Introduction
Glioma, which is refractory to tumor therapies with
high mortality and morbidity worldwide, is the most frequent
intracranial tumor in humans (1,2).
Despite the improvement in treatment technology, the disease still
has a poor prognosis and survival rate (3). Thereby, developing effective treatment
methods is necessary. The lack of sufficient vehicles that
specifically target glioma in the brain limits the development of
effective therapies. In recent years, bone marrow-derived
mesenchymal stem cells (BMSCs) were found to possess selective
tumor-tropic properties, which can migrate throughout the
experimental brain tumors (4).
BMSCs were proposed as a novel cellular vehicles for local delivery
of drugs or biological agents to gliomas due to their specific
glioma tropism (5,6). Therefore, BMSCs are promising gene
vehicles for the treatment of gliomas.
The brain is an immune-privileged site that lacks
lymphatic drainage due to the blood-brain barrier, which
accelerates the development and progression of gliomas.
Immunotherapy for the treatment of glioma has been reported
(7). Interleukin (IL)-18 with a
molecular weight of 18.3 kDa is a member of IL family that
specifically induces Th1 cytokine production, particularly
interferon (IFN)-γ from T cells and natural killer cells (8,9). IL-18
activates cytotoxic T activity, augment T cell proliferation,
enhance natural killer cytolytic activity and promote antigen
presentation (10–12). Furthermore, IL-18 induces the
production of IL-2, which has significant antitumor activity
against experimental and clinical model of glioma (13,14).
Thereby, IL-18 has been proposed as potential immunomodulator for
the treatment of malignant gliomas. IFNs containing IFN-α, IFN-β
and IFN-γ are a family of natural glycoproteins that activate
immune cells, including macrophages, T cells and natural killer
cells augmenting antitumor immunity and inhibiting oncogene
expression and tumor angiogenesis (15–17).
Of these IFNs, IFN-β has been widely studied in the treatment of
tumors. This glycoprotein displays potent antiproliferative
activity against melanoma cells (18). IFN-β also has a direct cytotoxic
effect on glioma cells, and the intratumoral IFN-β delivery
significantly enhances the cytotoxic T and natural killer activity
without any side-effect (19,20).
In summary, BMSCs possess specific glioma-tropic
properties, which are ideal vehicles for gene therapy of gliomas.
Our previous study demonstrated that IL-18-expressing BMSCs
effectively inhibits intracranial glioma in rats in vivo
(21). Considering the vital
function of IFN-β in inhibiting tumor growth, we speculated that
the co-expression of IL-18 and IFN-β displays more potent and
effective antitumor effect against glioma. To test this hypothesis,
we constructed a recombinant lentivirus co-expressing IL-18 and
IFN-β, which was then introduced into BMSCs to engineer the BMSCs
genetically to express both IL-18 and IFN-β. In the present study,
we explored the effect of these engineered BMSCs in glioma
treatment in vitro and in vivo in a rat intracranial
glioma model.
Materials and methods
Experimental animals
Adult male Fisher 344 rats (9–11 weeks old, weighing
180–220 g) were purchased from Gongdong Medical Laboratory Animal
Center (Guangzhou, China) and housed in the animal care facility
under standard protocols. All animal procedures were conducted
according to the institutional and the national guidelines approved
by the Institutional Animal Care and Use Committee of Xi'an
Jiaotong University.
Cell cultures
Rat 9L glioma cells were obtained from American Type
Culture Collection (ATCC; Manassas, VA, USA) and grown in
Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA,
USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen), 2
mM L-glutamine, and 1% streptomycin and penicillin. 293FT cells
(Invitrogen) were maintained in DMEM containing 10% fetal calf
serum and 1% streptomycin and penicillin. Primary rat BMSCs were
isolated and cultured as described in our previous study (21). Fisher 344 rats were sacrificed via
intraperitoneal injection of 10% chloral hydrate (0.4 ml/100 g).
The femurs and tibias were isolated, and the medullary cavity was
flushed with 5 ml of DMEM, followed by density gradient
centrifugation (Ficoll-Paque; BD Biosciences, Lincoln Park, NJ,
USA) at 2,000 rpm for 15 min. The mononuclear cells were then
harvested and resuspended in low glucose DMEM supplemented with 15%
FBS, 2 mM L-glutamine and 1% streptomycin and penicillin. All these
cells were cultured in a humidified incubator containing 5%
CO2 at 37°C.
Recombinant lentivirus construction and
infection
The full length cDNAs of IL-18 and IFN-β were
amplified using pCR3.1-IL-18 (New England Biolabs, Beverly, MA,
USA) and Ad-CMV-IFN-β (provided by the Department of Microbiology,
The Second Military Medical University, Shanghai, China),
respectively. These cDNA fragments were subcloned into the
lentiviral vector pLenti6/V5-DEST to construct the recombinant
vectors pLenti6/V5-DEST-IL-18, pLenti6/V5-DEST-IFN-β or
pLenti6/V5-DEST-IL-18-IFN-β. These vectors were packaged using
ViraPower™ Lentiviral Expression Systems (Invitrogen) according to
the manufacturer's instruction. After transfection with 293FT cells
for 48–72 h, the culture medium was harvested and filtered using a
0.45 µm filter (Amicon Ultra-15 100K; Millipore, Billerica, MA,
USA). These recombinant lentivirus were named LV-IL-18, LV-IFN-β
and LV-IL-18-IFN-β, respectively. For infection, BMSCs were seeded
into 24-well tissue plates at 1×106 cells/well for 24 h.
Subsequently, the lentiviral super-natants were added and incubated
for 24 h. Stable clones were selected with Blasticidin S
(Invitrogen). These genetically engineered BMSCs were named
BMSCs-IL-18, BMSCs-IFN-β and BMSCs-IL-18-IFN-β.
