Overview
Cancer is the leading cause of mortality from
non-infectious diseases worldwide, while mortalities associated
with infectious diseases have been declining (reported in 2014)
(1). Cancer is defined as a
multistep process involving the uncontrolled proliferation of
genetically altered cells capable of invading adjacent and distant
tissues, via utilization of the blood and lymphatic systems
(2,3). Certain types of cancer are more
prevalent in developing countries, mainly due to intrinsic factors
such as poverty, late or incorrect diagnosis (1,4),
infectious diseases, and other factors including obesity, physical
inactivity, alcoholism and smoking (5).
In 2015, >8.8 million cancer-associated
mortalities were reported worldwide; 70% of which occurred in low
and middle-income countries and 25% of these were induced by viral
infections, such as hepatitis virus or human papilloma virus (HPV)
(6,7). HPV-16 and HPV-18 are the main
biological factors associated with the development of cervical
cancer (CC), and are the third most common cause of malignant
tumors. According to the 2018 report of GLOBOCAN, CC remains the
fourth most common cause of cancer-associated mortalities among
women worldwide (6,7), and one of the most fatal types of
cancer among the female population in developing countries
(7–9). In Mexico, CC is the second most common
cause of cancer-associated mortality in women (2018), primarily due
to poor clinical diagnosis during the early stages of disease
(7,10). Previous reports between 2015 and 2017
revealed that HPV-16 and HPV-18 are responsible for 65–75% of
precancerous cervical lesions and are present in 99% of patients
with CC worldwide (9,11–13).
However, it has also been established that HPV infections do not
necessarily trigger CC (9,12,13). In
this sense, a significant number of factors are involved;
contraceptives pills, multiple sexual partners, multiple births,
obesity, smoking, alcoholism, poor diet, immunosuppressive cervical
microenvironment, abnormal vaginal microbiota, co-infections with
Chlamydia trachomatis or human immunodeficiency virus and
the presence of cervical cancer stem cells (CCSCs) (9,12,14–19).
Regarding abnormal vaginal microbiota, certain
published reports have indicated remarkable changes in the
microenvironment of patients during the late stages of the disease.
Therefore, patients positive for HPV exhibit more complex bacterial
diversity than a healthy subject. Reports on patients with HPV
frequently exhibit the presence of Lactobacillus iners;
however, Sneathia spp., Megasphaera elsdenii and
Shuttleworthia satelles were commonly found in cervical
intraepithelial neoplasia (CIN) cases (9,19,20–22).
Therefore, some of these microbes could be considered as a
microbiological marker associated with the HPV-infection and CIN.
Besides, Fusobacterium spp. is the most abundant
microorganism found in CC cases (19). Figs. 1
and 2 illustrate the changes in the
cervical microbiota through the progression of CIN I to CC.
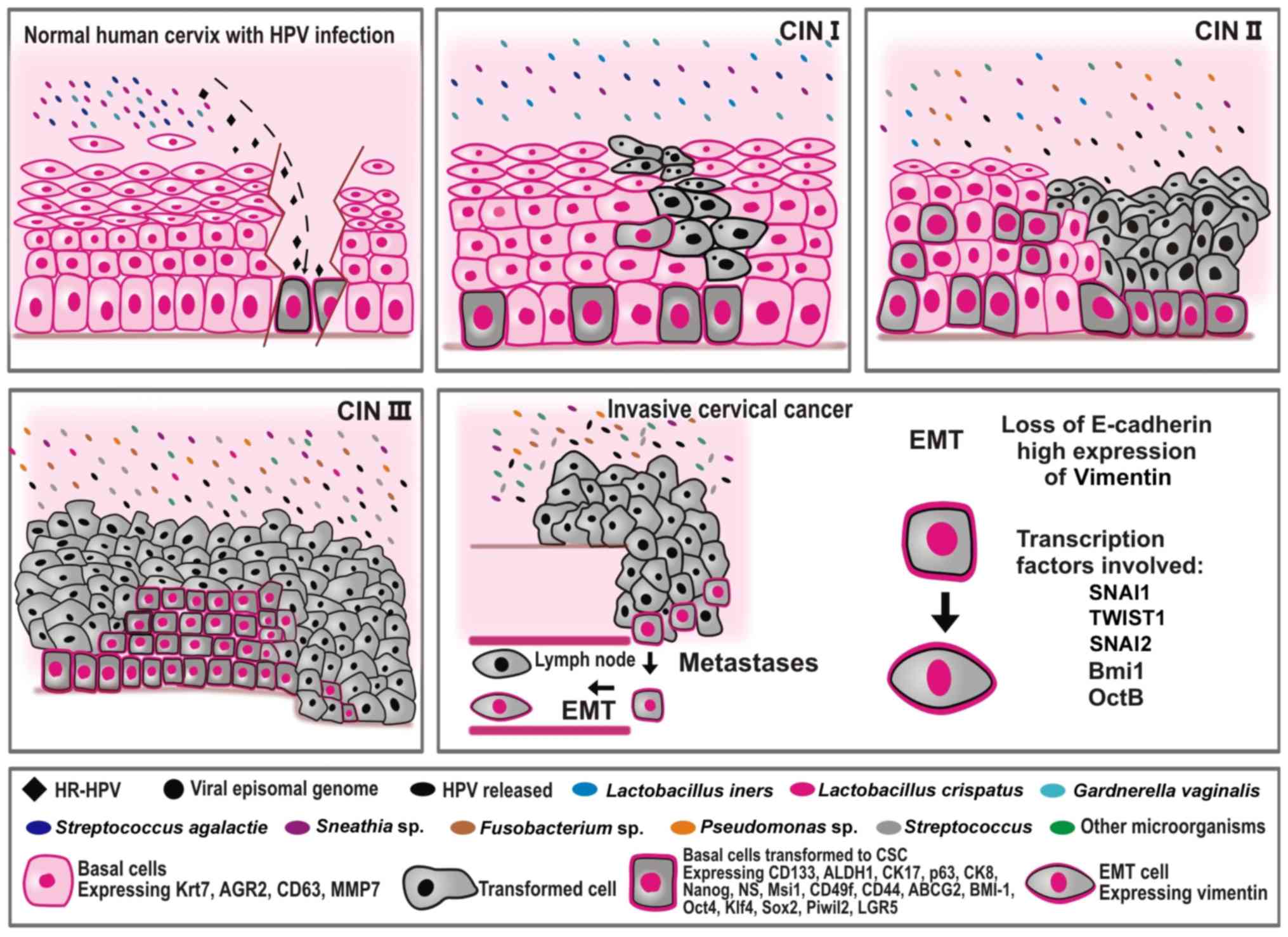 | Figure 1.An overview of factors involved in
the progression of CC. The figure illustrates cervical cancer
progression as a consequence of the multiple factors and steps
involved. CC is initiated by persistent infection with HR HPV (type
16 or type 18) that reaches the basal cells of the transformation
zone, turning it into CCSCs. The microorganisms present during this
stage of the recent infection in a healthy cervix are much less
diverse and much less complex than those reported in more advanced
stages of the disease, such as in CIN II, CIN III, and CC.
