1. Introduction
The detection of tumors at early stages allows
curative treatment to be administered before tumor progression
occurs (1); consequently, patients
live longer and fare better than those with advanced cancer
(2,3). The detection of such tumors is
challenging, which is particularly significant in the high-risk
population, in whom the incidence of disease is higher (1,4). Major
components of the early detection of cancer are education and
screening, but the most important aspect is the development of
early diagnostic tools. Tools that recognize the warning signs of
cancer so prompt action can be taken may ensure an early diagnosis,
and simple tests can identify individuals in a healthy population
who have the disease but have not developed symptoms (5). Diagnoses that are based on symptoms
are unacceptable for cancer, as they usually appear when the tumors
are sufficiently large in size (6).
An early diagnosis is paramount in breast cancer
(BC), since it is the most frequent tumor occurring in women in
industrialized and developing nations (2,6). Thus,
early detection remains the cornerstone of controlling BC to
improve patient outcomes and survival. Breast tumors express
aberrant levels of mutated or modified forms of proteins that are
associated with malignant growth. These proteins, called
tumor-associated antigens (TAAs) and tumor-associated carbohydrate
antigens (TACAs), are able to stimulate cellular and humoral immune
responses; TAAs are identified by serum antibodies (Abs) of
patients (7,8).
The only effective screening method for BC is
mammography. Mammography is expensive and is only cost-effective
and feasible in developed countries with good health
infrastructure. Many low- and middle-income nations must implement
low-cost screening, such as clinical breast examination and early
diagnostic tools (5,9). Ongoing studies are evaluating
inexpensive screening methods that can be implemented and sustained
in low-resource settings, based on the detection of antitumor
antigens by immunoglobulin M (IgM) Abs in the serum of female mice
with BC. In these studies, the patterns of antigen (Ag) recognition
by Abs in 2D immunoblots are identified and expressed as
immunological signatures, allowing certain patterns to be
correlated with resistance or susceptibility (10).
Two types of IgM exist: natural, which is present in
an organism without prior antigenic contact and is part of the
first-line defense; and adaptive, which develops after antigenic
challenge (11). Natural IgM also
has a significant function in maintaining tissue homeostasis,
promoting the phagocytic clearance of apoptotic cells and
preventing infectious and autoimmune diseases (12), and in recognizing and removing
precancerous and cancerous cells (13–18).
In the present review, we discuss the function of
natural IgM and adaptive IgM in eliminating cancer cells in the
early stages of BC and their potential as early diagnostic tools
and how, as components of an organism's defense, they can be used
to identify TAAs and TACAs.
2. IgM antibodies
IgM, which has μ heavy chains, is the first
class of antibody that is synthesized by and appears on the surface
of a developing B cell, although many B cells eventually switch to
other classes (19). It is also the
major class that is secreted into the blood in the early stages of
a primary antibody response on initial exposure to an Ag.
IgM is the first line of defense of an organism. In
its secreted form, IgM is a pentamer that comprises 5 4-chain
units, giving it a total of 10 Ag-binding sites and thus higher
valency than the structures of other immunoglobulins (Igs) and
allowing it to bind Ags with high avidity (20). Each pentamer contains one copy of
another polypeptide chain, called a J (joining) chain (21). IgM regulates B cell development
(22), facilitates the clearance of
apoptotic cells (23), modulates
inflammatory responses (24) and
autoimmune diseases (25) and
mediates the elimination of cancer cells (13).
The binding of an Ag to a single secreted pentameric
IgM molecule initiates the complement system. When the Ag resides
on the surface of an invading pathogen, senescent cells, cell
debris, or precancerous or cancer cells, this activation marks
pathogens and transformed cells for phagocytosis or kills them
directly (21).
Natural IgM antibodies
Natural Abs are predominantly IgM and to a lesser
extent IgA and IgG (26–28) and are polyreactive and of low
affinity (29). Natural IgM
circulates in healthy individuals in the absence of exogenous
antigenic stimulation or Ag-driven selection (30,31).
Natural IgM levels in the serum of newborns and in animals that are
grown under sterile conditions on an Ag-free diet do not differ
from those of normal animals (11).
Natural IgMs are also in humans (32).
Natural IgM has a significant function in primary
defense mechanisms (14,33,34).
They participate in the early recognition and elimination of
bacterial and viral invaders and altered self-material from an
organism, reacting with cell surface receptors and recognizing and
removing apoptotic and senescent cells, cell debris and self-Ags
(13,33,35–37).
Natural IgM auto-Abs help suppress pathogenic IgG auto-Ab responses
(38).
Natural IgM is associated with the recognition and
removal of precancerous and cancerous cells (13–18).
Natural IgM binds preferentially to post-transcriptionally modified
cell surface Ags that are tumor-specific, recognizing the conserved
structures of carbohydrate epitopes (14,39–42).
Carbohydrate epitopes that are recognized by natural IgM are stably
expressed in many tumors at various precursor stages. Unlike
epitope-based single-peptide chains, glycoepitopes share structural
homologies beyond the limits of the protein families; thus, they
can crossreact and constitute the preferred targets for natural IgM
Abs (35).
Natural IgM is produced by a small subset of B1
cells (CD5+) and B cells in the marginal zone (Mz) and
do not require affinity maturation to provide early protection
(43). B1 cells are
B220lowIgMhiCD23low/−CD43+IgDlow,
have the characteristics of activated cells, and have greater size
and cytoplasmic complexity than B2 cells (44).
Natural IgM Abs are germline-encoded and not
affinitymatured. Over 80% of natural IgM Abs are expressed by VH
genes of the VH3 family (45) and
have low affinity (kDa = 10−4 to 10−7
mol−1) (46). The
strength of the Ag-Ab interaction is enhanced by the potency of IgM
in engaging the complement pathway; unlike IgG, a single IgM
molecule can bind to C1q and activate the complement cascade
(18). Natural IgM is equipped with
a λ chain, unlike other Abs (14).
Adaptive IgM antibodies
Adaptive IgM is the first antibody to appear after
an immunological challenge, but its production normally falls
during the development of the IgG response. Consequently, IgM is
generally not considered to have a significant function in
long-term immunity, although it is effective in host defense
(19,47).
Long-lasting humoral immunity is typically
associated with the development of high-affinity Ab and isotype
switching (47). For example, IgM
that is induced by immunization differs from natural IgM with
regard to its structure in the Ag-binding centers, affinity,
specificity repertoire and spectrum of functions (11,48).
These IgM Abs constitute a small fraction of circulating molecules,
are monoreactive, and have higher affinity (10−7 to
10−11 mol−1), and their variable regions
contain point mutations. The half-life of monoreactive IgM is 35 h
(45,46).
Adaptive IgM is produced by B2 cells and follicular
B cells, which are typical of the adaptive immune response. B2
cells mediate T-dependent reactions of the germinal center (GC) and
effect the development of memory cells and highaffinity plasma
cells (29). Mature B2 cells
produce Ab after being stimulated, expanded, and selected in GCs in
the presence of T helper (Th) cells; thus, they are important in
the adaptive immune response, representing the first-line defense
against most infections and the only form of protection against
encapsulated bacteria. The adaptive immune response requires at
least 1 week to produce monospecific high-affinity Abs, first
generating IgM and then isotype-switching to IgG (49). B2 cells have a
B220hiIgMintCD23hiIgDhi
phenotype (44).