Quantitative real-time PCR (qRT-PCR)
The total RNA extracted using total RNA extraction
system (Qiagen, Dusseldorf, Germany) was reversely transcripted
into cDNA using M-MLV reverse transcriptase (Clontech, Palo Alto,
CA, USA) according to the manufacturer's instruction. qRT-PCR was
performed with SYBR-Green PCR master mix (Invitrogen) on a PTC-100
DNA thermal cycler (MG Research, Waltham, MA, USA).
Glyceraldehyde-3-phosphate dehydro-genase (GAPDH) was used as an
internal reference for the quantification of the relative mRNA
expression.
Western blot analysis
Equal amounts of proteins from different samples
were loaded on 12.5% SDS-PAGE, separated and transferred to
nitrocellulose membranes (Miltenyi Biotec, Auburn, CA, USA). The
membranes were blocked with non-fat milk and then incubated with
primary antibodies at 4°C overnight. Subsequently, secondary
antibodies coupled with horseradish peroxidase (Bioss, Beijing,
China) were added for 1 h at room temperature. The immunoreactive
proteins were detected using an enhanced chemiluminescence reagent
(GE Healthcare, Piscataway, NJ, USA). Primary antibodies, including
anti-IL-18, anti-IFN-β and anti-GAPDH were purchased from Santa
Cruz Biotechnology (Santa Cruz, CA, USA).
Enzyme-linked immunosorbent assay
(ELISA)
The concentrations of IL-18, IFN-β, IFN-γ and IL-2
in the cell cultured medium or tumor tissues were measured using
commercial ELISA kits (R&D Systems, Minneapolis, MN, USA)
according to the manufacturer's instructions.
Cell viability assay
Cell viability was detected by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay. In brief, 9L cells were seeded into 96-well plates
co-cultured with different genetically engineered BMSCs or
collected cell culture medium for 24 and 48 h. After replacement
with fresh medium, 20 µl of MTT [0.5 mg/ml in phosphate-buffered
saline (PBS)] was added per well and incubated for 4 h. The medium
was discarded and dimethyl-sulfoxide (150 µl/well) was added to
melt the formazan for 15 min. Absorbance was then determined at 450
nm.
Intracranial tumor implantation and
treatment
Rat 9L glioma cells were stereotactically implanted
into the rat brain according to previously described methods with
minor modification (22,23). In brief, rats were anesthetized and
placed in a stereotactic frame. The glioma cells (1×106)
resuspended into 10 µl of PBS were stereotactically injected into
the right corpus striatum with a Hamilton syringe at a depth of 5
mm. Three days after 9L cell implantation, the rats were treated
with intratumoral inoculations of 1×106 BMSCs,
BMSCs-IL-18, BMSCs-IFN-β and BMSCs-IL-18-IFN-β resuspended into 10
µl of PBS. The tumor volume was monitored using a 1.5 T MRI system
(General Electric, Syracuse, NY, USA) with 3-inch surface coil
every four days for four weeks. Tumor volumes (mm2) were
calculated as follows: (3/4π x length × width × height) × 1/8.
Immunohistochemical staining
After two weeks of treatment, brain tumor tissues
were removed from the rats, fixed in 4% paraformaldehyde,
dehydrated, frozen and serially cut into 10-µm thick sections using
a cryostat microtome. The sections were then incubated with
anti-CD4 and anti-CD8 monoclonal antibodies (Abcam, Cambridge, UK).
Immunostaining was performed using the avidin-biotin complex method
with DAB color development.
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assay
Brain tumor tissue sections were stained using the
TUNEL apoptosis kit (GenMed Scientifics, Inc., Arlinghton, MA, USA)
according to the manufacturer's instructions. In brief, cells were
fixed with 4% paraformaldehyde and incubated with TUNEL reaction
mixtures at 37°C for 1 h. The stained cells were visualized and
counted under a fluorescence microscope (Olympus, Tokyo, Japan).
The number of apoptotic cells was counted in random fields in a
blinded manner.
Statistical analysis
Data are expressed as means ± standard deviation.
One-way ANOVA was carried with SPSS software package version 11.5
(SPSS, Inc., Chicago, IL, USA) to analyze statistical differences.
The survival curve was calculated with the Kaplan-Meier method.
p<0.05 was considered to indicate a statistically significant
result.
