Introduction
Drug resistance and toxicity to normal cells limit
the effectiveness of currently used cancer chemotherapies (1–3),
increasing the necessity for the development of new antitumor
agents. Natural products, including traditional Chinese medicine
(TCM), has recently received great interest since these agents have
relatively few side-effects and have been used as important
alternative cancer remedies for many years (4,5).
Patrinia scabiosaefolia is a perennial plant natively
distributed in Eastern Asia. As a well-known Oriental folk
medicine, it has long been used in China for the treatment of
edema, appendicitis, endometritis and other inflammatory illnesses
(6). Patrinia scabiosaefolia
is also used as an important component in several TCM formulas to
clinically treat cancers (7,8).
However, the precise mechanism of its antitumor activity remains
largely unknown.
Angiogenesis is a physiological process involving
the growth of new blood vessels from the pre-existing vasculature.
Although angiogenesis plays an essential role in a wide range of
physiological processes such as wound healing, reproduction and
embryonic development, deregulation of this vital biological
process is strongly associated with cancer progression (9–13). In
the initial stage, tumor cells obtain oxygen and nutrients from
nearby blood vessels by simple passive diffusion. However, when a
tumor grows to reach a certain size, >2 mm3, oxygen
delivery by diffusion is no longer sufficient, causing tumor cells
to induce the sprouting of new blood vessels from pre-existing
vasculature, creating a blood supply system within the tumor that
is essential for continue growth of the tumor as well as providing
an avenue for hematogenous metastasis (14–17).
Induction of angiogenesis is mediated by a variety of molecules
released by tumor cells (18–20),
including vascular endothelial growth factor A (VEGF-A) which is
considered as one of the strongest stimulators of angiogenesis
(21–23). VEGF-A is commonly overexpressed in
many types of human cancer, and is correlated with tumor
progression, invasion and metastasis, and poorer survival and
prognosis in patients (24–27). VEGF-A exerts its biologic effect
primarily through interaction with specific receptors present on
the surface of vascular endothelial cells (ECs), which in turn
triggers a tyrosine kinase signaling cascade, inducing EC
proliferation, migration, survival, sprouting and eventually tube
formation (21,24,27,28).
Due to the key role of angiogenesis in the progression of solid
tumors, inhibition of tumor angiogenesis has become a major focus
of anticancer drug development.
To elucidate the mechanism of the tumorcidal
activity of Patrinia scabiosaefolia, using a colorectal
cancer (CRC) mouse xenograft model, the human colon carcinoma cell
line HT-29 and human umbilical vein endothelial cells (HUVECs), in
this study we evaluated its effect on tumor angiogenesis in
vivo and in vitro, and investigated the underlying
molecular mechanisms.
Materials and methods
Materials and reagents
Roswell Park Memorial Institute (RPMI)-1640 medium,
Dulbecco’s modified Eagle’s medium (DMEM), fetal bovine serum
(FBS), penicillin-streptomycin, Trypsin-EDTA, and TRIzol reagent
were purchased from Invitrogen Life Technologies (Carlsbad, CA,
USA). SuperScript II reverse transcriptase was obtained from
Promega Corporation (Madison, WI, USA). CD31 and VEGF-A antibodies,
and secondary antibodies were obtained from Cell Signaling
Technology, Inc. (Beverly, MA, USA). The In vitro
angiogenesis assay kit was purchased from Millipore (Billerica, MA,
USA). The Human VEGF-A ELISA kit was obtained from Shanghai Xitang
Biological Technology Ltd. (Shanghai, China). All other chemicals,
unless otherwise stated, were obtained from Sigma-Aldrich (St.
Louis, MO, USA).
Preparation of ethanol extract from
Patrinia scabiosaefolia
EEPS was prepared as described previously (8). For animal experiments, EEPS powder was
dissolved in saline to a working concentration of 250 mg/ml. In
cell-based experiments, EEPS powder was dissolved in 50% DMSO to a
stock concentration of 250 mg/ml and the working concentrations of
EEPS were made by diluting the stock solution in the culture
medium. The final concentration of DMSO in the medium for all cell
experiments was <0.5%.
Cell culture
Human colon carcinoma HT-29 cells and HUVECs were
obtained from the Cell Bank of the Chinese Academy of Science
(Shanghai, China). HUVECs or HT-29 cells were grown in RPMI-1640 or
DMEM, respectively. RPMI-1640 and DMEM were supplemented with 10%
(v/v) FBS, and 100 U/ml penicillin and 100 μg/ml streptomycin.
Cells were cultured at 37°C humidified incubator with 5%
CO2.
Animals
Male BALB/c athymic (nude) mice (with an initial
body weight of 20–22 g) were obtained from the Shanghai SLAC
Laboratory Animal Co., Ltd. (Shanghai, China) and housed under
pathogen-free conditions with controlled temperature (22°C),
humidity and a 12-h light/dark cycle. Food and water were given
ad libitum throughout the experiment. All animal treatments
were strictly in accordance with international ethical guidelines
and the National Institutes of Health Guide concerning the Care and
Use of Laboratory Animals, and the experiments were approved by the
Institutional Animal Care and Use Committee of Fujian University of
Traditional Chinese Medicine.