Fig. 1 shows the
various functions of natural and adaptive IgM, as identified in
murine models, and how IgM and IgG behave throughout the
development of a breast tumor. Natural and adaptive IgM levels are
constant from transformation until the tumor is established.
Whereas IgG is present only in the initial stages of adaptive
immunity, it becomes immunosuppressed when the breast tumor is
formed. In innate and adaptive immunity, natural IgM and adaptive
IgM, respectively, protect the organism from pathogenic infection,
cellular debris, senescence, and transformed cells using many
strategies, such as classical complement activation with C1q
(50). IgM has been proposed to
bind tightly to complement factor C1q and activate the complement
cascade (51). It also neutralizes
(52–54) and clears apoptotic cells by
phagocytosis (55), binding to
mannose-binding lectin (MBL), which interacts with apoptotic cells
(56), and directs the clearance of
immune complexes by binding to the putative Fca/mR receptor on
phagocytes.
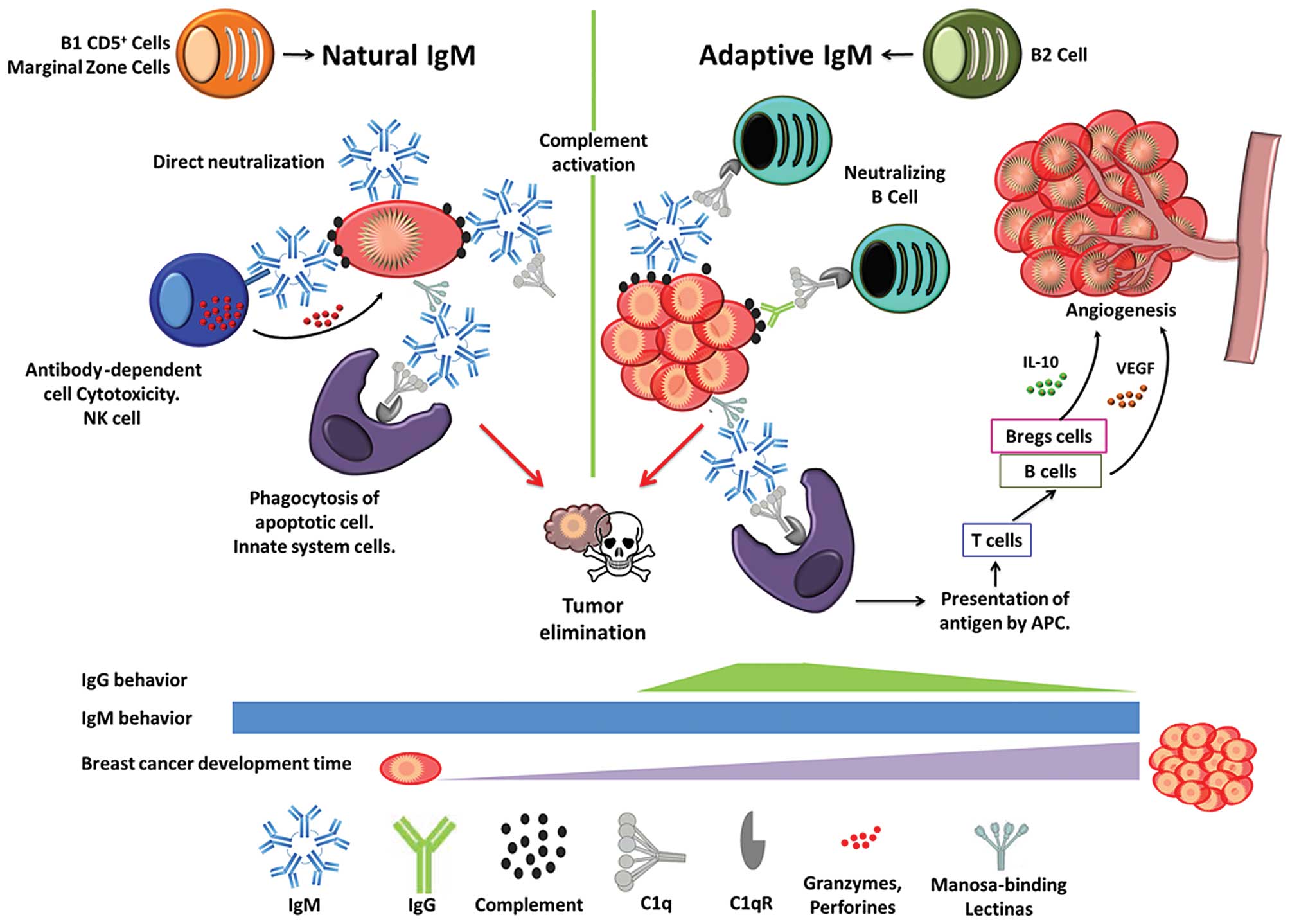 | Figure 1Natural IgM is produced by B1 cells
and marginal zone cells, and adaptive IgM is synthesized by B2
cells. Both types of IgM have several functions in the immune
response, eliminating tumor cells when they begin to transform
(natural IgM) and grow (adaptive IgM). But, when a tumor is
established, components of the immune system, such as B cells and
adaptive regulatory B cells (Bregs), secrete vascular endothelial
growth factor (VEGF) and IL-10, respectively, promoting
angiogenesis, inhibiting T cell responses, and accelerating
progression, all of which facilitate the spread of neoplastic
cells. IgG is present in the early stages of breast cancer but
becomes immunosuppressed over time, whereas IgM remains
constant. |
Natural and adaptive IgM molecules participate in
the recruitment of Ags into secondary lymphoid organs, priming
subsequent adaptive immune response (48,57).
This mechanism forms the link between the innate and adaptive
immune systems. In adaptive immunity, after IgM appears, T and B
cells are activated, and adaptive regulatory B cells (Bregs)
develop, the immunological mechanisms are damaged, and the
relationship between the tumor and immune response shifts toward a
state of conditioned immunosuppression (58,59),
causing late immune responses to fail to develop strategies that
eliminate tumor cells. B cells in the inflammatory infiltrate
effect the release of vascular endothelial growth factor (VEGF),
promoting angiogenesis and thus accelerating the spread of
neoplastic cells through the lymphatics to regional lymph nodes
(60). In addition, Bregs produce
IL-10, which has suppressive effects on systemic immunity, inhibits
T cell responses, and favors the induction of proinflammatory
factors and angiogenic molecules (58,61).
3. Immune surveillance: Mechanism to
eliminate cancer cells
In 1909, Paul Ehrlich postulated that the immune
system not only eliminates pathogenic bacteria but also suppresses
the growth of carcinomas with great frequency by generating Abs
against malignant cells (35).