Results
Stable expression of IL-18 and IFN-β in
engineered BMSCs
To detect whether IL-18 and IFN-β were successfully
introduced into BMSCs, we detected their gene expressions in these
engineered BMSCs. The results of qRT-PCR and western blot analysis
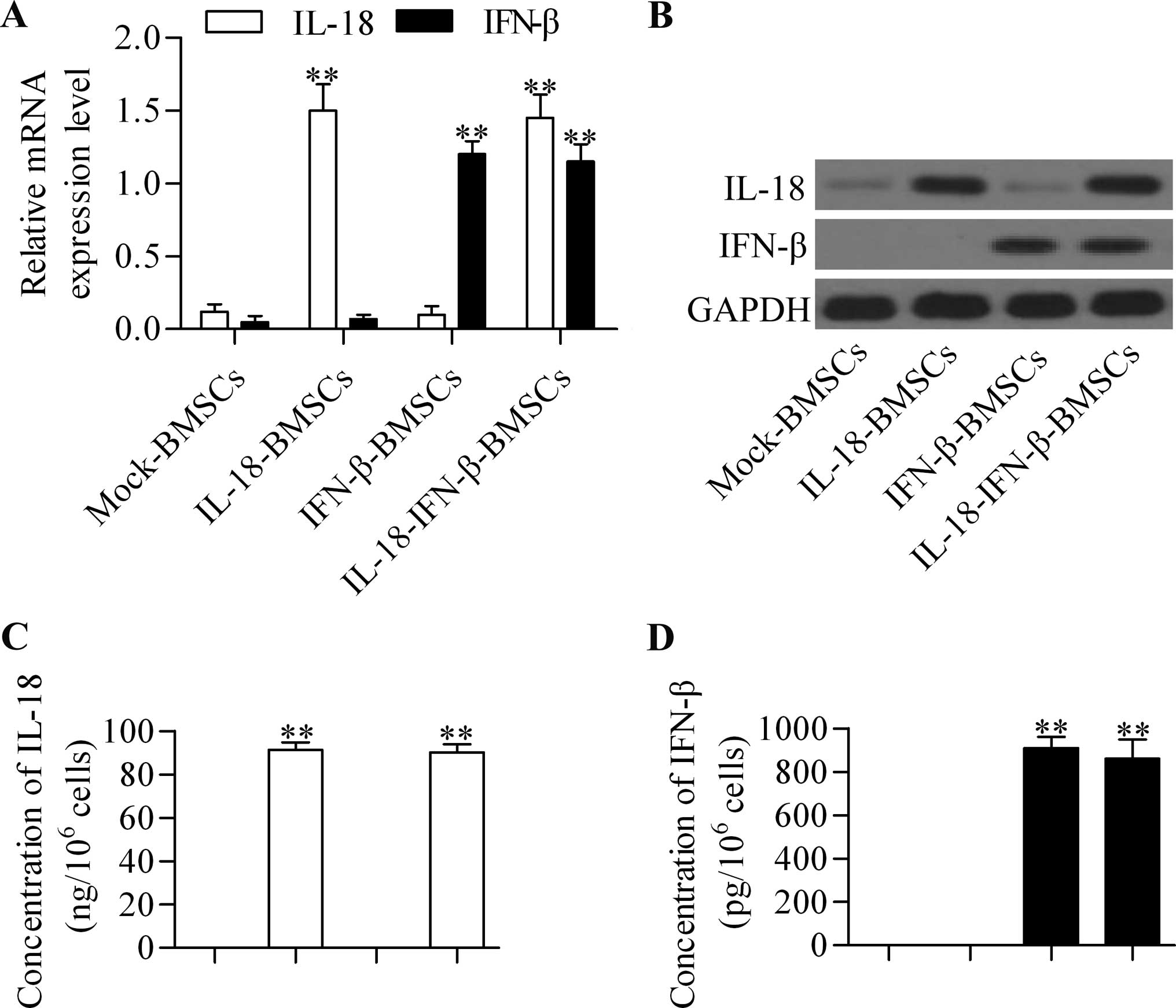
showed that the IL-18 and IFN-β were expressed in IL-18-BMSCs and
IFN-β-BMSCs, respectively, at the mRNA (Fig. 1A) and protein (Fig. 1B) levels. Moreover, co-expression of
IL-18 and IFN-β occurred in IL-18-IFN-β-BMSCs. We also examined the
secretion levels of these proteins in conditioned media by ELISA
method. The results exhibited that soluble IL-18 (Fig. 1C) and IFN-β (Fig. 1D) were detectable in the
supernatants from IL-18-BMSCs and IFN-β-BMSCs, respectively,
whereas both IL-18 and IFN-β were detected in the supernatants from
IL-18-IFN-β-BMSCs. These data indicated that the engineered BMSCs
were successfully established.
IL-18-IFN-β-BMSCs significantly inhibit
the growth of glioma cells in vitro
