Introduction
Thyroid cancer is the most common malignancy of the
endocrine system and accounts for ~1% of all malignant tumors
(1). In 2013, an estimated 60,220
Americans may be diagnosed with thyroid cancer, and the disease is
likely to account for ~1,850 mortalities (2). Papillary thyroid cancer (PTC) is the
most common subtype of this disease, accounting for >83% of all
such malignancies (3,4). Although the majority of PTC is
effectively managed by surgical resection followed by radioactive
iodine therapy, a subset of PTC is aggressive and refractory to
current therapeutic approaches. Doxorubicin (DOX), a broad-spectrum
anthracyclin, is the only FDA-approved chemotherapeutic agent for
patients with non-radioiodine avid thyroid cancer (5). However, the clinical impact of DOX on
thyroid cancer remains poor due to a high degree of chemoresistance
(5,6). In particular, in heart tissues, DOX
causes dilated cardiomyopathy, congestive heart failure and even
cardiac sudden death (7,8), which limits its clinical applications.
Therefore, improved therapeutic regimens that potentiate DOX
effects, allowing dose reduction are required to improve the
treatment of patients with the aggressive subtype of papillary
thyroid cancer.
NIN1/RPN12 binding protein 1 homolog (NOB1)
encodes a chaperone protein that joins the 20S proteasome with the
19S regulatory particle in the nucleus and is required for the
biogenesis and function of the 26S proteasome, which plays a role
in maintaining cellular homeostasis by controlling protein
degradation. The human ortholog of the NOB1 gene was
previously cloned (9). The
NOB1 gene, located on chromosome 16q22.1, encoding a 50-kDa
protein consisting of a PilT amino terminus (PIN) domain and a zinc
ribbon domain, is mainly expressed in the liver, lung and spleen
(9,10). NOB1 has been found to be
upregulated in various types of cancer, including ovarian cancer,
prostate and breast-infiltrating ductal carcinoma and non-small
cell lung cancer (11–14). In addition, the downregulation of
endogenous NOB1 inhibited tumor cell proliferation and colony
formation, and induced cell apoptosis and arrest in the G0/G1 stage
in various types of cancer (11,14–16).
For PTC, results of a previous study showed that the expression
level of NOB1 protein was increased in PTC tissue compared to
normal thyroid tissue and benign thyroid tumor tissue and its
expression was associated with UICC stage, tumor size and lymph
node metastasis (17). Findings of
a recent study demonstrated that adenovirus-mediated siRNA
targeting NOB1 (Ad/sh-NOB1) in human PTC cells inhibited
proliferation, migration and invasion in vitro, and
suppressed tumor growth in a mouse model, and enhanced the in
vitro and in vivo radiosensitivity of PTC cells
(18). However, to the best of our
knowledge, no previous studies have elucidated the role of NOB1 in
thyroid cancer chemoresistance.
The aim of the present study was to evaluate the
potency of Ad/sh-NOB1 in combination with DOX in inhibiting PTC
cell growth, proliferation and migration and invasion, and inducing
cell apoptosis and cell cycle arrest. In addition, tumor growth
ability in nude mice was detected to determine the combination
treatment effect on tumorigenesis in a nude mouse model.
Materials and methods
Reagents and antibody
Adenovirus expression vector carrying NOB1 siRNA
(Ad/sh-NOB1) and adenovirus expression vector carrying scramble
siRNA (Ad/sh-Scramble) were constructed and stored, as previously
described (18). Doxorubicin (DOX)
was obtained from Pfizer, Inc. (New York, NY, USA). Nonidet P-40
lysis buffer, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) was purchased from Sigma-Aldrich, St. Louis, MO, USA.
Stock solutions of propidium iodide (PI) and MTT were prepared by
dissolving 1 mg of each compound in 1 ml of phosphate-buffered
saline (PBS). The solution was protected from light, stored at 4°C
and used within one month.
For the western blot analysis, the antibodies used
were: mouse monoclonal anti-human NOB1 (Abcam, Cambridge, UK),
mouse monoclonal anti-human phosphorylated(p-) p38MAPK and mouse
monoclonal anti-human cyclin D1 (Cell Signaling Technology,
Danvers, MA, USA), mouse monoclonal anti-human p38MAPK, mouse
monoclonal anti-human p-ERK1/2, mouse monoclonal anti-human ERK1/2,
mouse monoclonal anti-human JNK, mouse monoclonal anti-human p-JNK,
mouse monoclonal anti-human P21, mouse monoclonal anti-human cyclin
D3 and mouse monoclonal anti-human GAPDH (Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA). Secondary antibodies HRP-conjugated
goat anti-mouse IgG was purchased from Amersham Biosciences
(Uppsala, Sweden).
Cell culture
The TPC-1 human PTC cell line, provided by the
Chinese Cell Bank of the Chinese Academy of Sciences (Shanghai,
China), was cultured in RPMI-1640 containing 10% fetal bovine serum
(FBS) (both from Gibco, Carlsbad, CA, USA), 100 U/ml penicillin and
100 mg/ml streptomycin (both from Sigma-Aldrich) at 37°C in a
humidified atmosphere of 5% CO2.
In vitro assay of chemosensitivity to
DOX
TPC-1 cells infected with Ad/sh-NOB1 or
Ad/sh-Scramble (MOI of 50 each), along with untreated cells were
seeded in a 96-well plates at a density of 5×103
cells/well and were cultured for 24 h. DOX was freshly prepared and
added to cells with final concentration series from 0 to 10,000 nM.
After incubation for 72 h, cell viability was determined by MTT
assay as described below. The IC50 values were
calculated from three independent experiments.
Proliferation assays
To measure the effect of Ad/sh-NOB1 combined with
DOX or as monotherapy on cell proliferation, MTT assay was
performed. Briefly, TPC-1 cells grown in monolayers were collected
and dispensed in 96-well culture plates in 100 μl of DMEM at a
concentration of 5×103 cells/well. After being cultured
for 24 h, the cells were treated with Ad/sh-NOB1 (MOI of 50 each),
Ad/sh-Scramble (MOI of 50 each), DOX (2X IC50) or
Ad/sh-NOB1 (MOI of 50 each) in combination with DOX (1X
IC50), respectively. After 48 h, the cells were washed
with PBS (pH 7.4) and incubated in 50 μl of 0.5 mg/ml MTT in
culture medium at 37°C for 4 h. Then 150 μl dimethyl sulfoxide
(DMSO) was added to dissolve the crystals. After 10 min at room
temperature, the absorbance was recorded at 570 nm in an ELISA
plate reader (Molecular Devices Corporation, Sunnyvale, CA, USA).
The mean proliferation of cells without any treatment was expressed
as 100%.
Colony formation assay
TPC-1 cells were seeded in 6-well culture plates at
1×104 cells/well. After being cultured for 24 h, the
cells were treated with Ad/sh-NOB1 (MOI of 50 each), Ad/sh-Scramble
(MOI of 50 each), DOX (2X IC50) or Ad/sh-NOB1 (MOI of 50
each) in combination with DOX (1X IC50), respectively.
