Introduction
The use of plant-derived natural products for
medical benefits is playing an important role globally. Several
anticancer drugs available on the market today such as Taxol,
Oncovin, Navelbine and Vumon trace their origins to plants
(1–3). In cancer therapy, the focus is on
strategies that suppress tumor growth through cell cycle disruption
(4) and activate the apoptotic
program in the cell (5).
Several essential oils from plants possess medical
benefits. Among the various constituents of essential oils, 1,
8-cineole has been shown to possess pharmacological effects. The
content of 1, 8-cineole in the essential oils varies in the
different Eucalyptus species, from 25 to 90% (6,7). 1,
8-cineole has been used as a percutaneous penetration enhancer
(8), as an antibacterial and
expectorant (6), anti-inflammatory
(9,10) or antihypertensive (11) agent. 1, 8-cineole was reported to
induce apoptosis in leukemia cell lines (12).
In this study, we found that 1, 8-cineole exerts
antitumor activity on human colon cancer cell lines HCT116 and RKO.
We investigated whether 1, 8-cineole induces apoptosis in two human
colorectal cancer cell lines and in a xenograft model.
Materials and methods
Reagents
1, 8-cineole (eucalyptol) was obtained from Sigma
(St. Louis, MO, USA). For cell culture experiment, 1, 8-cineole was
dissolved in ethanol and then in water at a concentration of 0.1
mg/ml. In an in vitro experiment, 5–50 mM of 1, 8-cineole
was added to the medium and cell viability assay and western blot
experiments were performed 24 h later.
Cells
The human CRC cell lines HCT116 and RKO were used
and tested for mycoplasma-free cell lines. The cancer cell lines
were subdivided into multiple tubes for stock in liquid nitrogen
immediately after possession. All cell lines were subjected to the
present experiment within 6 months of resuscitation. Stock cultures
were grown in high-glucose DMEM containing 10% FBS and 1%
antibiotics. The cells were grown in growth medium at 37°C in a 95%
air, 5% CO2-humidified incubator.
Cell viability assay
To measure the cytotoxicity of 1, 8-cineole against
these cancer cells, 3×103 cells were plated per well
onto 96-well plates. Following overnight culture, 1, 8-cineole and
oxaliplatin were added at specified concentrations. After 24 h
incubation, cell viability was measured by the mitochondrial
activity in reducing
2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium
monosodium salt (WST-8) to formazan using a Cell Counting kit-8
(Dojindo Laboratories, Kumamoto, Japan). Cells were incubated with
a reagent as per the manufacturer's instructions. Plates were read
at A450 on a spectrometer.
Cell proliferation assay
To measure the cell proliferation activity of 1,
8-cineole against HCT116 and RKO cancer cells, 3×103
cells were plated per well onto 96-well plates. Following overnight
culture, 1, 8-cineole was added at specified concentrations. After
24 h of incubation, cell proliferation was measured with a BrdU
assay kit (Roche Diagnostics, Penzberg, Germany). Cells were
incubated with a reagent as per the manufacturer's instructions.
Plates were read at A450 on a spectrometer.
Lactate dehydrogenase (LDH) activity
assay
In order to evaluate the activity of the cytoplasmic
enzyme lactate dehydrogenase (LDH) released from the cytosol when
cells were damaged under stress, HCT116 and RKO cells were seeded
(1×105 cells/ml) on 96-well plates. The LDH activity was
determined using a commercial kit (Takara Bio, Tokyo, Japan).
Absorbance values were then correlated with the number of viable
cells to predict the cytotoxic activity. Triton X-100 (1%) (Sigma)
was used as a positive control.
Apoptosis assay
The In Situ Cell Death Detection kit (Roche
Diagnostics, Basel, Switzerland) was used for the demonstration of
apoptotic cell death of cell culture and liver tissue. Cells
(3×104) were plated per well onto Lab-Tek II Chamber
Slides (Nalge Nunc International, Tokyo, Japan) and were incubated
with the terminal deoxynucleotidyl transferase-mediated dUTP nick
end labeling (TUNEL) reaction mixture according to the
manufacturer's recommendations.
Western blot analysis
For western blot analysis, total protein extracts of
HCT116 cells were obtained 24 h after 1, 8-cineole treatment and
separated by 10% SDS-PAGE and transferred to nitrocellulose
membrane (Millipore, Bedford, MA, USA). The following antibodies
were used as primary antibodies: cleaved caspase-3 (9661), cleaved
poly(ADP-ribose) polymerase (PARP) (5625p), survivin (2808p),
phosphoserine 473 Akt (9271), phospho p38 (4511p) and GAPDH (2118)
(Cell Signaling Technology, Beverly, MA, USA). Secondary goat
anti-rabbit antibody conjugated with horseradish peroxidase was
purchased from Cell Signaling Technology.