In vivo tumor xenograft study
HT-29 cells (1.5×106) mixed with Matrigel
(1:1) were subcutaneously injected into the right flank of the mice
to initiate tumor growth. After 5 days of xenograft implantation,
mice were randomly divided into 2 groups (n=6) and given
intragastric administration with 1.93 g/kg/day of EEPS or saline
daily, 5 days a week for 21 days. Tumor size was determined by
measuring the major (L) and minor (W) diameter with a caliper. The
tumor volume was calculated according to the following formula:
tumor volume = π/6 × L × W2.
Immunohistochemical staining
After being fixed with 10% formaldehyde for 12 h,
tumor samples were processed conventionally for paraffin-embedded
tumor slides. The slides were subjected to antigen retrieval, and
the endogenous peroxidase activity was quenched with hydrogen
peroxide. After blocking non-specific proteins with normal serum in
phosphate-buffered saline (PBS) (0.1% Tween-20), slides were
incubated with rabbit polyclonal antibodies against CD31 and VEGF-A
(1:200 dilution). After washing with PBS, slides were incubated
with a biotinylated secondary antibody followed by conjugated
horseradish peroxidase (HRP)-labeled streptavidin (Dako), and then
washed with PBS. The slides were then incubated with
diaminobenzidine (DAB) (Sigma) as the chromogen, followed by
counterstaining with diluted Harris’ hematoxylin (Sigma). After
staining, 5 high-power fields (x400) were randomly selected in each
slide, and the average proportion of positive cells in each field
was counted using a true color multi-functional cell image analysis
management system (Image-Pro Plus; Media Cybernetics, Rockville,
MD, USA). To rule out any nonspecific staining, PBS was used to
replace the primary antibody as a negative control.
Evaluation of cell viability by MTT
assay
Viability of HUVECS was assessed by the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
colorimetric assay. HUVECs were seeded into 96-well plates at a
density of 1×104 cells/well in 0.1 ml medium. The cells
were treated with various concentrations of EEPS for 24, 48 and 72
h respectively. At the end of the treatment, 100 μl MTT (0.5 mg/ml
in PBS) was added to each well, and the samples were incubated for
an additional 4 h at 37°C. The purple-blue MTT formazan precipitate
was dissolved in 100 μl DMSO. The absorbance was measured at 570 nm
using an ELISA reader (BioTek, Model ELX800; BioTek Instruments,
Inc., Winooski, VT, USA).
Migration assay of HUVECs
Migration of HUVECs was performed by the wound
healing method. HUVECs were seeded into 12-well plates at a density
of 2×105 cells/well in 1 ml medium. After 24 h of
incubation, cells were scraped away vertically in each well using a
P100 pipette tip. Three randomly selected views along the scraped
line were photographed for each well using a phase-contrast
inverted microscope at a magnification of ×100. Cells were then
treated with the indicated concentrations of EEPS for 24 h, and
another set of images was captured using the same method. A
reduction in the scraped area indicated migratory capability.
Tube formation assay of HUVECs
The tube formation of HUVECs was examined using the
ECMatrix assay kit (Millipore), following the manufacturer’s
instructions. Briefly, confluent HUVECs were harvested and diluted
(1×104 cells) in 50 μl of medium, containing indicated
concentrations of EEPS. The harvested cells were then seeded into
1:1 ECMatrix gel (v/v)-coated 96-well plates and incubated for 9 h
at 37°C. The network-like structures were examined using a
phase-contrast inverted microscope. The images were captured at a
magnification of ×100. The level of tube formation was quantified
by calculating the length of tubes in 3 randomly chosen fields from
each well.
Measurement of VEGF-A secretion in HT-29
cells by ELISA
HT-29 cells were seeded into 6-well plates at a
density of 2×105 cells/well in 2 ml medium and treated
with the indicated concentrations of EEPS for 24 h. The level of
VEGF-A in the media was measured using an ELISA kit (XiTang
Biological Technology Ltd., Shanghai, China) according to the
manufacturer’s instructions. The concentration of VEGF-A was
determined by comparison to serial dilutions of a purified standard
of VEGF-A.
RT-PCR analysis
Total RNA was isolated from tumor tissues or HT-29
cells with TRIzol reagent. Oligo(dT)-primed RNA (1 μg) was
reverse-transcribed with SuperScript II reverse transcriptase
(Promega Corporation) according to the manufacturer’s instructions.
The obtained cDNA was used to determine the mRNA amount of VEGF-A
by PCR with Taq DNA polymerase (Fermentas). GAPDH was used
as an internal control.
Statistical analysis
Data are presented as means ± SD for the indicated
number of independently performed experiments and were analyzed
using the SPSS Package for Windows (version 16.0). Statistical
analysis of the data was performed with the Student’s t-test and
ANOVA. Differences with P<0.05 were considered to be
statistically significant.
