SPANXA1 | F:
TTCCTCCTGTAGCGAACCACT |
|
| R:
TGCCACTGACATTGAAGAACC |
| β-actin | F:
CATGTACGTTGCTATCCAGGC |
|
| R:
CTCCTTAATGTCACGCACGAT |
Western blot analysis
Total protein was extracted from BCPAP cells using
RIPA lysis buffer (Beyotime Institute of Biotechnology), and its
concentration was determined using a bicinchoninic acid assay.
Extracted protein (~25 µg) in each group was denatured at 95°C for
10 min. Proteins were separated by 10% SDS-PAGE and transferred
onto PVDF membranes (100V; 60 min). The membranes were blocked for
2 h in Tris-buffered saline with Tween-20 (TBST) with 5% non-fat
milk at room temperature, and were subsequently incubated at 4°C
overnight with primary antibodies against PI3K (cat. no. 4228;
dilution, 1:1,000), phosphorylated-AKT (p-AKT; cat. no. 9271;
dilution, 1:500), AKT (cat. no. 9272; dilution, 1:500) and GADPH
(cat. no. 5174; dilution, 1:1,000). Following the incubation, the
membranes were washed three times using TBST and incubated with
horseradish peroxidase-conjugated anti-rabbit immunoglobulin G
secondary antibody (cat. no. 5151; dilution, 1:2,000) at room
temperature for 1 h. All antibodies were purchased from Cell
Signaling Technology, Inc. The membranes were washed with TBST, and
the blots were examined by enhanced chemiluminescence. Protein
expression levels were semi-quantified by densitometry analysis
using an imaging system (LI-COR, Inc.).
Statistical analysis
All statistical analysis was performed using SPSS
18.0 software (SPSS, Inc.). The results were presented as the mean
± standard deviation as appropriate. All calculated significances
are based on the one-way analysis of variance test and the Tukey's
post-hoc test. For direct comparisons between two groups, paired
Student's t-test was used. P<0.05 was considered to indicate a
statistically significant difference.
Results
Iodine at 1.0×10−6 mol/l
significantly promotes cell proliferation and inhibits
apoptosis
Cell proliferation, as evaluated by an MTT assay,
suggested that iodine at 1.0×10−4, 1.0×10−5,
1.0×10−6, 1.0×10−7 and 1.0×10−8
mol/l enhanced cell proliferation when compared with the control
group. Significant increases in cell proliferation compared with
the control were observed following treatment with
1.0×10−6 mol/l. However, cells incubated with iodine at
1.0×10−3 mol/l presented notably decreased cell growth