Fifty years later, Burnet and Thomas revised the topic of natural
immune protection against cancer. Burnet proposed that immune
tolerance, with regard to tumor cell-specific neo-Ags, could effect
an immunological reaction that prevents the development of cancers,
defined as the immune surveillance concept (62,63).
Transformed cells are removed by immune surveillance, comprising an
immediate immune response that provides Abs against malignant cells
and a secondary inherited immune response in which B cells are
derived (35,43,64,65).
The Ab response is known as the humoral arm of the immune system
and is a critical mechanism in the primary and secondary responses
against all types of nonself (bacteria, viruses, fungi and cancer)
(15).
Innate immunity is the first line of defense and
stimulates secondary adaptive immune responses (66), relying on Toll-like receptors
(TLRs), which recognize pathogen-associated molecular patterns.
These specific patterns are conserved and repetitive structures,
such as carbohydrates on glycoproteins and glycolipids (eg,
lipopolysaccharides) that are expressed independently of mutational
events (67) and detected
independently of T cells (35).
4. Autoantibodies and tumor-associated
antigens in breast cancer
BC is a heterogeneous disease with tumors that
express a variety of aberrant proteins (68). Natural and adaptive IgM can perceive
foreign TAAs that undergo post-translational modifications, and
natural IgM mediates the destruction of tumor tissues that
recognize TACAs (69). The presence
of post-translational modifications, such as glycosylation,
phosphorylation, oxidation, and proteolysis, can induce immune
responses by generating a new epitope, inducing its presentation by
major histocompatibility complex (MHC) molecules and stimulating T
cell receptors (70,71). Such modified proteins are wrongly
localized, mutated, insufficiently folded, or aberrantly expressed
and are associated with carcinogenic processes (eg, cell cycle
progression, signal transduction, proliferation and apoptosis)
(70,71). Cell surface glycans that are
secreted into the serum by malignant cells provide a mechanism of
tracking tumor burden. Many malignant cells, but not normal cells,
overexpress CD20, ECFR and HER2, rendering them commonly used
diagnostic markers, but not for early diagnosis (42,72).
Aberrant proteins have been used since the 1970's in
serological studies in BC patients that have suggested that some
portion of positive sera contains Abs that are directed toward
TAAs, but they did not specify the type of Ab. The serum of 28
patients with BC showed positive fluorescence against breast
carcinoma cells that were grown by tissue culture, while that of
donors was negative (73). In
another study, the serum from BC patients was tested for reactivity
to a human breast tumor cell line; 45% of patients had
complement-fixing Abs compared with 13% that had benign breast
disease (74). In contrast, the
serum of 55 BC patients of all ages and stages of disease harbored
significantly elevated IgA levels and decreased IgG content vs. The
control group (75). Ig levels are
significantly lower in breast tissue compared with benign tissue,
except for adaptive IgM, which is consistently higher in BC tissue
of patients with stage I and II disease (76).
There are several blood tests that identify tumor
Ags at high levels in patients with metastatic disease, but they
are too insensitive for use in the early detection and diagnosis of
BC (68). TAAs have modulate
transmembrane signaling, which is required for proliferation,
invasion and metastasis of tumor cells (72). Natural and adaptive IgM against TAAs
can be measured as an early sign of BC in vivo and detect
the disease earlier than current methods; furthermore, natural IgM
is detected in the asymptomatic stages of cancer, up to 5 years
before disease onset (77).
Although there are IgG-based diagnostic biomarkers for BC that are
under development and although hundreds of self-Ags and TAAs that
are recognized by auto-Abs have been identified, no definitive
Ab-based sero-logical markers for the early diagnosis of BC
exist.
Auto-Abs against TAAs in the serum of BC patients
can be easily detected and are inherently stable, persisting in the
serum for long periods, since they generally do not undergo
proteolysis, as do other polypeptides (71).
5. IgM antibodies directed against breast
cancer tumor antigens
Both IgM types must be considered to develop a tool
for an early diagnosis once the tumor has been established, since
natural and adaptive IgM has direct cytotoxic effects on tumor
cells. Certain IgM Abs have been isolated from the tumors of
patients, since they eliminate tumors by inducing apoptosis in
vivo (25,43) through the domain-independent pathway
of cell death, binding to surface receptors that induce cell stress
(13). Following, we discuss
several types of IgM Abs that are used in the diagnosis of BC.
FC-2.15
FC-2.15 is a murine monoclonal IgM Ab that was
raised against human BC. FC-2.15 recognizes BC cells and certain
normal cells, such as peripheral polymorphonuclear granulocytes
(PMNs); specifically, the carbohydrate moiety of certain
glycoproteins mediates the in vitro lysis of
Ag-2.15+ cells by human complement. FC-2.15 induces
antitumor responses and reversible neutropenia. In an analysis of
epitope specificity, FC-2.15 specifically recognized terminally
exposed Lewisx trisaccharide but not
sialyl-Lewisx, Lewisa, trifucosylated
Lewisy, blood groups Ag A and B, globo H, or
gangliosides. Lewisx in its mostly O-linked is present
in BC cells.
In contrast to other monoclonal Abs against
carbohydrates, which have affinity constants of 103 to
105 M−1, FC-2.15 has an affinity constant of
6.9×107 M−1, which might explain its potent
effects in vivo. The presence of Lex epitopes on BC cells
and peripheral PMNs explains the antitumor responses and
neutropenia that were observed in a trial of FC-2.15. Neutropenia
was inconsequential to the patients, although >90% of PMNs
disappeared from the peripheral blood, and the neutropenia resolved
rapidly after mAb infusion was halted, with the appearance of
juvenile myeloid forms after repeated courses of mAb, as myeloid
precursors are not lysed by FC-2.15 (78).
Natural IgM antibody SC-1
The natural IgM Ab SC-1 was isolated from a patient
with signet-ring cell carcinoma of the stomach (79). SC-1 binds to a tumor-specific
carbohydrate epitope of decay acceleration factor-B (DAF; also
called CD55), which is specifically expressed in the membrane of
stomach carcinoma cells, and induces apoptosis by crosslinking the
receptor in vitro and in experimental in vivo systems
(35,80). The apoptotic effects of two novel
sorafenib analogs, SC-1 and SC-43, in eliminating BC cells were
examined. Sorafenib, SC-1 and SC-43 induced apoptosis, concurrent
with downregulation of p-STAT3 and its downstream proteins, cyclin
D1 and survivin, dose-dependently in BC cell lines (HCC-1937,
MDA-MB-468, MDA-MB-231, MDA-MB-453, SK-BR3 and MCF-7). SC-1 and
SC-43 also stimulated apoptosis through SHP-1-dependent STAT3
inactivation and had more potent apoptotic effects than sorafenib
in human BC cells (81).
Monoclonal IgM antibody PAM-1
The fully human germline-encoded monoclonal IgM Ab
PAM-1 was isolated from a patient with gastric carcinoma. The
blockade of growth factor receptors, such as EGFR and FGFR, which
are often overexpressed in malignant cells, led to starvation and
cell death. PAM-1 binds to CFr-1 (cysteine-rich fibroblast growth
factor receptor). The post-transcriptionally modified CFr-1/PAM-1
receptor is expressed in nearly all epithelial cancers of every
type and origin and in the precursor stages but not in healthy
tissue. The binding of PAM-1 induces apoptotic events in
vitro and in vivo (35,80).