Results and Discussion
EEPS inhibits tumor growth in colorectal
cancer (CRC) xenograft mice
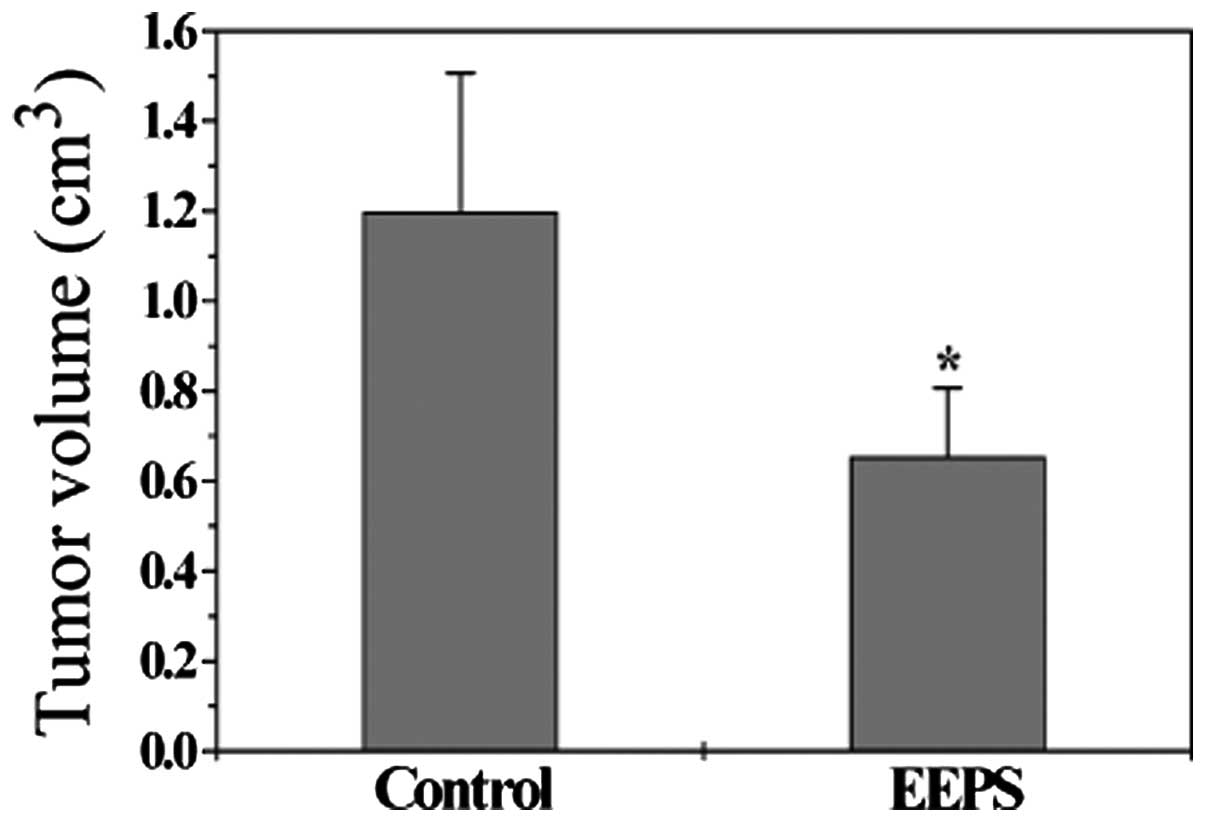
We evaluated the tumor growth in CRC xenograft mice
by measuring the tumor volume. Data in Fig. 1 indicate that EEPS treatment
significantly suppressed tumor growth, as compared with the control
group. The final tumor volume/mouse in the control and the
EEPS-treated group was 1.20±0.31 and 0.65±0.15 cm3,
respectively (P<0.05), suggesting that EEPS was effective in the
treatment of colorectal cancer.