compared with the control group (Fig.
1A). For apoptosis, these results were reversed. Apoptosis in
BCPAP cells incubated with iodine at 1.0×10−4,
1.0×10−5, 1.0×10−6, 1.0×10−7 and
1.0×10−8 mol/l was significantly decreased compared with
the control group; the lowest degree of apoptosis was observed in
response to 1.0×10−6 mol/l iodine. Apoptosis in samples
treated with iodine at 1.0×10−3 mol/l significantly
increased compared with the control (Fig. 1B).
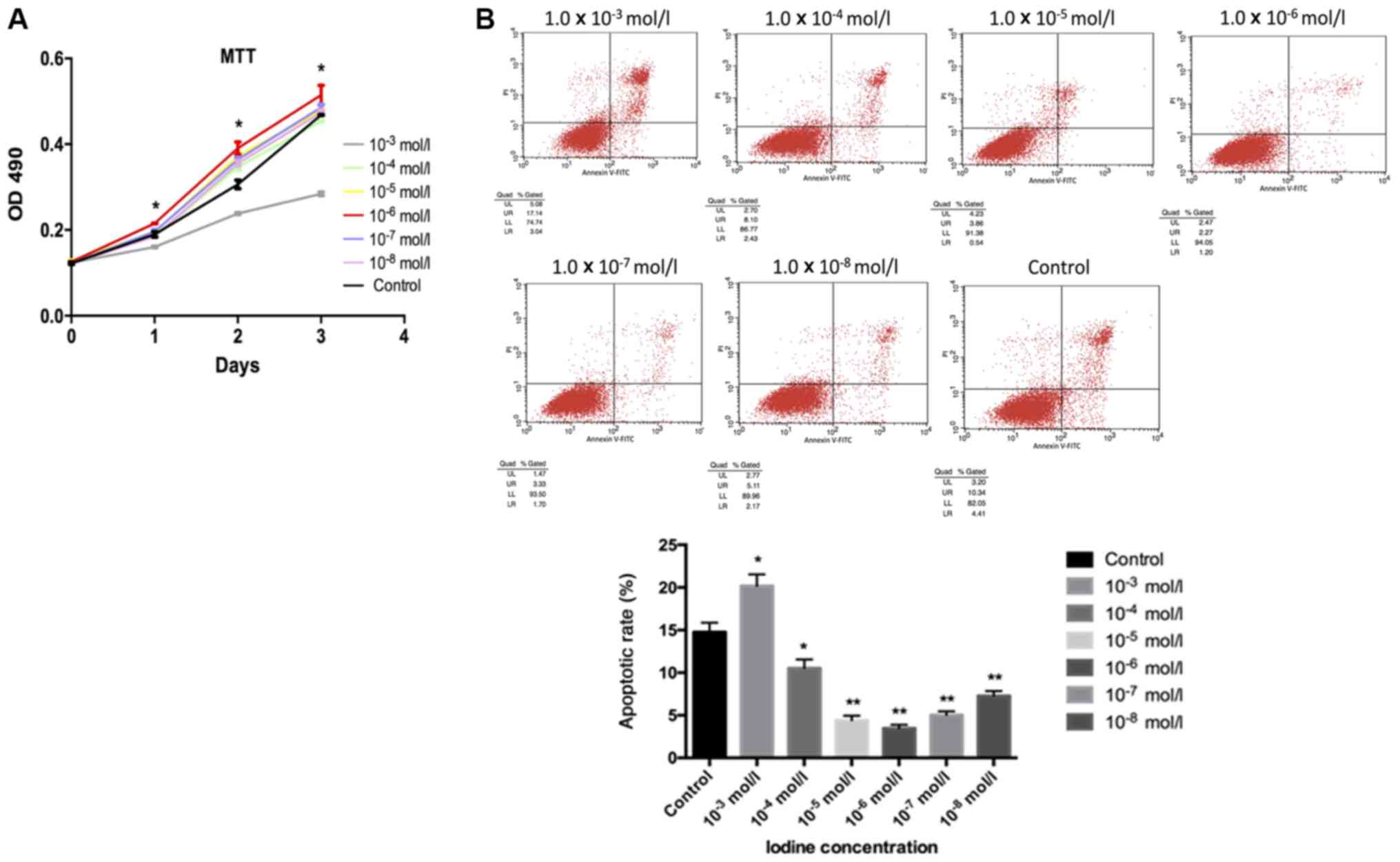 | Figure 1.Cell proliferation evaluated by MTT
assay and flow cytometry. (A) It was observed that iodine at lower
concentrations (1.0×10−4, 1.0×10−5,
1.0×10−6, 1.0×10−7 and 1.0×10−8
mol/l) contributed to the proliferation of BCPAP cells, while
iodine of a higher concentration (1.0×10−3 mol/l) had a
negative effect. Cells treated with iodine at 1.0×10−6
mol/l exhibited the most pronounced changes. (B) BCPAP cell
apoptosis was analysed by flow cytometry with different
concentrations of iodine. BCPAP cell apoptosis, when incubated with
lower concentrations of iodine (1.0×10−4,
1.0×10−5, 1.0×10−6, 1.0×10−7 and
1.0×10−8 mol/l) was significantly inhibited. Among
these, the highest inhibitory effect was observed with
1.0×10−6 mol/l iodine. A higher concentration of iodine
(1.0×10−3 mol/l) appeared to promote cell apoptosis.
*P<0.05 and **P<0.01 vs. the control group. OD, optical
density. |
High throughput RNA-sequencing shows
that iodine at 1.0×10−6 mol/l promotes thyroid cancer
development via SPANXA1
Our results indicated that iodine exhibits important
roles in thyroid cancer, particularly at 1.0×10−6 mol/l.
In order to investigate the molecular mechanism by which iodine
affects thyroid cancer cells, high throughput RNA-sequencing was
utilized to screen for differentially expressed genes in BCPAP
cells incubated in ordinary medium (control group) and cells
cultured in iodine-enriched medium (1.0×10−6 mol/l). The
differential expression of genes was confirmed via RT-qPCR
(Fig. 2). It was revealed that
SPANXA1 mRNA expression was significantly upregulated in BCPAP
cells when treated with iodine.
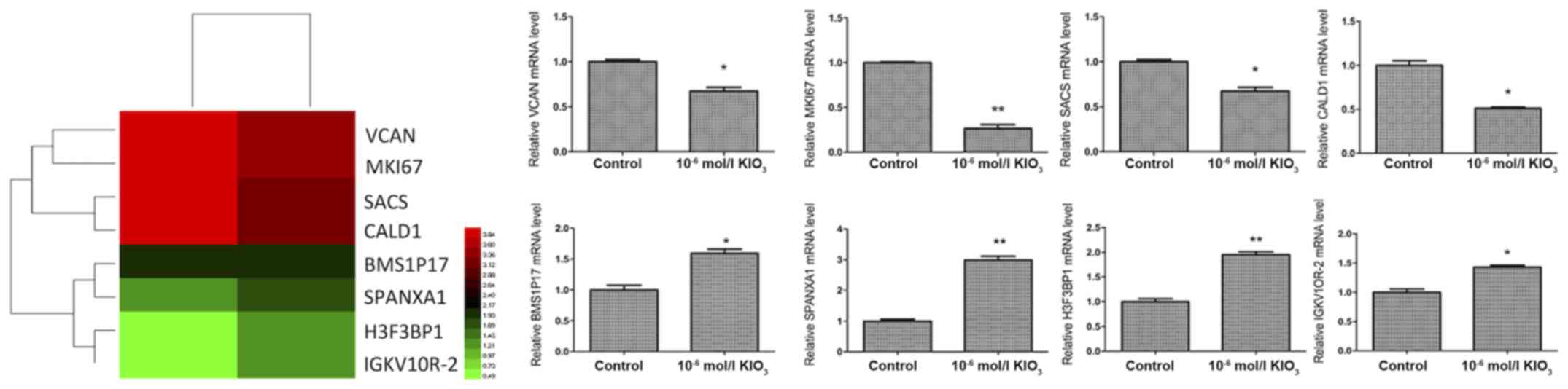 | Figure 2.High throughput RNA-sequencing data
shown as a heat map and expression of different genes in BCPAP
cells treated with iodine at 1.0×10−6 mol/l. Red
represents high expression, and green represents low expression.