After 14 days, the cells were washed, fixed in paraformaldehyde,
and stained with Giemsa for 10 min. Extra Giemsa was washed three
times by ddH2O, and the colonies were photographed using
a digital camera. The visible colonies in each group were
counted.
Cell cycle analysis
TPC-1 cells (5×105) were plated in 60-mm
dishes and treated with Ad/sh-NOB1 (MOI of 50 each), Ad/sh-Scramble
(MOI of 50 each), DOX (2X IC50) or Ad/sh-NOB1 (MOI of 50
each) in combination with DOX (1X IC50), respectively.
Forty-eight hours after treatment, the cells were collected by
trypsinization, fixed in 70% ethanol, and kept at −20°C overnight
for fixation. The cells were washed in PBS, resuspended in 1 ml of
PBS containing 100 μg/ml RNase and 40 μg/ml PI and incubated in the
dark at room temperature for 30 min. Cell distribution in the cell
cycle phases was analyzed from the DNA histogram with a FACSCalibur
flow cytometer and CellQuest software (both from Becton-Dickinson,
San Jose, CA, USA).
Cell apoptosis
Cell apoptosis was detected using flow cytometry. In
briefly, TPC-1 cells were treated with Ad/sh-NOB1 (MOI of 50 each),
Ad/sh-Scramble (MOI of 50 each), DOX (2X IC50) or
Ad/sh-NOB1 (MOI of 50 each) in combination with DOX (1X
IC50), respectively. After 48 h, 1×106 cells
were digested with 10 μg/ml RNase for 30 min at 37°C. Annexin
V-fluorescein isothiocyanate (0.5 μg/ml) and PI (0.6 μg/ml) were
added to a 250 μl aliquot of this cell suspension. After a 15-min
incubation in the dark at room temperature, the sample was read on
a Coulter EPICS XL flow cytometer (Beckman Coulter, Inc.,
Fullerton, CA, USA), and the data were analyzed using CellQuest
software (BD Biosciences, San Jose, CA, USA). Experiments were
performed in triplicate. In addition, caspase-3, −8 and −9 activity
were detected as an additional indicator of apoptosis.
Caspase activity
Caspase-3, −8 and −9 activity was measured using
caspase colorimetric protease assay kits (Millipore Corporation,
Billerica, MA, USA) according to the manufacturer’s
instructions.
Transwell migration and invasion
assays
To assess the effect of DOX and Ad/sh-NOB1 on cell
migration and invasion, the migration and invasion assays were
performed using Transwell insert chambers (Corning, Costar,
Cambridge, MA, USA). For the migration assay, TPC-1 cells were
treated with Ad/sh-NOB1 (MOI of 50 each), Ad/sh-Scramble (MOI of 50
each), DOX (2X IC50) or Ad/sh-NOB1 (MOI of 50 each) in
combination with DOX (1X IC50), respectively.
Forty-eight hours post-treatment, 1×105 cells were
plated into the upper chamber in serum-free RPMI-1640 medium.
Medium containing 20% FBS in the lower chamber served as the
chemoattractant. After being cultured for 24 h, the medium was
removed from the upper chamber by wiping with a cotton swab and
cells that migrated to the lower surface of filter were fixed in
70% ethanol for 30 min, followed by staining with 0.2% crystal
violet for 10 min. Cell migration was determined by counting five
random fields/filter under a light microscope (Olympus, Tokyo,
Japan).
For the invasion assay, after treatment,
3×105 cells were seeded in upper chambers precoated with
Matrigel (BD Biosciences, Bedford, MA USA) in serum-free medium in
triplicate, and the subsequent steps were similar to those of the
migration assay. The number of cells invading the Matrigel was
counted in five randomly selected fields using an inverted
microscope (Olympus). All the experiments were performed in
triplicate.
Measurement of matrix metalloproteinase-9
(MMP-9) and vascular endothelial growth factor (VEGF) level
MMP-9 and VEGF levels were determined by ELISA.
Briefly, TPC-1 cells were treated with Ad/sh-NOB1 (MOI of 50 each),
Ad/sh-Scramble (MOI of 50 each), DOX (2X IC50) or
Ad/sh-NOB1 (MOI of 50 each) in combination with DOX (1X
IC50) for 48 h in 24-well plates, and the culture media
were centrifuged to remove cell debris. PGE2 levels in the cell
supernatant were then measured by human MMP-9 ELISA kits (R&D
Systems, Shanghai, China) according to the manufacturer’s
instructions. VEGF levels in the cell supernatant were determined
by a human VEGF ELISA kit (Yanyu, Shanghai, China) according to the
manufacturer’s instructions.
Western blotting
Proteins were extracted from TPC-1 cells, and were
characterized using western blot analysis. Briefly, the cells were
lysed in lysis buffer. The homogenates were then centrifuged at
14,000 rpm at 4°C for 30 min to remove insoluble material, and the
supernatants were collected for protein concentration determination
using the BCA assay kit (Sigma-Aldrich). Cell extracts (20 μg of
protein) were separated on a sodium dodecyl sulfate-polyacrylamide
electrophoretic gel (SDS-PAGE) and transferred to nitrocellulose
membranes, which were blocked in 3% bovine serum albumin (BSA) in
PBST for 1 h at room temperature. The membrane was probed with
primary antibody overnight at 4°C. After extensive washing, the
membrane was incubated with secondary antibody for 1 h at room
temperature. The proteins detected by the enhanced protein bands
were visualized with enhanced chemiluminescent reagent (ECL;
Sigma-Aldrich). The integrated densities value (IDV) was analyzed
with computerized image analysis system (Fluor Chen 2.0) and
normalized with that of GAPDH.
Tumor xenograft in nude mice
Female BALB/c nude mice at 6–7 weeks old were
obtained from the Experimental Animal Center of the Jilin
University (Changchun, China) and maintained under specific
pathogen-free conditions and provided with food and water ad
libitum.
Exponentially growing TPC-1 cells were collected and
a tumorigenic dose of 2×106 cells was injected
intraperitoneally into 4- to 5-week-old female BALB mice. The tumor
volume was calculated using the formula: Volume = (length ×
width2)/2. When tumors grew to an average volume of 100
mm3, the mice were divided randomly into five groups
(n=10 mice/group). The control group received 1% PBS in deionized
water. The remaining four groups were treated with Ad/sh-Scramble
(1×108 PFU/dose), DOX (4 mg/kg body weight), Ad/sh-NOB1
(1×108 PFU/dose), or DOX plus Ad/sh-NOB1 (DOX, 2 mg/kg
body weight; Ad/sh-NOB1, 1×108 PFU/dose, respectively)
intraperitoneally on alternative days for 3 weeks. Tumor size was
measured using calipers prior to administration of the treatment
injections and on day 7, 14 and 21 of treatment. On day 21, the
animals were euthanized using chloroform, and tumor tissues were
isolated and weighed. In addition, spleen tissues of the mice were
collected and cultured for a splenocyte surveillance study, as
previously described (19). The
present animal study was performed following approval of the
protocol by the Jilin University Animal Care and Use Committee
(Changchun, Jilin, China)
Statistical analysis
Data from at least three independent experiments
were expressed as mean ± standard deviation (SD). One-way ANOVA
followed by Tukey’s post-hoc test were utilized to determine the
significant difference among multiple groups. Statistical analyses
were undertaken using GraphPad Prism version 5.01 (GraphPad
Software, San Diego, CA, USA) and the SPSS® statistical
package, version 19.0 (SPSS, Inc., Chicago, IL, USA) for
Windows®. P<0.05 was considered to indicate a
statistically significant result.