Animals
Seven-week-old male severe combined immunodeficiency
(SCID) mice (Clea, Tokyo, Japan), weighing 24–28 g, were utilized.
The mice were kept in a temperature-controlled room on a 12-h
light-dark cycle. They had free access to water and standard chow
throughout the experiment. After an acclimation period of ≥7 days,
the mice were separated into two groups as follows: control group,
mice without any treatment (n=10); and 1, 8-cineole group, mice
with 1, 8-cineole treatment (n=9). All animal experiments were
carried out in a humane manner after receiving approval from the
Institutional Animal Committee of Teikyo University and in
accordance with the Regulation for Animal Experiments of the
University and Fundamental Guidelines for Proper Conduct of Animal
Experiments and Related Activities in Academic Research
Institutions under the jurisdication of the Ministry of Education,
Culture, Sports, Science and Technology of Japan.
Xenograft
Cells (2×106) of RKO were injected
subcutaneously into the right flank of each mouse with a 27-gauge
needle. Tumors were detected by palpation and measured periodically
with calipers. Seven days after tumor injection, 50 mg/kg of 1,
8-cineole was injected subcutaneously every 3 days. Twenty-one days
after inoculation, mice were sacrificed and tumors were removed for
examination.
Statistical analysis
All data are expressed as the mean ± SD of samples.