*P<0.05 and **P<0.01 vs. the control group. KIO3, potassium
iodate; BMS1P17, BMS1, ribosome biogenesis factor pseudogene 17;
CALD1, caldesmon 1; H3F3BP1, H3 histone, family 3B pseudogene 1;
MKI67, marker of proliferation Ki-67; SACS, sacsin molecular
chaperone; SPANXA1, sperm protein associated with the nucleus,
X-linked, family member A1; VCAN, versican. |
Downregulation of SPANXA1 inhibits
cell proliferation, migration and invasion, and promotes cell
apoptosis in thyroid cancer cells incubated with an extra-low dose
of iodine
BCPAP cells incubated with iodine at
1.0×10-6 mol/l were selected to be transfected with 50
nmol/l siRNA-SPANXA1 to inhibit SPANXA1 expression (Fig. 3A). There was no notable difference in
efficiency between siRNA-NC group and siRNA-SPANXA1 group.
Therefore, the present study proposed that alterations in the
parameters assessed may be induced by siRNA-SPANXA1. After
transfection, SPANXA1 mRNA expression was downregulated
significantly, which was evaluated by RT-qPCR (Fig. 3B). As presented in Fig. 3C, the proliferation of transfected
BCPAP cells was lower when compared with the control group. The
results demonstrated that migration (Fig. 3D) and invasion (Fig. 3E) of the siRNA-SPANXA1-transfected
BCPAP cells were also significantly decreased compared with the
control group. The apoptotic rate of the siRNA-SPANXA1-transfected
BCPAP cells significantly increased when compared with the control
group (13.05 vs. 9.35%), which implied that downregulating SPANXA1
may significantly induce BCPAP cell apoptosis (Fig. 3F).
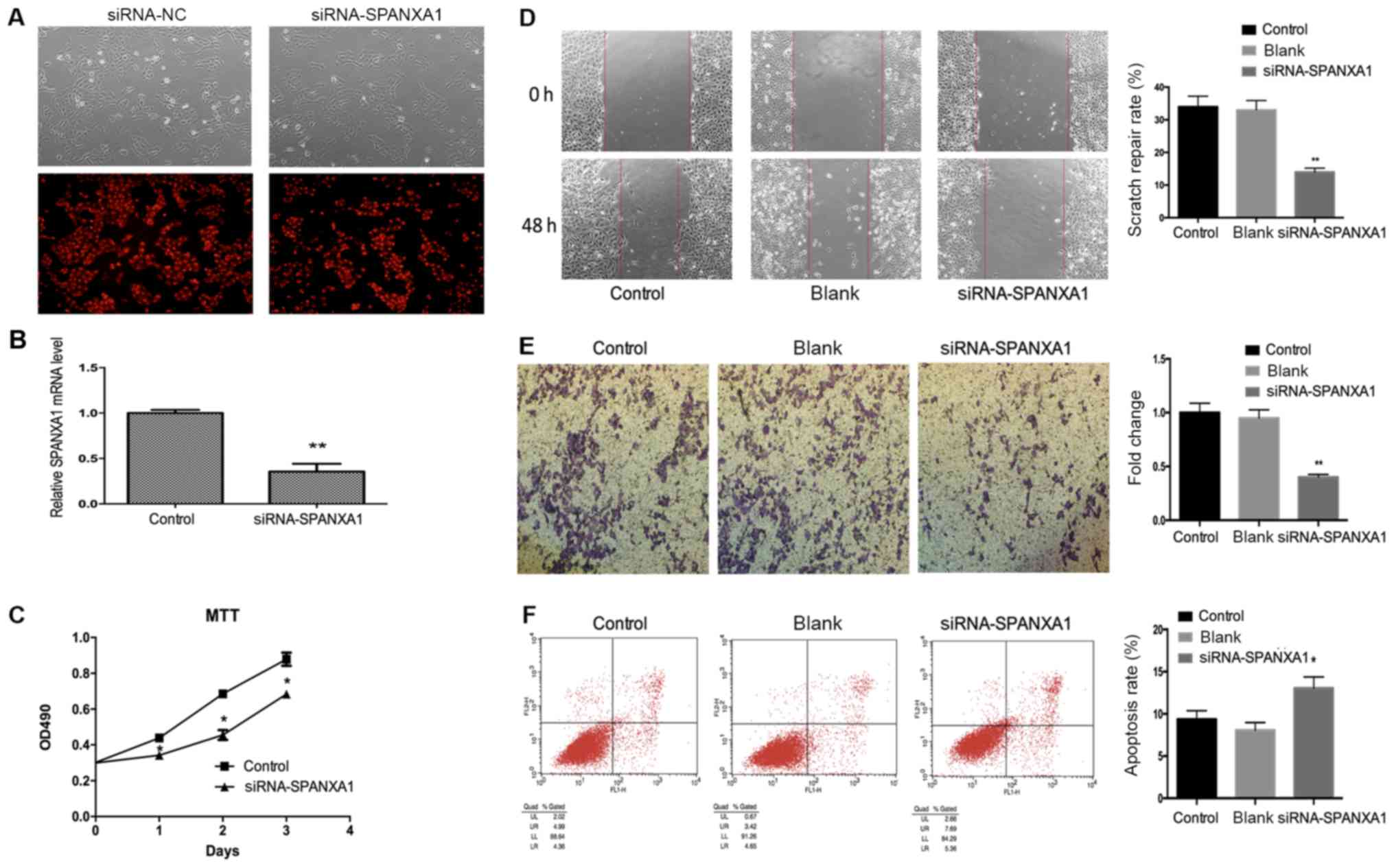 | Figure 3.Effects of SPANXA1 on BCAP cell
proliferation, migration, invasion and apoptosis. (A) Microscopy
images of cells transfected with 50 nmol/l siRNA and control cells
(magnification, ×400). (B) After transfection, SPANXA1 expression
was decreased. (C) Proliferation of BCPAP cells treated with iodine
at 1.0×10−6 mol/l as assessed by an MTT assay;
SPANXA1-knockdown BCPAP cells exhibited suppressed proliferation.