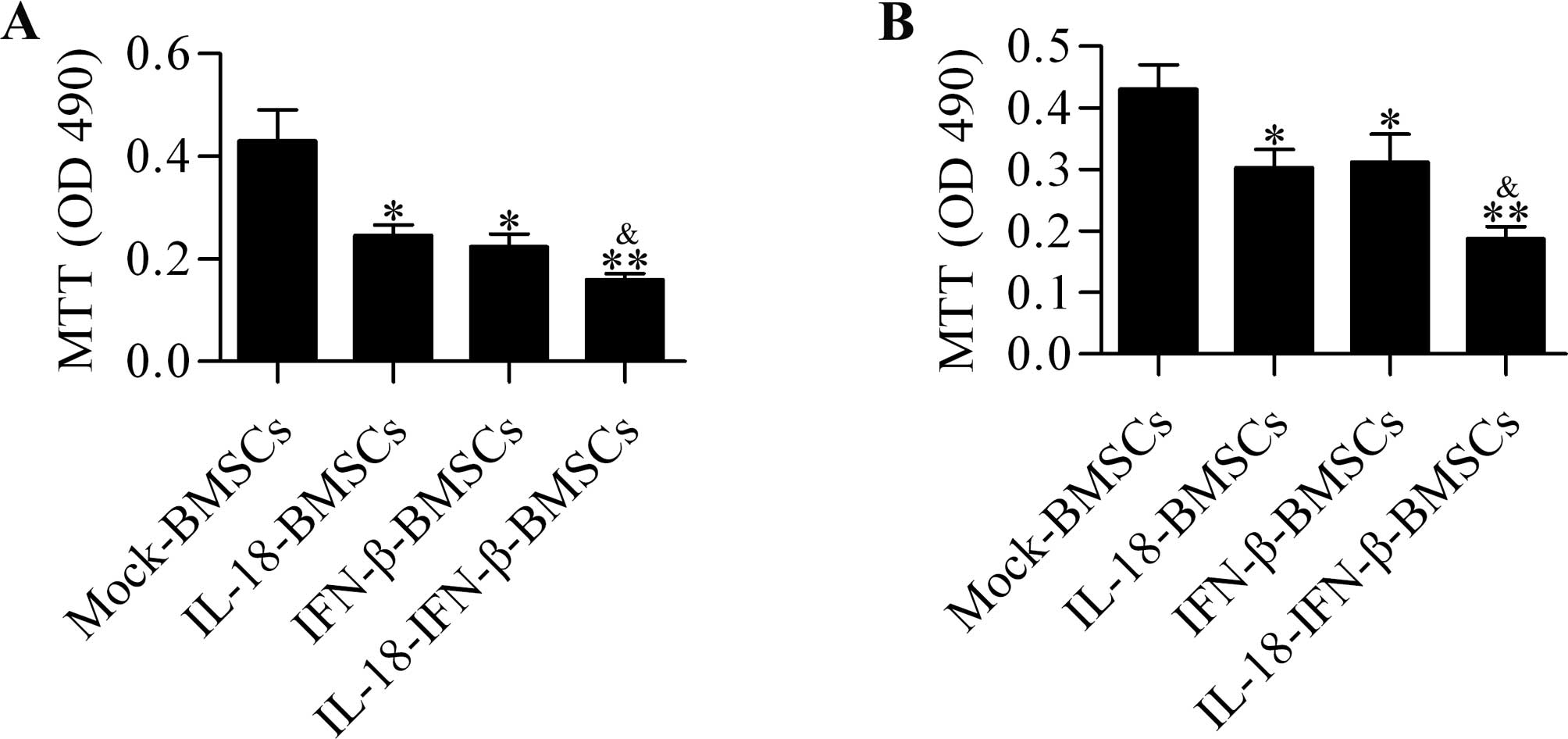
To determine the effect of these engineered BMSCs on
glioma cell growth, we performed the MTT assay. The results showed
that IL-18-BMSCs and IFN-β-BMSCs affected the viability of glioma
cells, but IL-18-IFN-β-BMSCs showed more significant inhibitory
effect compared with IL-18-BMSCs or IFN-β-BMSCs (Fig. 2A). Moreover, conditioned media from
IL-18-IFN-β-BMSCs displayed more potent inhibiting effect on cell
viability than those from IL-18-BMSCs or IFN-β-BMSCs (Fig. 2B). These results indicated that the
IL-18 and IFN-β secreted from these engineered BMSCs were
bioactive.
Treatment with IL-18-IFN-β-BMSCs
significantly prolongs survival and represses intracranial glioma
growth
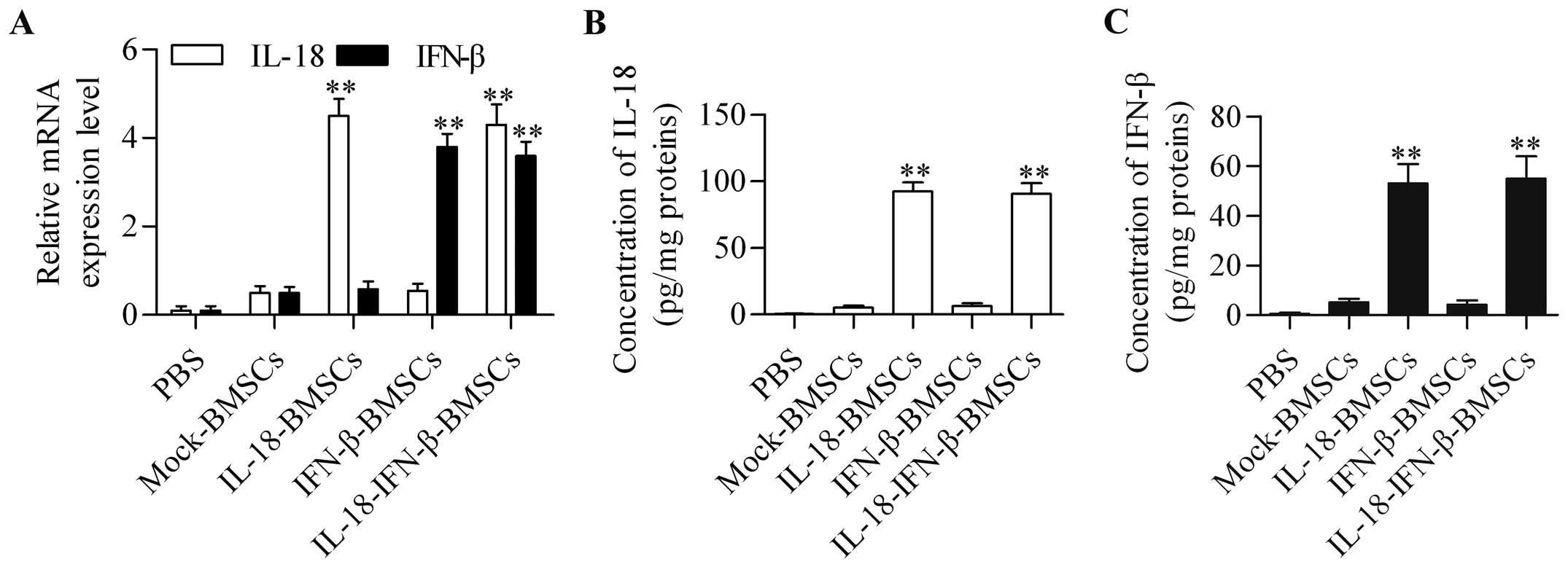
To determine whether IL-18-IFN-β-BMSCs provided an
effective therapeutic benefit against intracranial glioma growth,
these engineered BMSCs were delivered into established intracranial
gliomas in a rat model. The results showed that IL-18-IFN-β-BMSCs