Comparisons between various points were made using one-way ANOVA.
Comparisons between the two groups were made using Mann-Whitney U
test. Significant data were examined by the Bonferroni-Dunn
multiple comparisons post hoc test. In all cases, a P-value
<0.05 was considered statistically significant.
Results
Dose-dependent inhibition of
proliferation of human CRC cell lines by 1, 8-cineole
We initially determined whether 1, 8-cineole
treatment led to the inhibition of human CRC cell proliferation.
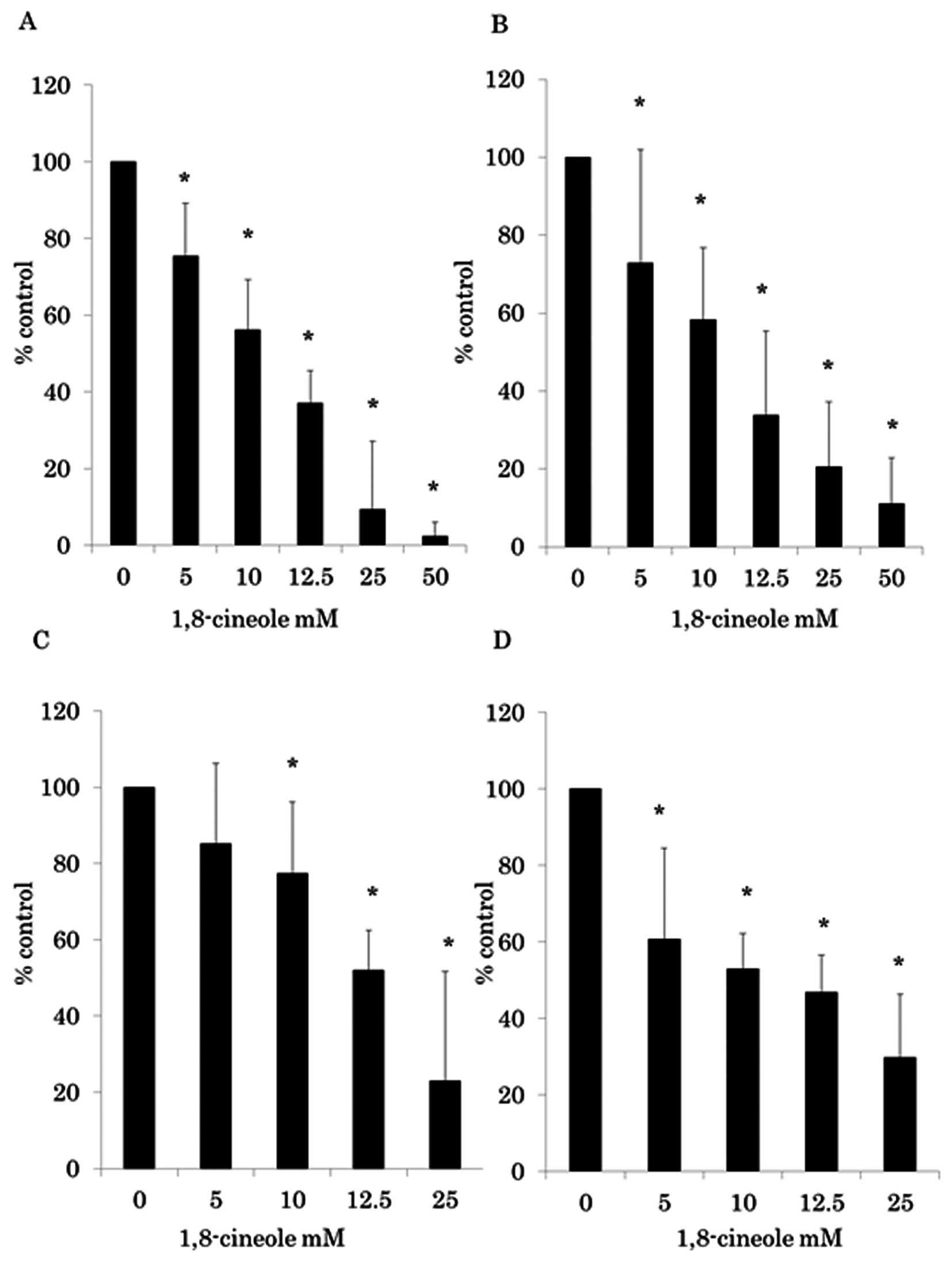
CRC cells were treated with various doses of 1, 8-cineole for 24 h
and cell viability was assayed using WST-8 assay (Fig. 1A and B) and BrdU assay (Fig. 1C and D). Fig. 1 shows that as the dose of 1,
8-cineole increased from 5 to 50 mM, cell growth inhibition
increased in a dose-dependent fashion in CRC cell lines HCT116 and
RKO. 1, 8-cineole-induced growth inhibition was found to be
statistically significant (p<0.01) (one-way ANOVA) in 5–50 mM of
1, 8-cineole compared to 0 mM in WST-8 assay and 5–25 mM in BrdU
assay.
1, 8-cineole does not induce
necrosis
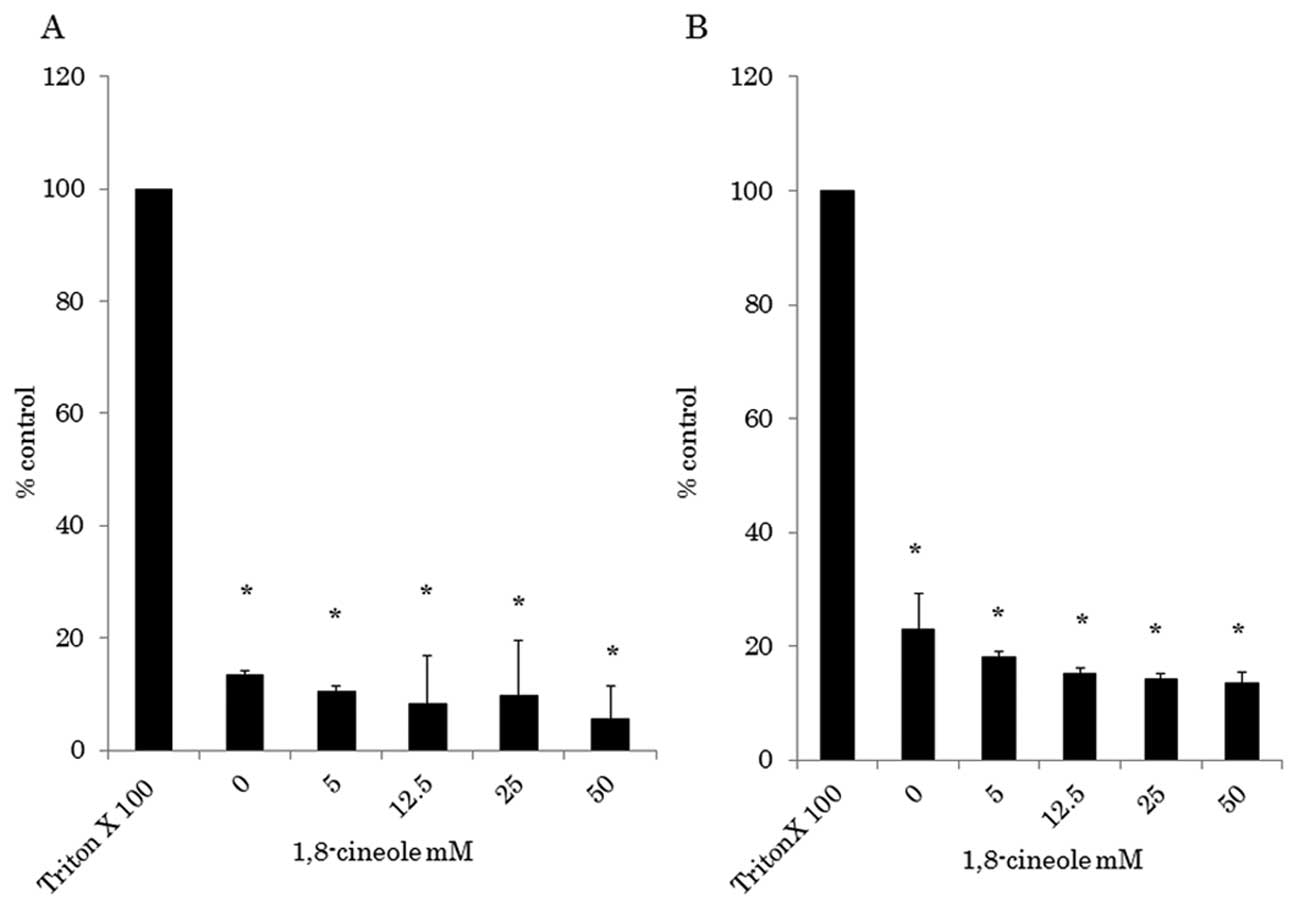
Fig. 2 shows the
cytoplasmic LDH released of HCT116 and RKO cells treated with each
sample at 5–50 mM of 1, 8-cineole. Compared to culture medium
(negative control), no statically significant increase in LDH
released by these cells was observed. However, all samples showed a
