Introduction
Bladder cancer is one of the most prevalent lethal
diseases among men worldwide (1).
The development of bladder tumors combines multiple progressive
processes that are influenced by a number of environmental factors
such as chemical carcinogens, several anticancer drugs and reactive
oxygen species (2–5). The principal proliferative type of
bladder tumors can be described as a transitional cell carcinoma
(TCC) that is a muscle-invasive bladder cancer (MIBC), which has
potential for migration and invasion (1,6,7).
Although many therapeutic advances have been achieved against MIBC,
effective treatment options remain limited. Therefore, the
identification of novel therapeutic targets is critical.
Cellular proliferation experiences a crucial
check-point at which the progression of the G1 to the S phase of
the cell cycle is regulated (8,9). The
progression of the cell cycle is directly driven by the action of
heterodimers assembled by cyclin-dependent kinases (CDKs) and
cyclins (activating subunits of CDKs) (8,9). After
the stimulation of growth signaling, the cells are categorized as
either a G1 or S phase progression, which is predominantly
regulated by cyclin D/CDK4/6 and cyclin E/CDK2 complexes (8,9).
Numerous studies have suggested negative regulatory roles for cell
cycle inhibitors that are involved in proliferation, migration and
invasion in the development of mammalian cells (8). p21WAF1 is a cell cycle
inhibitor that binds to CDK or CDK-cyclin complexes, and suppresses
cell cycle progression (8).
Matrix metalloproteinase-2 (MMP-2) is a type IV
collagenase that degrades the extracellular matrix resulting in the
migration and invasion of tumor cells (10,11).
Several lines of study have demonstrated that an elevated level of
MMP-2 expression is correlated with the progression of bladder
cancer, characterized by migration and invasion (12–14).
In addition, high levels of MMP-2 expression have been detected in
the serum and urine of patients with bladder cancer (12–14).
The MMP-2 promoter contains several functional cis-elements
including p53, AP-1, Ets-1, C/EBP, CREB, PEA3, Sp1, ATF2 and AP-2
that are involved in MMP-2 regulation (15).
MicroRNAs (miRs) are a class of small non-coding RNA
molecules that negatively regulate gene expression by controlling
either translational repression or mRNA degradation, which depends
on the degree of partial or perfect complementarity to the 3′
untranslated regions of their mRNAs (16). Cumulative studies have shown that
miRs reveal diverse biological and pathological functions including
the control of cellular proliferation, migration, differentiation
and apoptosis (16–18). Recent evidence has identified
several miRs as oncogenes or tumor suppressors (16–18).
Previous studies have shown that microRNA-20b (miR-20b) is
upregulated in human breast cancer, c-Myc-induced mouse mammary
tumors and ionizing radiation-induced rat mammary gland tissues
(19–21). In addition, several studies have
demonstrated that miR-20b serves as a survival and oncogenic factor
(19–23). Numerous studies have suggested that
miR-20b plays important roles in oxygen supply, PTEN inhibition,
VEGF regulation by HIF-1α and STAT3 and in transcriptional control
by early growth response-1 (19,21–23).
miR-20b has also been reported as a suppressive factor for the Th17
differentiation and pathogenesis of multiple sclerosis and
experimental autoimmune encephalomyelitis (24). However, little is known concerning
the suppressive effects of miR-20b on cell cycle-regulated
proliferation as well as on MMP-2-mediated migration and invasion
in cancer cells.
In the present study, we report the novel molecular
mechanisms involved in the inhibition of proliferation, migration
and invasion by miR-20b in bladder cancer EJ cells. The present
study is the first report concerning the role of miR-20b in its
function as a tumor suppressor in cancer cells. Based on
miRNA-target prediction analyses, our results demonstrated that
miR-20b inhibited the proliferation of EJ cells and induced
p21WAF1-mediated G1 phase cell cycle arrest. In
addition, miR-20b inhibited the migration and invasion of EJ cells
via decreased MMP-2 expression by downregulating the activation of
Sp-1.
Materials and methods
Materials
Polyclonal antibodies to cyclin E, CDK2, CDK6,
cyclin D1, p53, p21WAF1, p27KIP1 and GAPDH
were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
The polyclonal MMP-2 antibody was obtained from Chemicon
International (Billerica, MA, USA). miR-20b
(5′-CAAAGUGCUCAUAGUGCAGGUAG-3′) and miR-20b inhibitor were designed
and synthesized by Genolution (Seoul, Korea).
Cell cultures
Human bladder carcinoma cell lines (EJ, 5637 and
T24) were purchased from the American Type Culture Collection
(ATCC; Manassas, VA, USA). The cells were maintained in Dulbecco's
modified Eagle's medium (DMEM) (4.5 g glucose/liter) supplemented
with 10% fetal calf serum, L-glutamine and antibiotics (Biological
Industries, Beit Haemek, Israel) at 37°C in a 5% CO2
humidified incubator. Normal human urothelial cells (HUCs) were
obtained from ScienCell Research Laboratories (Carlsbad, CA, USA).
The cells were cultured in urothelial cell medium supplemented with
urothelial cell growth supplement and penicillin/streptomycin
solution according to the protocol of the manufacturer.
Quantitative real-time RT-PCR
(qRT-PCR)
To quantify miRNA expression, real-time PCR
amplification was subjected to a Rotor-Gene™ 6000, as previously
described (25). Real-time PCR
assays were carried out in microreaction tubes (Corbett Research,
Mortlake, Australia) using a miScript PCR Starter kit (Qiagen
Korea, Seoul, Korea). For amplification of the target miRNAs,
forward primers for miR-20b (5′-CAAAGUGCUCAUAGUGCAGGUAG-3′) were
designed. The PCR reaction was performed in a final volume of 20
μl (10 μl 2X QuantiTect SYBR-Green PCR Master Mix, 2
μl 10X miScript Universal Primer, 2 μl 10 pmol
forward primer, 2 μl template cDNA and RNase-free water).
Real-time PCR conditions were as follows: one cycle of initial
activation for 15 min at 95°C, followed by 50 cycles of 15 sec at
94°C for denaturation, annealing for 30 sec at 55°C and extension
for 30 sec at 70°C. The melting program was performed at 70–99°C at
a heating rate of 1°C/5 sec. Spectral data were captured and
determined using Rotor-Gene Real-Time Analysis Software 6.0 Build
14. All of the reactions were carried out in triplicate. In this
experiment, U6 was used as a control for the normalization of the
real-time PCR results in miRNA quantification studies using the
miScript PCR system.
Bioinformatics analysis
In order to identify the potential targets of
miR-20b, we used the miRanda (http://www.microrna.org/microrna/home.do) algorithm to
search the human genome based on an NCBI mRNA database (http://www.ncbi.nlm.nih.gov/) for signaling pathway
annotation and putative functional annotation.