expressed both high mRNA (Fig. 3A)
and secreted protein levels of IL-18 and IFN-β (Fig. 3B and C) in the tumor mass compared
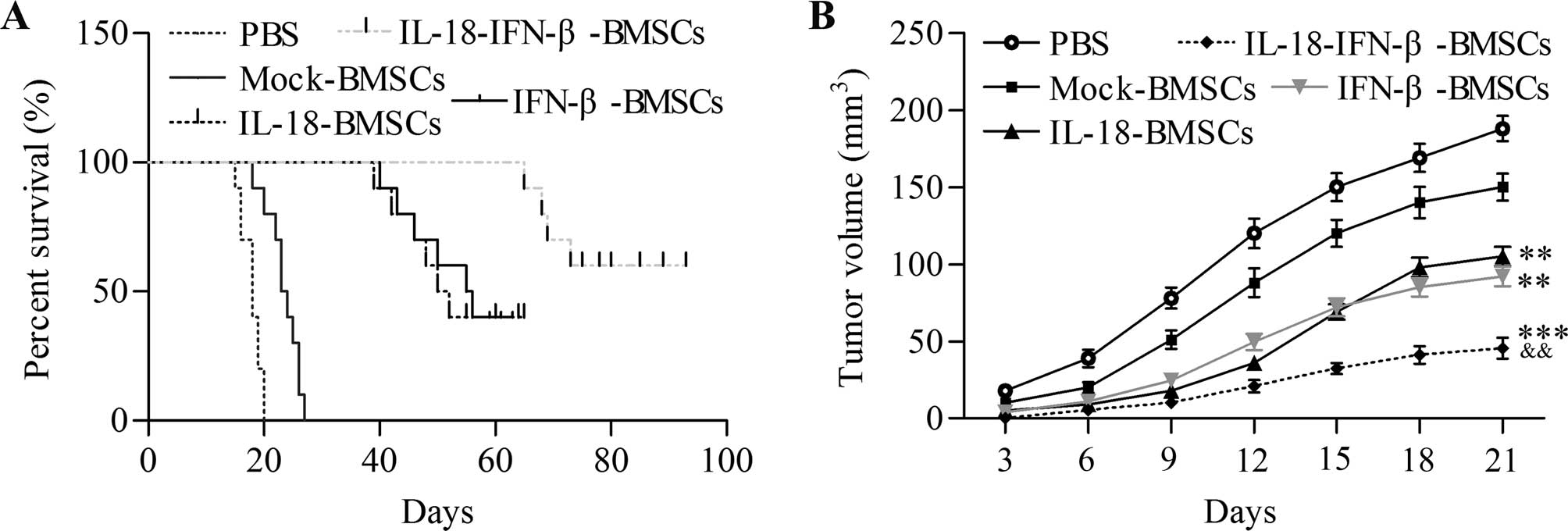
with other groups. We also found that IL-18-IFN-β-BMSC treatment
significantly prolonged the survival compared with IL-18-BMSC- and
IFN-β-BMSC-treated groups (Fig.
4A). Furthermore, their effect on tumor volume was assessed.
IL-18-IFN-β-BMSC treatment showed great inhibition of tumor growth
when compared with other treatment groups (Fig. 4B).
IL-18-IFN-β-BMSCs induce potent glioma
cell apoptosis
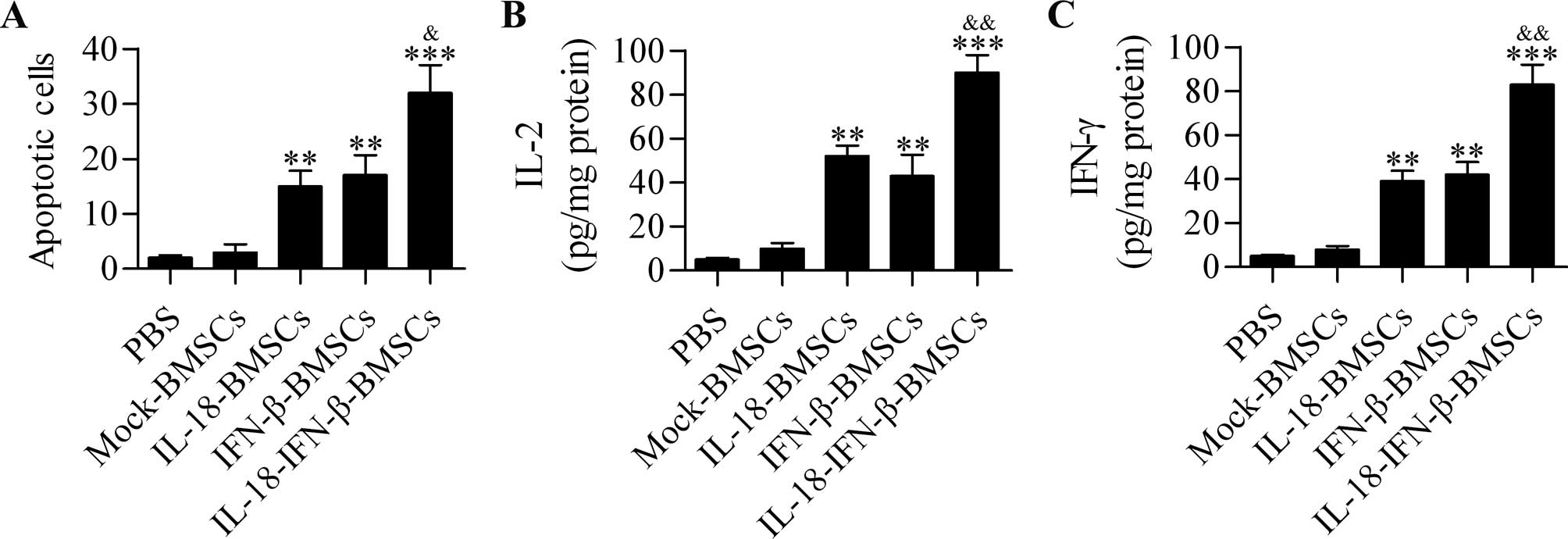
To assess apoptosis induction by different
engineered BMSCs, apoptotic cells in tumors were identified by the
TUNEL assay after treatment. The quantitative data indicated that
IL-18-BMSC and IFN-β-BMSC treatments induced considerable cell
apoptosis compared with the control group, whereas
IL-18-IFN-β-BMSCs induced tumor cell apoptosis more potently than
the other groups (Fig. 5A).