Lactobacillus spp. are the principal microorganisms observed
in a healthy cervix and in CIN stage I, and are the first line of
defense against pathogenic microorganisms. As the CIN advances,
more toxic microorganisms are observed in the cervix such as
Fusobacterium, Sneathia and Streptococcus. HPV, human
papilloma virus; CIN, cervical intraepithelial neoplasia; CC,
cervical cancer; CCSCs, cervical cancer stem cells; CSCs, cancer
stem cells; SNAI1, snail family transcriptional repressor 1; SNAI2,
snail family transcriptional repressor 2; TWIST1, twist family bHLH
transcription factor 1; BMI1, BMI1 polycomb ring finger
proto-oncogene; Oct 4, octamer-binding transcription factor 4B;
Krt7, keratin 7; AGR2, anterior gradient protein 2 homolog; Sox2,
sex determining region Y-box 2; Klf4, Krüppel-like factor 4; ALDH1,
aldehyde dehydrogenase 1; CD, cluster of differentiation; CK,
cytokeratin; NS, Nucleostemin; MSI1, musashi RNA binding protein 1;
ABCG2, adenosine triphosphate-binding cassette subfamily G member
2; PIWIL2, piwi like RNA-mediated gene silencing 2; LGR5,
Leucine-rich repeat-containing G-protein-coupled receptor 5; HR,
high-risk; EMT, epithelial-mesenchymal transition; MMP, matrix
metalloproteinase; ABCG2, adenosine triphosphate-binding cassette
subfamily G member 2. |
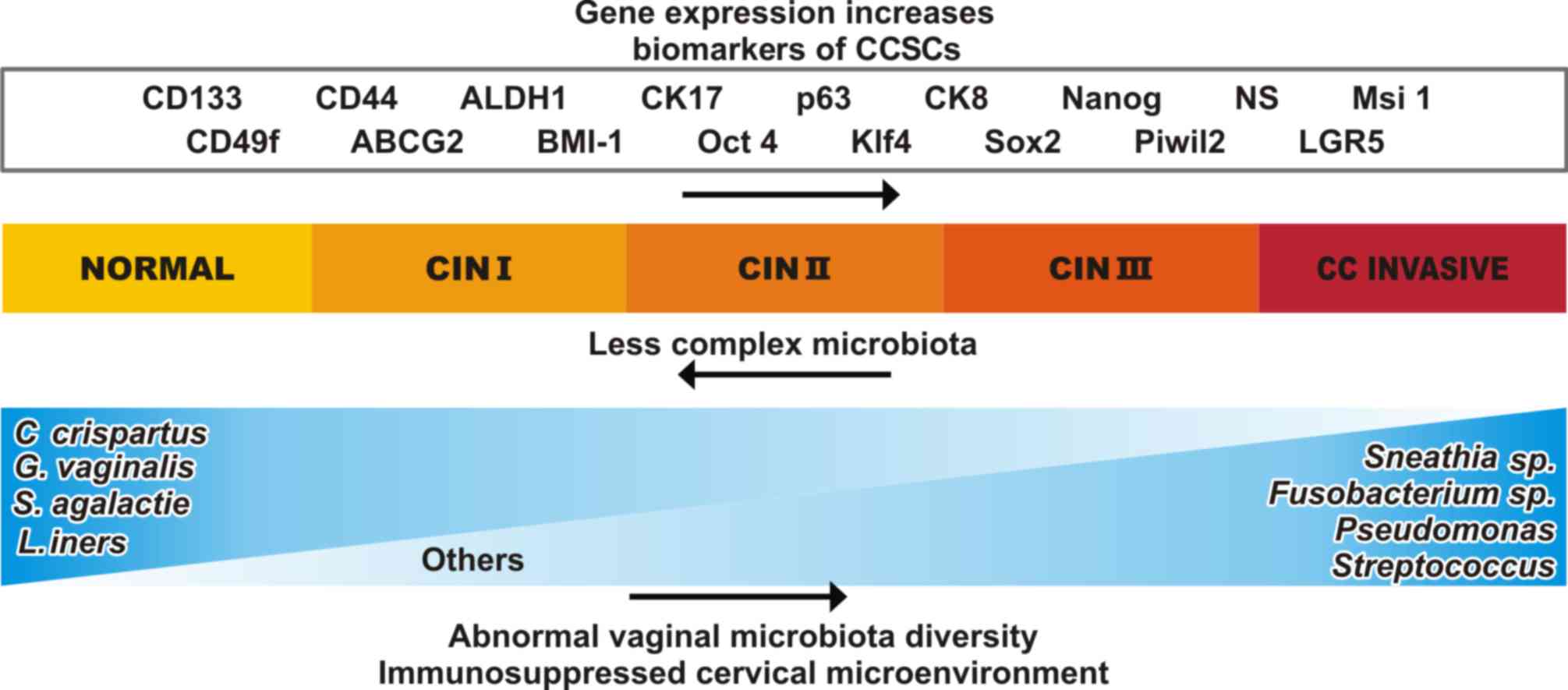 | Figure 2.Major changes occur in the expression
of genes and the cervical microenvironment as CIN I progress to CC.
It has been observed that certain biomarkers of CCSCs exhibit
increased expression and concentration levels, such as NANOG, SOX2
and KLF4, and there is also an increase in markers that are
associated with CCSCs such as, CD133, Cd44, ALDH1, CK17, p63, CK8,
NS, MSI1, CD49f, ABCG2, BMI1, PIWIL2 and LGR5. Many of these
markers have been reported in other types of CSCs. In the cervical
microenvironment, the complexity and quantity of the microbiota are
altered. As the disease progresses, the genus Lactobacillus
tends to disappear and instead, microorganisms as
Fusobacterium sp., Sneathia sp., Pseudomonas
and Streptococcus appear. CIN, cervical intraepithelial
neoplasia; CC, cervical cancer; CCSCs, cervical cancer stem cells;
CD, cluster of differentiation; CK, cytokeratin; NS, Nucleostemin;
MSI1, musashi RNA binding protein 1; PIWIL2, piwi like RNA-mediated
gene silencing 2; LGR5, Leucine-rich repeat-containing
G-protein-coupled receptor 5; BMI1, BMI1 polycomb ring finger
proto-oncogene; ABCG2, adenosine triphosphate-binding cassette
subfamily G member 2; KLF4, Krüppel-like factor 4. |
Although a number of genetic and molecular events
require further clarification in cervical carcinogenesis, it is
currently accepted that HPV viruses depend on epithelial cell
differentiation to establish their progeny and spread their viral
genes (11–13). HPV has a more significant opportunity
to infect basal cells with stem cell-like properties and integrate
its viral DNA into the genome of these cells, driving the oncogenic
transformation into CCSCs (13,23). The
infection occurs where the ectocervix and endocervix join, known as
the transformation zone (TZ) or the squamocolumnar junction
(12,13,24). The
TZ has a unique morphology and genetic expression profile,
expressing keratin 7, anterior gradient 2, CD63 and matrix
metalloproteinase 7, as well as other genes (24). Another essential factor in cervical
carcinogenesis is the epithelial-mesenchymal transition (EMT). EMT
is a crucial process for the generation of invasive cells and
metastasis and it is regulated by snail family transcriptional
repressors 1 (SNAI1) and 2 (SNAI2), and twist family bHLH
transcription factor 1 (TWIST1) transcription factors. The EMT is a
rich source of CSCs and its induction promotes metastasis, tumor
cell invasion and drug resistance. EMT-specific changes include the
loss of E-cadherin expression and upregulation of N-cadherin and
Vimentin (25–28).
On the other hand, the presence of CCSCs is another
factor that is equally important for triggering the development of
CC, as they provide several malignant characteristics to the tumor,
such as the proliferation and metastasis capacity, resistance to
radiation and cisplatin-based chemotherapy. Currently there are no
therapies capable of eliminating CCSCs, and their presence results
in poor prognosis in CC. In this sense, development of dual
therapies against CC could eliminate both primary cancer cells and
CCSCs. Specific biomarkers from CSCs are being considered as
promising targets for the development of new therapies in many
types of cancer. In the present review, the leading factors
involved in cervical cancer development were investigated, with a
particular emphasis on specific biomarkers reported in CCSC.
HPV: A key factor in CC development
HPV consists of double-stranded and circularized DNA
that contains non-coding, long control regions and eight open
reading frames that encode viral proteins located in the ‘early’
and ‘late’ regions (8,29). The early stages of HPV DNA
replication occur in proliferative basal epithelial cells, yielding
low viral copy numbers (8). However,
the progeny of basal cells replicates and move into the spinous
cells layer where viruses mature; allowing the expression of late
capsid proteins and the release of infectious virions within
desquamated cells (8).
Role of viral proteins
Early HPV genes are involved in the synthesis of
proteins associated with replication and maintenance of the viral
genome. In addition, late genes encode proteins associated with the
formation of the viral coat.
E1 is a DNA helicase required for viral replication
in host cells. E1 establishes the viral genome as a multicopy
episome in the nucleus of cells and can arrange protein-protein and
protein-nucleic acid interactions. In this sense, p80, human
SWI/SNF related, matrix associated, actin dependent regulator of
chromatin, subfamily b, member 1, histone H1, E1 binding protein
and p56, are essential in order to link the DNA replication network
of the host cell with the replication origin of the virus. Given
the role that E1 serves in viral DNA replication and its immediate
interaction with the host machinery, there has been speculation
regarding its participation in the development of cervical cancer
(30–33).