Migration of BCPAP cells with downregulated SPANXA1 was evaluated
using (D) a Transwell assay and (E) invasion was analysed using a
scratch-wound assay. Downregulation of SPANXA1 contributed to the
inhibition of migration and invasion of iodine-treated BCPAP cells.
Magnification, ×20. (F) Cell apoptosis results were obtained by
flow cytometry and suggested that downregulation of SPANXA1
increased apoptosis in iodine-treated BCPAP cells
(1.0×10−6 mol/l). *P<0.05 and **P<0.01 vs. the
control group. NC, negative control; SPANXA1, sperm protein
associated with the nucleus, X-linked, family member A1; si, small
interfering; OD, optical density. |
Iodine promotes thyroid cancer
development via SPANXA1 through the PI3K/AKT signalling
pathway
As presented in Fig.
4A, PI3K and p-AKT expression was downregulated following
SPANXA1 knockdown. PI3K and p-AKT expression were detected in the
control and iodine treatment (1.0×106 mol/l) groups. It
was observed that PI3K and p-AKT expression levels were upregulated
in the iodine treatment group compared with the control group
(Fig. 4B). In addition, LY294002, a
specific PI3K inhibitor, was added to the iodine-treated cells
(1.0×10−6 mol/l; with high expression of SPANXA1). The
effects of the inhibitor on the PI3K/AKT signalling pathway were
confirmed by an MTT assay (Fig. 4C),
scratch (Fig. 4D) and Transwell
(Fig. 4E) assays, as well as by flow
cytometry (Fig. 4F). Downregulation
of the PI3K/AKT signalling pathway inhibited cell proliferation,
migration, invasion and promoted cell apoptosis. Our results
suggested that the inhibition of the PI3K/AKT signalling pathway
reversed the SPANXA1-promoted development of BCPAP cells
pre-treated with iodine.
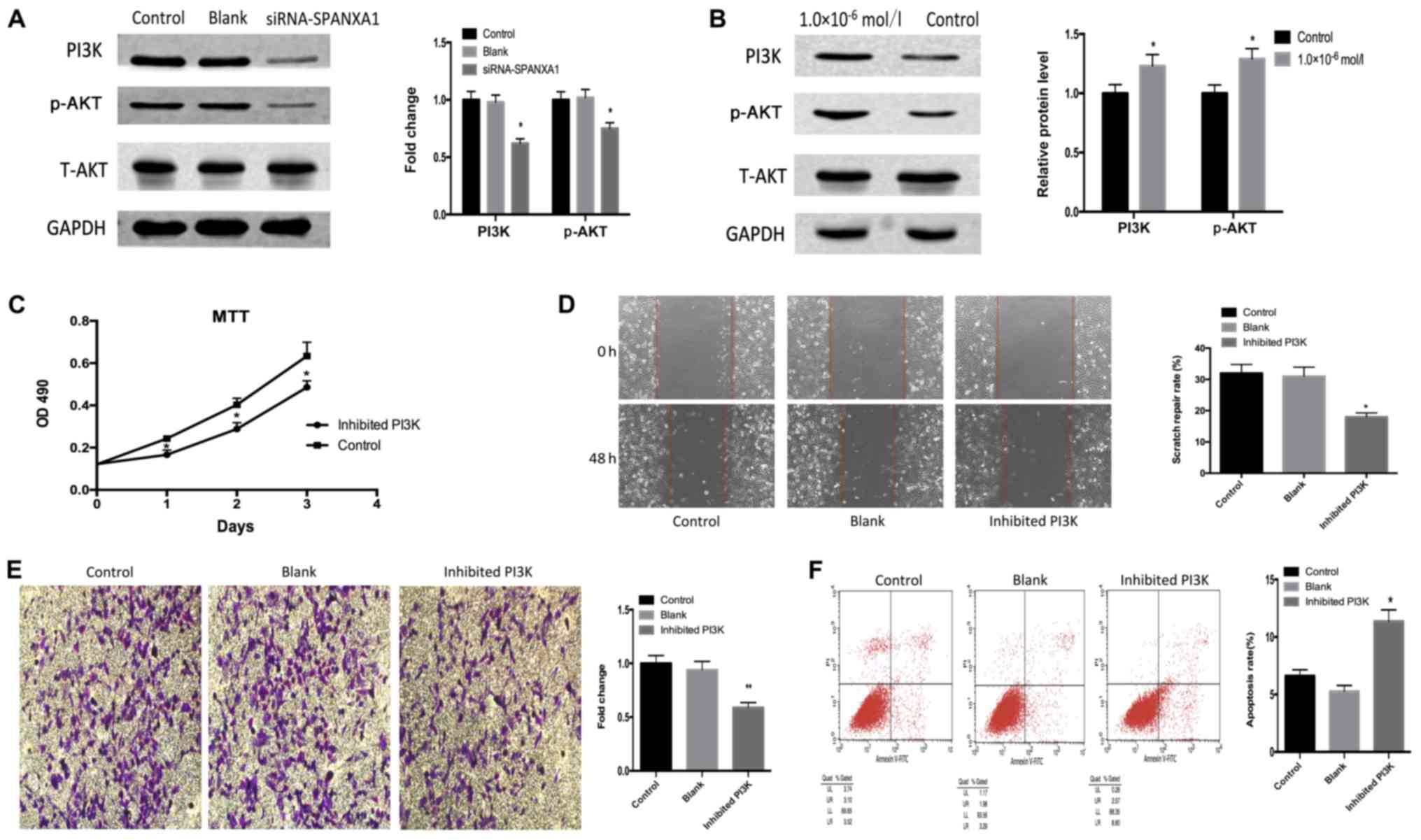 | Figure 4.Effects of the PI3K/AKT signalling
pathway. (A) Downregulation of SPANXA1 suppresses PI3K and p-AKT
expression in iodine-treated BCPAP cells (1.0×10−6
mol/l). (B) PI3K and p-AKT protein expression were increased in
BCPAP cells incubated with iodine at 1.0×10−6 mol/l.
Applying PI3K inhibitor LY294002 to iodine-treated cells suppressed
biological behaviours, including cell proliferation, invasion and
migration, but promoted apoptosis, as demonstrated by (C) MTT, (D)
wound healing and (E) Transwell assays, as well as (F) flow
cytometry, respectively. Magnification, ×20. *P<0.05 and
**P<0.01 vs. the control group. SPANXA1, sperm protein
associated with the nucleus, X-linked, family member A1; p,
phosphorylated; PI3K, phosphoinositide 3-kinase; t, total. |
Discussion
Thyroid cancer is a common endocrine cancer and its
etiological factors include radiation, the environment and genetics
(1,10,13). An
epidemiological study showed that high urinary iodine is a risk