IL-18-IFN-β-BMSCs induce strong
production of IL-2 and IFN-γ
To further verify that IL-18-IFN-β-BMSCs potentiated
strong antitumor effect, we detected the expression of IL-2 and
IFN-γ, which were closely related producing anti-tumor effect
(24) in tumor tissues with ELISA
method. The tumor tissues from IL-18-BMSC- or IFN-β-BMSC-treated
rats displayed higher levels of IL-2 and IFN-γ than those from the
control group. Significant differences were also observed between
rats treated with IL-18-IFN-β-BMSCs and IL-18-BMSCs or IFN-β-BMSCs
(Fig. 5B and C).
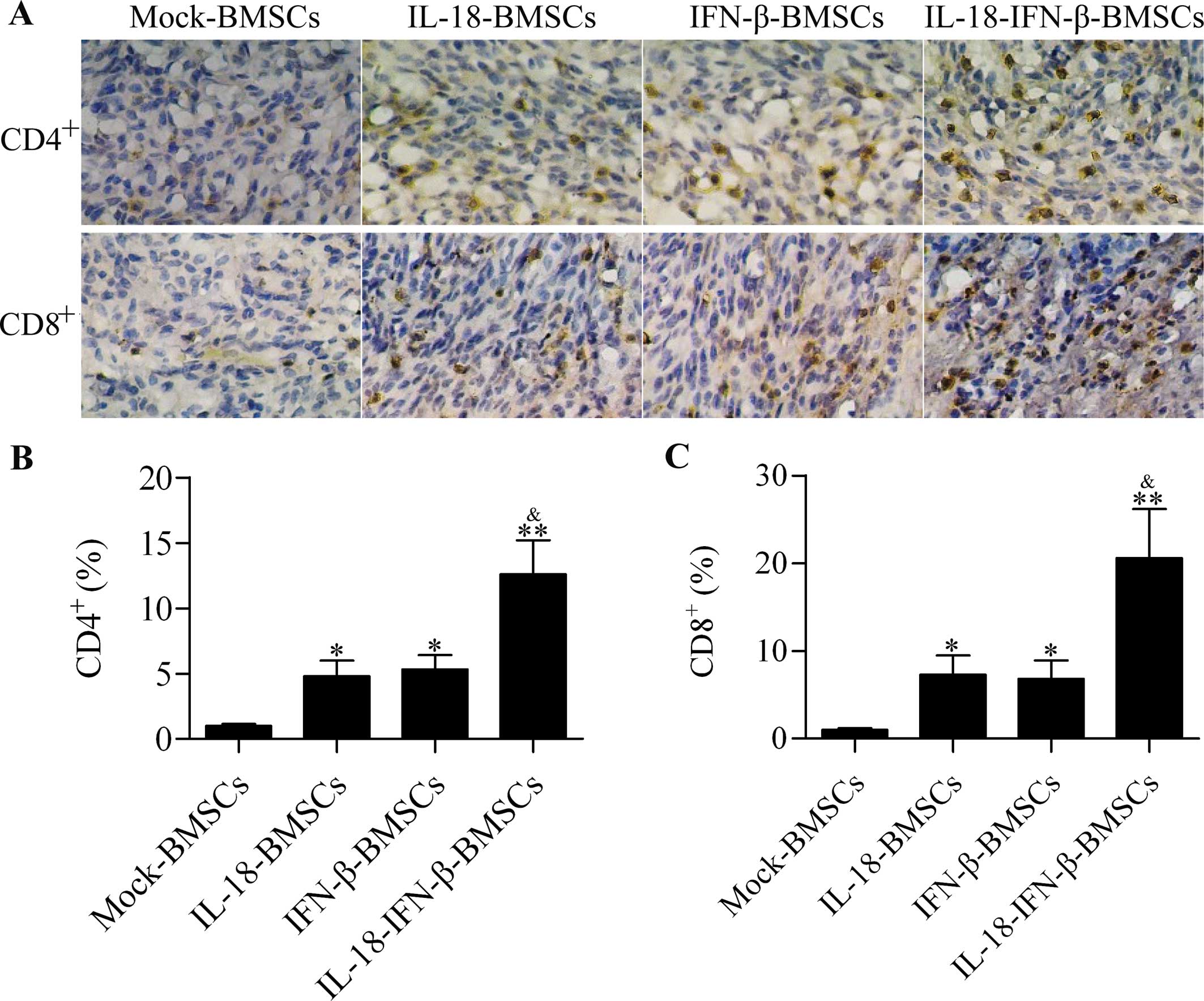
IL-18-IFN-β-BMSCs induces enhanced
CD4+ and CD8+ T-cell infiltration in
intracranial glioma
Tumor-infiltrating T cells in glioma patient are
beneficial for prolonging survival (25,26).
Negligible CD4+ and CD8+ T-cell infiltrations
were observed in mock-BMSC-treated groups. A certain degree of
tumor-infiltrating CD4+ and CD8+ T cells was
also found in animals receiving IL-18-BMSCs or IFN-β-BMSCs
treatment. IL-18-IFN-β-BMSC treatments induced more robust
CD4+ and CD8+ T-cell infiltration in
intracranial glioma than the other groups (Fig. 6A–C).
Discussion
The use of BMSCs as a gene transfer vehicle in the
treatment of BMSCs has been extensively investigated (27). BMSCs expressing herpes simplex
virus-thymidine kinase suppress glioma tumor growth in a mouse
model (28). BMSCs expressing
soluble tumor necrosis factor-related apoptosis-inducing ligand
(TRAIL) induce substantial tumor cell apoptosis in intracra-nial
glioma in a mouse model and significantly improve the survival rate
(29). Moreover, TRAIL-engineered
BMSCs exhibit cytotoxic effects on glioma cells in vitro
(30). In the present study, we
demonstrated that BMSCs co-expressing IL-18 and IFN-β displayed a
more potent antitumor effect than BMSCs solely expressing IL-18 or
IFN-β. This result implies a significant synergistic effect of
IL-18 and IFN-β in glioma treatment. The present study, not only
confirmed that BMSCs served as a powerful delivery vehicle for
glioma treatment, but also provided novel insight for the
improvement of glioma gene therapy using BMSCs.