Results
Effects of DOX and Ad/sh-NOB1 alone or in
combination on cell proliferation and colony formation of TPC-1
cells
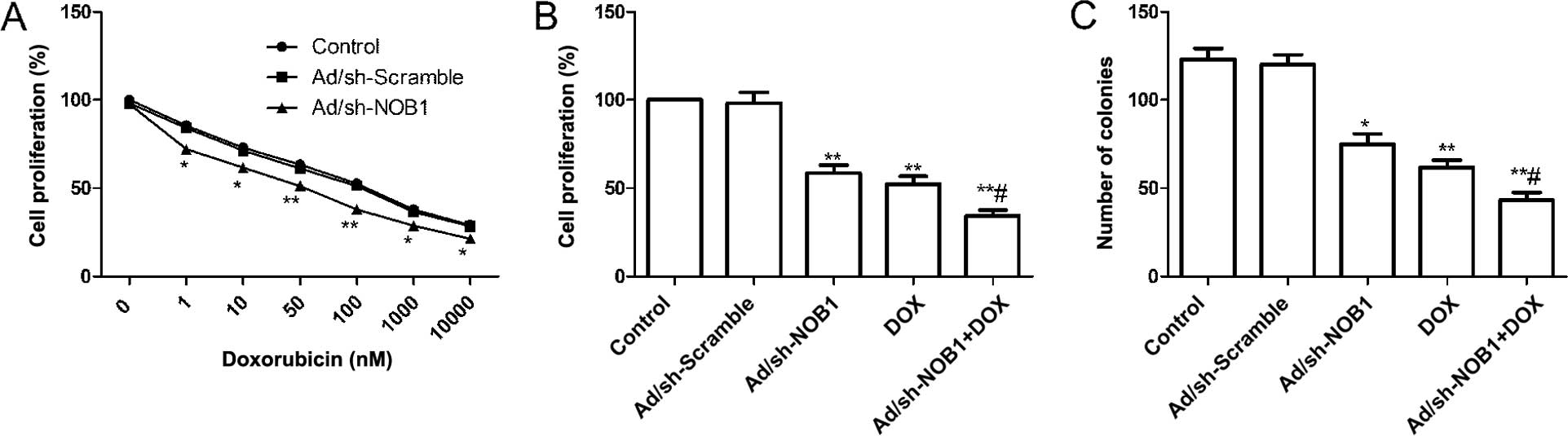
TPC-1 cells were infected with Ad/sh-NOB1 or
Ad/sh-Scramble, and then incubated with DOX (0–10,000 nM). MTT
assays were performed following treatment and the IC50
for DOX was calculated for each of the groups treated. It was found
that downregulation of NOB1 by Ad/sh-NOB1 resulted in a 50.56%
reduction of the DOX IC50 (75.97±6.12 nM) in TPC-1 cells
as compared to cells infected Ad/sh-Scramble (150.25±11.58 nM)
(Fig. 1A). Thus, a reduced NOB1
expression sensitized TPC-1 cells to DOX.
To evaluate the effect of DOX combined with
Ad/sh-NOB1 or independently on proliferation of PTC cells, an MTT
assay was performed 48 h after TPC-1 cells were treated with DOX
and Ad/sh-NOB1 alone or in combination. The results showed a
significant decrease in cell proliferation as compared to the
control and Ad/sh-Scramble treatment group (Fig. 1B, P<0.01). Compared to the
monotherapy group, DOX in combination with Ad/sh-NOB1 obviously
reduced cell proliferation (Fig.
1B, P<0.05).
The effects of DOX and Ad/sh-NOB1 alone or
combination on the TPC-1 cell colony formation cycles of cells were
then analyzed. As shown Fig. 1C,
colony number tumor cell have significantly reduction in DOX and
Ad/sh-NOB1 alone or combination groups compared with control and
Ad/sh-Scramble group, (P<0.05, Fig.
1C). The DOX in combination with Ad/sh-NOB1 resulted in a
greater reduction of colony number compared to either DOX or
Ad/sh-NOB1 (P<0.05, Fig.
1C).
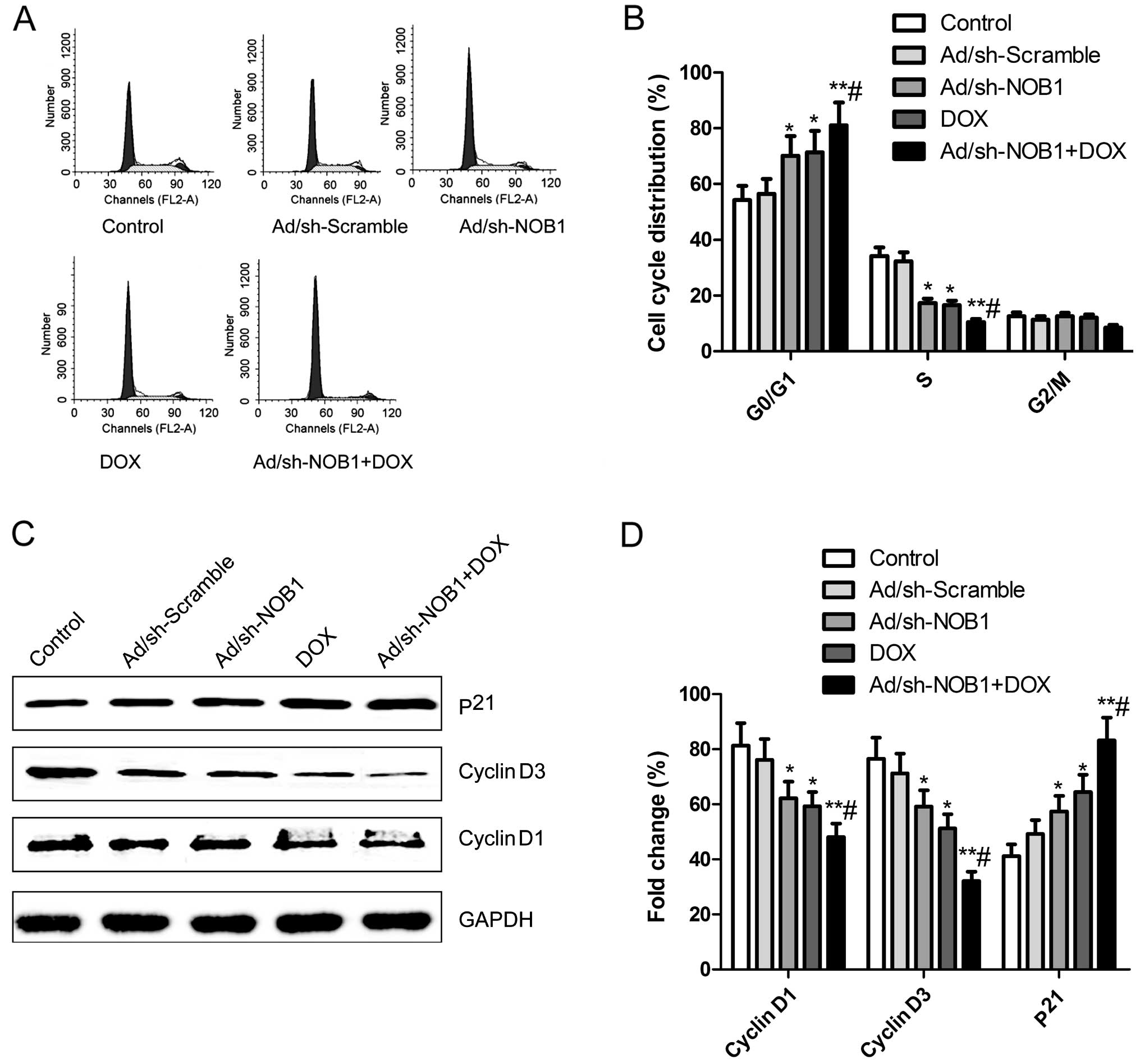