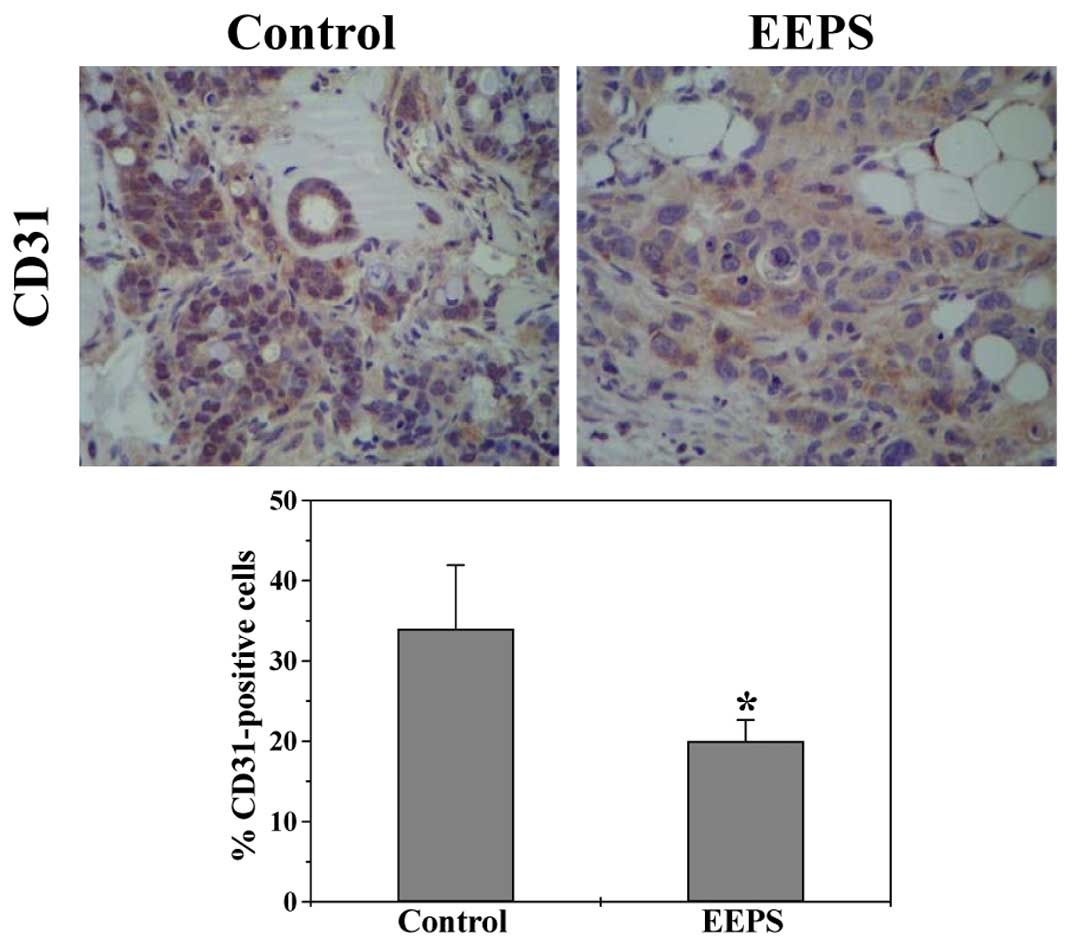
EEPS displays anti-angiogenic activity in
CRC xenograft mouse tumors and in human umbilical vein endothelial
cells
Angiogenesis plays an important role in the
progression and metastasis of cancer, which thus has become an
attractive target for anticancer therapy. The expression of
endothelial cell-specific marker CD31 in CRC tumor tissues was
determined by immunohistochemical staining (IHS) to evaluate the
effect of EEPS on intratumoral microvessel density (MVD), an
indicator of new vessel growth. As shown in Fig. 2, the percentage of CD31-positive