CFR-1/PAM-1 receptor expression in the precancerous
stages of BC was analyzed by immunohistochemistry and compared with
normal breast tissue and adenocarcinomas. The CFR-1/PAM-1 receptor
was expressed in nearly all precancerous stages and carcinomas,
whereas normal breast tissue was negative. The unique expression of
this CFR-1/PAM-1 receptor renders PAM-1 Ab an ideal diagnostic tool
and therapeutic agent for precancerous and cancerous epithelial
lesions in BC (16).
Natural IgM antibody SAM-6
The levels of cell surface-associated chaperone
GRP78 are high in BC cells. GRP78 is a ubiquitously expressed
member of the heat-shock protein 70 (HSP70) family and governs
cellular homeostasis by preventing stress-induced apoptosis. In
malignant cells, which are permanently exposed to environmental
stress, GRP78 is overexpressed, and its levels increase in the
cytoplasm and on the cell membrane (41). Thus, GRP78 promotes tumor
proliferation, survival, metastases and resistance to many
therapies.
The fully human monoclonal IgM Ab SAM-6 binds to a
new variant of GRP78, which has a molecular weight of 82 kDa. The
epitope is an O-linked carbohydrate moiety that is specific to
malignant cells (13). SAM-6 is
internalized through endocytosis and mediates the lethal
accumulation of oxidized lipoproteins, followed by apoptosis.
Modified protective molecules, such as GRP78-SAM-6, are excellent
targets for specific Abs that can neutralize the protective effects
of tumor cells, disable mechanisms of drug resistance, and directly
kill cancer cells by inducing apoptosis (13,41).
SAM-6 induces apoptosis and the accumulation of
neutral and polar lipids in tumor cells but not normal cells. The
non-physiological intracellular accumulation of neutral lipids,
such as triglycerides and cholesterol, is cytotoxic and can lead to
lipoptosis (35,65). SAM-6 binds to a cell surface
receptor on malignant cells and oxidized low-density lipoprotein
(LDL). Shortly after the internalization of Ab/oxidized
LDL/receptor complexes and the formation of lipid depots,
cytochrome c is released by mitochondria and subsequently,
the initiators caspase-8 and caspase-9 and effectors caspase-3 and
caspase-6 are activated. Thus, SAM-6 induces a near-intrinsic form
of apoptosis by overfeeding malignant cells with lipoproteins
(80,82).
Murine monoclonal IgM antibody 3EL.2
Mammary serum Ag (MSA) belongs to the molecular
family of breast mucins (MUCs) and is a macromolecular glycolic
protein with a molecular weight of >300,000 kDa. MSA is targeted
by the murine monoclonal IgM Ab 3EL.2. The diagnostic value of MSA
in identifying BC has been studied in 56 healthy patients and 43
subjects with benign BC, in whom this Ag was abnormally elevated.
Thus, 3EL.2 is useful in the clinic, is a good indicator of the
extent of disease, and might have significant prognostic value
(83).
P10s
Abs that recognize such glycosphingolipids (GSLs) as
GD2, GM2 and LewisY (LeY) mediate complement-dependent
cytotoxicity and have been suggested to be more cytotoxic to tumor
cells than Abs that recognize proteins Ag or TACAs (84), which kill tumor cells by
Ab-dependent cellular cytotoxicity. Carbohydrate mimetic peptides
(CMPs) of TACAs induce IgM that targets TACAs in BC.
Preexisting ganglioside-reactive IgM has been
detected in normal healthy individuals. Circulating gangliosides
from tumors might be perceived as danger signals by the host's
immune system, as evidenced by the endogenous antiganglioside
immune response to gangliosides. Then, endogenous IgM against
gangliosides might facilitate the elimination of these signals in
BC in order to restore the immune competence of the host (69).
P10s-WRYTAPVHLGDG, a CMP that induces primarily weak
anti-GD2 IgM responses that are crossreactive with several
gangliosides, including GD3, GM2 and GD1a, has been developed. P10s
was derived from a sequence (P10-GVVWRY-TAPVHLGDG) that was
selected by panning a peptide library against the GD2-binding mAb
ME36.1. The P10 peptide mediates antitumor responses (85). This sequence was further optimized
by molecular modeling to overlap its binding interface more with
the ME36.1 paratope, thus yielding P10s (86).
Anti-mucin IgM
MUCs are highly glycosylated proteins that are
expressed in cancers of epithelial origin in an underglycosylated
form and have been used to develop several tests for cancer
detection. MUCs are components of mammary cell-cell junctions and
mediate ICAM-1-initiated signal transduction. Polymorphic
epithelial mucin (PEM, or MUC1 with different epitopes: CA 15.3 and
CA 27.29) and MUC16 (CA 125) are the most extensively studied MUCs,
although the latter is more frequently used for ovarian cancer than
for BC.
Recent evidence indicates that MUC-1 Ag induces
apoptosis in T-lymphocytes, providing insight into the mechanisms
of escape from immune surveillance by tumors (87).
MUC1 was examined to determine the incidence of
naturally occurring MUC1 Ab in patients with early BC and correlate
these Abs in pretreatment serum to disease outcome. IgG and IgM
against MUC1 were measured by ELISA in pretreatment serum samples
from 154 patients with BC and 302 controls. A positive test result
for both anti-MUC1 IgG and IgM in pretreatment serum was associated
with a significant benefit for disease-specific survival in BC
patients. Patients with early BC with a natural humoral response to
MUC1 are less likely to develop metastases and have better
disease-specific survival. MUC1 Abs may control hematogenic tumor
dissemination and outgrowth by aiding in the destruction of
circulating or established MUC1-expressing tumor cells (88,89).
MUC1 is a high-molecular-weight molecule with
multiple tandem repeats (VTSAPDTRPAPGSTAP-PAHG). The most
immunogenic motif is APDTRPA, which harbors the epitope that is
recognized by various monoclonal Abs, normal sera and cytotoxic
T-cells. Mice that are vaccinated with MUC1 peptide that contains
1.5 tandem repeats and is conjugated to keyhole limpet hemocyanin
(KLH) and mixed with QS-21 induce high-titer Ab (but no evidence of
T-cell immunity) against MUC1 and MUC1-expressing tumor cells.
Furthermore, these vaccinations conferred protection to these mice
when they were challenged with MUC1-expressing tumor cells
(90).
Anti-CEA IgM
Carcinoembryonic Ag (CEA) was described in 1965 and
was the first tumor Ag to be identified. CEA is a glycoprotein that
belongs to the Ig family of genes and is detected in the serum of
cancer patients by radioimmunoassay or ELISA. However, its clinical
value is limited due to a high false-positive rate in normal
populations and its low diagnostic sensitivity and specificity.