factor for thyroid tumorigenesis (28). Another study demonstrated that iodine
regulates G2/M progression, induced by C-C motif ligand
21/C-C motif chemokine 7 interactions in primary cultures of
thyroid cancer cells with RET/PTC expression (29). A study using rats revealed that low
and high iodine diets reduced the expression of p14ARF and
p16INK4a, and promoted thyroid cancer development (24). Therefore, the association between
iodine and thyroid cancer has become a key area of interest within
the field. However, the underlying mechanisms by which iodine
affects thyroid cancer cells remain unclear.
The present study investigated how iodine affected
the physiological features of thyroid cancer cells in vitro,
including proliferation and apoptosis. The results indicated that
iodine served a dual role in the proliferation and apoptosis of
thyroid cancer cells. Compared with the control group, extra-high
doses of iodine (1.0×10−3 mol/l) inhibited cell
proliferation and promoted cell apoptosis, while extra-low doses of
iodine (1.0×10−4−1.0×10−8 mol/l) exhibited
opposing effects. In particular, proliferation in cells incubated
with iodine at 1.0×10−6 mol/l increased significantly.
The experimental outcomes suggested that extra-low doses of iodine
may be unfavourable for patients with thyroid cancer. Therefore,
iodine intake should be restricted, since extra iodine may expedite
tumour progression.
Thyroid cancer cells cultured with iodine at
1.0×10−6 mol/l were selected for subsequent experiments,
investigating prominent biological alternations induced by iodine.
High throughput RNA-sequencing results indicated that the level of
SPANXA1 was increased in thyroid cancer cells treated with this
specific concentration of iodine. It should be noted that the
effects of the SPANX gene family on tumour cells have been reported
for various cancers, including melanoma (30), myeloma and haematological
malignancies (31). High SPANX gene
expression promotes the progression of various cancers under most
circumstances. For instance, experimental evidence suggested that
SPANX-A/C/D promotes breast cancer progression by regulating
complementary cellular functions to induce its invasiveness
(32). Furthermore, there may be a
potential association between SPANX overexpression and prostate
cancer development (33). To the
best of our knowledge, less attention has been paid to the SPANXA1
gene in previous cancer research. In melanoma and glioblastoma cell
lines, SPANXA1 is the most frequently expressed SPANX variant
(34). Previous studies demonstrated
that SPANXA1 expressed in thyroid tissue and thyroid cancer
(35,36). Our findings regarding the connection
between SPANXA1 and thyroid cancer complemented the exploration of
thyroid cancer from a genic point of view.