cells in the EEPS-treated mouse tumors was significantly reduced
(P<0.05), demonstrating the in vivo anti-angiogenic
activity of EEPS.
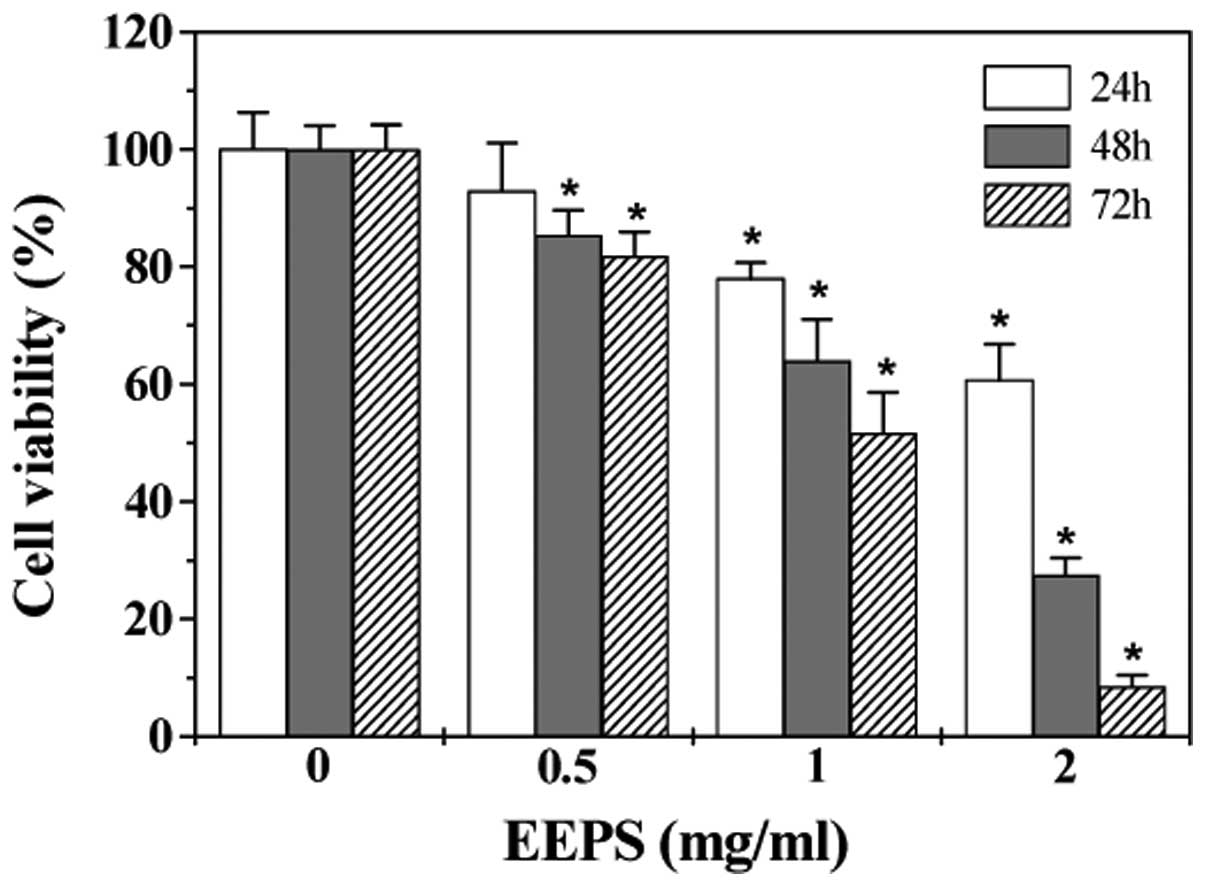
Angiogenesis typically consists of several features,
including proliferation, migration and eventually the capillary
tube formation of endothelial cells (ECs). To further evaluate the
anti-angiogenic effect of EEPS we modeled each of these processes
with HUVECs in vitro. As shown in Fig. 3, EEPS treatment dose- and
time-dependently decreased the proliferation (viability) of HUVECs