Elevated CEA levels are not specific for BC (50% of cases), CEA is
expressed in many types of neoplasia and is detected by anti-CEA
IgM. Nevertheless, in breast tumors, CEA is more prevalent in
ductal versus lobular carcinomas. CEA is found in patients with
ductal carcinoma in situ, suggesting that it is an early marker of
tumorigenesis (91,92).
I antigen and IgM antibodies
The ABH blood group Ags, which are expressed in
normal epithelial cells, are downregulated in various carcinomas.
BC cells produce I Ag or I Ag-like substances that influence serum
anti-I. I Ags are precursors to ABH and accumulate in cancer cells.
The levels of I Ag rise in the serum of individuals with breast
carcinoma, based on their anti-I scores and IgM concentrations.
These alterations in cold hemagglutinins could be a host response
to the production of I Ag by BC cells and is thus important in
understanding immune modulation of breast carcinoma. However, the
concentration of IgM in patients with BC was similar to that of
controls (93).
Anti-malignin antibodies in serum
(AMAS)
Malignin is a 10-kDa polypeptide in the cytoplasmic
and outer membranes of all malignant cells. Anti-malignin Ab (AMA)
is an IgM that is spontaneously produced by the host against the
oncoprotein malignin when neoplastic transformation occurs; because
AMAs are IgM molecules, they are an indicator of 'early'
transformation that is useful for the early detection of
cancer.
Elevated AMA serum concentrations have been measured
using a commercial reagent. The AMA test has a sensitivity and
specificity of 95% on first determination and >99% on repeat
measurements and is a promising diagnostic tool for the early
detection of cancer, the monitoring of treatment responses, and the
screening of asymptomatic populations (94).
In tumor marker assays with AMAs, CEA (CA15-3,
CA19-9 and CA125) and biopsies were examined following a suspicious
mammogram to determine whether tumor markers aided in the diagnosis
and could be used to monitor residual disease. In the present
study, by AMA test, 3 of 154 healthy volunteers were AMA-positive.
After further examination, 2 were positive for cancer and 1 had a
history of ulcerative colitis. Tumor biopsies of 43 suspicious
patients by mammography revealed that 32 were cancerous and 11 were
benign by pathology. Furthermore, 31 of 32 cancer patients were
positive for AMA versus 4 of 11 pathological benign cases (95).
Sialyl Tn antigen and IgM antibodies
Several structurally similar blood group-related
carbohydrate Ags, including Thomsen-Freidenreich (TF), Tn, and
sialyl Tn (sTn), that are attached to the protein backbones of
glycoproteins are promising targets, based on their widespread
distribution on the cell surface of human tumors. The disaccharide
sTn [NeuAca (2→6) GalNAca-0-ser/Thr] is O-linked with serine and
threonine residues on mucins. Greater sTn expression in tumors
might be linked to a poorer prognosis in BC. Abnormal glycosylation
of tumor cell MUCs results in shorter and fewer carbohydrate
chains, increasing the exposure of Ags, such as sTn, which might
upregulate sTn in tumors compared with normal cells.
The induction of IgM and IgG against synthetic
sTn(c) was measured before and after immunization with clustered
sTn-KLH [sTn(c)-KLH] conjugate plus QS-21 in 27 patients with BC,
all of whom developed significant IgM and IgG titers against
sTn(c). Furthermore, IgM reactivity against LSC tumor cells was
observed in patients, indicating the production of IgM and IgG.
Thus, immunization with sTn(c)-KLH conjugate plus QS-21 is well
tolerated and immunogenic in high-risk patients with BC (96).
IgM and IgG complexes
The occurrence of circulating immune complexes
(CICs) is considered a marker of tumor burden. CICs are formed by
Abs or Ags with and without complement. CICs that comprise two
classes of Ig (Ig-Ig) or an Ig class of and C3 (Ig-C) are
collectively referred to as two-component-determined CICs (TCICs).
Ig/Ig TCICs may reveal alterations in immune regulation in
patients. IgM and IgG TCICs were measured in the sera of patients
with BC, in whom IgM/IgG TCICs and IgG/IgM TCICs were detected.
Downregulation of IgM/IgG TCICs was a common feature in patients,
whereas IgG/IgM TCIC levels were significantly higher, lower, or
unchanged with respect to the control. Total serum IgM differed
significantly in BC patients (1.20±0.94 mg/ml) vs. healthy controls
(0.99±0.53 mg/ml). These results suggested that IgM and IgG TCICs
have a significant function in immune regulation during the course
of malignancies and are hallmarks of cancer pathogenesis. Decreased
IgM/IgG TCIC levels, accompanied by IgG/IgM-TCICs, constitute a
peculiar trait in malignancies (97).
6. Conclusions
Certain IgMs, natural and adaptive, have been
isolated from the tumors of patients with cancer; they eliminate BC
tumors through various mechanisms, such as apoptosis and
complement. Natural IgM has a direct cytotoxic effect on tumor
cells, it recognizes tumor-modified cell surfaces that develop
during tumorigenesis, and it activates complement to destroy
nascent transformed cells. The first Ig that is produced after an
immune challenge is adaptive IgM, which must be considered as an
early diagnostic tool. Auto-Abs that target TAAs could serve as
molecular signatures for the early diagnosis and prognosis of
patients with BC by serology to increase the sensitivity and
specificity of diagnostic markers for BC patients. Most studies
have focused on the development of IgG-like biomarkers for BC
treatment; however, IgG is subject to immunoregulation, which can
manifest as immunosuppression, whereas natural IgM is not. IgM as a
diagnostic tool can be coupled with early mammography, magnetic
resonance imaging, or Doppler ultrasound to detect cancer.
Acknowledgments
Financial support was provided by Grants #151747
(P.O.-S.) from the Consejo Nacional de Ciencia y Tecnología, México
and #IN201715-3 (P.O.-S.) from Programa de Apoyo a Proyectos de
Investigación e Innovación Tecnológica (PAPITT), DGAPA Universidad
Nacional Autónoma de México (UNAM). M. Díaz-Zaragoza acknowledges
the scholarship and financial support provided by the National
Council of Science and Technology (CONACyT) and the Program in
Biological Sciences, UNAM. We thank Jorge Lopez for his support
with the writing of this manuscript.
References
|
1
|
Etzioni R, Urban N, Ramsey S, McIntosh M,
Schwartz S, Reid B, Radich J, Anderson G and Hartwell L: The case
for early detection. Nat Rev Cancer. 3:243–252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Levenson VV: Biomarkers for early
detection of breast cancer: What, when, and where? Biochim Biophys
Acta. 1770:847–856. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Panieri E: Breast cancer screening in
developing countries. Best Pract Res Clin Obstet Gynaecol.
26:283–290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Desmetz C, Mange A, Maudelonde T and
Solassol J: Autoantibody signatures: Progress and perspectives for
early cancer detection. J Cell Mol Med. 15:2013–2024. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
WHO: Early detection of cancer. http://www.who.int/cancer/detection/en/.
|
|
6
|
Lu H, Goodell V and Disis ML: Humoral
immunity directed against tumor-associated antigens as potential
biomarkers for the early diagnosis of cancer. J Proteome Res.