Cell proliferation
Cell proliferation was analyzed using a modification
of the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphe-nyltetrazolium
bromide (MTT) assay, which is based on the conversion of
tetrazolium salt
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2-tetrazolium
to a formazan product by mitochondrial dehydrogenase (26). The formazan product was determined
by quantifying its absorbance at 490 nm. The morphology of cell
proliferation was photographed using phase-contrast microscopy.
Transfection
Cells were transfected with miR-20b and the miR-20b
inhibitor using Lipofectamine 2000 transfection reagent according
to the manufacturer's protocol (Invitrogen). After transfection for
the indicated times, the cells were studied via MTT assay,
immunoblotting, zymography, electrophoretic mobility shift assay
(EMSA), and invasion and wound-healing migration assays.
Cell cycle analysis via
fluorescence-activated cell sorter (FACS)
Cells were harvested and fixed in 70% ethanol. After
washing the cells with ice-cold phosphate-buffered saline (PBS),
they were incubated with RNase (1 mg/ml), DNA intercalating dye and
propidium iodide (50 mg/ml). The analysis of cell cycle
distribution was determined by a Becton-Dickinson FACStar flow
cytometer equipped with Becton-Dickinson Cell Fit software.
Immunoblot analysis
After transfection for 48 h, the cells were rinsed
twice with ice-cold PBS and freeze-thawed in 250 μl lysis
buffer [containing, in mmol/l, HEPES (pH 7.5) 50, NaCl 150, EDTA 1,
EGTA 2.5, DTT 1, β-glycerophosphate 10, NaF 1,
Na3VO4 0.1 and phenylmethylsulfonyl fluoride
0.1 and 10% glycerol, 0.1% Tween-20, 10 μg/ml of leupeptin
and 2 μg/ml of aprotinin], and then scraped into 1.5-ml
tubes. The lysates were then centrifuged at 12,000 rpm for 20 min
at 4°C. The protein concentration of the supernatant was quantified
using a Bradford reagent method (Bio-Rad). Cellular proteins were
electrophoresed on a 0.1% SDS-10% polyacrylamide gel (SDS-PAGE)
under denaturing conditions and transferred to nitrocellulose
membranes (Hybond, Amersham Corp.). The blots were blocked in 10
mmol/l Tris-HCl (pH 8.0), 150 mmol/l NaCl and 5% (wt/vol) non-fat
dry milk and then the membranes were incubated with primary
antibodies at 4°C overnight, which was followed by incubation with
peroxidase-conjugated secondary antibodies for 90 min, followed by
visualization of the immunocomplexes using a chemiluminescence
reagent kit (Amersham Corp.). The experiments were repeated at
least 3 times.
Immunoprecipitation and immune complex
kinase assays
Cell lysates were prepared using an ice-cold lysis
buffer [containing, in mM/l, HEPES (pH 7.5) 50, NaCl 150, EDTA 1,
EGTA 2.5, DTT 1, β-glycerophosphate 10, NaF 1,
Na3VO4 0.1 and phenylmethylsulfonyl fluoride
0.1 and 10% glycerol, 0.1% Tween-20, 10 μg/ml of leupeptin
and 2 μg/ml of aprotinin) and were sonicated at 4°C [Micro
Ultrasonic cell disrupter (from Kontes], at 30% power, 2 times for
10 sec, each time. After centrifugation at 10,000 × g for 5 min,
the lysates were clarified and the supernatants were precipitated
by adding protein A-Sepharose beads precoated with saturating
amounts of the indicated antibodies at 4°C for 2 h. When monoclonal
antibodies were used, protein A-Sepharose was pre-incubated with
rabbit anti-mouse immunoglobulin G (Jackson ImmunoResearch
Laboratories). The immunoprecipitated proteins on the beads were
washed 4 times with 1 ml of lysis buffer and twice with a kinase
buffer (containing, in mM/l, HEPES 50, MgCl2 10, DTT 1,
β-glycerophosphate 10, NaF 1 and sodium orthovanadate 0.1). The
final pellet was re-suspended in 25 μl of kinase buffer
containing either 1 μg of glutathione S-transferase
(GST)-pRb C-terminal (pRb amino acids 769–921) fusion protein
(Santa Cruz Biotechnology), or 5 μg of histone H1
(Life Technologies, Inc.), 20 μM/l ATP and 5 μCi of
[γ32P] ATP (4,500 μCi/mmol; ICN), and were then
incubated for 20 min at 30°C with occasional mixing. Twenty-five
microliters of 2X concentrated Laemmli sample buffer was added to
terminate the reaction, which was then resolved on either 10 or
12.5% SDS-polyacrylamide gels. The migration of histone
H1 or GST-pRb was determined using Coomassie blue
staining. Phosphorylated pRb and histone H1 were
visualized (27).
Wound-healing migration assay
Cells (3×105) were seeded into 6-well
plates in 2 ml medium and were grown to 90% confluency. A clear
area was damaged with a 2-mm-wide tip. After 3 washes with PBS, the
plate was incubated at 37°C in serum-free medium. The migration of
cells into the wounded area was analyzed and photographed using an
inverted microscope (magnification, ×40).
Invasion assay
Invasion assays were performed using an invasion
assay kit (Cell Biolabs, USA) according to the manufacturer's
instructions. Cells (2.5×104) were re-suspended in
serum-free medium and plated in the upper chamber. Medium with 10%
FBS was added to the lower portion of the chamber as a
chemoattractant. After 24 h of incubation, the cells were allowed
to pass through a polycarbonate membrane bearing 8-μm sized
pores with a thin layer of an ECM matrix-like material. The cells
on the lower surface of the membrane were stained and photographed.
The invasive ability of the cells was evaluated using a commercial
cell invasion assay kit (Chemicon International).
Zymography
Culture supernatants were resolved in a
polyacrylamide gel containing 1 mg/ml gelatin. The gel was then
washed with 2.5% Triton X-100 at room temperature for 2 h, followed
by incubation at 37°C overnight in a buffer containing 10 mM
CaCl2, 150 mM NaCl and 50 mM Tris-HCl, pH 7.5. The gel
was stained using 0.2% Coomassie blue and photographed on a light
box. Areas of gelatinase activity were determined as a clear band
in a dark blue field.
Nuclear extracts and EMSA
After centrifugation, the cell pellets were
re-suspended in 1 ml of buffer A containing 10 mM HEPES (pH 7.9),
10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM DTT and 0.5 mM PMSF. They
were then incubated on ice for 15 min and vortexed in the presence
of 0.5% Nonidet NP-40. The pellets were centrifuged and extracted
in 1 ml of buffer B containing 20 mM HEPES pH 7.9, 0.4 M NaCl, 1 mM
EDTA, 1 mM EGTA, 1 mM DTT and 1 mM PMSF for 15 min at 4°C. Protein
concentrations were measured via the Bio-Rad protein assay.