To confirm the role of SPANXA1, its expression was
downregulated in thyroid cancer cells through transfection. It was
found that transfected thyroid cancer cells exhibited reduced cell
proliferation, migration and invasion and increased thyroid cancer
cell apoptosis. These findings indicated that SPANXA1 was one of
the key genes, which enhanced the process of tumour growth in cells
treated with an extra-low dose of iodine. High levels of SPANX in
tumour cells have been detected in various cancers (32,33). The
use of SPANX as a prognostic marker may be promising; however,
little is known about the cellular behaviour during the regulation
of SPANX expression. In this context, the tumour-suppressive role
of downregulated SPANX was supported by transfection experiments.
In conjunction with increased gene expression in thyroid cancer
cells treated with iodine, these results may suggest the
involvement of SPANXA1 in iodine-mediated thyroid cancer
development, yet the potential effects of SPANXA1 on other tumour
cells require further investigation.
After documenting the high prevalence of SPANXA1 in
iodine-treated thyroid cancer, the signalling pathway by which
SPANXA1 exerts its oncogenic action on thyroid cancer cells treated
with extra-low doses of iodine was elucidated. PI3K/AKT, a
classical signalling pathway, is involved in the regulation of
various human cancers (37).
Activating PI3K stimulates the phosphorylation of AKT and therefore
influences the biological behaviour of tumours (38,39). It
has been previously reported that the PI3K/AKT signalling pathway
was connected to thyroid cancers (40,41). It
was demonstrated that leptin is involved in thyroid cancer
pathogenesis through the PI3K/AKT signalling pathway via the
obesity receptor (42). Another
study proposed that miR-34a regulates growth arrest specific 1
expression to promote proliferation and suppress apoptosis of
thyroid cancer cells via the PI3K/AKT/BAD signalling pathway
(43). A previous study established
that p-53-inducible gene 3 plays an oncogenic role in thyroid
cancer via the regulation of the PI3K/AKT/PTEN signalling pathway
(44).
The PI3K/AKT signalling pathway is involved in the
thyroid autoregulation induced by iodide (45), and therefore, it is proposed that the
PI3K/AKT signalling pathway is highly likely to be involved in
thyroid cancer development in iodine-rich environments. To confirm
this, the association between SPANXA1 expression and protein
targets of the PI3K/AKT signalling pathway were examined in thyroid
cancer cells. PI3K and p-AKT exhibited increased expression in
thyroid cancer cells treated with an extra-low dose of iodine.
SPANXA1 gene silencing in these cells resulted in the
downregulation of PI3K and p-AKT expression. These results revealed
that the PI3K/AKT signalling pathway was involved in the
progression of SPANXA1-mediated thyroid cancer cells, pre-treated
with extra-low doses of iodine. Data analysis was further conducted
to elucidate whether the PI3K/AKT signalling pathway is the key
signalling pathway through which SPANXA1 affects thyroid cancer
cells treated with extra-low doses of iodine. Inhibiting PI3K,
without changing SPANXA1 expression, reduced cell proliferation,
migration and invasion, and promoted apoptosis. This highlighted
the importance of the PI3K signalling pathway in the development of
iodine-treated thyroid cancer cells.
In summary, extra-low doses of iodine promoted the
proliferation and inhibited the apoptosis of thyroid cancer cells,
particularly at 1.0×10−6 mol/l. The molecular mechanisms
behind this phenomenon were examined by a series of experiments
using thyroid cancer cells treated with iodine at this specific
concentration. The SPANXA1 gene was discovered to be responsible
for the development of iodine-treated thyroid cancer cells. The
high expression of SPANXA1 promoted cell proliferation and
inhibited cell apoptosis. In addition, PI3K/AKT was proposed to be
a key signalling pathway through which SPANXA1 mediates its
effects. The results suggested that SPANXA1 may be a biomarker in
thyroid cancer and may help in developing effective dietary plans
for patients with thyroid cancer, in which patients should limit
excessive iodine intake.
Acknowledgements
Not applicable.
Funding
This research was supported by the National Natural
Science Foundation of China (grant no. 81673108 to HQ) and the
Science and Technology Planning Project of Harbin, China (grant no.
2016RAXYJ088 to HQ).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HQ designed the present study. XY participated in
study design and drafted the manuscript. JS and JH performed the
literature research and statistical analysis. LS and HW performed
the western blot experiments. DZ and QF analysed the data. JL
performed the reverse transcription-quantitative PCR
experiments.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nikiforov YE, Steward DL, Robinson-Smith
TM, Haugen BR, Klopper JP, Zhu Z, Fagin JA, Falciglia M, Weber K
and Nikiforova MN: Molecular testing for mutations in improving the
fine-needle aspiration diagnosis of thyroid nodules. J Clin
Endocrinol Metab. 94:2092–2098. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ahn HS, Kim HJ and Welch HG: Korea's
thyroid-cancer ‘epidemic’-screening and overdiagnosis. N Engl J
Med. 371:1765–1767. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oh CM, Won YJ, Jung KW, Kong HJ, Cho H,
Lee JK, Lee DH and Lee KH; Community of Population-Based Regional
Cancer Registries, : Cancer statistics in Korea: Incidence,
mortality, survival, and prevalence in 2013. Cancer Res Treat.