Effects of DOX and Ad/sh-NOB1 alone or in
combination on cell cycle of TPC-1 cells
The effects of DOX and Ad/sh-NOB1 alone or in
combination on the cell cycles of TPC-1 cells were analyzed using
flow cytometry. It was found that TPC-1 cells treated with DOX and
Ad/sh-NOB1 alone or in combination had an increased percentage of
arrest at the G0/G1 phase, and a decreased percentage of arrest at
the S stage compared to the control and Ad/sh-Scramble groups
(P<0.05, Fig. 2A and B). The DOX
combination with Ad/sh-NOB1 resulted in a greater effect at the
G0/G1 phase and S stage (Fig. 2A and
B, P<0.01).
We analyzed the effects of DOX and Ad/sh-NOB1 alone
or in combination on the expression of cell cycle relevant
proteins, such as cyclin D1 and D3 and p21 by western blotting. As
shown in Fig. 2C and D, p21
expression was significantly increased, while cyclin D1 and D3
expression was significantly decreased in the DOX and Ad/sh-NOB1
alone or combination group compared to the control and
Ad/sh-Scramble groups (P<0.05). Compared to the DOX or
Ad/sh-NOB1 groups, the combination treatment group showed a
synergistic effect on p21, cyclin D1 and D3 protein expression
(P<0.05, Fig. 2C and D).
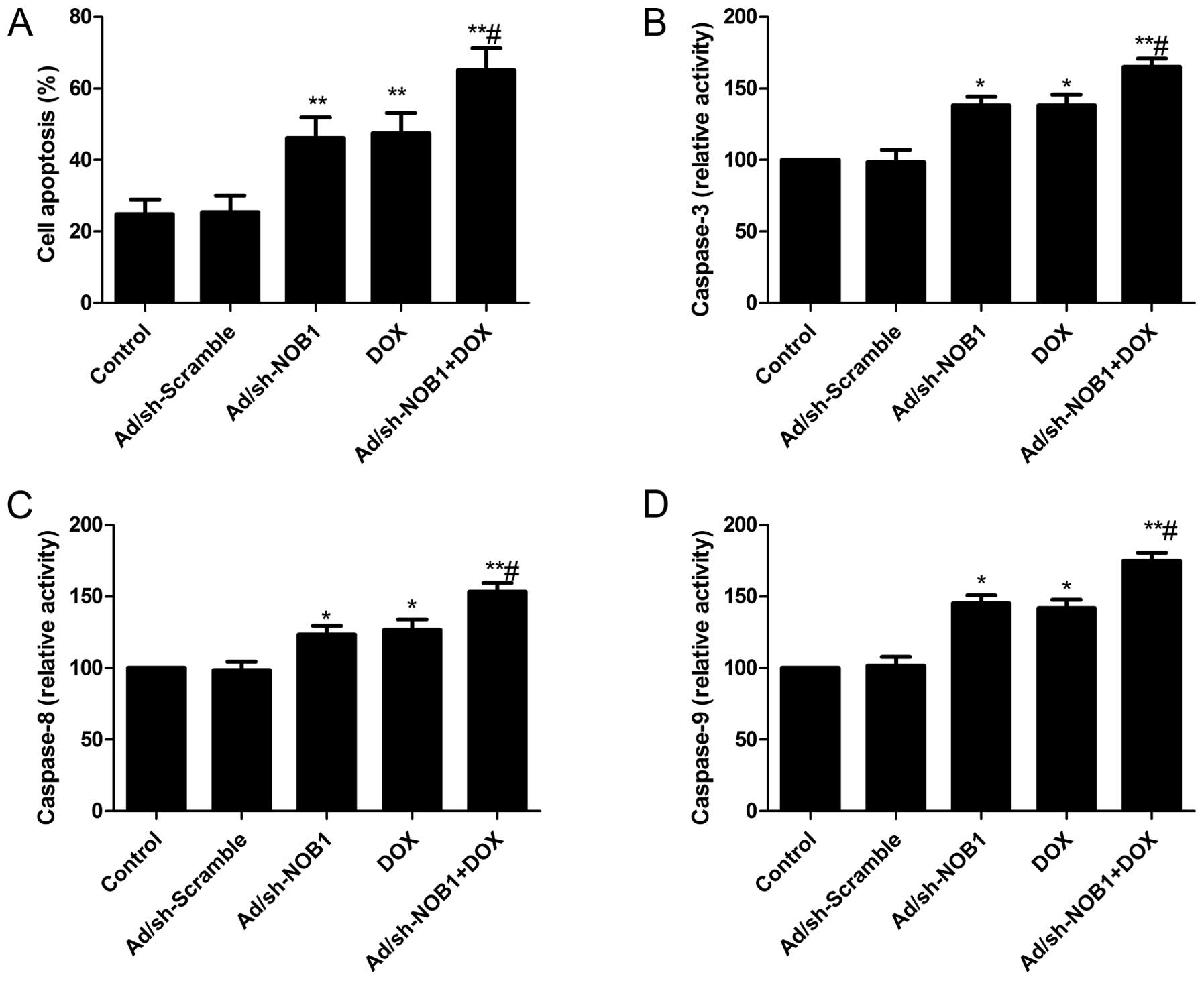
Effects of DOX and Ad/sh-NOB1 alone or in
combination on cell apoptosis of TPC-1 cells
To investigate whether DOX and Ad/sh-NOB1 alone or
in combination induced apoptosis, cell apoptosis was analyzed by
flow cytometry following treatment. It was found that TPC-1 cells
treated with DOX and Ad/sh-NOB1 alone or in combination
significantly induced cell apoptosis compared with the control and
Ad/sh-Scramble groups (P<0.05, Fig.