E2 is considered the central transcriptional
regulator of the papilloma viruses as it interacts with E1 when DNA
replication begins, and because this protein scatters the viral
episomal genomes during the division of the infected cells. E2
mediates the interactions between the viral genome and chromatin
adapter proteins at the point of mitotic division and indirectly
regulates transcription of E6 and E7 producing several effects on
the cell cycle that affect proliferation, differentiation,
apoptosis and senescence (11,34–36).
E4 is associated with successful virus release and
can also be used as a biomarker of active HPV infections. E4 is
found between the early replication of HPV genes; however, this
protein is also expressed during the late stages of the infection
(37–40).
The E5 protein presents oncogenic activities in
cultured cells and animals, and it is thought to play an important
role in the productive virus cycle. The E5 protein has also been
reported to modulate the activity of cellular proteins, interacting
with targets such as Bax or NF-κB, which induce cell proliferation,
apoptosis and senescence (41–43).
E6 is expressed when transformed cells migrate to
the spinous cell layer. E6 stimulates protein synthesis by
increasing translation through rapamycin (mTOR) complex 1, which
enhances the 5′ mRNA cap translation initiation-complex. It was
reported that loss of E6 resulted in poor maintenance of the HPV
genome due to the failure in p53 degradation. E6 avoids apoptotic
responses by binding to procaspase-8, and it is also able to
inhibit the responses of interferon by downregulating multiple
interferon response genes (44–47).
E7 induces HPV viral replication by reprograming the
cellular environment; together with E6, it induces a potent
transformation activity in the host cells. The influence of E7 is
observed in many cellular processes including viral replication,
transformation, cell cycle and cell death, through direct or
indirect interactions with a large number of proteins (48–52). E6
and E7 induce the degradation of p53 tumor suppressor protein via
the ubiquitin-proteasome pathway, triggering uncontrolled
proliferation of the infected cell population (15,47,48).
L1 protein is comprised of the icosahedral surface
of the HPV virions and it is an initial point of contact between
host cells and the virus. L1 is flexible enough to release the
viral genome into a new target cell; however, assembly of the new
virions occurs only in fully differentiated keratinocytes that are
ready to flake (53–56). Furthermore, L2 also participates in
papillomavirus assembly, initiating the infection process. L2 helps
HPV DNA encapsidation and it is a significant component of the
late-stage proteins. Therefore, L1 and L2 proteins are involved in
virion assembly and in early events of infection; for this reason,
they could be used as potential vaccine antigens (29,57–59).
CSCs
The term ‘cell’, originally proposed in 1665 by the
English scientist Robert Hooke, has been redefined (60). Currently, researchers recognize the
existence of a group of cells with the capacity to renew with high
plasticity. SCs differentiate into the most appropriate lineage,
depending on the stimuli that they receive from the surrounding
microenvironment. In addition to self-renewing, SCs can generate
specialized cells with limited proliferative capacity (61).
The function of this subset of cells is to maintain
tissue homeostasis during daily turnover and regenerate tissue
injuries (62). With the discovery
of humans SCs, even in adults, two new challenges have emerged: i)
Developing laboratory protocols that allow the isolation of SCs in
sufficient amounts; and ii) understanding the molecular mechanisms
that define the fate of these cells (27,63,64). The
identification and isolation of SCs have been performed taking
advantage of molecular biomarkers consisting of differentially
expressed proteins (13). However,
the expression levels of these markers change significantly
depending on the environmental conditions, particularly in in
vitro cultures (64,65). Malignant SCs are called CSCs; these
cells share some characteristics with normal SCs; such as
self-renewal, differentiation, high expression of telomerase,
apoptosis evasion and the ability to migrate (66). CSCs are also able to transport
substances like drugs throughout the membrane, via the membrane
transport protein adenosine triphosphate-binding cassette subfamily
G member 2 (ABCG2), which leads to the recurrence of disease in
patients following surgery and chemotherapy (13,23,24,27,52).
Theories regarding the origin of
CSCs
Given the similarities between SCs and CSCs, several
theories exist regarding the origin of CSCs. DNA mutations can
affect somatic cells and SCs; therefore, essential alterations can
target SCs and produce CSCs with high tumorigenic capacity. Another
possibility is regarding the DNA mutations that target stemness
genes in malignant progenitor cells, transforming progenitor cells
into CSC (13,24). However, the possibility exists that
CSCs could be present in a dormant state until the initiation of
carcinogenesis (23,24,67).
CSC and metastases
As aforementioned, CSCs are involved in promoting
cancer metastasis due to their migratory and invasion capacity to
distant organs. Previous studies have demonstrated that
tumor-associated CSCs are associated with adverse outcomes and high
rates of metastasis (23,25,26,68,69). EMT
is considered to be the primary source of metastatic cells. The
transformation of this cell is induced by transcription factors,
such as SNAI1, SNAI2, TWIST1 and BMI1 polycomb ring finger
proto-oncogene (BMI1), which are highly expressed in CSCs. The EMT
produces a loss of epithelial adhesion and apical-basal polarity,
allowing the release of transformed cells with CSC characteristics
into the circulatory system (25–27).
Fig. 1 illustrates the cells
undergoing EMT, resulting in metastasis.
Biomarkers in CCSCs
Certain specific markers have been reported for
CSCs, including CD44, CD90, CD133, CD271, epithelial cell adhesion
molecule and aldehyde dehydrogenase 1 (ALDH1). However, there is no
set of universal biomarkers to identify and isolate CSCs (23,27,65–74).
Therefore, the main method to study CSCs is through the ‘side
population’ (SP), which is a small subpopulation within the tumor
mass present in some cases (up to 20%) (75). The SP exhibits CSC-like
characteristics, such as the ability to initiate tumors,
development of resistance to chemotherapeutic drugs, and its
potential as a predictor of patient outcome. Therefore the SP could
be an alternative source for studying CSCs with unknown biomarkers
(75,76). Analyses on the basis of the ability
of the cells to efflux the fluorescent dye Hoechst and provide a
system to identify multipotent SCs (76). The spheroid cell formation assay can
also be used as it is based on the capacity of CSCs to grow and
form spheres in non-adherent conditions. The principal focus is to
analyze and compare the expression of surface markers via
fluorescence-activated cell sorting (FACS), confocal microscopy,
immunohistochemistry (IHC), reverse transcription (RT)-quantitative
PCR and the isolation of CSCs for tumorigenic efficiency tests and
tumor subpopulations analysis in animal models (25,27,65,67).
Therefore, CCSC populations exhibit characteristic
expression profiles and cell surface markers that (on tumor tissues
of patients) make their isolation possible in vitro and
in vivo, as well as their evaluation, and research into the
progression of cancer will make creating dual therapies directed
towards specific targets of the CCSC possible in the near future.
Among the main proteins expressed in CCSC are cytokeratin (CK) and
CDs.
CKs −5, −8, −13, −17, −18 and −19 are proteins
expressed in reserve cells and the immature squamous metaplastic
cells of the cervix. CK19 was described by Wang et al
(77), where it was revealed that
the expression of CK19 in CC was significantly higher than that
observed in patients with benign lesions. In addition, high levels
of CK19 expression were identified by RT-PCR in the CCSCs of
sentinel lymph nodes from patients with CC (77). In a study by Ikeda et al
(78), CK8 and CK17 were
investigated by performing IHC on the tissues of patients with
different grades of CIN and CC. Therefore, the authors concluded
that CK8 and CK17 were expressed in CIN and CC tissues, and that
CK17 was associated with metastatic processes and the development
of highly malignant diseases (78).
Thus, CK17 and CK19 could be considered as biomarkers of CCSCs via
the primary cultivation of CC clinical samples (79). In addition, analysis of main and side
population using Hoechst 33342 dye, flow cytometry sorting method,
as well as tumor formation in nude mice revealed that the side
population presented higher tumorigenicity and CSC characteristics
(79).
CD44 and CD133 proteins have been broadly accepted
as general CSC markers in many types of tumor. CD44 and CD133
transmembrane glycoproteins are involved in normal cellular
processes (13) and also in cancer
cell migration, aggregation and tumor development (13,24,72).