statistically significant difference (p<0.01) in LDH released
when compared to 1% Triton X-100 (positive control). Damaged cells
or those under stress can release cytoplasmic LDH and other
substances into the medium due to disruption of the cytoplasmic
membrane and cell necrosis (13).
Therefore, it is possible that the cytotoxic activity of the
samples was not based on the mechanism of cell death by necrosis.
Consequently, these results indicate that the cytotoxic activity of
the samples can involve pathways inducing cell death by
apoptosis.
Induction of apoptosis via activation of
caspase-dependent pathway by 1, 8-cineole
In subsequent experiments, we determined the
mechanism of the observed suppressive effect of 1, 8-cineole by
WST-8 and BrdU assays. The overexpression of survivin has been
observed to cooperate with survival pathways, including the
phosphatidylinositol-3-kinase (PI3K)/Akt pathway. Suppression of
survivin and constitutive activated Akt and activation of p38 was
observed in 12.5–50 mM of 1, 8-cineole. These molecules induced
increments of cleaved PARP and caspase-3 in 1, 8-cineole treatment
(Fig. 3A). TUNEL staining of RKO
cells showed apoptotic cells in 50 mM of 1, 8-cineole (Fig. 3B).
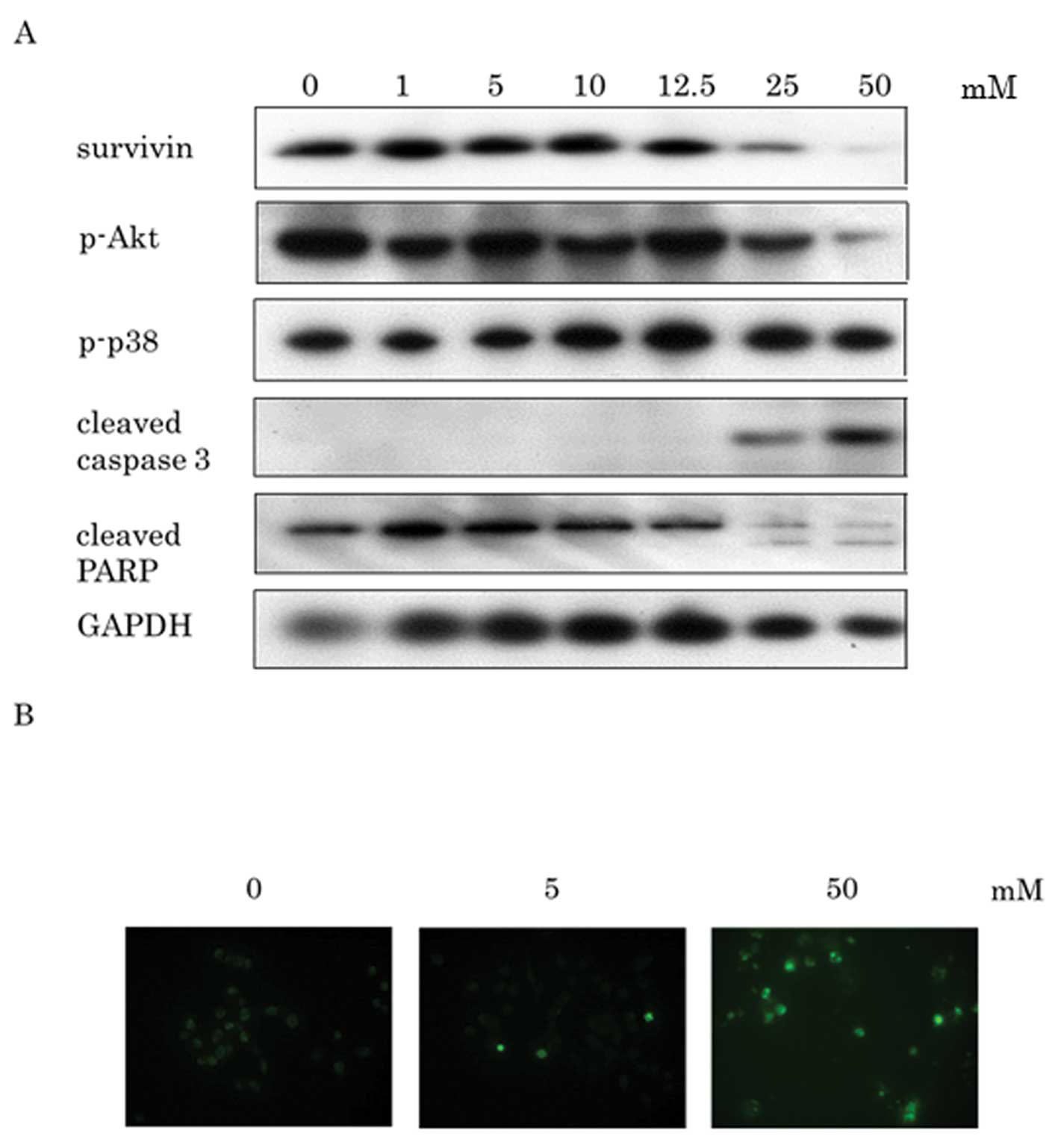 | Figure 3(A) RKO cells were treated with 0–50
mM of 1, 8-cineole for 24 h. After cell lysis, equal amounts of