(P<0.05, compared to untreated control cells). Moreover, EEPS
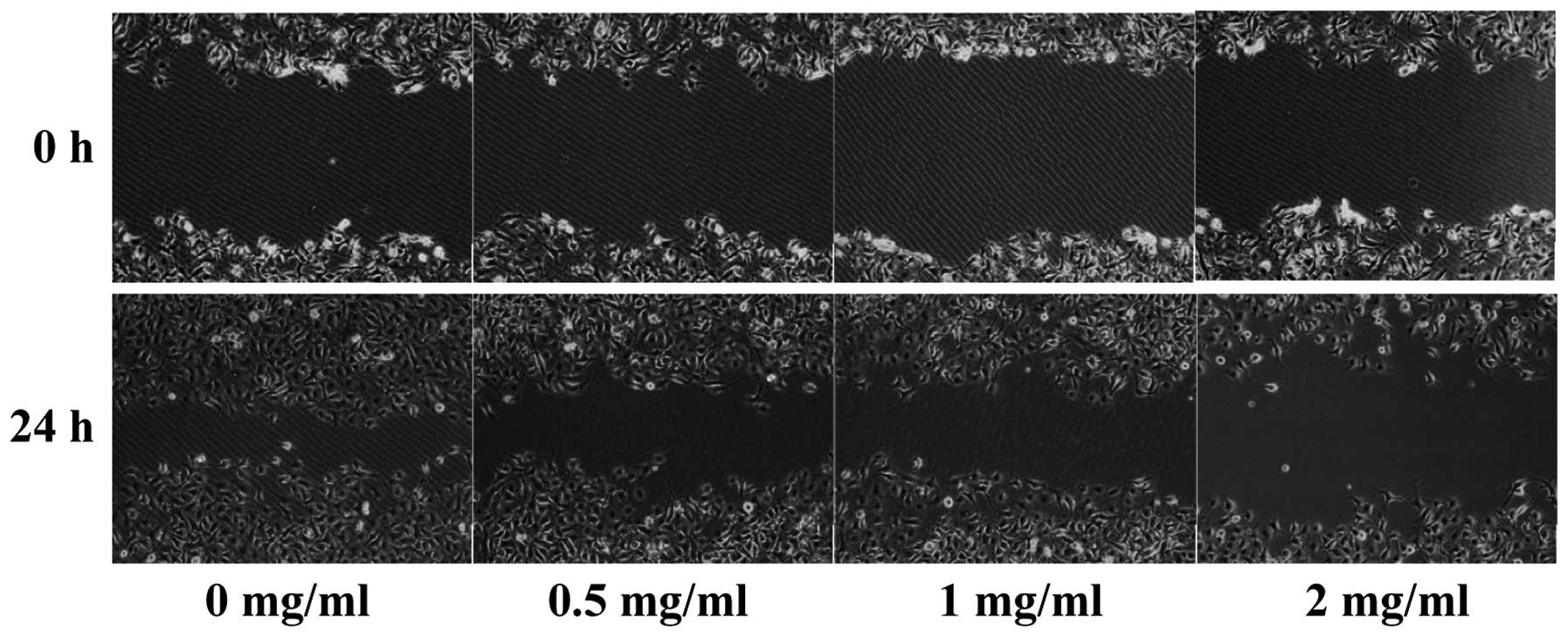
treatment inhibited HUVEC migration after monolayer wounding
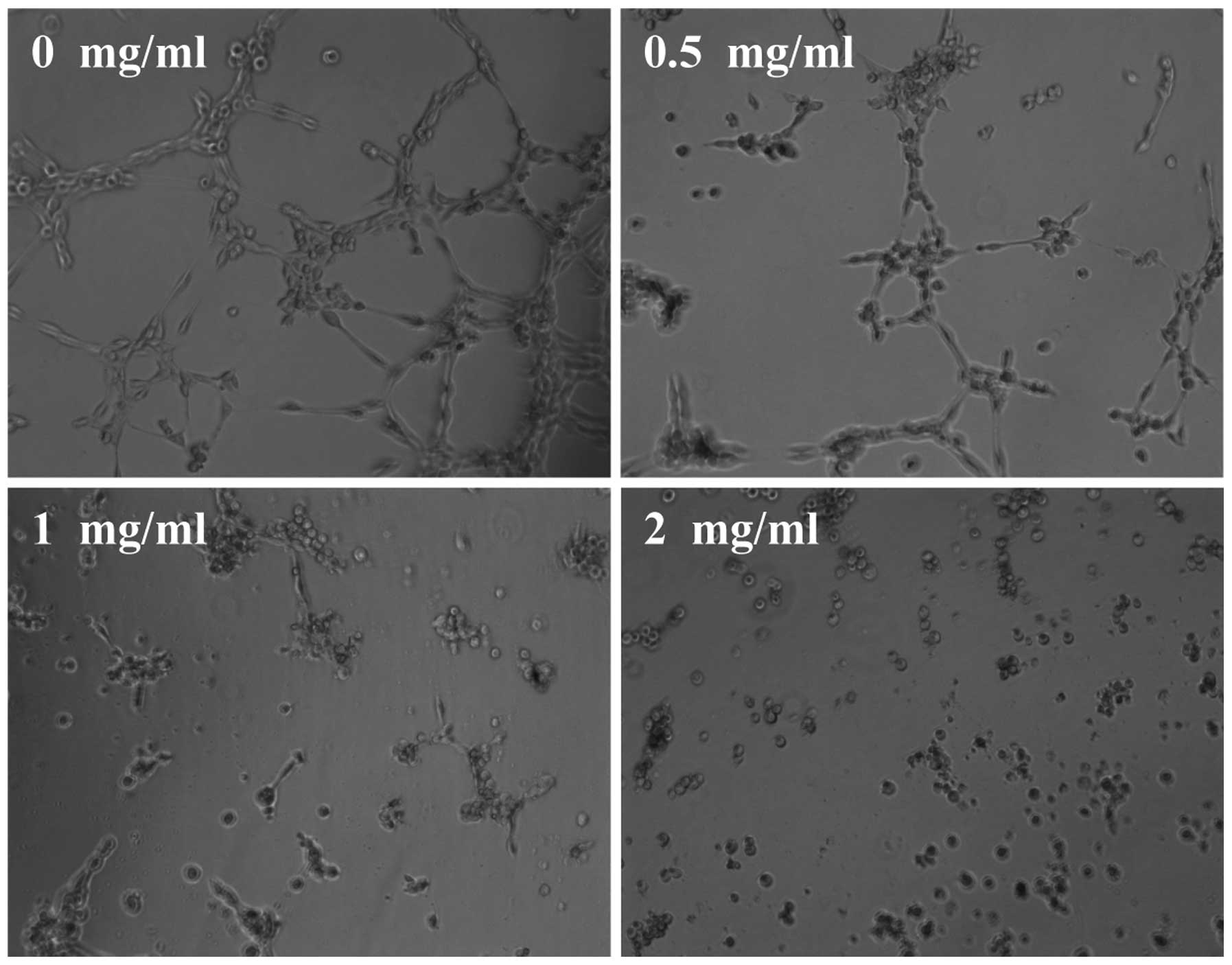
(Fig. 4). Furthermore, we examined
the effect of EEPS on capillary tube formation of HUVECs using an
extracellular matrix. As shown in Fig.
5, untreated HUVECs formed elongated tube-like structures;
which, however, was profoundly inhibited by EEPS treatment. Taken
together, it is suggested that the antitumor angiogenesis activity
of EEPS could have contributed to its inhibitory effect on
colorectal tumor growth.