7:1388–1394. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Disis ML, Pupa SM, Gralow JR, Dittadi R,
Menard S and Cheever MA: High-titer HER-2/neu protein-specific
antibody can be detected in patients with early-stage breast
cancer. J Clin Oncol. 15:3363–3367. 1997.PubMed/NCBI
|
|
8
|
Carter P, Smith L and Ryan M:
Identification and validation of cell surface antigens for antibody
targeting in oncology. Endocr Relat Cancer. 11:659–687. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
WHO: Breast cancer: prevention and
control. http://www.who.int/cancer/detection/breastcancer/en/.
|
|
10
|
Díaz-Zaragoza M, Hernández R and
Ostoa-Saloma P: 2D immunoblots show differential response of mouse
IgG and IgM antibodies to antigens of mammary carcinoma 4 T1 cells.
Cancer Cell Int. 14:92014. View Article : Google Scholar
|
|
11
|
Klimovich VB: IgM and its receptors:
Structural and functional aspects. Biochemistry (Mosc). 76:534–549.
2011. View Article : Google Scholar
|
|
12
|
Reynolds AE, Kuraoka M and Kelsoe G:
Natural IgM is produced by CD5− plasma cells that occupy
a distinct survival niche in bone marrow. J Immunol. 194:231–242.
2015. View Article : Google Scholar
|
|
13
|
Vollmers HP and Brändlein S: Natural
antibodies and cancer. N Biotechnol. 25:294–298. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brändlein S, Pohle T, Ruoff N, Wozniak E,
Müller-Hermelink HK and Vollmers HP: Natural IgM antibodies and
immunosurveillance mechanisms against epithelial cancer cells in
humans. Cancer Res. 63:7995–8005. 2003.PubMed/NCBI
|
|
15
|
Vollmers HP and Brändlein S: The 'early
birds': Natural IgM antibodies and immune surveillance. Histol
Histopathol. 20:927–937. 2005.PubMed/NCBI
|
|
16
|
Brändlein S, Eck M, Ströbel P, Wozniak E,
Müller-Hermelink HK, Hensel F and Vollmers HP: PAM-1, a natural
human IgM antibody as new tool for detection of breast and prostate
precursors. Hum Antibodies. 13:97–104. 2004.
|
|
17
|
Vollmers HP and Brändlein S: Death by
stress: Natural IgM-induced apoptosis. Methods Find Exp Clin
Pharmacol. 27:185–191. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Manson JJ, Mauri C and Ehrenstein MR:
Natural serum IgM maintains immunological homeostasis and prevents
autoimmunity. Springer Semin Immunopathol. 26:425–432. 2005.
View Article : Google Scholar
|
|
19
|
Racine R, McLaughlin M, Jones DD, Wittmer
ST, MacNamara KC, Woodland DL and Winslow GM: IgM production by
bone marrow plasmablasts contributes to long-term protection
against intracellular bacterial infection. J Immunol.
186:1011–1021. 2011. View Article : Google Scholar :
|
|
20
|
Bendtzen K, Hansen MB, Ross C and Svenson
M: High-avidity autoantibodies to cytokines. Immunol Today.
19:209–211. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Alberts B, Johnson A, Lewis J, Raff M,
Roberts K and Walter P: Molecular Biology of the Cell. B Cells and
Antibodies. 4th edition. Garland Science; New York, NY: 2002
|
|
22
|
Lim HW, Hillsamer P, Banham AH and Kim CH:
Cutting edge: Direct suppression of B cells by CD4+
CD25+ regulatory T cells. J Immunol. 175:4180–4183.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shaw PX, Hörkkö S, Chang MK, Curtiss LK,
Palinski W, Silverman GJ and Witztum JL: Natural antibodies with
the T15 idiotype may act in atherosclerosis, apoptotic clearance,
and protective immunity. J Clin Invest. 105:1731–1740. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang M, Austen WG Jr, Chiu I, Alicot EM,
Hung R, Ma M, Verna N, Xu M, Hechtman HB, Moore FD Jr, et al:
Identification of a specific self-reactive IgM antibody that
initiates intestinal ischemia/reperfusion injury. Proc Natl Acad
Sci USA. 101:3886–3891. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ray SK, Putterman C and Diamond B:
Pathogenic autoantibodies are routinely generated during the
response to foreign antigen: A paradigm for autoimmune disease.
Proc Natl Acad Sci USA. 93:2019–2024. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Casali P and Schettino EW: Structure and
function of natural antibodies. Curr Top Microbiol Immunol.
210:167–179. 1996.PubMed/NCBI
|
|
27
|
Coutinho A, Kazatchkine MD and Avrameas S:
Natural autoantibodies. Curr Opin Immunol. 7:812–818. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Panda S and Ding JL: Natural antibodies
bridge innate and adaptive immunity. J Immunol. 194:13–20. 2015.
View Article : Google Scholar
|
|
29
|
Lopes-Carvalho T and Kearney JF:
Development and selection of marginal zone B cells. Immunol Rev.
197:192–205. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Madi A, Hecht I, Bransburg-Zabary S, Merbl
Y, Pick A, Zucker-Toledano M, Quintana FJ, Tauber AI, Cohen IR and
Ben-Jacob E: Organization of the autoantibody repertoire in healthy
newborns and adults revealed by system level informatics of antigen
microarray data. Proc Natl Acad Sci USA. 106:14484–14489. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Merbl Y, Zucker-Toledano M, Quintana FJ
and Cohen IR: Newborn humans manifest autoantibodies to defined
self molecules detected by antigen microarray informatics. J Clin
Invest. 117:712–718. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schettino EW, Chai SK, Kasaian MT,
Schroeder HW Jr and Casali P: VHDJH gene sequences and antigen
reactivity of monoclonal antibodies produced by human B-1 cells:
Evidence for somatic selection. J Immunol. 158:2477–2489.
1997.PubMed/NCBI
|
|
33
|
Bohn J: Are natural antibodies involved in
tumour defence? Immunol Lett. 69:317–320. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Boes M: Role of natural and immune IgM
antibodies in immune responses. Mol Immunol. 37:1141–1149. 2000.
View Article : Google Scholar
|
|
35
|
Vollmers HP and Brändlein S: Natural IgM
antibodies: From parias to parvenus. Histol Histopathol.
21:1355–1366. 2006.PubMed/NCBI
|
|
36
|
Baumgarth N: The double life of a B-1
cell: Self-reactivity selects for protective effector functions.
Nat Rev Immunol. 11:34–46. 2011. View Article : Google Scholar
|
|
37
|
Nagele EP, Han M, Acharya NK, DeMarshall
C, Kosciuk MC and Nagele RG: Natural IgG autoantibodies are
abundant and ubiquitous in human sera, and their number is
influenced by age, gender, and disease. PLoS One. 8:e607262013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Boes M, Schmidt T, Linkemann K, Beaudette
BC, MarshakRothstein A and Chen J: Accelerated development of IgG
autoantibodies and autoimmune disease in the absence of secreted
IgM. Proc Natl Acad Sci USA. 97:1184–1189. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hensel F, Brändlein S, Eck M, Schmidt K,
Krenn V, Kloetzer A, Bachi A, Mann M, Müller-Hermelink HK and
Vollmers HP: A novel proliferation-associated variant of CFR-1
defined by a human monoclonal antibody. Lab Invest. 81:1097–1108.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hensel F, Hermann R, Schubert C, Abé N,
Schmidt K, Franke A, Shevchenko A, Mann M, Müller-Hermelink HK and
Vollmers HP: Characterization of glycosylphosphatidylinositollinked
molecule CD55/decay-accelerating factor as the receptor for
antibody SC-1-induced apoptosis. Cancer Res. 59:5299–5306.