Therefore, these glycoproteins can be used as surface markers to
isolate several types of CSCs including breast, prostate, pancreas,
colorectal, gastric and cervical cancer (13,72,80–82). On
the other hand, CD49f is a highly expressed protein in CCSC and it
is advantageous in its identification and isolation.
López et al (74) analyzed the presence of certain
surface markers in sphere cells derived from HeLa, SiHa, CaSki and
C-41 cell lines grown at low density (1,000 cells/ml) in serum-free
medium and discovered an increase in CD49f and CD133 positive cells
when compared with the monolayer cells.
Ortiz-Sánchez et al (69) reported characteristic phenotypes of
putative CCSC including, CD49f+, CK17, p63+, AII+ and ALDH; the
sphere culture exhibited a stemness state characterized by the
presence of OCT4, NANOG and β-catenin. In addition, it was observed
that the presence of CD49f and AII was associated with the
possibility that HR-HPV can infect normal cervical cells (SCs).
Ortiz-Sánchez et al (69)
also demonstrated the high tumorigenic capacity of
ALDHbright cells when compared with ALDHlow
cells.
Tyagi et al (83) demonstrated the role of viral
oncoprotein E6 in SC signaling and the maintenance of stemness in
CC. CSCs that expressed a set of phenotypic markers including
CD49f, ABCG2, CD71 and CD133 were isolated from primary cervical
tumors and cancer-derived cell lines grown as spheres. In addition,
transcripts of self-renewal and stemness markers including OCT4,
SOX2, NANOG, leucine-rich repeat-containing G protein-coupled
receptor 1 (LGR1) and CD133 were identified, along with the
overexpression of E6, Hes family bHLH transcription factor 1, a
protein involved in enhancing self-renewal properties and the
ability of tumorsphere formation (83).
Hou et al (84) reported that there was a correlation
between CCSC markers in patient tissue samples and the prediction
of CIN prognosis. The authors analyzed paraffin-embedded surgical
samples through IHC, snap-frozen CC samples and normal cervical
samples using RT-PCR for CD49f, SOX2, ALDH1 and musashi RNA binding
protein 1 (MSI1). In this sense, patients with tumors classified as
high MSI1 and low CD49f expression had the poorest prognosis,
whereas tumors without MSI1 and CD49f upregulation had the best
prognosis in CC. Hou et al (84) reported, for the first time, clinical
evidence regarding CCSC markers associated with the prognosis of
patients with CC.
Other proteins differentially expressed in CCSC are
NANOG, Nucleostemin (NS), MSI1, TWIST, nestin, ALDH1, BMI1,
piwi-like RNA-mediated gene silencing 2 (PIWIL2), TIMP
metallopeptidase inhibitor 4 (TIMP4), LGR5, OCT and SOX.
Ye et al (85)
reported the role of NANOG, NS and MSI1 in cervix carcinogenesis
and progression of cervical carcinoma by performing IHC analysis on
235 paraffin-embedded samples with normal cervical epithelia,
CIN-I, -II and -III; and CC. The CINs were staged as follows
(86): I, low-grade lesion with
mildly atypical cellular changes in the lower third of the
epithelium (mild dysplasia); CIN II, high-grade lesion with
moderately atypical cellular changes confined to the basal
two-thirds of the epithelium (moderate dysplasia); and CIN III,
high-grade lesion with severely atypical cellular changes
encompassing the full thickness of the epithelium (severe
dysplasia). Ye et al (85)
observed high levels of expression of the three proteins in CC, CIN
-II and -III, and low expression levels in CIN I and normal
cervical epithelia. However, there was no correlation among NANOG,
NS and MSI1 expression levels and the prognosis of CC. In addition,
NANOG, NS and MSI1 also have a critical role in the carcinogenesis
of glioma, liver, gastric and other types of cancer (85).
In 2011, Li and Zhou (87) revealed the activation of
Wnt/β-catenin and Akt signaling pathways in TWIST-overexpressing
cells that had CSC characteristics, such as tumorsphere formation
and ALDH1 and CD44 expression. The study reported that TWIST is an
inducer of morphological changes associated with EMT. In addition,
the spheroid cells gained expression of human actin α-cardiac
muscle 1 (also known as α-smooth muscle actin) and Vimentin
mesenchymal markers. Functional analysis demonstrated that spheroid
cells are more resistant than monolayer cells to paclitaxel
(87). On the other hand, knockdown
of β-catenin expression by small interfering RNA transfection and
Akt signaling pathway inhibition by the PI3K/Akt inhibitor
wortmannin, suppressed the expression of CD44 (87).
Sato et al (88), determined the role of Nestin in CIN
and CC via IHC and in situ hybridization analysis. Nestin is
usually expressed in the brain; however, it was also identified in
CSCs, several metastasized carcinomas and in types of cancer with
poor prognosis. In this study, Sato et al (88) analyzed tissue samples from 26 cases
of each stage of CIN and 55 of CC. The results revealed low Nestin
expression levels in the basal squamous epithelium of CIN I;
however, in CIN II Nestin was present in 65% of the cases. Nestin
was found in most of the cases of CIN III localized in the squamous
epithelium.
Furthermore, Nestin was detected in all of the
invasive CC samples. In addition, Sato et al (88) analyzed the effects of Nestin
overexpression via transfection of ME-180 cells; a metastatic
cancer cell line derived from the cervix. Overexpression of Nestin
induced a higher capacity to form spheres, produce a
CD44high/CD24low pattern associated with the biomarkers found in
breast CSCs, increased expression of ALDH, NANOG and OCT4.
According to these results, the authors concluded that Nestin may
be involved in the progression of CIN to CC and could be associated
with the regulation of CSCs due to its ability to stimulate sphere
formation in vitro (88).
ALDH1 is a cytosolic isoenzyme normally involved in
retinol oxidation to produce retinoic acid. Liu and Zheng (89) reported high levels of ALDH1 in CCSCs.
The authors observed that patients exhibited high ALDH1 activity
level in a subpopulation of CC cases with a great capacity for
self-renewal, high differentiation potential, and high
tumorigenicity, just like CSCs. The authors used tools such as FACS
and functional assays in xenografted NOD/SCID mice, as well as
cultures of CC cells expressing high and low levels of ALDH1 in
serum-free media to promote tumor sphere cell formation. The
authors concluded that ALDH1 may function as a CSCC marker. In the
study by Liu and Zheng (89), cells
with high ALDH1 activity were resistant to cisplatin and produced a
high expression of OCT4, NANOG, Krüppel-like factor 4 (KLF4) and
BMI1.
Liu et al (90) demonstrated that SOX2-positive CC
cells share all the characteristics with CSCs including
self-renewal, differentiation and tumor-initiating properties.
Additionally, SOX2-positive cervical cells increased the levels of
OCT4, BMI1 and ALDH1 stemness markers, as well as Vimentin, SNAI1
and β-catenin (mesenchymal SCs markers) (91). For this reason, the authors concluded
that SOX2 could be a critical factor in self-renewal, pluripotency
and also a stemness factor needed for SCs and CCSCs differentiation
(90).
Lizarraga et al (92) described the role of TIMP4 in the
stemness of CC cells. TIMP4 is a tissue inhibitor that has been
overexpressed in a number of cancer cell lines and in nude mice, to
evaluate its function in carcinogenesis. The results by Lizarraga
et al (92) revealed a faster
tumor formation in nude mice that overexpressed TIMP4 in CC cells.
Notably, activation of NF-κB signaling pathway and the increasing
CSC population led to a high expression of pluripotency markers as
OCT3/4 and SOX2, the EMT markers SNAI1 and Vimentin and the drug
efflux transporters markers ABCG1 and ABCG2 (92).
Feng et al (93) demonstrated the role of PIWIL2 in CC
tumorigenesis. They observed that PIWIL2 expression was present in
the HPV+ CC cell lines HeLa, SiHa and CaSki, and was
undetectable in HPV− cancer cell line C33A. Knockdown of
PIWIL2 by short hairpin RNA in HeLa and SiHa cells decreased the
tumorigenic, proliferation and chemoresistant capacity of these
cells. On the other hand, overexpression of PIWIL2 in HaCat cells
activated tumor-initiating capabilities and cMyc, KLF4, NANOG, OCT4
and SOX2 cell reprogramming factors were upregulated (93). Feng et al (93) also demonstrated that PIWIL2
reactivation by E6 and E7 oncoproteins is essential in the
transformation of cervical epithelial cells into CSCs. PIWIL2 was
highly expressed in CIN II, CIN III and CC, but its expression in
healthy tissue and CIN I was low. PIWIL2 also suppressed the
expression of P53 and P21 in CC cell lines, inducing cervical
carcinogenesis (93).