The nuclear extract (10–20 μg) was
pre-incubated at 4°C for 30 min with the 100-fold excess of an
unlabeled oligonucleotide spanning the MMP-2 cis-element of
interest. The sequences were as follows: Sp-1,
GCCCATTCCTTCCGCCCCCAGATGAAGCAG; ATF2, GATCCAGCTTGATGACGTCAGCCG. The
reaction mixture was then further incubated at 4°C for 20 min in a
buffer (25 mM HEPES buffer pH 7.9, 0.5 mM EDTA, 0.5 mM DTT, 0.05 M
NaCl and 2.5% glycerol) with 2 μg of poly(dI-dC) and 5 fmol
(2×104 cpm) of a Klenow end-labeled (32P-ATP)
30-mer oligonucleotide, which corresponded to the DNA binding site
in the MMP-2 promoter. The reaction mixture was electrophoresed at
4°C on a 6% polyacrylamide gel using a TBE (89 mM Tris, 89 mM boric
acid and 1 mM EDTA) running buffer. The gel was washed dried and
exposed to X-ray film overnight (27).
Statistical analysis
Where appropriate, data are expressed as the mean ±
SE. Data were evaluated by factorial ANOVA and a Fisher's least
significant difference test where appropriate. Statistical
significance was set at P<0.05.
Results
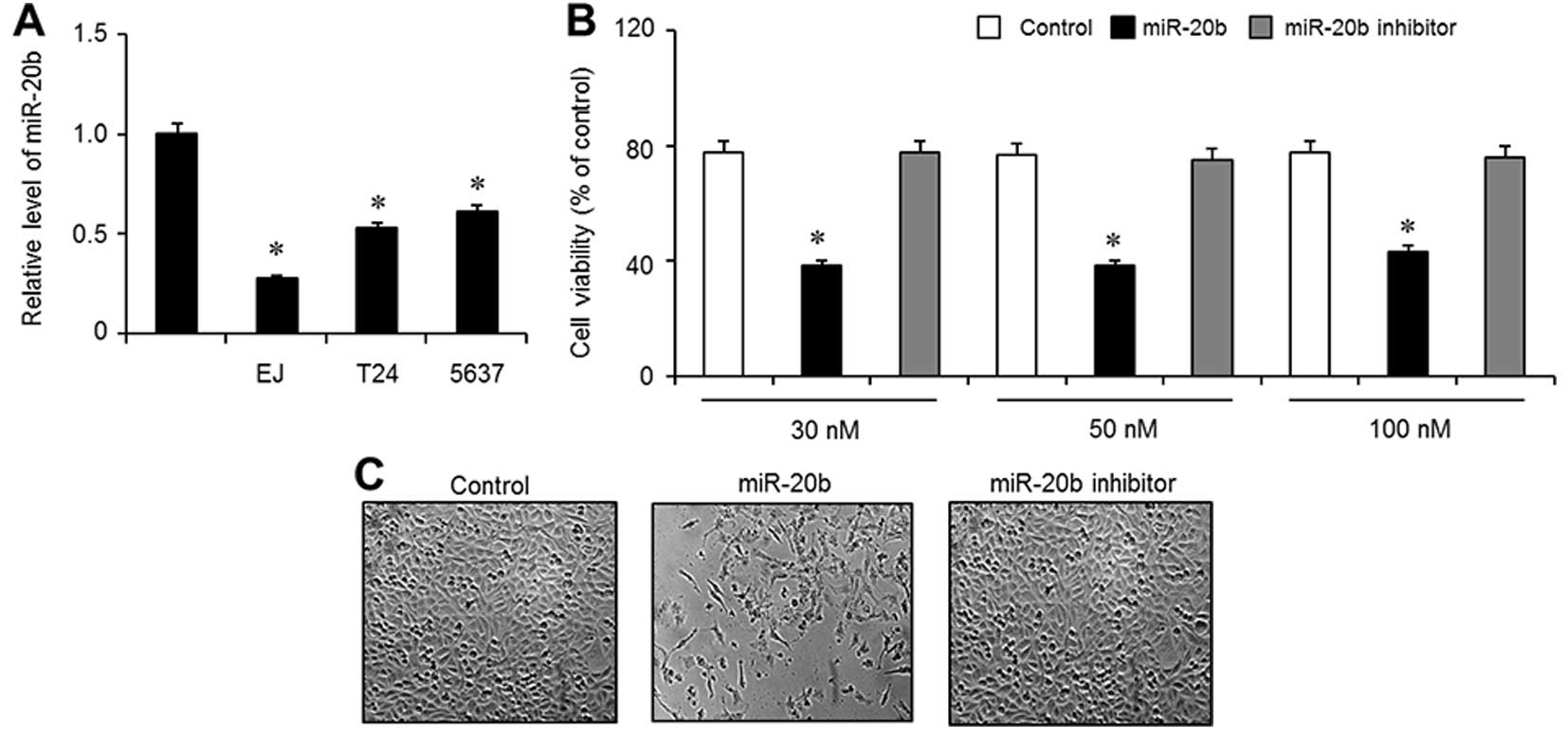
Expression of miR-20b is downregulated in
bladder cancer cell lines
To examine the expression level of miR-20b in
bladder cancer cell lines, 3 types of bladder cancer cell lines
(EJ, T24 and 5637) and normal HUCs were employed using quantitative
real-time PCR. As shown in Fig. 1A,
compared with the HUCs, the expression levels of miR-20b were
significantly downregulated in all 3 examined bladder cancer cell
lines (EJ, T24 and 5637). Since miR-20b expression was decreased in
the EJ cells, we selected this cell line for further investigation.
Next, to investigate the function of miR-20b in the proliferation
of the EJ cells, the cells were transfected with Lipofectamine 2000
only (control), miR-20b mimics (miR-20b) or a negative control
(miR-20b inhibitor) at various concentrations (30–100 nM).
Overexpression of miR-20b significantly reduced the proliferation
of the EJ cells within 48 h, as evidenced by the MTT assay
(Fig. 1B). The different
concentrations of miR-20b (30–100 nM) showed almost the same
antiproliferative effect in the EJ cells (Fig. 1B). Based on this observation, 30 nM
of miR-20b was used for the subsequent experiments. Similar results
were confirmed by phase-contrast images (Fig. 1C).