3A). Treatment with a combination of DOX and Ad/sh-NOB1 led to
a marked increase in apoptotic cells compared to single DOX or
Ad/sh-NOB1 treatment groups (P<0.05, Fig. 3A).
To examine the possible mechanism of induction of
cell apoptosis of the combination of DOX and Ad/sh-NOB1, caspase-3,
−8 and −9 activity was determined by ELISA. The results showed that
DOX and Ad/sh-NOB1 alone or combined significantly increased
caspase-3, −8 and −9 activity in TPC-1 cells, compared to the
control and Ad/sh-Scramble groups (P<0.05, Fig. 3B and D). Compared to DOX or
Ad/sh-NOB1 alone, caspase-3, −8 and −9 activity was significantly
increased in the combination treatment group (P<0.05, Fig. 3B and D).
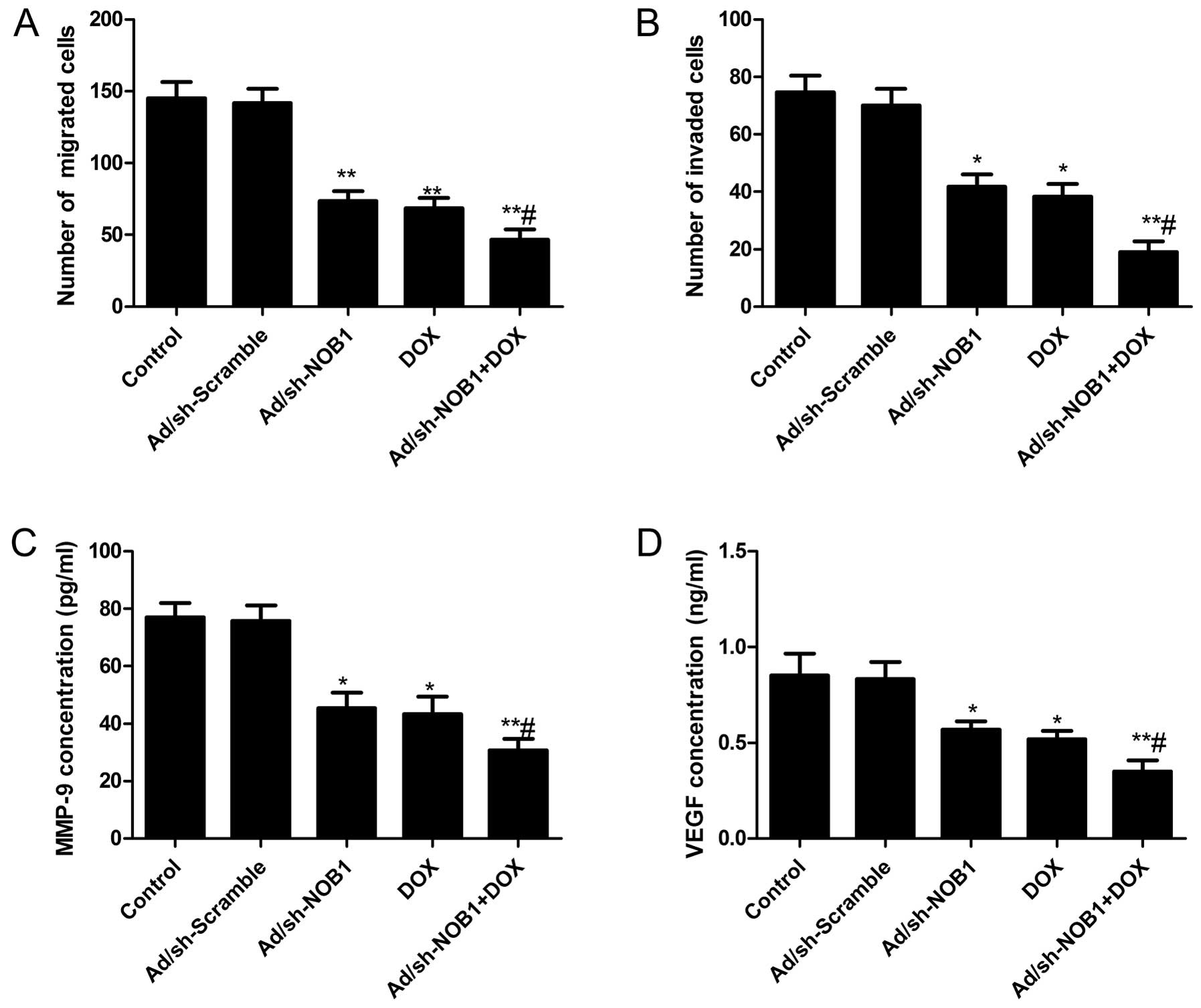
Effects of DOX and Ad/sh-NOB1 alone or in
combination on cell migration and invasion of TPC-1 cells
To determine the effect of DOX and Ad/sh-NOB1 alone
or as combined treatment on PTC cell migration and invasion, a
Transwell assay was performed. The Transwell assay (without
Matrigel) showed that the migratory speed of TPC-1 cells was
markedly slower in the DOX and Ad/sh-NOB1 alone or combined groups
than that of the control and Ad/sh-Scramble groups (Fig. 4A, P<0.05). Compared to the DOX
and Ad/sh-NOB1 group, migration was significantly reduced in the
combination group. Transwell matrix penetration (coated with
Matrigel) assay showed that DOX and Ad/sh-NOB1 alone or in
combination markedly reduced the invasiveness of TPC-1 cells as
compared to the control and Ad/sh-Scramble groups (Fig. 4B, P<0.05). DOX in combination
with Ad/sh-NOB1 resulted in the greatest reduction of invation of
TPC-1 cells (Fig. 4B,
P<0.05).
We also analyzed the effects of DOX and Ad/sh-NOB1
alone or in combination on the level of MMP-9 and VEGF by ELISA. It
was found that MMP-9 and VEGF were significantly decreased in the
DOX and Ad/sh-NOB1 alone or combinatorial treatment group compared
to the control and Ad/sh-Scramble groups (P<0.05, Fig. 4C and D). The combinatorial treatment
group showed synergistic inhibition of the MMP-9 and VEGF
level.