proteins were separated by SDS-PAGE, transferred to Immobilon
membrane and immunoblotted with antibodies against survivin, p-Akt,
p-p38, cleaved caspase-3, cleaved PARP and GAPDH as indicated. 1,
8-cineole treatment caused suppression of survivin,
dephosphorylation of constitutive phosphorylation of Akt, p38
activation and cleavage of PARP and caspase-3. (B) TUNEL staining
of RKO cells, which were treated with 0, 5 and 50 mM of 1,
8-cineole. Stained cells indicate TUNEL positive apoptotic
cells. |
1, 8-cineole inhibits tumor growth of RKO
xenografts
We evaluated the potential effectiveness of 1,
8-cineole in a xenograft model of RKO, which was subcutaneously
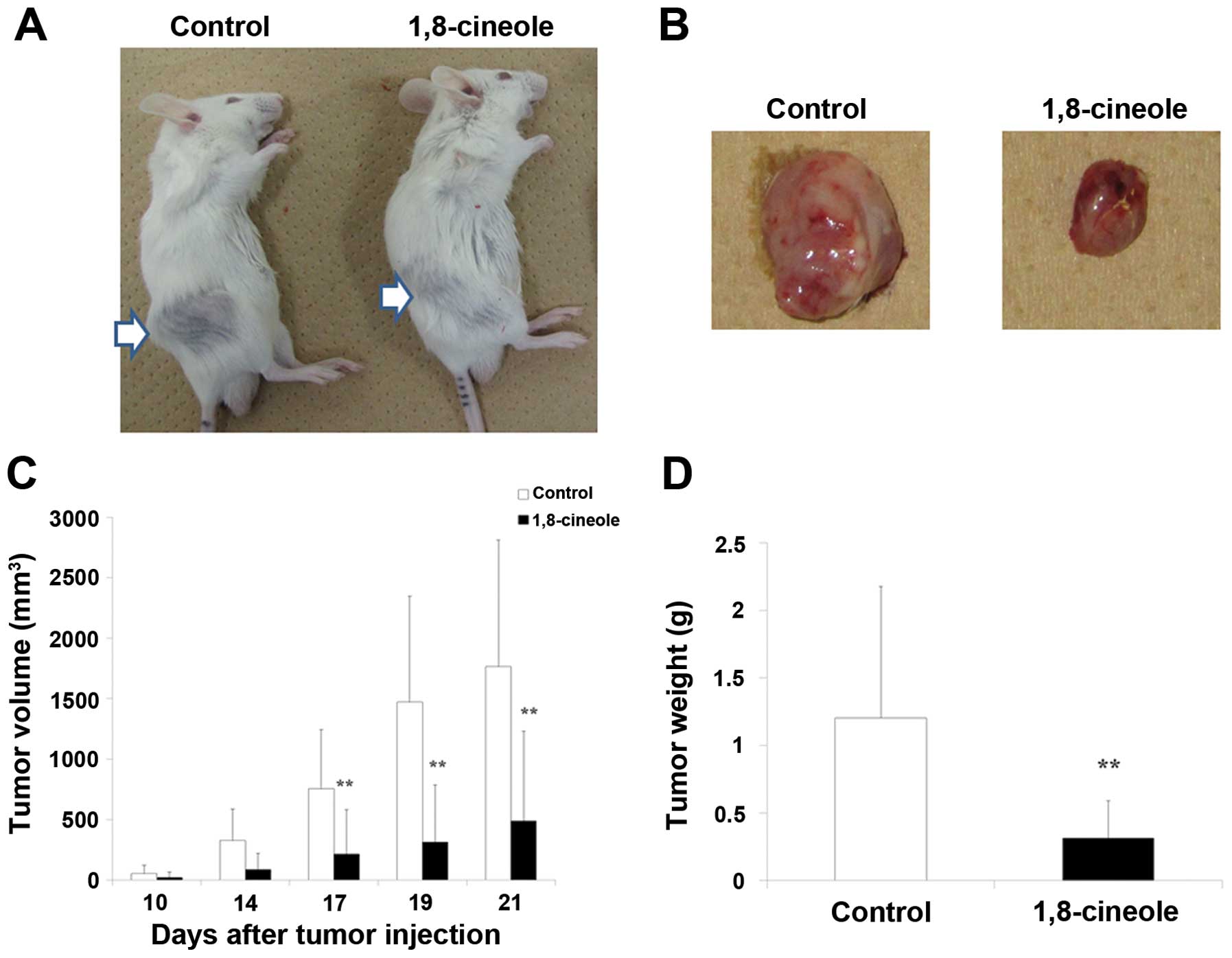
injected into the right flank of each mouse. Fig. 4A shows representative mice of each
group. No skin damage was observed in the 1, 8-cineole group.
Fig. 4B shows tumors removed from
the representative control and mice with 1, 8-cineole groups.
Fig. 4C indicates tumor growth
curve. Fig. 4D indicates tumor
weight in the 2 groups. Tumor growth was significantly inhibited in
the 1, 8-cineole group, compared to the control group.
Discussion
Essential oils and their components have been
investigated regarding their effect against a variety of human
cancer cell lines. Lemon balm was verified to show cytotoxic
activity against some human cancer cell lines and a mouse cell line
(14). The genus Eucalyptus is