Overexpression of IL-18 induces cell apoptosis of
human tongue squamous cell carcinoma cells (31) by regulating glycogen synthase
kinase-3β signaling (32).
Adenovirus-mediated expression of IL-18 induces significant cell
apoptosis and inhibits angiogenesis against melanoma in mice
(33). In addition, IFN-β inhibits
the proliferation and angiogenesis of malignant glioma cells
(34) and displays potent
antiproliferative activity against melanoma and glioma cells
(18,35,36).
In the present study, we found that IL-18-BMSCs or IFN-β-BMSCs
successfully expressed IL-18 or IFN-β and significantly inhibited
the cell growth of glioma cells. Furthermore, the condition medium
from these cultured cells also inhibited cell growth of glioma
cells, thus implying that the secreted IL-18 or IFN-β from
engineered BMSCs was bioactive. Surprisingly, IL-18-IFN-β-BMSCs
co-expressing IL-18 or IFN-β exhibited a more significant growth
inhibition on glioma cells than those solely expressing BMSCs.
These data suggest that combined IL-18 and IFN-β treatment may have
potent antitumor effect on glioma. Similarly, co-expression of
IL-18 and Fas enhanced the apoptosis of glioma cells (37). Thereby, the combined utilization of
antitumor drugs or biological agents may produce more satisfactory
antitumor effect.
The sole use of IL-18 or IFN-β as immunostimulatory
cytokine in the treatment of various types of cancers, including
gliomas, has been widely studied reporting that IL-18 is a powerful
adjuvant for immunotherapy and vaccine therapy and is safe for
clinical use (38,39). Intracerebral injection of
recombinant IL-18 inhibits the gliomas located in the mouse brain
(40), whereas intramuscular
injection promotes antigen-specific immune responses mediated by
CD4+/CD8+ T cells against brain tumors
(41). The glioma cells
co-expression of IL-18 and Fas shows reduced tumorigenicity
(37). Dendritic cells loaded with
tumor lysates and IL-18 elicit CD8+ cytotoxic T cell
response against glioma in patients (42). Similarly, dendritic cells that are
genetically engineered to express IL-18 combined with the
administration of IL-12 significantly induce tumor-specific
CD4+/CD8+ T cells and natural killer cells
against glioma cells (43).
Treatment of adeno-associated virus encoding human IFN-β completely
prevents tumor growth and improves the survival of glioma mouse
model (44,45). Co-administration of IFN-β and
dendritic cells accelerates cell apoptosis and induces specific
T-cell responses against glioma cell (16). The intratumoral injection of
liposomes containing IFN-β reduces tumor growth and induces massive
T cell infiltration within the residual tumor (19). To date, the gene transfer of IL-18
or IFN-β using BMSCs has rarely been investigated. In our previous
study, we demonstrated that adenoviral-mediated IL-18 expression in
BMSCs effectively suppressed the growth of glioma in rats (21), thereby providing a promising new
treatment option for malignant glioma. In the present study, we
investigated the treatment effect of the BMSCs co-expression of
IL-18 or IFN-β on glioma. The data demonstrated that this
co-expression displayed more signifi-cant inhibition effect on
glioma, including prolonged survival and reduced tumor growth in
intracranial glioma in rats, compared with BMSCs solely expressing
IL-18 or IFN-β. The underlying mechanism may be that IL-18 and
IFN-β synergistically promoted antitumor immunity, in which
antitumor cytokines IL-2 and IFN-γ production and CD4+
and CD8+ T-cell infiltration in intracranial glioma were
significantly promoted.
The present study is the first to investigate the
effect of BMSCs that were genetically engineered with IL-18 and
IFN-β for the treatment of experimental glioma in a rat model. We
found that BMSCs co-expressing IL-18 and IFN-β significantly
induced cell apoptosis, prolonged survival and reduced tumor growth
of intracranial glioma. Their co-expression elicits more potent
antitumor immune response than BMSCs solely expressing IL-18 or
IFN-β. Our results suggested that delivery of IL-18 and IFN-β by
BMSCs may be an excellent and promising approach for the
development of an effective treatment protocol for glioma
therapy.
Acknowledgments
The present study was supported by the Natural
Science Foundation of Shaanxi Province of China (no.
2014JM2-8179).
Abbreviations:
|
BMSCs
|
bone marrow-derived mesenchymal stem
cells
|
|
IL-18
|
interleukin-18
|
|
IFN-β
|
interferon-β
|
References
|
1
|
Mao XG, Zhang X and Zhen HN: Progress on
potential strategies to target brain tumor stem cells. Cell Mol
Neurobiol. 29:141–155. 2009. View Article : Google Scholar
|
|
2
|
Maher EA, Furnari FB, Bachoo RM, Rowitch
DH, Louis DN, Cavenee WK and DePinho RA: Malignant glioma: Genetics
and biology of a grave matter. Genes Dev. 15:1311–1333. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sanai N and Berger MS: Glioma extent of
resection and its impact on patient outcome. Neurosurgery.
62:753–764; discussion 264–266. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bexell D, Gunnarsson S, Tormin A, Darabi
A, Gisselsson D, Roybon L, Scheding S and Bengzon J: Bone marrow
multipotent mesenchymal stroma cells act as pericyte-like migratory
vehicles in experimental gliomas. Mol Ther. 17:183–190. 2009.
View Article : Google Scholar
|
|
5
|
Gunnarsson S, Bexell D, Svensson A, Siesjö
P, Darabi A and Bengzon J: Intratumoral IL-7 delivery by
mesenchymal stromal cells potentiates IFNgamma-transduced tumor
cell immuno-therapy of experimental glioma. J Neuroimmunol.