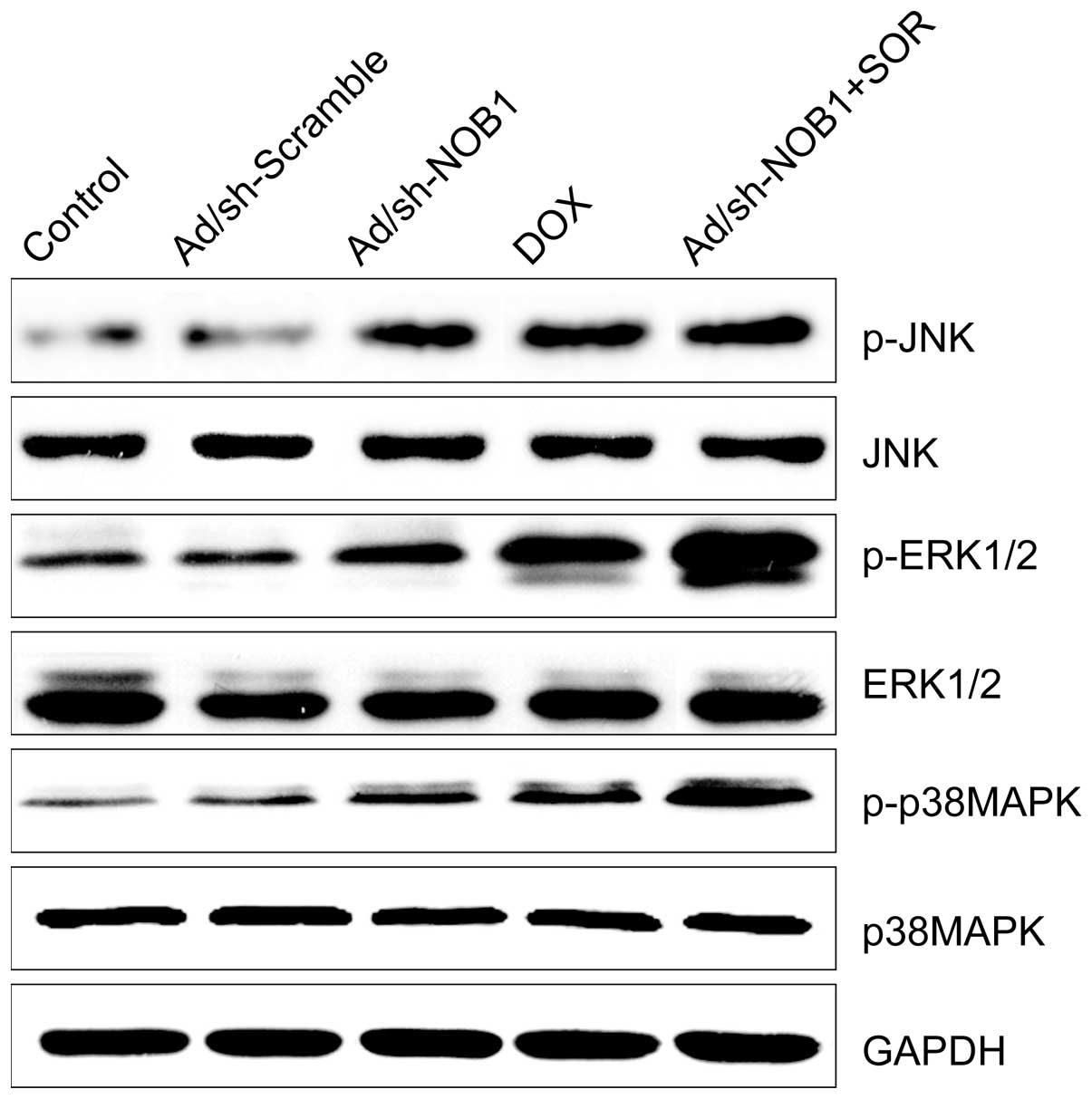
Effects of DOX and Ad/sh-NOB1 alone or in
combination on the p38MAPK pathway
It was previously shown that DOX stimulated the
p38MAPK pathway and induced cell apoptosis (20). Therefore, we examined whether this
mediated signaling pathway was enhanced by DOX in combination with
AD/sh-NOB1. Measurements of the phosphorylation/activation pattern
of p38MAPK, ERK1/2 and JNK were performed by western blotting 12 h
after treatment. It was shown that DOX and Ad/sh-NOB1 alone or in
combination resulted in a marked addition of phosphorylated
p38MAPK, ERK1/2 and JNK as compared to the control and
Ad/sh-Scramble groups, without altering the total protein levels of
p38MAPK, ERK1/2 and JNK in each group (Fig. 5). Compared to the monotherapy group,
the DOX and Ad/sh-NOB1 combinatorial treatment group increased
phosphorylated p38MAPK, ERK1/2 and JNK protein expression,
demonstrating a synergistic effect.
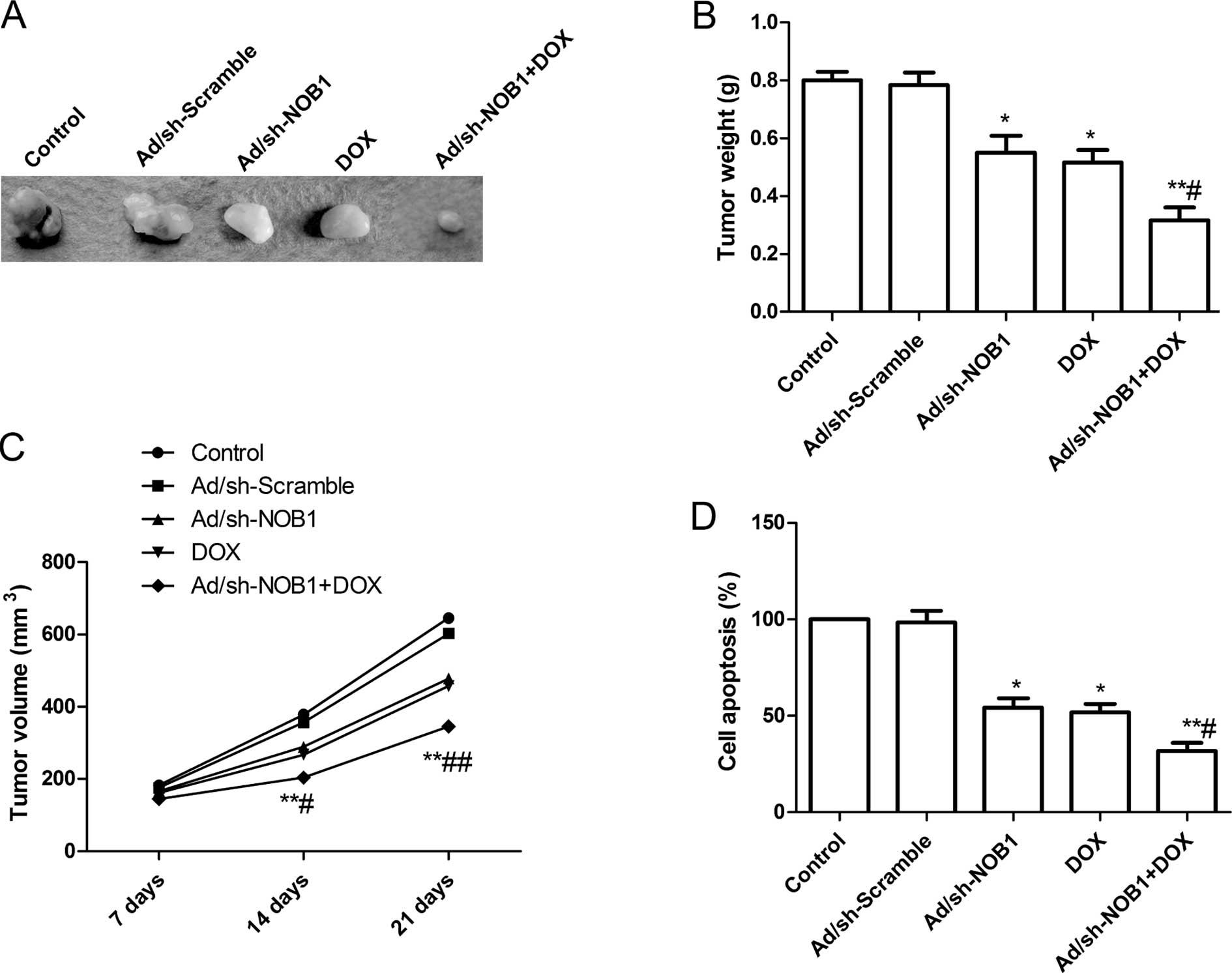
Effects of DOX and Ad/sh-NOB1 alone or in
combination on tumor growth in vivo
We assessed the in vivo therapeutic efficacy
of DOX and Ad/sh-NOB1 alone or in combination on female BALB mice
bearing TPC-1 tumor cells. Tumor growth was monitored for 21 days.