Cao et al (94) previously reported the principal role
of LGR5 in CC for the activation of Wnt/β-catenin signaling
pathway. In addition, this study reported the role of LGR5 in CCSCs
via overexpressing and silencing its effect in CC cell lines. Thus,
they showed that overexpression of LGR5 induces CSC characteristics
including tumorsphere formation, increased tumorigenic capacities
in vivo, chemoresistance to cisplatin, augmented cell
migration and invasion, and upregulate the expression of stem
cell-associated transcription factors in vitro.
LGR5-overexpression in HeLa and SiHa cells was revealed as being
correlated with higher expression levels of BMI1, NANOG, OCT4 and
KLF4, when compared with control or silenced cells (94). Figs. 1
and 2 summarize leading factors
involved in the progression of CC, the significant changes in
genetic expression from CCSC as well as the changes in microbiota
population, throughout CC development and progression.
CSC signaling pathway
SCs and CSCs share Hedgehog (Hh), Notch, Wnt, NF-κB
and PI3K/Akt/mTOR signaling pathways. Hh is an essential pathway
for self-renewal and cell fate; it is associated with
tumorigenesis, development and the progression of certain types of
cancer including the maintenance of CSCs (95). The Hh signaling pathway drives
stemness in CSCs via the genetic regulation of OCT4, SOX2 and BMI1
(96,97). Regarding CC, Hh has been associated
with poor outcomes in irradiated patients and evidence has
suggested that Hh is involved in the repopulation of cervical cells
following chemoradiation (98).
The Notch signaling pathway regulates proliferation,
stem cell maintenance, cell fate specification, differentiation and
angiogenesis (99). In addition, the
Notch signaling pathway is involved in cell-cell communication
through transmembrane ligands and receptors (100). The canonical pathway involves five
canonical Notch ligands: ∆-like canonical Notch ligand (DLL) 1,
DLL3, DLL4, Jagged1 and Jagged2, as well as four receptor paralogs.
Cancer cells express different Notch receptors and ligands
(100) and the noncanonical pathway
may also have some relevance in cancer. Thus, both pathways can
control several types of tumor-associated cells, including CSCs or
immune cells. Furthermore, expression of different Notch paralogs
varies between different types of tumor (101). In CC, Notch can exhibit different
roles depending on the disease progression: i) Maintaining immature
epithelium by preventing terminal differentiation; ii) increasing
NOTCH expression during the progress of CIN into CC; iii)
regulation of Notch signaling pathway by E6 in CC cell lines; and
iv) silencing Jagged1 in CaSki cells inhibits its tumorigenic
capacity (101,102).
The Wnt signaling pathway is involved in cell
proliferation and differentiation during embryogenesis. From Wnt
signaling cascades, the canonical path is the best studied for its
involvement in cancer development. There are many reports regarding
CSC contribution to the maintenance of these cells through the
Wnt-β-catenin signaling pathway (103–106).
In CC, apoptosis is induced and tumor growth is inhibited if the
Wnt signaling pathway is repressed. On the other hand,
overactivation of Wnt/β-catenin signaling pathway is associated
with cervical tumorigenesis with HPV infection (105,107).
In the basal layer, Wnt ligands are required for sustaining the
undifferentiated state of SCs. The majority of genetic mutations in
colorectal cancer activate the Wnt signaling pathway, and CSCs are
the most susceptible to transformation by these mutations (28,108).
NF-κB serves a pivotal role in HPV infected cells.
This signaling pathway is involved in cancer development by
regulating several oncogenic genes (109). The NF-κB signaling pathway has two
routes: The canonical pathway that depends on the inhibitor of
NF-κB kinase complex (IκB), and the non-canonical pathway, which is
activated when the homodimer of IkB, inhibitor of nuclear factor
kappa-B kinase subunit α (IKKα) is phosphorylated. IKKα is part of
the IκB complex, and is associated with the growth, metastases and
stemness of several types of cancer (110,111).
In the PI3K/Akt/mTOR signaling pathway, PI3K and
mTOR serve a critical role in cell proliferation, angiogenesis,
metabolism, differentiation and survival (112). This signaling pathway is usually
activated when mTOR is not correctly regulated in cancer
conditions. PI3K overexpression has been reported in ovarian cancer
and CC (113,114). PI3K/Akt/mTOR is an essential
signaling pathway to regulate self-renewal and the maintenance of
stemness in SCs and CSCs. The role of CSCs is well recognized in
prostate cancer (66,115); however, the mechanism through
PI3K/Akt/mTOR signaling regulates CSC is unknown.
Conclusions
A large number of studies have sought to expand the
current knowledge of the molecular pathogenesis of CC and the
progression of viral infections leading to this invasive cancer.
The majority of these studies have attempted to verify that the
cause of CC is the HPV infection; however, recent studies have
aimed to determine the factors and changes at the molecular level
that are involved with the stemness and development of CC. In this
sense, a growing amount of evidence has identified the CCSCs as a
novel, fundamental and strategic key factor to be considered in
cancer development, chemotherapy resistance and cancer regression.
Regarding the origin of CCSCs, the best-accepted hypothesis
involves the transformation of SCs, via HPV infection by using
their E6 and E7 oncoproteins. However, there are a large number of
proteins whose natural activity is the maintenance of stemness in
healthy cells. Nevertheless, SOX2, NANOG, OCT4, Klf4 and Nestin
proteins are also involved in maintaining the stemness of CSCs.
ABCG2, SNAI1, Vimentin and LGR5 are other proteins
that are also involved in maintenance, although it is not their
primary role. This evidence confirms the existence of a complex
network of regulation in CC development and progression. Thus,
searching biomarkers at each disease stage is essential to improve
our current knowledge for the development and application of novel
therapies and to allow for an accurate CC diagnoses.
Currently, several research groups are making a
great effort to identify new targets involved in the stemness of
cervical cancer cells. Therefore, genes, proteins and signaling
pathways are under consideration. In this sense, the information
described in the present review collates some potential candidates
to accomplish the difficult task of controlling CCSC development.
Thus, CD44, CD133 and CD49f could be an excellent target to direct
the therapeutic efforts and block Hedgehog, PI3K/Akt/mTOR, Wnt or
Notch signaling pathways in CCSC.
Acknowledgements
The authors would like to thank Dr Michelle Quezada
from The University of Newcastle (Newcastle, UK) and Mr Yael Vargas
for their assistance in editing the manuscript, and also Mr
Humberto Vallejo for his support with the figure design.
Funding
No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
GMA, EOS, LRZ, CRS, EEI and JO were all responsible
for reviewing the cited literature and writing the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Barondess JA: Scanning the chronic disease
terrain: Prospects and opportunities. Trans Am Clin Climatol Assoc.
125:45–56. 2014.PubMed/NCBI
|
|
2
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sanchez-Vega F, Mina M, Armenia J, Chatila
WK, Luna A, La KC, Dimitriadoy S, Liu DL, Kantheti HS, Saghafinia
S, et al: Oncogenic signaling pathways in the cancer genome atlas.
Cell. 173:321–337.e10. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sahasrabuddhe V, Luhn P and Wentzensen N:
Human papillomavirus and cervical cancer: Biomarkers for improved
prevention efforts. Future Microbiol. 6:1083–1098. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Latino-Martel P, Cottet V, Druesne-Pecollo
N, Pierre FH, Touillaud M, Touvier M, Vasson MP, Deschasaux M, Le
Merdy J, Barrandon E and Ancellin R: Alcoholic beverages, obesity,
physical activity and other nutritional factors, and cancer risk: A
review of the evidence. Crit Rev Oncol Hematol. 99:308–323. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
International agency for Research on
Cancer. Global Cancer Observatory. http://gco.iarc.fr
|
|
8
|
Scheurer ME, Tortolero-Luna G and
Adler-Storthz K: Human papillomavirus infection: Biology,
epidemiology, and prevention. Int J Gynecol Cancer. 15:727–746.