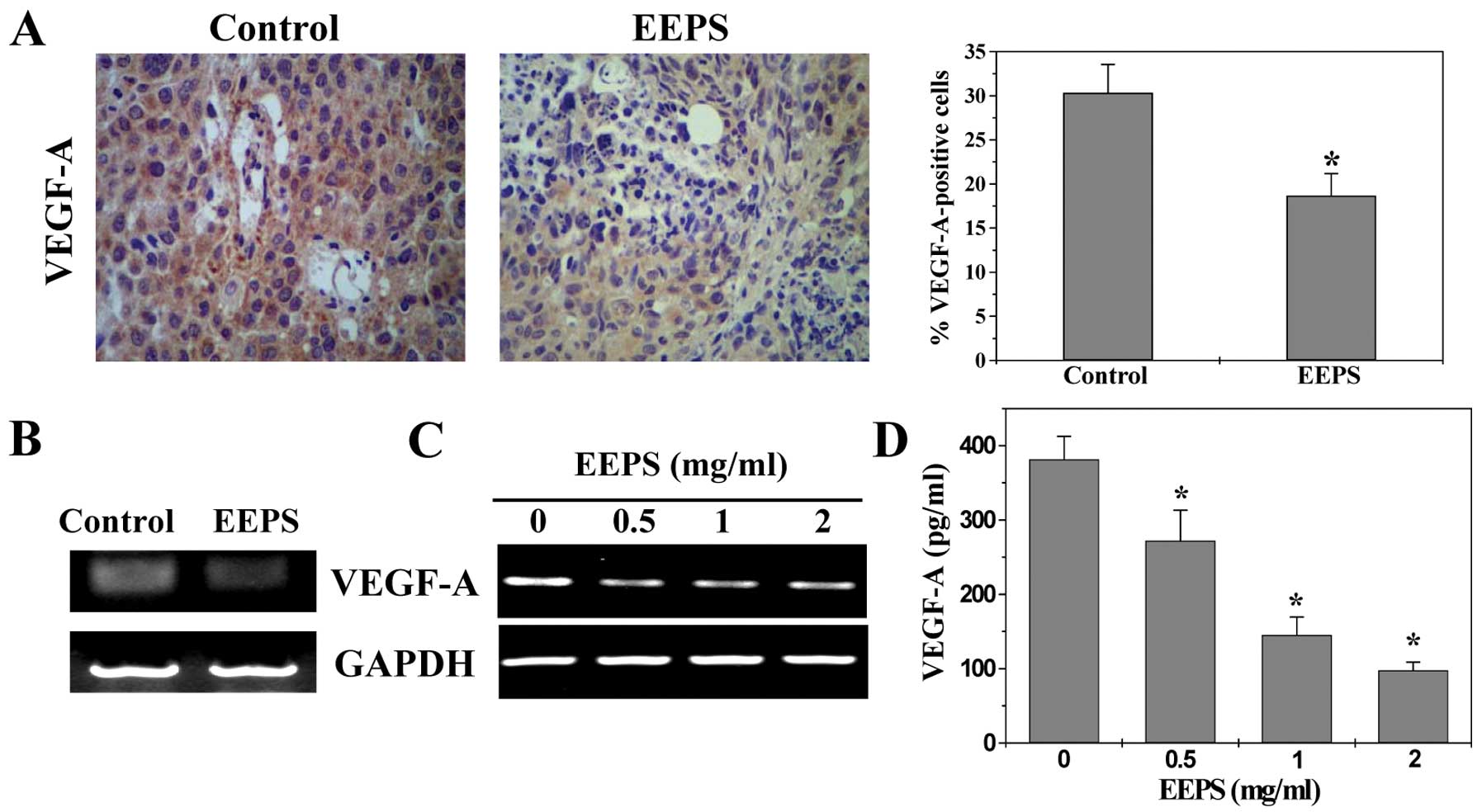
EEPS decreases VEGF-A expression in CRC
xenograft mouse tumors and human colon carcinoma HT-29 cells
The process of tumor angiogenesis is tightly
regulated by a variety of molecules released by tumor cells.
Vascular endothelial growth factor A (VEGF-A) is considered as one
of the most potent angiogenic stimulators. Overexpression of VEGF-A
is found in numerous types of cancer and commonly suggests poor
prognosis. VEGF-A functions primarily via a paracrine pathway,
binding to its specific receptors located on vascular ECs after
secretion from tumor cells, leading to a series of angiogenic
processes. To further investigate the mechanism of the
anti-angiogenic activity of EEPS, we examined its effect on the
expression of VEGF-A. As shown in Fig.
6, EEPS treatment obviously decreased the mRNA and protein
expression levels of VEGF-A both in CRC tumor tissues and in HT-29
cells (P<0.01).
In conclusion, here for the first time we
demonstrated that Patrinia scabiosaefolia inhibits
colorectal cancer growth via inhibition of tumor angiogenesis,
which may in part explain its anticancer activity.
Acknowledgements
This study was sponsored by the National Natural
Science Foundations of China (nos. 81173272 and 81073097).
Abbreviations:
|
EEPS
|
ethanol extract of Patrinia
scabiosaefolia
|
|
CRC
|
colorectal cancer
|
|
IHS
|
immunohistochemical staining
|
|
VEGF-A
|
vascular endothelial growth factor
A
|
|
MVD
|
microvessel density
|
References
|
1
|
Gorlick R and Bertino JR: Drug resistance
in colon cancer. Semin Oncol. 26:606–611. 1999.
|
|
2
|
Boose G and Stopper H: Genotoxicity of
several clinically used topoisomerase II inhibitors. Toxicol Lett.