On day 21, the mice were anaesthetized and decapitated, and then
tumors were excised, weighed and measured. The tumor weight was
significantly reduced in the DOX and Ad/sh-NOB1 alone or
combinatorial groups compared to that of the control (PBS group)
and Ad/sh-Scramble groups (P<0.05; Fig. 6A and B). Compared to other groups,
the DOX in combination with Ad/sh-NOB1 group showed maximally
reduced weight. The tumor volume was determined at different time
points (7, 14 and 21 day). The tumor volume in the DOX and
Ad/sh-NOB1 alone or combinatorial groups were significantly
(P<0.05) reduced compared to the Ad/sh-Scramble and control
groups (PBS group) at the different time points (P<0.05,
Fig. 6C). DOX in combination with
Ad/sh-NOB1 led to a significant inhibition of tumor volume compared
with other treatment groups (P<0.05, Fig. 6C).
In addition, we employed MTT assays to modulate
splenocyte proliferation to demonstrate the antitumor activities.
We found that splenocyte cell proliferation in the DOX and
Ad/sh-NOB1 alone or combinatorial groups were significantly
decreased as compared to the control and Ad/sh-Scramble groups
(P<0.05, Fig. 6D). The
combination treatment led to a significant inhibition of splenocyte
cell proliferation compared with other treatment groups (P<0.05,
Fig. 6D). These results suggested
that the most prominent tumor decrease occurred in the DOX in
combination with Ad/sh-Scramble group, demonstrating a synergistic
effect in vivo.
Discussion
It is well known that drug resistance is a major
hindrance encountered in cancer chemotherapy. During tumorigenesis
or the course of chemotherapy, alteration of the gene expression
may occur and influence cell sensitivity to chemotherapeutic drugs
(21). In the present study, we
found that the downregulation of NOB1 expression by Ad/sh-NOB1
sensitized TPC-1 cells to doxorubicin (DOX). In addition, our in
vitro studies demonstrated that Ad/sh-NOB1 in combination with
a reduced dose of DOX significantly suppressed cell proliferation,
migration and invasion, and induced cell apoptosis and cell cycle
arrest at G0/G1 stage compared to an increased dose of DOX.
Additionally, further studies with in vivo mouse models
confirmed that this combination significantly the suppressed tumor
growth of PTC compared to a high dose of DOX. Therefore,
combination of Ad/sh-NOB1 and DOX may serve as a novel therapeutic
strategy for further evaluation in clinical trials for the
treatment of PTC.
NOB1 is an important component of 26S proteasome,
and favors 26S proteasome biogenesis (22). The 26S proteasome involved in the
ubiquitin-proteasome pathway (UPP), required for the degradation of
cyclic proteins and regulates multiple aspects of the cell cycle
progression in eukaryotes (23,24).
Previous studies have demonstrated that inhibition of 26S
proteasomes is an effective anticancer therapeutic approach in
vitro and in vivo. For example, bortezomib, one of the
first proteasome inhibitors, which was designed to inhibit the
activity of the 26S proteasome by binding to the N-terminal
threonine residues at the active site of the catalytic region
(25), has been shown to act as an
effective drug against various human cancers and solid tumor types
(25–27). Therefore, NOB1, a key factor in the
UPP and proteasome complex, has received attention, and has been
found to play role in tumor development and progression (11–18).
In a recent study (18), we showed
that blockade of NOB1 using Ad/sh-NOB1 via direct intratumoral
injections significantly reduced PTC tumor growth in nude mice.
However, complete suppression of tumor growth was not observed. RNA
interference does not completely block gene expression,
particularly when the target mRNA is expressed at abnormally high
levels (28). Findings of our
recent study showed that Ad/sh-NOB1 combined with irradiation
treatment led to a significant inhibition of tumor growth in nude
mice compated to Ad/sh-NOB1 treatment, which suggested that
Ad/sh-NOB1 combined with other drugs or irradiation may lead to a
stronger antitumor effect for human thyroid cancer. Therefore, in
the present study, we used Ad/sh-NOB1 combined with DOX for this
purpose. Our results show that the most prominent tumor decrease
occurred in DOX combined with the Ad/sh-Scramble group as compared
to Ad/sh-NOB1 and DOX as monotherapy, demonstrating a synergistic
effect in vivo. These findings together with those of our
previous study (17,18) have shown that the downregulation of
NOB1 expression by Ad/sh-NOB1 suppressed the tumor growth of PTC
in vitro and in vivo, and enhanced the radio- and
chemosensitivity of PTC cells.
The MAPK pathway plays a crucial role in signal
transduction and mediates cell proliferation, differentiation,
inflammation and apoptosis, which consists of extracellular
signal-regulated kinase (ERK), p38 and Jun N-terminal kinase (JNK)
(29). It has been demonstrated
that DOX suppressed tumor growth via activation of the MAPK pathway
(19). In particular, it has been
shown that knockdown of NOB1 expression inhibited tumor growth, to
some extent, by activating the MAPK pathway (30,31).
Our recently study also showed that adenovirus-mediated siRNA
targeting NOB1 could active MAPK pathway (18). Therefore, whether mediated MAPK
pathway was enhanced by DOX in combination with Ad/sh-NOB1 was
determined. In the present study, we found that Ad/sh-NOB1 in
combination with DOX treatment resulted in a marked increase of
phosphorylated f p38MAPK, ERK1/2 and JNK relative to DOX and
Ad/sh-NOB1 group, without altering the total protein levels of f
p38MAPK, ERK1/2 and JNK in each group. These findings suggest that
Ad/sh-NOB1 in combination with DOX demonstrated the synergistic
effect on inhibiting the tumor growth of PTC in vitro and
in vivo, at least in part through activation of the MAPK
pathway.