48:436–450. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Davies L and Welch HG: Current thyroid
cancer trends in the United States. JAMA Otolaryngol Head Neck
Surg. 140:317–322. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Solis OE, Mehta RI, Lai A, Mehta RI,
Farchoukh LO, Green RM, Cheng JC, Natarajan S, Vinters HV,
Cloughesy T and Yong WH: Rosette-forming glioneuronal tumor: A
pineal region case with IDH1 and IDH2 mutation analyses and
literature review of 43 cases. J Neurooncol. 102:477–484. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zarkesh M, Zadeh-Vakili A, Akbarzadeh M,
Fanaei SA, Hedayati M and Azizi F: The role of matrix
metalloproteinase-9 as a prognostic biomarker in papillary thyroid
cancer. BMC Cancer. 18:11992018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Choi C, Thi Thao Tran N, Van Ngu T, Park
SW, Song MS, Kim SH, Bae YU, Ayudthaya PDN, Munir J, Kim E, et al:
Promotion of tumor progression and cancer stemness by MUC15 in
thyroid cancer via the GPCR/ERK and integrin-FAK signaling
pathways. Oncogenesis. 7:852018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim M, Jeon MJ, Oh HS, Park S, Song DE,
Sung TY, Kim TY, Chung KW, Kim WB, Shong YK, et al: Prognostic
implication of N1b classification in the eighth edition of the
tumor-node-metastasis staging system of differentiated thyroid
cancer. Thyroid. 28:496–503. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu X, Quiros RM, Gattuso P, Ain KB and
Prinz RA: High prevalence of BRAF gene mutation in papillary
thyroid carcinomas and thyroid tumor cell lines. Cancer Res.
63:4561–4567. 2003.PubMed/NCBI
|
|
11
|
Kim S, Chung JK, Min HS, Kang JH, Park DJ,
Jeong JM, Lee DS, Park SH, Cho BY, Lee S and Lee MC: Expression
patterns of glucose transporter-1 gene and thyroid specific genes
in human papillary thyroid carcinoma. Nucl Med Mol Imaging.
48:91–97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kitahara CM: New evidence on the
association between prediagnostic thyroid-stimulating hormone
levels and thyroid cancer risk. Cancer Epidemiol Biomarkers Prev.
26:1163–1164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Williams D: Radiation carcinogenesis:
Lessons from Chernobyl. Oncogene. 27 (Suppl 2):S9–S18. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pellegriti G, Frasca F, Regalbuto C,
Squatrito S and Vigneri R: Worldwide increasing incidence of
thyroid cancer: Update on epidemiology and risk factors. J Cancer
Epidemiol. 2013:9652122013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Teng W, Shan Z, Teng X, Guan H, Li Y, Teng
D, Jin Y, Yu X, Fan C, Chong W, et al: Effect of iodine intake on
thyroid diseases in China. N Engl J Med. 354:2783–2793. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim JS: Reply to: Papillary thyroid
microcarcinoma in developing country scenario with endemic iodine
deficiency. Surgery. 162:1912017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zamrazil V, Cerovska J, Bílek R, Simecková
A, Vrbíková J, Dvoráková M, Hníková O, Janecková M and Tomiska F:
The effect of insufficient iodine intake on the size and function
of the thyroid gland. Bratisl Lek Listy. 96:609–612. 1995.(In
Czech). PubMed/NCBI
|
|
18
|
Fiore E, Latrofa F and Vitti P: Iodine,
thyroid autoimmunity and cancer. Eur Thyroid J. 4:26–35. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu XH, Chen GG, Vlantis AC and van
Hasselt CA: Iodine mediated mechanisms and thyroid carcinoma. Crit
Rev Clin Lab Sci. 46:302–318. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dijkstra B, Prichard RS, Lee A, Kelly LM,
Smyth PP, Crotty T, McDermott EW, Hill AD and O'Higgins N: Changing
patterns of thyroid carcinoma. Ir J Med Sci. 176:87–90. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Maier J, van Steeg H, van Oostrom C,
Paschke R, Weiss RE and Krohn K: Iodine deficiency activates
antioxidant genes and causes DNA damage in the thyroid gland of
rats and mice. Biochim Biophys Acta. 1773:990–999. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gerard AC, Humblet K, Wilvers C, Poncin S,
Derradji H, de Ville de Goyet C, Abou-el-Ardat K, Baatout S,
Sonveaux P, Denef JF and Colin IM: Iodine-deficiency-induced long
lasting angiogenic reaction in thyroid cancers occurs via a
vascular endothelial growth factor-hypoxia inducible
factor-1-dependent, but not a reactive oxygen species-dependent,
pathway. Thyroid. 22:699–708. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Aschebrook-Kilfoy B, Grogan RH, Ward MH,
Kaplan E and Devesa SS: Follicular thyroid cancer incidence
patterns in the United States, 1980–2009. Thyroid. 23:1015–1021.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun R, Wang J, Li X, Li L, Yang J, Ren Y,
Xi Y and Sun C: Effect of iodine intake on p14ARF and p16INK4a
expression in thyroid papillary carcinoma in rats. Med Sci Monit.