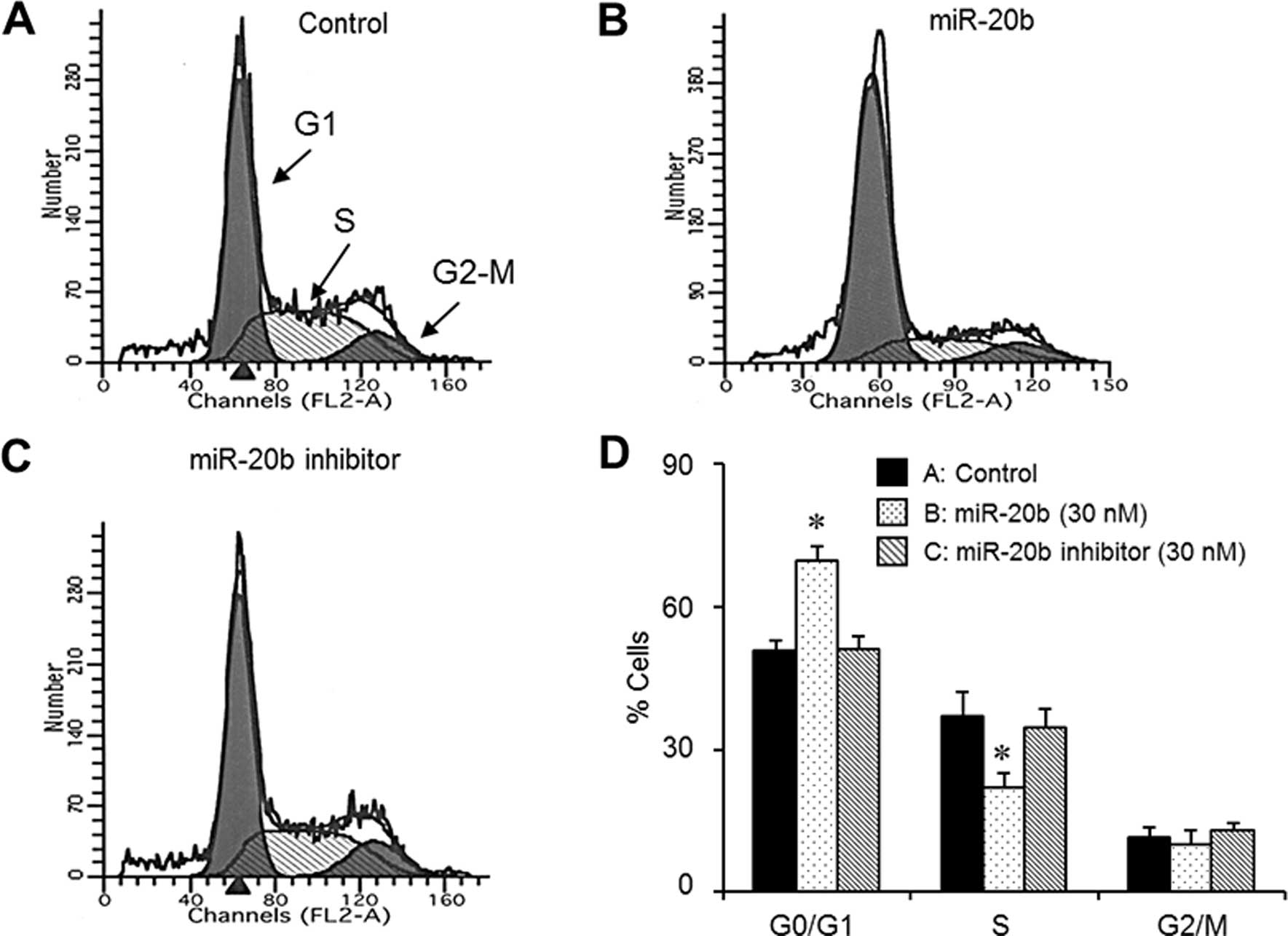
miR-20b induces G1 phase cell cycle
arrest in EJ cells
Flow cytometric analysis was used to investigate
whether the miR-20b-mediated inhibition of cell proliferation was
due to the modulation of the cell cycle. After transfection of
miR-20b (30 nM) into the EJ cells for 48 h, DNA content analysis
was analyzed. Cell cycle distribution analysis demonstrated that
the miR-20b (30 nM)-transfected cells induced a significant
accumulation of cells in the G1 phase (Fig. 2A–D). In addition, the G1 phase cell
cycle arrest was followed by a corresponding decrease in the
population of the S phase population in the presence of miR-20b
(Fig. 2A–D). To verify whether the
observed G1 phase cell cycle arrest of miR-20b transfectants was
involved in the regulation of the cell cycle machinery, the levels
of cell cycle-regulatory molecules were examined. First, we used
bioinformatics analysis from the public miRNA database (miRanda,
http://www.microrna.org/microrna/home.do) to identify
the targets of miR-20b. We found that miR-20b targeted G1 phase
cell cycle regulators, including cyclin D1, CDK2 and CDK6 (Fig. 3E). Immunoblot analysis indicated
that the overexpression of miR-20b resulted in decreased expression
of cyclin D1, CDK2 and CDK6 (Fig.
3A). However, the expression level of cyclin E was not changed
in the miR-20b-transfected cells (Fig.
3A). The kinase activities of the CDKs are an essential step in
the formation of the cyclin/CDK complexes, which result in the cell
cycle progression through sequential checkpoints (8,9).
Therefore, we evaluated the effect of miR-20b on the kinase
activities associated with CDK2 or CDK6. The transfection of
miR-20b into the EJ cells suppressed the kinase activities of both
CDK2 and CDK6 immunoprecipitates (Fig.
3C).