2015. View Article : Google Scholar
|
|
9
|
Kyrgiou M, Mitra A and Moscicki AB: Does
the vaginal microbiota play a role in the development of cervical
cancer? Transl Res. 179:168–182. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Secretariat of Health, Mexico, .
Statistics of breast cancer and uterine cervical cancer. https://www.gob.mx/salud/acciones-y-programas/informacion-estadistica2015
|
|
11
|
Domínguez-Catzín V, Reveles-Espinoza AM,
Sánchez-Ramos J, Cruz-Cadena R, Lemus-Hernández D and Garrido E:
HPV16-E2 protein modifies self-renewal and differentiation rate in
progenitor cells of human immortalized keratinocytes. Virol J.
14:652017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chhabra R: Cervical cancer stem cells:
Opportunities and challenges. J Cancer Res Clin Oncol.
141:1889–1897. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yao T, Lu R, Zhang Y, Zhang Y, Zhao C, Lin
R and Lin Z: Cervical cancer stem cells. Cell Prol. 48:611–625.
2015. View Article : Google Scholar
|
|
14
|
Ncube B, Bey A, Knight J, Bessler P and
Jolly PE: Factors associated with the uptake of cervical cancer
screening among women in portland, Jamaica. N Am J Med Sci.
7:104–113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jungbauer F, Aderhold C, Birk R, Hoermann
K, Kramer B, Kuhlin B, Thorn C, Umbreit C and Lammert A:
Communicate or Die-A Model for HPV+ and HPV- CSCs and their
interactions with SDF-1α. Anticancer Res. 37:4827–4836.
2017.PubMed/NCBI
|
|
16
|
American Cancer Society: What are the risk
factors for cervical cancer? http://www.cancer.org/cancer/cervicalcancer/moreinformation/cervicalcancerpreventionandearlydetection2014
|
|
17
|
Alfaro KM, Gage JC, Rosenbaum AJ, Ditzian
LR, Maza M, Scarinci IC, Miranda E, Villalta S, Felix JC, Castle PE
and Cremer ML: Factors affecting attendance to cervical cancer
screening among women in the Paracentral Region of El Salvador: A
nested study within the CAPE HPV screening program. BMC Public
Health. 15:10582015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ngugi CW, Boga H, Muigai AW, Wanzala P and
Mbithi JN: Factors affecting uptake of cervical cancer early
detection measures among women in Thika, Kenya. Health Care Women
Int. 33:595–613. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Audirac-Chalifour A, Torres-Poveda K,
Bahena-Román M, Téllez-Sosa J, Martínez-Barnetche J,
Cortina-Ceballos B, López-Estrada G, Delgado-Romero K,
Burguete-García AI, Cantú D, et al: Cervical microbiome and
cytokine profile at various stages of cervical cancer: A pilot
study. PLoS One. 11:e01532742016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mitra A, Maclntyre DA, Lee YS, Smith A,
Marchesi JR, Lehne B, Bhatia R, Lyons D, Paraskevaidis E, Li JV, et
al: Cervical intraepithelial neoplasia disease progression is
associated with increased vaginal microbiome diversity. Sci Rep.
5:168652015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mitra A, Maclntyre DA, Marchesi JR, Lee
YS, Benett PR and Kyrgiou M: The vaginal microbiota, human
papillomavirus infection and cervical intraepithelial neoplasia:
What do we know and where are we going next? Microbiome. 4:582016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang X, Da M, Zhang W, Qi Q, Zhang C and
Han S: Role of Lactobacillus in cervical cancer. Cancer Manag Res.
10:1219–1229. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang R and Rofstad E: Cancer stem cells
(CSCs), cervical CSCs and targeted therapies. Oncotarget.
8:35351–35367. 2017.PubMed/NCBI
|
|
24
|
Rao QX, Yao TT, Zhang BZ, Lin RC, Chen ZL,
Zhou H, Wang LJ, Lu HW, Chen Q, Di N and Lin Z: Expression and
functional role of ALDH1 in cervical carcinoma cells. Asian Pac J
Cancer Prev. 13:1325–1331. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
López J, Ruíz G, Organista-Nava J,
Gariglio P and García-Carrancá A: Human papillomavirus infections
and cancer stem cells of tumors from the uterine cervix. Open Virol
J. 6:232–240. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin J, Liu X and Ding D: Evidence for
epithelial-mesenchymal transition in cancer stem-like cells derived
from carcinoma cell lines of the cervix uteri. Int J Clin Exp
Pathol. 8:847–855. 2015.PubMed/NCBI
|
|
27
|
Yang MH, Imrali A and Heeschen C:
Circulating cancer stem cells: The importance to select. Chin J
Cancer Res. 27:437–449. 2015.PubMed/NCBI
|
|
28
|
Batlle E and Clevers H: Cancer stem cells
revisited. Nat Med. 23:1124–1134. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang JW and Roden RB: L2, the minor capsid
protein of papillomavirus. Virology. 445:175–186. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kuo SR, Liu JS, Broker TR and Chow LT:
Cell-free replication of the human papillomavirus DNA with
homologous viral E1 and E2 proteins and human cell extracts. J Biol
Chem. 269:24058–24065. 1994.PubMed/NCBI
|
|
31
|
Sanders CM, Kovalevskiy OV, Sizov D,
Lebedev AA, Isupov MN and Anston AA: Papillomavirus E1 helicase
assembly maintains an asymmetric state in the absence of DNA and
nucleotide cofactors. Nucleic Acids Res. 35:6451–6457. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Egawa N, Nakahara T, Ohno S,
Narisawa-Saito M, Yugawa T, Fujita M, Yamato K, Natori Y and Kiyono
T: The E1 protein of human papillomavirus type 16 is dispensable
for maintenance replication of the viral genome. J Virol.
86:3276–3283. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bergvall M, Melendy T and Archambault J:
The E1 proteins. Virology. 445:35–56. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chin MT, Hirochika R, Hirochika H, Broker
TR and Chow LT: Regulation of human papillomavirus type 11 enhancer
and E6 promoter by activating and repressing proteins from the E2
open reading frame: Functional and biochemical studies. J Virol.
62:2994–3002. 1988.PubMed/NCBI
|
|
35
|
Hou SY, Wu SY, Zhou T, Thomas MC and
Chiang CM: Alleviation of human papillomavirus E2-mediated
transcriptional repression via formation of a TATA binding protein
(or TFIID)-TFIIB-RNA polymerase II-TFIIF preinitiation complex. Mol
Cell Biol. 20:113–125. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
McBride AA: The Papillomavirus E2
proteins. Virology. 445:57–79. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Davy C and Doorbar J: G2/M cell cycle
arrest in the life cycle of viruses. Virology. 368:219–226. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Borgogna C, Zavattaro E, de Andrea M,
Griffin HM, Dell'Oste V, Azzimonti B, Landini MM, Peh WL, Pfister
H, Doorbar J, et al: Characterization of beta papillomavirus E4
expression in tumours from Epidermodysplasia Verruciformis patients
and in experimental models. Virology. 423:195–204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Griffin H, Wu Z, Marnane R, Dewar V,
Molijin A, Quint W, Van Hoof C, Struyf F, Colau B, Jenkins D and
Doorbar J: E4 antibodies facilitate detection and type-assignment
of active HPV infection in cervical disease. PLoS One.
7:e499742012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Doorbar J: The E4 protein; structure,
function and patterns of expression. Virology. 445:80–98. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang Y, Lehman JM and Petti LM: Apoptosis
of mortal human fibroblasts transformed by the bovine
papillomavirus E5 oncoprotein. Mol Cancer Res. 1:122–136.
2002.PubMed/NCBI
|
|
42
|
Venuti A, Paolini F, Nasir L, Corteggio A,
Roperto S, Campo MS and Borzacchiello G: Papillomavirus E5: The
smallest oncoprotein with many functions. Mol Cancer. 10:1402011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Di Maio D and Petti LM: The E5 proteins.