native to Australia and this plant is mainly cultivated for paper,
pharmaceutical and cosmetics industries. Several species of
Eucalyptus are also used in traditional medicine as antiseptics and
against infections of the upper respiratory tract such as the
common cold, influenza and sinus congestion (15,16).
Essential oils from this species have a therapeutic application in
the treatment of pulmonary infections by inhalation (17) and the monoterpene extracted from
Eucalyptus species exhibit antibacterial activity (18). Ashour (19) showed cytotoxic activities of
volatile oils and extracts from Eucalyptus species against the
human breast adenocarcinoma cell line MCF7.
Among the various constituents of essential oils
extracted from Eucalyptus species, 1, 8-cineole is one of the main
constituent in the essential oils from Eucalyptus species (6,7). 1,
8-cineole has been shown to possess several pharmacological
effects. The cytotoxic activities against tumor cell lines were
previously only reported in leukemia cell lines (12). In this study, we revealed the
cytotoxic activities of 1, 8-cineole against human colorectal
cancer for the first time.
The mechanism of cell killing by 1, 8-cineole is not
fully understood. Suppression of growth by 1, 8-cineole in the
leukemia cell lines was reported to the induction of apoptosis
(12). Survivin is a type of
inhibitor of apoptosis (IAP) protein and helps repress apoptosis.