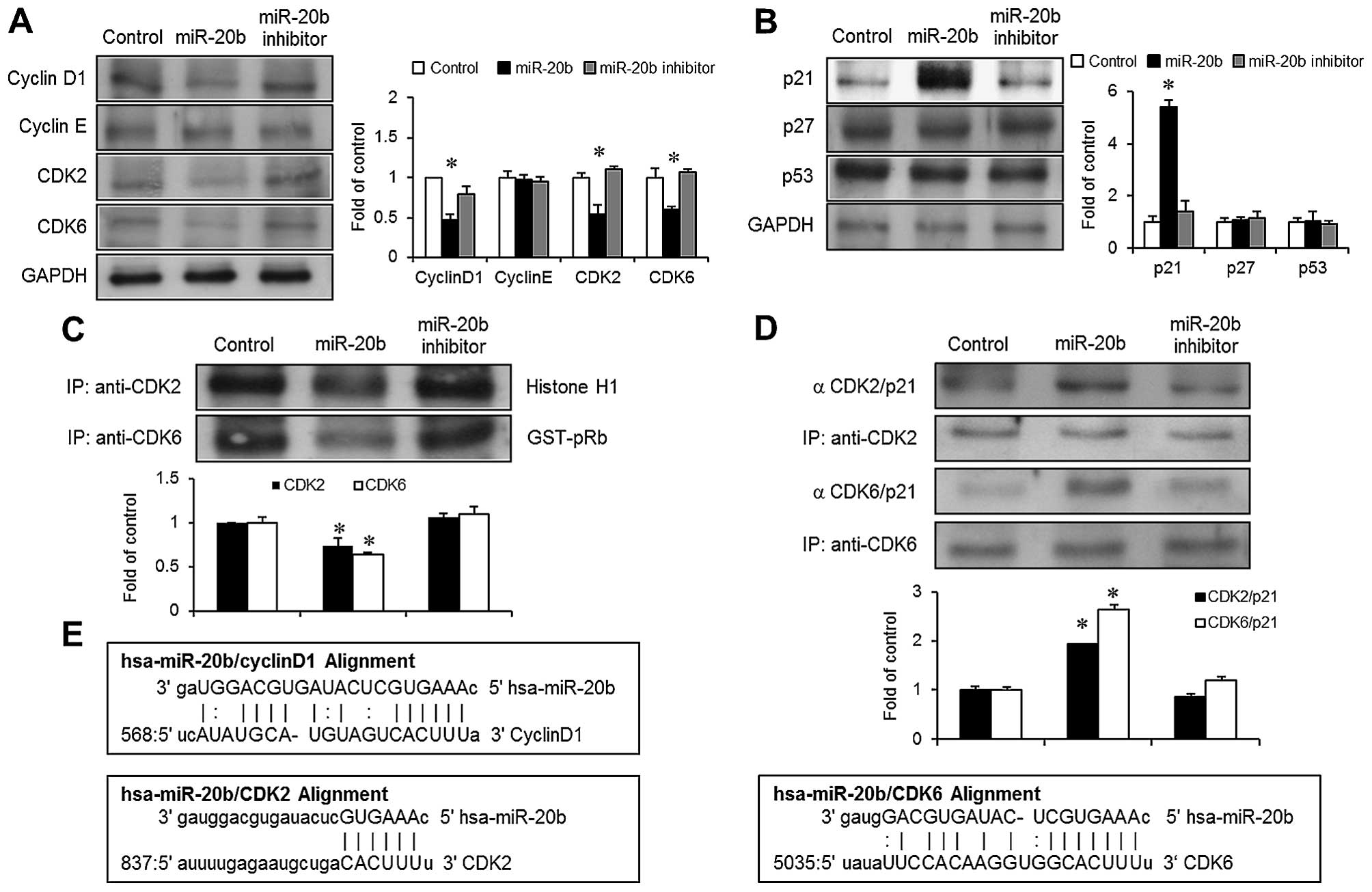 | Figure 3miR-20b overexpression inhibits the
expression of cyclin D1, CDK2 and CDK6, and stimulates the
expression of p21WAF1 in EJ cells. (A and B) After 48 h
of transfection, the cells were harvested for immunoblot analysis
using specific antibodies against cyclin D1, cyclin E, CDK2, CDK6,
p21WAF1, p27KIP1 and p53. GAPDH was used as a
loading control. (C) Cells were transfected with the indicated
genes for 48 h and were then collected. Immunoprecipitation was
performed using anti-CDK2 and anti-CDK6 antibodies on total cell
lysates. The kinase reaction was analyzed using histone H1 (for
CDK2) or GST-pRb (for CDK4) as the substrate. (D) Total cell
lysates were immunoprecipitated with anti-CDK2 and anti-CDK6
antibodies, which were analyzed by immunoblot analysis using an
anti-p21WAF1 antibody. The expression of
immunoprecipitated CDK2 and CDK6 was used as internal proteins. (E)
Diagram of the miR-20b base-pairing site in the 3′-UTR of cyclin
D1, CDK2 and CDK6 mRNA. All the results are expressed as the means
± SE from 3 triplicate experiments. *P<0.01 compared
with the control. |
miR-20b induces the G1 phase cell cycle
through the upregulation of p21WAF1
The effect of miR-20b on CDK inhibitors (CKIs),
which blocks the progression of the G1 to the S phase of the cell
cycle (8) was investigated.
Immunoblot analysis indicated that miR-20b overexpression
stimulated the induction of p21WAF1 in the EJ cells
(Fig. 3B). However, the expression
levels of another CKI p27KIP protein and
tumor-suppressor p53 protein were not altered in the
miR-20b-transfected cells (Fig.
3B). An immunoprecipitation experiment was employed to further
investigate whether the suppression of cyclin/CDK complexes induced
by miR-20b was due to the interaction between p21WAF1
and CDKs. Immunoprecipitated levels of both p21WAF1/CDK2
and p21WAF1/CDK6 were increased in the
miR-20b-transfected cells (Fig.
3D). These results indicated that p21WAF1 plays an
important role in miR-20b-mediated G1 phase cell cycle arrest
inhibiting CDK activities in bladder cancer EJ cells.
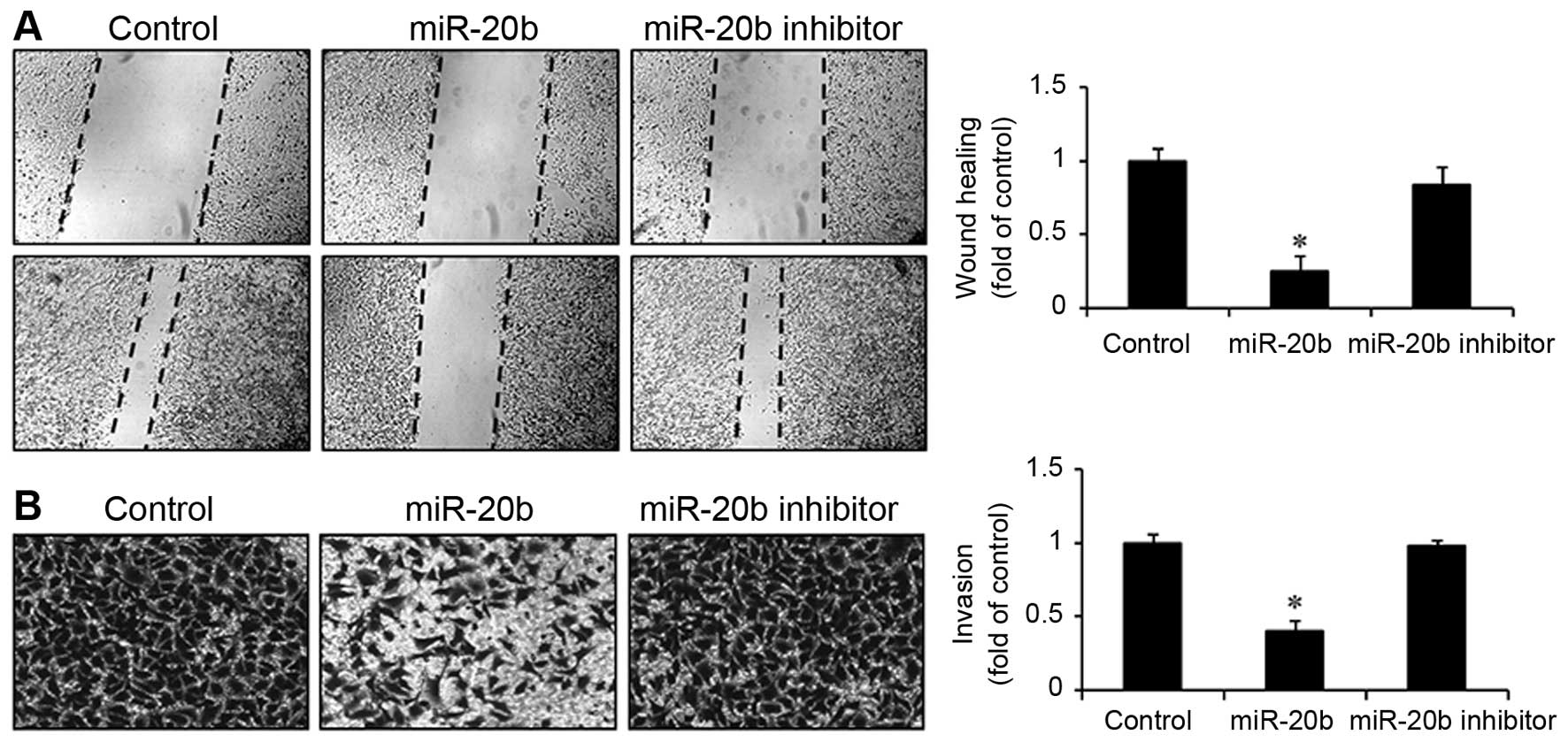
miR-20b inhibits the wound-healing
migration and invasion of EJ cells
Next, we examined the role of miR-20b in the
migration and invasion of bladder cancer EJ cells using a
wound-healing migration assay and a Boyden chamber invasion assay.