Taken together, the findings reported herein present
evidence that Ad/sh-NOB1 in combination with DOX significantly
inhibited cell proliferation, migration and invasion, induced cell
apoptosis in vitro, and suppressed tumor growth in
vivo, compared to either Ad/sh-NOB1 or DOX, demonstrating a
synergistic effect. In addition, this combination also activated
the p38MAPK signaling pathway, which contributes to inhibition of
tumor growth. These findings suggest that the combination of DOX
and Ad/sh-NOB1 is a promising drug candidate for the treatment of
PTC.
Acknowledgements
This study was supported by the Science and
Technology Research and Innovation Team funded of Jilin Provincial
(JL20130518).
References
|
1
|
Brown RL, de Souza JA and Cohen EE:
Thyroid cancer: burden of illness and management of disease. J
Cancer. 2:193–199. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pitoia F, Bueno F, Urciuoli C, Abelleira
E, Cross G and Tuttle RM: Outcomes of patients with differentiated
thyroid cancer risk-stratified according to the American thyroid
association and Latin American thyroid society risk of recurrence
classification systems. Thyroid. 23:1401–1407. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thompson L: World Health Organization
classification of tumours: pathology and genetics of head and neck
tumours. Ear Nose Throat J. 85:742006.PubMed/NCBI
|
|
4
|
Nikiforova MN and Nikiforov YE: Molecular
genetics of thyroid cancer: implications for diagnosis, treatment
and prognosis. Expert Rev Mol Diagn. 8:83–95. 2008. View Article : Google Scholar
|
|
5
|
Carvalho C, Santos RX, Cardoso S, et al:
Doxorubicin: the good, the bad and the ugly effect. Curr Med Chem.
16:3267–3285. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gottesman MM, Fojo T and Bates SE:
Multidrug resistance in cancer: role of ATP-dependent transporters.
Nat Rev Cancer. 2:48–58. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Silber JH and Barber G:
Doxorubicin-induced cardiotoxicity. N Engl J Med. 333:1359–1360.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
King PD and Perry MC: Hepatotoxicity of
chemotherapy. Oncologist. 6:162–176. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y, Ni J, Zhou G, et al: Cloning,
expression and characterization of the human NOB1 gene. Mol Biol
Rep. 32:185–189. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arcus VL, Bäckbro K, Roos A, Daniel EL and
Baker EN: Distant structural homology leads to the functional
characterization of an archaeal PIN domain as an exonuclease. J
Biol Chem. 279:16471–16478. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin Y, Peng S, Yu H, et al: RNAi-mediated
downregulation of NOB1 suppresses the growth and colony-formation
ability of human ovarian cancer cells. Med Oncol. 29:311–317. 2012.
View Article : Google Scholar
|
|
12
|
Che JP, Li W, Yan Y, et al: Expression and
clinical significance of the nin one binding protein and p38 MAPK
in prostate carcinoma. Int J Clin Exp Pathol. 6:2300–2311.
2013.PubMed/NCBI
|
|
13
|
Li XY, Luo QF, Li J, et al: Clinical
significance of NOB1 expression in breast infiltrating ductal
carcinoma. Int J Clin Exp Pathol. 6:2137–2144. 2013.PubMed/NCBI
|
|
14
|
Li Y, Ma C, Qian M, Wen Z, Jing H and Qian
D: Downregulation of NOB1 suppresses the proliferation and tumor
growth of non-small cell lung cancer in vitro and in vivo. Oncol
Rep. 31:1271–1276. 2014.PubMed/NCBI
|
|
15
|
Huang WY, Chen DH, Ning L and Wang LW:
siRNA mediated silencing of NIN1/RPN12 binding protein 1 homolog
inhibits proliferation and growth of breast cancer cells. Asian Pac
J Cancer Prev. 13:1823–1827. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lu Z, Guo Q, Shi A, et al: Downregulation
of NIN/RPN12 binding protein inhibit the growth of human
hepatocellular carcinoma cells. Mol Biol Rep. 39:501–507. 2012.
View Article : Google Scholar
|
|
17
|
Lin S, Meng W, Zhang W, et al: Expression
of the NOB1 gene and its clinical significance in papillary thyroid
carcinoma. J Int Med Res. 41:568–572. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Meng W, Wang PS, Liu J, et al:
Adenovirus-mediated siRNA targeting NOB1 inhibits tumor growth and
enhances radiosensitivity of human papillary thyroid carcinoma in
vitro and in vivo. Oncol Rep. 32:2411–2420. 2014.PubMed/NCBI
|
|
19
|
Abe S, Nishimoto Y, Isu K, Ishii T and
Goto T; Japanese Musculoskeletal Oncology Group. Preoperative
cisplatin for initial treatment of limb osteosarcoma: its local
effect and impact on prognosis. Cancer Chemother Pharmacol.
50:320–324. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao BX, Sun YB, Wang SQ, et al: Grape
seed procyanidin reversal of P-glycoprotein associated multi-drug
resistance via down-regulation of NF-κB and MAPK/ERK mediated YB-1
activity in A2780/T cells. PLoS One. 8:e710712013. View Article : Google Scholar
|
|
21
|
el-Deiry WS: Role of oncogenes in
resistance and killing by cancer therapeutic agents. Curr Opin
Oncol. 9:79–87. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tone Y and Toh-E A: Nob1p is required for
biogenesis of the 26S proteasome and degraded upon its maturation
in Saccharomyces cerevisiae. Genes Dev. 16:3142–3157. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tone Y, Tanahashi N, Tanaka K, Fujimuro M,
Yokosawa H and Toh-E A: Nob1p, a new essential protein, associates
with the 26S proteasome of growing Saccharomyces cerevisiae cells.
Gene. 243:37–45. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu G, Bernaudo S, Fu G, et al: Cyclin G2
is degraded through the ubiquitin-proteasome pathway and mediates
the antiproliferative effect of activin receptor-like kinase 7. Mol
Biol Cell. 19:4968–4979. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pandit B and Gartel AL: Thiazole
antibiotic thiostrepton synergize with bortezomib to induce
apoptosis in cancer cells. PLoS One. 6:e171102011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Piperdi B, Ling YH, Liebes L, Muggia F and
Perez-Soler R: Bortezomib: understanding the mechanism of action.
Mol Cancer Ther. 10:2029–2030. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dick LR and Fleming PE: Building on
bortezomib: second-generation proteasome inhibitors as anti-cancer
therapy. Drug Discov Today. 15:243–249. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Elbashir SM, Harborth J, Weber K and
Tuschl T: Analysis of gene function in somatic mammalian cells
using small interfering RNAs. Methods. 26:199–213. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dhillon AS, Hagan S, Rath O and Kolch W:
MAP kinase signalling pathways in cancer. Oncogene. 26:3279–3290.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou J, Xu T, Yan Y, et al: MicroRNA-326
functions as a tumor suppressor in glioma by targeting the Nin one
binding protein (NOB1). PLoS One. 8:e684692013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li W, Liu M, Feng Y, et al: Downregulated
miR-646 in clear cell renal carcinoma correlated with tumour
metastasis by targeting the nin one binding protein (NOB1). Br J
Cancer. 111:1188–1200. 2014. View Article : Google Scholar : PubMed/NCBI
|