1999.PubMed/NCBI
|
|
41
|
Rauschert N, Brändlein S, Holzinger E,
Hensel F, MüllerHermelink H-K and Vollmers HP: A new tumor-specifc
variant of GRP78 as target for antibody-based therapy. Lab Invest.
88:375–386. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kobata A and Amano J: Altered
glycosylation of proteins produced by malignant cells, and
application for the diagnosis and immunotherapy of tumours. Immunol
Cell Biol. 83:429–439. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Milner ECB, Anolik J, Cappione A and Sanz
I: Human innate B cells: A link between host defense and
autoimmunity? Springer Semin Immunopathol. 26:433–452. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Merino MC and Gruppi A: Origen y
desarrollo de linfocitos B1, una población celular involucrada en
defensa y autoinmunidad. Med (Buenos Aires). 165–172. 2006.
|
|
45
|
Notkins AL: Polyreactivity of antibody
molecules. Trends Immunol. 25:174–179. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou Z-H, Tzioufas AG and Notkins AL:
Properties and function of polyreactive antibodies and polyreactive
antigen-binding B cells. J Autoimmun. 29:219–228. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Manz RA, Hauser AE, Hiepe F and Radbruch
A: Maintenance of serum antibody levels. Annu Rev Immunol.
23:367–386. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Baumgarth N, Chen J, Herman OC, Jager GC
and Herzenberg LA: The role of B-1 and B-2 cells in immune
protection from influenza virus infection. Curr Top Microbiol
Immunol. 252:163–169. 2000.PubMed/NCBI
|
|
49
|
Carsetti R, Rosado MM and Wardmann H:
Peripheral development of B cells in mouse and man. Immunol Rev.
197:179–191. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ogden CA, Kowalewski R, Peng Y, Montenegro
V and Elkon KB: IgM is required for efficient complement mediated
phagocytosis of apoptotic cells in vivo. Autoimmunity. 38:259–264.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Quartier P, Potter PK, Ehrenstein MR,
Walport MJ and Botto M: Predominant role of IgM-dependent
activation of the classical pathway in the clearance of dying cells
by murine bone marrowderived macrophages in vitro. Eur J Immunol.
35:252–260. 2005. View Article : Google Scholar
|
|
52
|
Ochsenbein AF, Fehr T, Lutz C, Suter M,
Brombacher F, Hengartner H and Zinkernagel RM: Control of early
viral and bacterial distribution and disease by natural antibodies.
Science. 286:2156–2159. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Baumgarth N, Herman OC, Jager GC, Brown
LE, Herzenberg LA and Chen J: B-1 and B-2 cell-derived
immunoglobulin M antibodies are nonredundant components of the
protective response to influenza virus infection. J Exp Med.
192:271–280. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jayasekera JP, Moseman EA and Carroll MC:
Natural antibody and complement mediate neutralization of influenza
virus in the absence of prior immunity. J Virol. 81:3487–3494.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen Y, Khanna S, Goodyear CS, Park YB,
Raz E, Thiel S, Grönwall C, Vas J, Boyle DL, Corr M, et al:
Regulation of dendritic cells and macrophages by an anti-apoptotic
cell natural antibody that suppresses TLR responses and inhibits
inflammatory arthritis. J Immunol. 183:1346–1359. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Nauta AJ, Raaschou-Jensen N, Roos A, Daha
MR, Madsen HO, Borrias-Essers MC, Ryder LP, Koch C and Garred P:
Mannosebinding lectin engagement with late apoptotic and necrotic
cells. Eur J Immunol. 33:2853–2863. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Boes M, Esau C, Fischer MB, Schmidt T,
Carroll M and Chen J: Enhanced B-1 cell development, but impaired
IgG antibody responses in mice deficient in secreted IgM. J
Immunol. 160:4776–4787. 1998.PubMed/NCBI
|
|
58
|
Rodríguez RC and Padilla CR: Compromiso
del Sistema Inmune en Pacientes con Cáncer de Mama. Cancerología.
3:191–197. 2008.In spanish.
|
|
59
|
Zhang X: Regulatory functions of
innate-like B cells. Cell Mol Immunol. 10:113–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Coronella JA, Spier C, Welch M, Trevor KT,
Stopeck AT, Villar H and Hersh EM: Antigen-driven oligoclonal
expansion of tumor-infiltrating B cells in infiltrating ductal
carcinoma of the breast. J Immunol. 169:1829–1836. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Llanes-Fernández L, Alvarez-Goyanes RI,
Arango-Prado MC, Alcocer-González JM, Mojarrieta JC, Pérez XE,
López MO, Odio SF, Camacho-Rodríguez R, Guerra-Yi ME, et al:
Relationship between IL-10 and tumor markers in breast cancer
patients. Breast. 15:482–489. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Burnet FM: Immunological surveillance in
neoplasia. Transplant Rev. 7:3–25. 1971.PubMed/NCBI
|
|
63
|
Dunn GP, Old LJ and Schreiber RD: The
immunobiology of cancer immunosurveillance and immunoediting.
Immunity. 21:137–148. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Karin M, Lawrence T and Nizet V: Innate
immunity gone awry: Linking microbial infections to chronic
inflammation and cancer. Cell. 124:823–835. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Vollmers HP and Brändlein S: Natural IgM
antibodies: The orphaned molecules in immune surveillance. Adv Drug
Deliv Rev. 58:755–765. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hoebe K, Janssen E and Beutler B: The
interface between innate and adaptive immunity. Nat Immunol.
5:971–974. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Janeway CA Jr: Pillars article:
approaching the asymptote? Evolution and revolution in immunology.
Cold Spring Harb Symp Quant Biol. 54:1–13. 1989.reprinted in J
Immunol 191: 4475–4487, 2013. View Article : Google Scholar
|
|
68
|
Molina R, Barak V, van Dalen A, Duffy MJ,
Einarsson R, Gion M, Goike H, Lamerz R, Nap M, Sölétormos G, et al:
Tumor markers in breast cancer- European Group on Tumor Markers
recommendations. Tumour Biol. 26:281–293. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Monzavi-Karbassi B, Hennings LJ, Artaud C,
Liu T, Jousheghany F, Pashov A, Murali R, Hutchins LF and
Kieber-Emmons T: Preclinical studies of carbohydrate mimetic
peptide vaccines for breast cancer and melanoma. Vaccine.