As shown in Fig. 4A, the
transfection of miR-20b into EJ cells inhibited cell migration into
the wounded areas within 48 h and caused a significant decrease in
the wound-closure rate compared with both the control and miR-20b
inhibitor transfectants. Consistently, the overexpression of
miR-20b showed a significant reduction in invasiveness through
Matrigel-coated plates by comparison with either the control or
miR-20b inhibitor-transfected cells (Fig. 4B). These results demonstrated that
the overexpression of miR-20b inhibited the migration and invasion
of bladder cancer EJ cells.
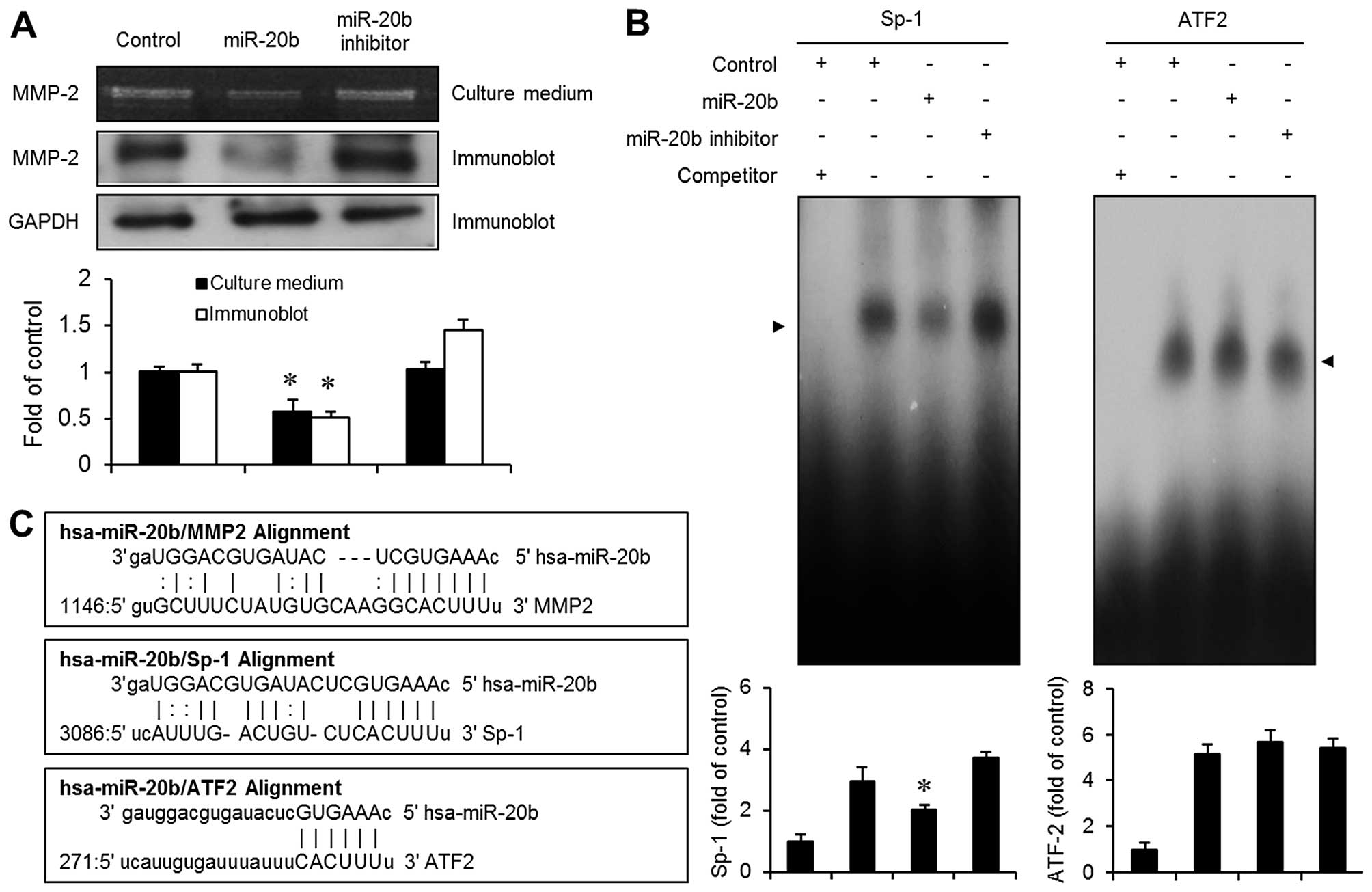
miR-20b transfectants exhibit suppression
of MMP-2 expression via the activation of transcription factor
Sp-1
To identify the potential target genes for the
inhibition of cell migration and invasion induced by miR-20b, we
used miRNA target prediction miRanda algorithms. Since the
expression of MMP-2 is known to be involved in regulating the
migration and invasion of tumor cells (10,11),
we focused on MMP-2 as a potential predicted target gene (Fig. 5C). We next examined the expression
of MMP-2 in the miR-20b-transfected cells. Gelatin zymography assay
showed that the transfection of miR-20b into EJ cells resulted in
an apparent reduction in MMP-2 expression (Fig. 5A). Furthermore, overexpression of
miR-20b markedly inhibited the expression level of MMP-2 protein in
the EJ cells as determined by immunoblot analysis (Fig. 5A). In addition, we found that
miR-20b potentially targets the transcription factors Sp-1 and
ATF2, which are involved in the regulation of MMP-2 in tumor cells
(15), using bioinformatics
analysis of the miRNA target prediction (Fig. 5C). To further investigate whether
the inhibitory effect of miR-20b on MMP-2 expression involves the
potential target genes Sp-1 and ATF2, an EMSA assay was performed.