116:7–16. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Longley DB, Allen WL and Johnston PG: Drug
resistance, predictive markers and pharmacogenomics in colorectal
cancer. Biochim Biophys Acta. 1766:184–196. 2006.PubMed/NCBI
|
|
4
|
Newman DJ, Cragg GM and Snader KM: The
influence of natural products upon drug discovery. Nat Prod Rep.
17:215–234. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ju HK, Baek SH, An RB, Bae K, Son KH, Kim
HP, Kang SS, Lee SH, Son JK and Chang HW: Inhibitory effects of
nardostachin on nitric oxide, prostaglandin E2, and tumor necrosis
factor-α production in lipopolysaccharide-activated macrophages.
Biol Pharm Bull. 26:1375–1378. 2003.PubMed/NCBI
|
|
7
|
Chiu LC, Ho TS, Wong EY and Ooi VE: Ethyl
acetate extract of Patrinia scabiosaefolia downregulates
anti-apoptotic Bcl-2/Bcl-X(L) expression, and induces apoptosis in
human breast carcinoma MCF-7 cells independent of caspase-9
activation. J Ethnopharmacol. 105:263–268. 2006.PubMed/NCBI
|
|
8
|
Peng J, Chen YQ, Lin JM, Zhuang ZQ, Xu W,
Hong ZF and Sferra TJ: Patrinia scabiosaefolia extract
suppresses proliferation and promotes apoptosis by inhibiting the
STAT3 pathway in human multiple myeloma cells. Mol Med Rep.
4:313–318. 2011.
|
|
9
|
Folkman J: Tumor angiogenesis: therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Folkman J and Shing Y: Angiogenesis. J
Biol Chem. 267:10931–10934. 1992.
|
|
11
|
Folkman J: Angiogenesis in cancer,
vascular, rheumatoid and other disease. Nat Med. 1:27–31. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Folkman J: Angiogenesis. Annu Rev Med.
57:1–18. 2006. View Article : Google Scholar
|
|
13
|
Cook KM and Figg WD: Angiogenesis
inhibitors: current strategies and future prospects. CA Cancer J
Clin. 60:222–243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer-related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar
|
|
15
|
Whiteside TL: The tumor microenvironment
and its role in promoting tumor growth. Oncogene. 27:5904–5912.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jain RK: Transport of molecules in the
tumor interstitium: a review. Cancer Res. 47:3039–3051.
1987.PubMed/NCBI
|
|
17
|
Folkman J: How is blood vessel growth
regulated in normal and neoplastic tissue? GHA Clowes memorial
Award lecture. Cancer Res. 46:467–473. 1986.PubMed/NCBI
|
|
18
|
Strömblad S and Cheresh DA: Integrins,
angiogenesis and vascular cell survival. Chem Biol. 3:881–885.
1996.PubMed/NCBI
|
|
19
|
Breier G and Risau W: The role of vascular
endothelial growth factor in blood vessel formation. Trends Cell
Biol. 6:454–456. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Weidner N, Semple JP, Welch WR and Folkman
J: Tumor angiogenesis and metastasis - correlation in invasive
breast carcinoma. N Engl J Med. 324:1–8. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ferrara N: Role of vascular endothelial
growth factor in physiologic and pathologic angiogenesis:
therapeutic implications. Semin Oncol. 29:10–14. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jain RK: Tumor angiogenesis and
accessibility: role of vascular endothelial growth factor. Semin
Oncol. 29:3–9. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kaya M, Wada T, Akatsuka T, Kawaguchi S,
Nagoya S, Shindoh M, Higashino F, Mezawa F, Okada F and Ishii S:
Vascular endothelial growth factor expression in untreated
osteosarcoma is predictive of pulmonary metastasis and poor
prognosis. Clin Cancer Res. 6:572–577. 2000.PubMed/NCBI
|
|
26
|
Maeda K, Chung YS, Ogawa Y, Takatsuka S,
Kang SM, Ogawa M, Sawada T and Sowa M: Prognostic value of vascular
endothelial growth factor expression in gastric carcinoma. Cancer.
77:858–863. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ishigami SI, Arii S, Furutani M, Niwano M,
Harada T, Mizumoto M, Mori A, Onodera H and Imamura M: Predictive
value of vascular endothelial growth factor (VEGF) in metastasis
and prognosis of human colorectal cancer. Br J Cancer.
78:1379–1384. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gille H, Kowalski J, Li B, LeCouter J,
Moffat B, Zioncheck TF, Pelletier N and Ferrara N: Analysis of
biological effects and signaling properties of Flt-1 (VEGFR-1) and
KDR (VEGFR-2). A reassessment using novel receptor-specific
vascular endothelial growth factor mutants. J Biol Chem.
276:3222–3230. 2001. View Article : Google Scholar
|