IAPs are highly expressed in most malignancies and play key roles
in the survival of neoplastic cells by negatively regulating
apoptosis (20). Several studies
show that high survivin expression in tumors is associated with
lower apoptosis index, increased resistance to chemotherapeutic
agents and poor patient survival (21). Specific inhibition of survivin
expression in multiple cancer cell lines by a survivin antisense
oligonucleotide induced caspase-3-dependent apoptosis, cell cycle
arrest in the G2-M phase and multinucleated cells
(21). Inhibition of survivin
expression using 1, 8-cineole mediates the apoptosis and growth
arrest in human colorectal cancers. p38 MAP kinase is a member of
the MAP kinase family and is activated by a variety of cellular
stresses (22–26). Activated p38 MAP kinase appears to
have multiple targets in the apoptotic pathway. Activated p38
mediates apoptosis by the downregulation of survivin in colon
cancer cells (27,28). The phosphatidylinositol-3-kinase
(PI3K)/Akt pathway is a cell signaling pathway and signaling
through the PI3K/Akt signaling axis promotes growth and survival
and undergoes apoptosis (29). The
overexpression of PI3K/Akt pathway was observed in many human
cancers (30). Our results show
that human CRC cell line RKO contains constitutively active
PI3K/Akt pathway. RKO expressed phosphoserine 473-Akt
constitutively and treatment with 1, 8-cineole dephosphorylated
Akt. 1, 8-cineole treatment activated p38 and dephosphorylated Akt,
which induced caspase-3 cleavage and resultant cleavage of PARP and
finally caused apoptosis.
In a xenotransplant mouse model, 1, 8-cineole
therapy showed tumor shrinkage comparable to the control group.
Throughout the experiments, the animals had no weight loss in 1,
8-cineole and control groups (data not shown). Moreover, we could
not observe any skin damage in the 1, 8-cineole group, which was
injected locally.
In conclusion, these findings demonstrate that 1,
8-cineole might exert its antitumor activity by triggering
apoptosis in human colorectal cancer cells in vitro and
in vivo. 1, 8-cineole shows promise as a strong and safe
chemotherapeutic agent for colorectal cancer.
Acknowledgements
The authors thank Satoko Nakabayashi for technical
assistance. This study was supported in part by grants-in-aid from
the Ministry of Education, Culture, Sports, Science and Technology
of Japan (MEXT).
References
|
1
|
Pezzuto JM: Plant-derived anticancer
agents. Biochem Pharmacol. 53:121–133. 1997. View Article : Google Scholar
|
|
2
|
Kinghorn AD, Farnsworth NR, Doel Soejarto
D, et al: Novel strategies for the discovery of plant-derived
anticancer agents. Pure Appl Chem. 71:1611–1618. 1999. View Article : Google Scholar
|
|
3
|
Lee KH: Anticancer drug design based on
plant-derived natural products. J Biomed Sci. 6:236–250.
1999.PubMed/NCBI
|
|
4
|
Moalic S, Liagre B, Corbiére C, et al: A
plant steroid, diosgenin, induces apoptosis, cell cycle arrest and
COX activity in osteosarcoma cells. FEBS Lett. 506:225–230. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan S, Cherney B, Reinhold W, et al:
Disruption of p53 function in immortalized human cells does not
affect survival or apoptosis after taxol or vincristine treatment.
Clin Can Res. 4:1047–1054. 1998.PubMed/NCBI
|
|
6
|
Giamakis A, Kretsi O, Chinou IB, et al:
Eucalyptus camaldulensis: volatiles from immature flowers
and high production of 1,8-cineole and b-pinene by in vitro
cultures. Phytochemistry. 58:351–355. 2001. View Article : Google Scholar
|
|
7
|
Tsiri DO, Kretsi O, Chinou IB, et al:
Composition of fruit volatiles and annual changes in the volatiles
of leaves of Eucalyptus camandulensis Dehn. Growing in
Greece. Flav Fragr J. 18:244–247. 2003. View Article : Google Scholar
|
|
8
|
Levinson KK, Takayama K, Okabe K, et al:
Formulation optimization of indomethacine gels containing a
combination of three kinds of cyclic monoterpenes as percutaneous
penetration enhancers. J Pharm Sci. 83:1367–1372. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Juergens UR, Stober M, Schmidt-Schilling
L, et al: Anti-inflammatory effects of encalyptol (1,8-cineole) in
bronchial asthma:inhibition of arachidonic acid metabolism in human
blood monocytes ex vivo. Eur J Med Res. 17:407–412. 1998.PubMed/NCBI
|
|
10
|
Juergens UR, Stober M and Vetter H:
Inhibition of cytokine production and arachidonic acid metabolism
by eucalyptol (1,8-cineole) in human blood monocytes in vitro. Eur
J Med Res. 17:508–510. 1998.PubMed/NCBI
|
|
11
|
Lahlou S, Figueierdo AF, Magalhaes PJ, et
al: Cardiovascular effects of 1,8-cineole, a terpenoid oxide
present in many plant essential oils, in normotensive rats. Can J
Physiol Pharmacol. 80:1125–1131. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Moteki H, Hibasami H, Yamada Y, et al:
Specific induction of apoptosis by 1,8-cineole in two human
leukemia cell lines, but not a in human stomach cancer cell line.