Nuclear extracts from the miR-20b-transfected cells showed
decreased activation of the Sp-1 binding motif (Fig. 5B). However, no specific binding
activity of ATF2 was observed in the miR-20b transfectants
(Fig. 5B). These results suggested
that the activation of Sp-1 binding is involved in the
miR-20b-induced inhibition of MMP-2 expression in bladder cancer EJ
cells.
Discussion
Transitional cell carcinoma (TCC) is one of the
leading causes of cancer-related mortality worldwide (1,6,7).
Therefore, therapies that can improve the survival of patients with
TCC are desirable. MicroRNAs (miRs) regulate a number of genes
involved in biological processes, such as the proliferation,
migration and apoptosis of tumor cells (16–18).
Therefore, current miR-targeted therapies that impede the function
of tumor-specific molecules have attracted considerable
attention.
It was previously reported that miR-20b was
overexpressed in c-Myc-induced mouse mammary tumors, ionizing
radiation-induced rat mammary gland tissues and human breast cancer
(19–21). Moreover, numerous studies have
identified miR-20b as a factor in tumorigenesis, survival and
oncogenesis (19–23). Although miR-20b has been associated
with the inhibitory effect of Th17 differentiation and immune
diseases such as multiple sclerosis and experimental autoimmune
encephalomyelitis, the tumor suppressor effect of miR-20b on cancer
cells has not previously been studied. In the present study, we
demonstrated the novel role of miR-20b in the proliferation,
migration and invasion of bladder cancer cells.
The results from the present study showed
downregulation of miR-20b expression in bladder cancer cells
compared with normal human urothelial HCC cells. Based on these
results, we postulated that miR-20b plays a critical role in the
proliferation and migration of bladder cancer cells. First, we
initiated a study to ascertain whether miR-20b inhibits the
proliferation of bladder cancer EJ cells when using a low
expression level of miR-20b. Overexpression of miR-20b drastically
inhibited the proliferation of EJ cells, which was attributed to G1
phase cell cycle arrest. To further investigate the exact mechanism
of the miR-20b-induced inhibition of cell proliferation, we used a
predicted target analysis of miRs. The cell cycle regulators,
including cyclin D1, CDK2 and CDK6, were identified as target genes
of miR-20b. We next examined the effect of miR-20b on the levels of
CDKs and cyclins responsible for G1 to S phase progression.
Consistent with the analysis of the miR-20b target genes, the
transfection of miR-20b into EJ cells significantly suppressed the
expression levels of cyclin D1, CDK2 and CDK6, yet not of cyclin E,
followed by inhibition of CDK2 and CDK6 kinase activity. In
addition, the data revealed that p21WAF1 expression was
evidently upregulated during G1 phase cell cycle arrest in the EJ
cells transfected with miR-20b. However, the expression levels of
p27KIP1 and p53 remained unchanged in the presence of
miR-20b. These results suggest that miR-20b inhibited the
proliferation of EJ cells via p21WAF1-mediated G1 phase
cell cycle arrest by suppressing the expression of cyclins and
CDKs. As far as we could ascertain, this is the first integrative
study demonstrating the relevance of each element of the
CKI-cyclin-CDK machinery during miR-20b-induced G1 phase cell cycle
arrest.
The main processes involved in metastatic potential
are known to be the migration and invasion of tumor cells (10,11).
In the present study, we found that miR-20b overexpression
inhibited the migration and invasion of bladder cancer cells. In
fact, it has been suggested that the expression of MMP-2
accelerates the degradation of ECM leading to the migration and
invasion of tumor cells, resulting in the promotion of tumor
metastasis (10–14). Previous studies describing analyses
of the MMP-2 promoter identified several essential functional
elements including p53, AP-1, Ets-1, C/EBP, CREB, PEA3, Sp1, ATF2
and AP-2 sites on astroglioma cell lines (15). Transcription factor Sp1 reportedly
promotes the level of MMP-2 in human tumor cells (15). Although it is well accepted that
MMP-2 facilitates tumor metastasis, little is known concerning the
role of miRs in mediating MMP-2 regulation in tumor cells. Based on
bioinformatic analysis, subsequent experiments prompted us to
examine whether or not MMP-2 is involved in inhibiting the
migration and invasion induced by miR-20b in bladder cancer EJ
cells. In the present study, we demonstrated that the transfection
of miR-20b into EJ cells inhibited the expression of MMP-2. We also
identified Sp-1 as the potential functional cis-element of
miR-20b-induced suppression of MMP-2 expression in the bladder
cancer cells. The data showed that miR-20b inhibited MMP-2
expression via a reduction in Sp-1 binding activation.
Bioinformatic analysis identified several putative
binding motifs, such as cyclin D1, CDK2, CDK6, MMP-2 and Sp-1,
which regulate miR-20b-inhibited proliferation, migration and
invasion of bladder cancer EJ cells. Our hypothesis was confirmed
by immunoblot and EMSA experiments performed with EJ cells, in
which miR-20b overexpression was negatively correlated with the
expression of cyclin D1, CDK2, CDK6, MMP-2 and Sp-1, with the noted
exception of ATF2. Cumulative studies have demonstrated that a
single miR binds to more than 200 gene targets, and these targets
could play important roles in diverse functions including those
involving the cell cycle, signaling pathways, apoptosis,
transcription factors and receptors (16–18).
Based on predicted-target analysis of miR-20b, our studies revealed
that overexpression of miR-20b directly targets cell cycle
regulators and migration-related factors that result in an apparent
suppression of proliferation, migration and invasion in bladder
cancer cells. These results suggest that target analysis of miRs
may reflect the involvement of several cellular mechanisms such as
proliferation, migration and invasion of tumor cells.
The evidence demonstrates that miR-20b is aberrantly
downregulated in bladder cancer cell lines. miRNA-target prediction
analyses showed several putative binding motifs present in the
miR-20b gene. Consistently, the results of the present study
indicated that miR-20b inhibits the proliferation of bladder cancer
EJ cells via p21WAF1-mediated G1 phase cell cycle arrest
by blocking cyclin/CDK complexes. Furthermore, our data revealed
that miR-20b suppressed the migration and invasion of EJ cells at
least in part through decreasing the MMP-2 expression by targeting
transcription factor Sp-1. The present findings suggest that
miR-20b may be a potential therapeutic target for the treatment of
bladder cancer. Further in vivo study is needed to
investigate the efficacy of miR-20b in the treatment of bladder
cancer.