Virology. 445:99–114. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Butz K, Ristriani T, Hengstermann A, Denk
C, Scheffner M and Hoppe-Seyler F: siRNA targeting of the viral E6
oncogene efficiently kills human papillomavirus-positive cancer
cells. Oncogene. 22:5938–5945. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ansari T, Brimer N and Vande Pol SB:
Peptide interactions stabilize and restructure human papillomavirus
type 16 E6 to interact with p53. J Virol. 86:11386–11391. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zanier K, ould M'hamed ould Sidi A,
Boulade-Ladame C, Rybin V, Chapelle A, Atkinson A, Kieffer B and
Travé G: Solution structure analysis of the HPV16 E6 oncoprotein
reveals a self-association mechanism required for E6-mediated
degradation of p53. Structure. 20:604–617. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Vande Pol SB and Klingelhutz AJ:
Papillomavirus E6 oncoproteins. Virology. 445:115–137. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
McLaughlin-Drubin ME, Bromberg-White JL
and Meyers C: The role of the human papillomavirus type 18 E7
oncoprotein during the complete viral life cycle. Virology.
338:61–68. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
McLaughlin-Drubin ME, Huh KW and Münger K:
Human papillomavirus type 16 E7 oncoprotein associates with E2F6. J
Virol. 82:8695–8705. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
McLaughlin-Drubin ME and Münger K: The
human papillomavirus E7 oncoprotein. Virology. 384:335–344. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
McLaughlin-Drubin ME, Crum CP and Münger
K: Human papillomavirus E7 oncoprotein induces KDM6A and KDM6B
histone demethylase expression and causes epigenetic reprogramming.
Proc Natl Acad Sci USA. 108:2130–2135. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
McLaughlin-Drubin ME, Meyers J and Munger
K: Cancer associated human papillomaviruses. Curr Opin Virol.
2:459–466. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Schäfer F, Florin L and Sapp M: DNA
binding of L1 is required for human papillomavirus morphogenesis in
vivo. Virology. 295:172–181. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Doorbar J: The papillomavirus life cycle.
J Clin Virol. 32 (Suppl 1):S7–S15. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Day PM, Lowy DR and Schiller JT: Heparan
sulfate-independent cell binding and infection with
furin-precleaved papillomavirus capsids. J Virol. 82:12565–12568.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Buck CB, Day PM and Trus BL: The
papillomavirus major capsid protein L1. Virology. 445:169–174.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kirnbauer R, Chandrachud LM, O'Neil BW,
Wagner ER, Grindlay GJ, Armstrong A, McGarvie GM, Schiller JT, Lowy
DR and Campo MS: Virus-like particles of bovine papillomavirus type
4 in prophylactic and therapeutic immunization. Virology.
219:37–44. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Rubio I, Seitz H, Canali E, Sehr P, Bolchi
A, Tommasino M, Ottonello S and Müller M: The N-terminal region of
the human papillomavirus L2 protein contains overlapping binding
sites for neutralizing, cross-neutralizing and non-neutralizing
antibodies. Virology. 409:348–359. 2001. View Article : Google Scholar
|
|
59
|
Doorbar J, Quint W, Banks L, Bravo IG,
Stoler M, Broker TR and Stanley MA: The biology and life-cycle of
human papillomaviruses. Vaccine. 30 (Suppl 5):F55–F70. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kutschera U: Founding fathers: The cell
was defined 150 years ago. Nature. 480:4572011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Nava MM, Raimondi MT and Pietrabissa R:
Controlling self-renewal and differentiation of stem cells via
mechanical cues. J Biomed Biotechnol 2012. 7974102012.
|
|
62
|
Ge Y and Fuchs E: Stretching the limits:
from homeostasis to stem cell plasticity in wound healing and
cancer. Nat Rev Genet. 19:311–325. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Maruyama T: Stem/progenitor cells and the
regeneration potentials in the human uterus. Reprod Med Biol.
9:9–16. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zapata AG, Alfaro D and García-Ceca J:
Biology of stem cells: The role of microenvironments. Adv Exp Med
Biol. 741:135–151. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yang B, Lu Y, Zhang A, Zhou A, Zhang L,
Zhang L, Gao L, Zang Y, Tang X and Sun L: Doxycycline induces
apoptosis and inhibits proliferation and invasion of human cervical
carcinoma stem cells. PLoS One. 10:e01291382015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Aponte PM and Caicedo A: Stemness in
cancer: Stem cells, cancer stem cells, and their microenvironment.
Stem Cells Int 2017. 56194722017.
|
|
67
|
Clevers H: The cancer stem cell: Premises,
promises and challenges. Nat Med. 17:313–319. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kobayashi NC and Noronha SM: Cancer stem
cells: A new approach to tumor development. Rev Assoc Med Bras
(1992). 61:86–93. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Ortiz-Sánchez E, Santiago-López L,
Cruz-Domínguez VB, Toledo-Guzmán ME, Hernández-Cueto D,
Muñiz-Hernández S, Garrido E, Cantú De León D and García-Carra A:
Characterization of cervical cancer stem celllike cells:
Phenotyping, stemness, and human papillomavirus co-receptor
expression. Oncotarget. 7:31943–31954. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Chiou SH, Yu CC, Huang CY, Lin SC, Liu CJ,
Tsai TH, Chou SH, Chien CS, Ku HH and Lo JF: Positive correlations
of Oct-4 and Nanog in oral cancer stem-like cells and high-grade
oral squamous cell carcinoma. Clin Cancer Res. 14:4085–4095. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Chen YC, Chen YW, Hsu HS, Tseng LM, Huang
PI, Lu KH, Chen DT, Tai LK, Yung MC, Chang SC, et al: Aldehyde
dehydrogenase 1 is a putative marker for cancer stem cells in head
and neck squamous cancer. Biochem Biophys Res Commun. 385:307–313.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhang Q, Shi S, Yen Y, Brown J, Ta JQ and
Le AD: A subpopulation of CD133(+) cancer stem-like cells
characterized in human oral squamous cell carcinoma confer
resistance to chemotherapy. Cancer Lett. 289:151–160. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Murillo-Sauca O, Chung MK, Shin JH,
Karamboulas C, Kwok S, Jung Y, Oakley R, Tysome JR, Farnebo LO,
Kaplan MJ, et al: CD271 is a functional and targetable marker of
tumor-initiating cells in head and neck squamous cell carcinoma.
Oncotarget. 5:6854–6866. 2014.PubMed/NCBI
|
|
74
|
López J, Poitevin A, Mendoza-Martínez V,
Pérez-Plasencia C and García-Carrancá A: Cancer-initiating cells
derived from established cervical cell lines exhibit stem-cell
markers and increased radioresistance. BMC Cancer. 12:482012.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Wu C and Alman BA: Side population cells
in human cancers. Cancer Lett. 268:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Richard V, Nair MG, Santhosh Kumar TR and
Pillai MR: Side population cells as prototype of chemoresistant,
tumor-initiating cells. Biomed Res Int. 2013:517237. 2013.
View Article : Google Scholar
|
|
77
|
Wang HY, Sun JM, Lu HF, Shi DR, Ou ZL, Ren
YL and Fu SQ: Micrometastases detected by cytokeratin 19 expression
in sentinel lymph nodes of patients with early-stage cervical
cancer. Int J Gynecol Cancer. 16:643–648. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Ikeda K, Tate G, Suzuki T and Mitsuya T:
Coordinate expression of cytokeratin 8 and cytokeratin 17
immunohistochemical staining in cervical intraepithelial neoplasia
and cervical squamous cell carcinoma: An immunohistochemical
analysis and review of the literature. Gynecol Oncol. 108:598–602.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wang Y, Wang M, Zeng Q, Lv Y and Bao B:
Isolation and biological characteristics of human cervical cancer
side population cells. Int J Clin Exp Pathol. 10:869–876. 2017.
|
|
80
|
Takaishi S, Okumura T, Tu S, Wang S,
Shibata W, Vingneshwaran R, Gordon SA, Shimada Y and Wang TC:
Identification of gastric cancer stem cells using the cell surface
marker CD44. Stem Cells. 27:1006–1020. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Su YJ, Lai HM, Chang YW, Chen GY and Lee
JL: Direct reprogramming of stem cell properties in colon cancer
cells by CD44. EMBO J. 30:3186–3199. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Hiraga T, Ito S and Nakamura H: Cancer
stem-like cell marker CD44 promotes bone metastases by enhancing
tumorigenicity, cell motility, and hyaluronan production. Cancer
Res. 73:4112–4122. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Tyagi A, Vishnoi K, Mahata S, Verma G,
Srivastava Y, Masaldan S, Roy BG, Bharti AC and Das BC: Cervical
cancer stem cells selectively overexpress HPV oncoprotein E6 that
controls stemness and self-renewal through upregulation of HES1.