Oncol Rep. 9:757–760. 2002.PubMed/NCBI
|
|
13
|
Korzeniewski C and Callewaert DM: An
enzyme-release assay for natural cytotoxicity. J Immunol Methods.
64:313–320. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
De Sousa AC, Alviano DS, Blank AF, et al:
Melissa officinalis L. Essential oil: antitumoral and
antioxidant activities. J Pharm Pharmacol. 56:677–681. 2004.
|
|
15
|
Silva J, Abebe W, Sousa SM, et al:
Analgestic and anti-inflammatory effects of essential oils of
Eucalyptus. J Ethnopharmacol. 89:277–283. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Williams LR, Stockley JK, Yan W, et al:
Essential oils with high antimicrobial activity for therapeutic
use. Int J Aromatherapy. 8:30–40. 1998. View Article : Google Scholar
|
|
17
|
Low D, Rawal BD and Griffin WJ:
Antibacterial action of the essential oils of some Australian
Myrtaceae with special references to the activity of
chromatographic fractions of oil of Eucalyptus citriodora.
Planta Med. 26:184–185. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ramezani H, Singh HP, Batish DR, et al:
Antifungal activity of the volatile oil of Eucalyptus
citriodora. Fitoterapia. 73:261–262. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ashour HM: Antibacterial, antifungal and
anticancer activities of volatile oils and extracts from stems,
leaves and flowers of Eucalyptus sideroxylon and
Eucalyptus torquata. Cancer Biol Ther. 7:399–403. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Altieri DC: Validating survivin as a
cancer therapeutic target. Nat Rev Cancer. 3:46–54. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Carrasco RA, Stamm NB, Marcusson E, et al:
Antisense inhibition of survivin expression as a cancer
therapeutic. Mol Cancer Ther. 10:221–232. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rouse J, Cohen P, Trigon S, et al: A novel
kinase cascade triggered by stress and heat shock that stimulates
MAPKAP kinase-2 and phosphorylation of the small heat shock
proteins. Cell. 78:1027–1037. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Han J, Lee JD, Bibbs L, et al: A MAP
kinase targeted by endotoxin and hyperosmolarity in mammalian
cells. Science. 265:808–811. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee JC, Laydon JT, McDonnell PC, et al: A
protein kinase involved in the regulation of inflammatory cytokine
biosynthesis. Nature. 372:739–746. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Freshney NW, Rawlinson L, Guesdon F, et
al: Interleukin-1 activates a novel protein kinase cascade that
results in the phosphorylation of Hsp27. Cell. 78:1039–1049. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Raingeaud J, Gupta S, Rogers JS, et al:
Pro-inflammatory cytokines and environmental stress cause p38
mitogen-activated protein kinase activation by dual phosphorylation
on tyrosine and threonine. J Biol Chem. 270:7420–7426. 1995.
View Article : Google Scholar
|
|
27
|
Hsiao PW, Chang CC, Liu HF, et al:
Activation of p38 mitogen-activated protein kinase by celecoxib
oppositely regulates survivin and gamma-H2AX in human colorectal
cancer cells. Toxicol Appl Pharmacol. 222:97–104. 2007. View Article : Google Scholar
|
|
28
|
Hsu YF, Sheu JR, Lin CH, et al:
Trichostatin A and sirtinol suppressed survivin expression through
AMPK and p38 MAPK in HT29 colon cancer cells. Biochem Biophys Acta.
1820:104–115. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Toker A and Yoeli-Lerner M: Akt signaling
and cancer: surviving but not moving on. Cancer Res. 66:3963–3966.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Altomare DA and Testa JR: Perturbations of
the AKT signaling pathway in human cancer. Oncogene. 24:7455–7464.
2005. View Article : Google Scholar : PubMed/NCBI
|