Acknowledgments
This study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea government
(MSIP) (no. 2014007036).
References
|
1
|
Jemal A, Siegel R, Ward E, Murray T, Xu J
and Thun MJ: Cancer statistics, 2007. CA Cancer J Clin. 57:43–66.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Metts MC, Metts JC, Milito SJ and Thomas
CR Jr: Bladder cancer: A review of diagnosis and management. J Natl
Med Assoc. 92:285–294. 2000.PubMed/NCBI
|
|
3
|
Cohen SM, Shirai T and Steineck G:
Epidemiology and etiology of premalignant and malignant urothelial
changes. Scand J Urol Nephrol Suppl. 205:105–115. 2000. View Article : Google Scholar
|
|
4
|
Vineis P, Talaska G, Malaveille C, Bartsch
H, Martone T, Sithisarankul P and Strickland P: DNA adducts in
urothelial cells: Relationship with biomarkers of exposure to
arylamines and polycyclic aromatic hydrocarbons from tobacco smoke.
Int J Cancer. 65:314–316. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Miyajima A, Nakashima J, Yoshioka K,
Tachibana M, Tazaki H and Murai M: Role of reactive oxygen species
in cis-dichlorodiammineplatinum-induced cytotoxicity on bladder
cancer cells. Br J Cancer. 76:206–210. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Levi F, La Vecchia C, Randimbison L and
Franceschi S: Incidence of infiltrating cancer following
superficial bladder carcinoma. Int J Cancer. 55:419–421. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Black PC and Dinney CP: Bladder cancer
angiogenesis and metastasis - translation from murine model to
clinical trial. Cancer Metastasis Rev. 26:623–634. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sherr CJ and Roberts JM: CDK inhibitors:
Positive and negative regulators of G1-phase progression. Genes
Dev. 13:1501–1512. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sherr CJ: Cancer cell cycles. Science.
274:1672–1677. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Matrisian LM: Metalloproteinases and their
inhibitors in matrix remodeling. Trends Genet. 6:121–125. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liotta LA: Tumor invasion and metastases -
role of the extracellular matrix: Rhoads Memorial Award lecture.
Cancer Res. 46:1–7. 1986.PubMed/NCBI
|
|
12
|
Sier CF, Casetta G, Verheijen JH, Tizzani
A, Agape V, Kos J, Blasi F and Hanemaaijer R: Enhanced urinary
gelatinase activities (matrix metalloproteinases 2 and 9) are
associated with early-stage bladder carcinoma: A comparison with
clinically used tumor markers. Clin Cancer Res. 6:2333–2340.
2000.PubMed/NCBI
|
|
13
|
Gerhards S, Jung K, Koenig F, Daniltchenko
D, Hauptmann S, Schnorr D and Loening SA: Excretion of matrix
metalloproteinases 2 and 9 in urine is associated with a high stage
and grade of bladder carcinoma. Urology. 57:675–679. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kawamura K, Kamiya N, Suyama T, Shimbo M,
Oosumi N, Suzuki H, Ueda T, Tobe T, Igarashi T and Ito H: In situ
gelatinolytic activity correlates with tumor progression and
prognosis in patients with bladder cancer. J Urol. 172:1480–1484.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qin H, Sun Y and Benveniste EN: The
transcription factors Sp1, Sp3, and AP-2 are required for
constitutive matrix metalloproteinase-2 gene expression in
astroglioma cells. J Biol Chem. 274:29130–29137. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar
|
|
18
|
Koturbash I, Zemp FJ, Pogribny I and
Kovalchuk O: Small molecules with big effects: The role of the
microRNAome in cancer and carcinogenesis. Mutat Res. 722:94–105.
2011. View Article : Google Scholar
|
|
19
|
Zhou W, Shi G, Zhang Q, Wu Q, Li B and
Zhang Z: MicroRNA-20b promotes cell growth of breast cancer cells
partly via targeting phosphatase and tensin homologue (PTEN). Cell
Biosci. 4:622014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun Y, Wu J, Wu SH, Thakur A, Bollig A,
Huang Y and Liao DJ: Expression profile of microRNAs in c-Myc
induced mouse mammary tumors. Breast Cancer Res Treat. 118:185–196.
2009. View Article : Google Scholar
|
|
21
|
Li D, Ilnytskyy Y, Kovalchuk A, Khachigian
LM, Bronson RT, Wang B and Kovalchuk O: Crucial role for early
growth response-1 in the transcriptional regulation of miR-20b in
breast cancer. Oncotarget. 4:1373–1387. 2013.PubMed/NCBI
|
|
22
|
Lei Z, Li B, Yang Z, Fang H, Zhang GM,
Feng ZH and Huang B: Regulation of HIF-1alpha and VEGF by miR-20b
tunes tumor cells to adapt to the alteration of oxygen
concentration. PLoS One. 4:e76292009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cascio S, D'Andrea A, Ferla R, Surmacz E,
Gulotta E, Amodeo V, Bazan V, Gebbia N and Russo A: miR-20b
modulates VEGF expression by targeting HIF-1 alpha and STAT3 in
MCF-7 breast cancer cells. J Cell Physiol. 224:242–249.
2010.PubMed/NCBI
|
|
24
|
Zhu E, Wang X, Zheng B, Wang Q, Hao J,
Chen S, Zhao Q, Zhao L, Wu Z and Yin Z: miR-20b suppresses Th17
differentiation and the pathogenesis of experimental autoimmune
encephalomyelitis by targeting RORγt and STAT3. J Immunol.
192:5599–5609. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yun SJ, Jeong P, Kim WT, Kim TH, Lee YS,
Song PH, Choi YH, Kim IY, Moon SK and Kim WJ: Cell-free microRNAs
in urine as diagnostic and prognostic biomarkers of bladder cancer.
Int J Oncol. 41:1871–1878. 2012.PubMed/NCBI
|
|
26
|
Moon SK, Jung SY, Choi YH, Lee YC,
Patterson C and Kim CH: PDTC, metal chelating compound, induces G1
phase cell cycle arrest in vascular smooth muscle cells through
inducing p21Cip1 expression: Involvement of p38 mitogen
activated protein kinase. J Cell Physiol. 198:310–323. 2004.
View Article : Google Scholar
|
|
27
|
Lee SJ, Cho SC, Lee EJ, Kim S, Lee SB, Lim
JH, Choi YH, Kim WJ and Moon SK: Interleukin-20 promotes migration
of bladder cancer cells through extracellular signal-regulated
kinase (ERK)-mediated MMP-9 protein expression leading to nuclear
factor (NF-κB) activation by inducing the up-regulation of
p21WAF1 protein expression. J Biol Chem. 288:5539–5552.
2013. View Article : Google Scholar :
|