Clin Cancer Res. 22:4170–4184. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hou T, Zhang W, Tong C, Kazobinka G, Huang
X, Huang Y and Zhang Y: Putative stem cell markers in cervical
squamous cell carcinoma are correlated with poor clinical outcome.
BMC Cancer. 15:7852015. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Ye F, Zhou C, Cheng Q, Shen J and Chen H:
Stem-cell-abundant proteins nanog, nucleostemin and musashi1 are
highly expressed in malignant cervical epithelial cells. BMC
Cancer. 8:1082008. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
The 1988 Bethesda System for reporting
cervical/vaginal cytological diagnoses. National Cancer Institute
Workshop. JAMA. 262:931–934. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Li J and Zhou BP: Activation of β-catenin
and Akt pathways by Twist are critical for the maintenance of EMT
associated cancer stem cell-like characters. BMC Cancer. 11:492011.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Sato A, Ishiwata T, Matsuda Y, Yamammoto
T, Asakura H, Takeshita T and Naito Z: Expression and role of
nestin in human cervical intraepithelial neoplasia and cervical
cancer. Int J Oncol. 41:441–448. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Liu SY and Zheng PS: High aldehyde
dehydrogenase activity identifies cancer stem cells in human
cervical cancer. Oncotarget. 4:2462–2475. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Liu XF, Yang WT, Xu R, Liu JT and Zheng
PS: Cervical cancer cells with positive Sox2 expression exhibit the
properties of cancer stem cells. PLoS One. 9:e870922014. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Mei W, Lin X, Kapoor A, GU Y, Zhao K and
Tang D: the contributions of prostate cancer stem cells in prostate
cancer initiation and metastasis. Cancers (Basel). 11(pii):
E4342019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Lizarraga F, Espinosa M, Ceballos-Cancino
G, Vazquez-Santillan K, Bahena-Ocampo I, Schwarz-Cruz Y Celis A,
Vega-Gordillo M, Garcia Lopez P, Maldonado V and Melendez-Zajgla J:
Tissue inhibitor of metalloproteinases-4 (TIMP-4) regulates
stemness in cervical cancer cells. Mol Carcinog. 55:1952–1961.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Feng D, Yan K, Zhou Y, Liang H, Liang J,
Zhao W, Dong Z and Ling B: Piwil2 is reactivated by HPV
oncoproteins and initiates cell reprogramming via epigenetic
regulation during cervical cancer tumorigenesis. Oncotarget.
7:64575–64588. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Cao HZ, Liu XF, Yang WT, Chen Q and Zheng
PS: LGR5 promotes cancer stem cell traits and chemoresistance in
cervical cancer. Cell Death Dis. 8:e30392017. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Jiang J and Hui CC: Hedgehog signaling in
development and cancer. Dev Cell. 15:801–812. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Batsaikhan BE, Yoshikawa K, Kurita N,
Iwata T, Takasu C, Kashihara H and Shimada M: Cyclopamine decreased
the expression of Sonic Hedgehog and its downstream genes in colon
cancer stem cells. Anticancer Res. 34:6339–6344. 2014.PubMed/NCBI
|
|
97
|
Cochrane CR, Szczepny A, Watkins DN and
Cain JE: Hedgehog signaling in the maintenance of cancer stem
cells. Cancers (Basel). 7:1554–1585. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Rofstad EK, Sundfør K, Lyng H and Tropé
CG: Hypoxia-induced treatment failure in advanced squamous cell
carcinoma of the uterine cervix is primarily due to hypoxia-induced
radiation resistance rather than hypoxia-induced metastasis. Br J
Cancer. 83:354–359. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Hitoshi S, Alexson T, Tropepe V, Donoviel
D, Elia AJ, Nye JS, Conlon RA, Mak TW, Bernstein A and van der Kooy
D: Notch pathway molecules are essential for the maintenance, but
not the generation, of mammalian neural stem cells. Genes Dev.
16:846–858. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Gordon WR, Vardar-Ulu D, Histen G,
Sanchez-Irizarry C, Aster JC and Blacklow SC: Structural basis for
autoinhibition of Notch. Nat Struct Mol Biol. 14:295–300. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Takebe N, Miele L, Harris PJ, Jeong W,
Bando H, Kahn M, Yang SX and Ivy SP: Targeting Notch, Hedgehog, and
Wnt pathways in cancer stem cells: Clinical update. Nat Rev Clin
Oncol. 12:445–464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Venkatesh V, Nataraj R, Thangaraj G,
Karthikeyan M, Gnanasekaran A, Kaginelli SB, Kuppanna G, Kapalla CG
and Basalingappa KS: Targeting Notch signalling pathway of cancer
stem cells. Stem Cell Invest. 5:52018. View Article : Google Scholar
|
|
103
|
Zhu AJ and Watt FM: Beta-catenin
signalling modulates proliferative potential of human epidermal
keratinocytes independently of intercellular adhesion. Development.
126:2285–2298. 1999.PubMed/NCBI
|
|
104
|
Andrade AC, Nilsson O, Barnes KM and Baron
J: Wnt gene expression in the post-natal growth plate: Regulation
with chondrocyte differentiation. Bone. 40:1361–1369. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Blanpain C, Horsley V and Fuchs E:
Epithelial stem cells: Turning over new leaves. Cell. 128:445–458.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
van der Flier LG and Clevers H: Stem
cells, self-renewal, and differentiation in the intestinal
epithelium. Annu Rev Physiol. 71:241–260. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Ji J, Wei X and Wang Y: Embryonic stem
cell markers Sox-2 and OCT4 expression and their correlation with
WNT signal pathway in cervical squamous cell carcinoma. Int J Clin
Exp Pathol. 7:2470–2476. 2014.PubMed/NCBI
|
|
108
|
Barker N, Ridgway RA, van Es JH, van de
Wetering M, Begthel H, van de Born M, Danenberg E, Clarke AR,
Sanson OJ and Clevers H: Crypt stem cells as the cells-of-origin of
intestinal cancer. Nature. 457:608–611. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Gonzalez-Torres C, Gaytan-Cervantes J,
Vazquez-Santillan K, Mandujano-Tinoco EA, Ceballos-Cancino G,
Garcia-Venzor A, Zampedri C, Sanchez-Maldonado P, Mojica-Espinosa
R, Jimenez-Hernandez LE and Maldonado V: NF-κB participates in the
stem cell phenotype of ovarian cancer cells. Arch Med Res.
48:343–351. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Affara NI and Coussens LM: IKKalpha at the
crossroads of inflammation and metastasis. Cell. 129:25–26. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Lüningschrör P, Kaltschmidt B and
Kaltschmidt C: Knockdown of IKK1/2 promotes differentiation of
mouse embryonic stem cells into neuroectoderm at the expense of
mesoderm. Stem Cell Rev Rep. 8:1098–1108. 2012. View Article : Google Scholar
|
|
112
|
Porta C, Paglino C and Mosca A: Targeting
PI3K/Akt/mTOR signaling in cancer. Front Oncol. 4:642014.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Shayesteh L, Lu Y, Kuo WL, Baldocchi R,
Godfrey T, Collins C, Pinkel D, Powell B, Mills GB and Gray JW:
PIK3CA is implicated as an oncogene in ovarian cancer. Nat Genet.
21:99–102. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
114
|
Ma YY, Wei SJ, Lin YC, Lung JC, Chang TC,
Whang-Peng J, Liu JM, Yang DM, Yang WK and Shen CY: PIK3CA as an
oncogene in cervical cancer. Oncogene. 19:2739–2744. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Xia P and Xu X: PI3K/Akt/mTOR signaling
pathway in cancer stem cells: From basic research to clinical
application. Am J Cancer Res. 5:1602–1609. 2015.PubMed/NCBI
|