25:3022–3031. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Anderson KS, Ramachandran N, Wong J,
Raphael JV, Hainsworth E, Demirkan G, Cramer D, Aronzon D, Hodi FS,
Harris L, et al: Application of protein microarrays for multiplexed
detection of antibodies to tumor antigens in breast cancer. J
Proteome Res. 7:1490–1499. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Tan HT, Low J, Lim SG and Chung MCM: Serum
autoantibodies as biomarkers for early cancer detection. FEBs J.
276:6880–6904. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Shishido SN, Varahan S, Yuan K, Li X and
Fleming SD: Humoral innate immune response and disease. Clin
Immunol. 144:142–158. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Priori ES, Seman G, Dmochowski L, Gallager
HS and Anderson DE: Immunofluorescence studies on sera of patients
with breast carcinoma. Cancer. 28:1462–1471. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chan SP, Maca RD, Levine PH and Ting RC:
Immunologic studies of human breast cancer. I. Serum reactivity
against a lymphoid cell line (Belev) derived from a breast cancer
patient as detected by complement-fixation test. J Natl Cancer
Inst. 47:511–517. 1971.PubMed/NCBI
|
|
75
|
Roberts MM, Bathgate EM and Stevenson A:
Serum immunoglobulin levels in patients with breast cancer. Cancer.
36:221–224. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Roberts MM, Bass EM, Wallace IW and
Stevenson A: Local immunoglobulin production in breast cancer. Br J
Cancer. 27:269–275. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Fernández Madrid F: Autoantibodies in
breast cancer sera: Candidate biomarkers and reporters of
tumorigenesis. Cancer Lett. 230:187–198. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Capurro M, Bover L, Portela P, Livingston
P and Mordoh J: FC-215, a monoclonal antibody active against human
breast cancer, specifically recognizes Lewis (x) hapten. Cancer
Immunol Immunother. 45:334–339. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hensel F, Hermann R, Brändlein S, Krenn V,
Schmausser B, Geis S, Müller-Hermelink HK and Vollmers HP:
Regulation of the new coexpressed CD55 (decay-accelerating factor)
receptor on stomach carcinoma cells involved in antibody
SC-1-induced apoptosis. Lab Invest. 81:1553–1563. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Pohle T, Brändlein S, Ruoff N,
Müller-Hermelink HK and Vollmers HP and Vollmers HP: Lipoptosis:
Tumor-specific cell death by antibody-induced intracellular lipid
accumulation. Cancer Res. 64:3900–3906. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Liu CY, Tseng LM, Su JC, Chang KC, Chu PY,
Tai WT, shiau CW and Chen KF: Novel sorafenib analogues induce
apoptosis through SHP-1 dependent STAT3 inactivation in human
breast cancer cells. Breast Cancer Res. 15:R632013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Brändlein S, Rauschert N, Rasche L,
Dreykluft A, Hensel F, Conzelmann E, Müller-Hermelink HK and
Vollmers HP: The human IgM antibody SAM-6 induces tumor-specific
apoptosis with oxidized low-density lipoprotein. Mol Cancer Ther.
6:326–333. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Verring A, Clouth A, Ziolkowski P and
Oremek GM: Clinical usefulness of cancer markers in primary breast
cancer. ISRN Pathol. 2011:8176182011. View Article : Google Scholar
|
|
84
|
Ragupathi G, Liu NX, Musselli C, Powell S,
Lloyd K and Livingston PO: Antibodies against tumor cell
glycolipids and proteins, but not mucins, mediate
complement-dependent cytotoxicity. J Immunol. 174:5706–5712. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wondimu A, Zhang T, Kieber-Emmons T,
Gimotty P, Sproesser K, Somasundaram R, Ferrone S, Tsao CY and
Herlyn D: Peptides mimicking GD2 ganglioside elicit cellular,
humoral and tumor-protective immune responses in mice. Cancer
Immunol Immunother. 57:1079–1089. 2008. View Article : Google Scholar
|
|
86
|
Pashov A, Monzavi-Karbassi B and
Kieber-Emmons T: Immune surveillance and immunotherapy: Lessons
from carbohydrate mimotopes. Vaccine. 27:3405–3415. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Hadden JW: The immunology and
immunotherapy of breast cancer: An update. Int J Immunopharmacol.
21:79–101. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
von Mensdorff-Pouilly S, Verstraeten AA,
Kenemans P, Snijdewint FG, Kok A, van Kamp GJ, Paul MA, Van Diest
PJ, Meijer S and Hilgers J: Survival in early breast cancer
patients is favorably infuenced by a natural humoral immune
response to polymorphic epithelial mucin. J Clin oncol. 18:574–583.
2000.PubMed/NCBI
|
|
89
|
Gilewski T, Adluri S, Ragupathi G, Zhang
S, Yao TJ, Panageas K, Moynahan M, Houghton A, Norton L and
Livingston PO: Vaccination of high-risk breast cancer patients with
mucin-1 (MUC1) keyhole limpet hemocyanin conjugate plus QS-21. Clin
Cancer Res. 6:1693–1701. 2000.PubMed/NCBI
|
|
90
|
Adluri S, Gilewski T, Zhang S, Ramnath V,
Ragupathi G and Livingston P: Specificity analysis of sera from
breast cancer patients vaccinated with MUC1-KLH plus QS-21. Br J
Cancer. 79:1806–1812. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Conry RM, Allen KO, Lee S, Moore SE, Shaw
DR and LoBuglio AF: Human autoantibodies to carcinoembryonic
antigen (CEA) induced by a vaccinia-CEA vaccine. Clin Cancer Res.
6:34–41. 2000.PubMed/NCBI
|
|
92
|
Albanopoulos K, Armakolas A,
Konstadoulakis MM, leandros E, Tsiompanou E, Katsaragakis S,
Alexiou D and Androulakis G: Prognostic significance of circulating
antibodies against carcinoembryonic antigen (anti-CEA) in patients
with colon cancer. Am J Gastroenterol. 95:1056–1061. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Dube VE, Haid M, Chmiel JS and Anderson B:
Serum cold agglutinin and IgM levels in breast carcinoma. Breast
Cancer Res Treat. 4:105–108. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Botti C, Martinetti A, Nerini-Molteni S
and Ferrari L: Antimalignin antibody evaluation: A possible
challenge for cancer management. Int J Biol Markers. 12:141–147.
1997.
|
|
95
|
Thornthwaite JT: Anti-malignin antibody in
serum and other tumor marker determinations in breast cancer.
Cancerlett. 148:39–48. 2000.
|
|
96
|
Gilewski TA, Ragupathi G, Dickler M,
Powell S, Bhuta S, Panageas K, Koganty RR, Chin-Eng J, Hudis C,
Norton L, et al: Immunization of high-risk breast cancer patients
with clustered sTn-KLH conjugate plus the immunologic adjuvant
QS-21. Clin Cancer Res. 13:2977–2985. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Yang TC, Li H, Huang GN and Wang SY:
Detection of IgM and IgG complexes provides new insight into immune
regulation of patients with malignancies: A randomized controlled
trial. Int Immunopharmacol. 7:1433–1441. 2007. View Article : Google Scholar : PubMed/NCBI
|