21:2288–2293. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guan H, Ji M, Bao R, Yu H, Wang Y, Hou P,
Zhang Y, Shan Z, Teng W and Xing M: Association of high iodine
intake with the T1799A BRAF mutation in papillary thyroid cancer. J
Clin Endocrinol Metab. 94:1612–1617. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Saiselet M, Floor S, Tarabichi M, Dom G,
Hébrant A, van Staveren WC and Maenhaut C: Thyroid cancer cell
lines: An overview. Front Endocrinol (Lausanne). 3:1332012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cao X, Ma W, Liu L, Xu J, Wang H, Li X,
Wang J, Zhang J, Wang Z and Gu Y: Analysis of potassium iodate
reduction in tissue homogenates using high performance liquid
chromatography-inductively coupled plasma-mass spectrometry. J
Trace Elem Med Biol. 32:1–6. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang F, Wang Y, Wang L, Wang X, Sun C,
Xing M and Zhao W: Strong association of high urinary iodine with
thyroid nodule and papillary thyroid cancer. Tumour Biol.
35:11375–11379. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang YY, Liu ZB, Ye XG and Ren WM: Iodine
regulates G2/M progression induced by CCL21/CCR7 interaction in
primary cultures of papillary thyroid cancer cells with RET/PTC
expression. Mol Med Rep. 14:3941–3946. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Salemi M, Calogero AE, Vicari E, Migliore
E, Zaccarello G, Cosentino A, Amore M, Tricoli D, Castiglione R,
Bosco P and Rappazzo G: A high percentage of skin melanoma cells
expresses SPANX proteins. Am J Dermatopathol. 31:182–186. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Z, Zhang Y, Liu H, Salati E,
Chiriva-Internati M and Lim SH: Gene expression and immunologic
consequence of SPAN-Xb in myeloma and other hematologic
malignancies. Blood. 101:955–960. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Maine EA, Westcott JM, Prechtl AM, Dang
TT, Whitehurst AW and Pearson GW: The cancer-testis antigens
SPANX-A/C/D and CTAG2 promote breast cancer invasion. Oncotarget.
7:14708–14726. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Salemi M, Calogero AE, Zaccarello G,
Castiglione R, Cosentino A, Campagna C, Vicari E and Rappazzo G:
Expression of SPANX proteins in normal prostatic tissue and in
prostate cancer. Eur J Histochem. 54:e412010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zendman AJ, Zschocke J, van Kraats AA, de
Wit NJ, Kurpisz M, Weidle UH, Ruiter DJ, Weiss EH and van Muijen
GN: The human SPANX multigene family: Genomic organization,
alignment and expression in male germ cells and tumor cell lines.
Gene. 309:125–133. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu K, Ganesan K, Tan LK, Laban M, Wu J,
Zhao XD, Li H, Leung CH, Zhu Y, Wei CL, et al: A precisely
regulated gene expression cassette potently modulates metastasis
and survival in multiple solid cancers. PLoS Genet. 4:e10001292008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Roth RB, Hevezi P, Lee J, Willhite D,
Lechner SM, Foster AC and Zlotnik A: Gene expression analyses
reveal molecular relationships among 20 regions of the human CNS.
Neurogenetics. 7:67–80. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fresno Vara JA, Casado E, de Castro J,
Cejas P, Belda-Iniesta C and Gonzalez-Baron M: PI3K/Akt signalling
pathway and cancer. Cancer Treat Rev. 30:193–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dillon RL, White DE and Muller WJ: The
phosphatidyl inositol 3-kinase signaling network: Implications for
human breast cancer. Oncogene. 26:1338–1345. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wen D, Deng L, Zhou M, Guo S, Shang L, Xu
G and Dong S: A biofuel cell with a single-walled carbon
nanohorn-based bioanode operating at physiological condition.
Biosens Bioelectron. 25:1544–1547. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu R, Liu D, Trink E, Bojdani E, Ning G
and Xing M: The Akt-specific inhibitor MK2206 selectively inhibits
thyroid cancer cells harboring mutations that can activate the
PI3K/Akt pathway. J Clin Endocrinol Metab. 96:E577–E585. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xing M: Genetic alterations in the
phosphatidylinositol-3 kinase/Akt pathway in thyroid cancer.
Thyroid. 20:697–706. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Uddin S, Bavi P, Siraj AK, Ahmed M,
Al-Rasheed M, Hussain AR, Ahmed M, Amin T, Alzahrani A, Al-Dayel F,
et al: Leptin-R and its association with PI3K/AKT signaling pathway
in papillary thyroid carcinoma. Endocr Relat Cancer. 17:191–202.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ma Y, Qin H and Cui Y: MiR-34a targets
GAS1 to promote cell proliferation and inhibit apoptosis in
papillary thyroid carcinoma via PI3K/Akt/Bad pathway. Biochem
Biophys Res Commun. 441:958–963. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xu J, Cai J, Jin X, Yang J, Shen Q, Ding X
and Liang Y: PIG3 plays an oncogenic role in papillary thyroid
cancer by activating the PI3K/AKT/PTEN pathway. Oncol Rep.
34:1424–1430. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Serrano-Nascimento C, da Silva Teixeira S,
Nicola JP, Nachbar RT, Masini-Repiso AM and Nunes MT: The acute
inhibitory effect of iodide excess on sodium/iodide symporter
expression and activity involves the PI3K/AKT signalling pathway.
Endocrinology. 155:1145–1156. 2014. View Article : Google Scholar : PubMed/NCBI
|