218:140–144. 2010. View Article : Google Scholar
|
|
6
|
Matuskova M, Hlubinova K, Pastorakova A,
Hunakova L, Altanerova V, Altaner C and Kucerova L: HSV-tk
expressing mesenchymal stem cells exert bystander effect on human
glio-blastoma cells. Cancer Lett. 290:58–67. 2010. View Article : Google Scholar
|
|
7
|
Suryadevara CM, Verla T, Sanchez-Perez L,
Reap EA, Choi BD, Fecci PE and Sampson JH: Immunotherapy for
malignant glioma. Surg Neurol Int. 6(Suppl 1): S68–S77. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okamura H, Tsutsi H, Komatsu T, Yutsudo M,
Hakura A, Tanimoto T, Torigoe K, Okura T, Nukada Y, Hattori K, et
al: Cloning of a new cytokine that induces IFN-gamma production by
T cells. Nature. 378:88–91. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hwang KS, Cho WK, Yoo J, Seong YR, Kim BK,
Kim S and Im DS: Adenovirus-mediated interleukin-18 mutant in vivo
gene transfer inhibits tumor growth through the induction of T cell
immunity and activation of natural killer cell cytotoxicity. Cancer
Gene Ther. 11:397–407. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ossendorp F, Mengedé E, Camps M, Filius R
and Melief CJ: Specific T helper cell requirement for optimal
induction of cytotoxic T lymphocytes against major
histocompatibility complex class II negative tumors. J Exp Med.
187:693–702. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kohyama M, Saijyo K, Hayasida M, Yasugi T,
Kurimoto M and Ohno T: Direct activation of human CD8+
cytotoxic T lymphocytes by interleukin-18. Jpn J Cancer Res.
89:1041–1046. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xia D, Li F and Xiang J: Engineered fusion
hybrid vaccine of IL-18 gene-modified tumor cells and dendritic
cells induces enhanced antitumor immunity. Cancer Biother
Radiopharm. 19:322–330. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Glick RP, Lichtor T, Mogharbel A, Taylor
CA and Cohen EP: Intracerebral versus subcutaneous immunization
with allogeneic fibroblasts genetically engineered to secrete
interleukin-2 in the treatment of central nervous system glioma and
melanoma. Neurosurgery. 41:898–906; discussion 906–907. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sobol RE, Fakhrai H, Shawler D, Gjerset R,
Dorigo O, Carson C, Khaleghi T, Koziol J, Shiftan TA and Royston I:
Interleukin-2 gene therapy in a patient with glioblastoma. Gene
Ther. 2:164–167. 1995.PubMed/NCBI
|
|
15
|
Hertzog PJ, Hwang SY and Kola I: Role of
interferons in the regulation of cell proliferation,
differentiation, and development. Mol Reprod Dev. 39:226–232. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nakahara N, Pollack IF, Storkus WJ,
Wakabayashi T, Yoshida J and Okada H: Effective induction of
antiglioma cytotoxic T cells by coadministration of interferon-beta
gene vector and dendritic cells. Cancer Gene Ther. 10:549–558.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Williams RF, Myers AL, Sims TL, Ng CY,
Nathwani AC and Davidoff AM: Targeting multiple angiogenic pathways
for the treatment of neuroblastoma. J Pediatr Surg. 45:1103–1109.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kageshita T, Mizuno M, Ono T, Matsumoto K,
Saida T and Yoshida J: Growth inhibition of human malignant
melanoma transfected with the human interferon-beta gene by means
of cationic liposomes. Melanoma Res. 11:337–342. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Natsume A, Mizuno M, Ryuke Y and Yoshida
J: Antitumor effect and cellular immunity activation by murine
interferon-beta gene transfer against intracerebral glioma in
mouse. Gene Ther. 6:1626–1633. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Natsume A, Tsujimura K, Mizuno M,
Takahashi T and Yoshida J: IFN-beta gene therapy induces systemic
antitumor immunity against malignant glioma. J Neurooncol.
47:117–124. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu G, Jiang XD, Xu Y, Zhang J, Huang FH,
Chen ZZ, Zhou DX, Shang JH, Zou YX and Cai YQ: Adenoviral-mediated
interleukin-18 expression in mesenchymal stem cells effectively
suppresses the growth of glioma in rats. Cell Biol Int. 33:466–474.
2009. View Article : Google Scholar
|
|
22
|
Olson JJ, Friedman R, Orr K, Delaney T and
Oldfield EH: Enhancement of the efficacy of x-irradiation by
pentobarbital in a rodent brain-tumor model. J Neurosurg.
72:745–748. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Farrell CL, Stewart PA and Del Maestro RF:
A new glioma model in rat: The C6 spheroid implantation technique
permeability and vascular characterization. J Neurooncol.
4:403–415. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen W, Wang J, Shao C, Liu S, Yu Y, Wang
Q and Cao X: Efficient induction of antitumor T cell immunity by
exosomes derived from heat-shocked lymphoma cells. Eur J Immunol.
36:1598–1607. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lohr J, Ratliff T, Huppertz A, Ge Y,
Dictus C, Ahmadi R, Grau S, Hiraoka N, Eckstein V, Ecker RC, et al:
Effector T-cell infiltration positively impacts survival of
glioblastoma patients and is impaired by tumor-derived TGF-β. Clin
Cancer Res. 17:4296–4308. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kraman M, Bambrough PJ, Arnold JN, Roberts
EW, Magiera L, Jones JO, Gopinathan A, Tuveson DA and Fearon DT:
Suppression of antitumor immunity by stromal cells expressing
fibroblast activation protein-alpha. Science. 330:827–830. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nakamizo A, Marini F, Amano T, Khan A,
Studeny M, Gumin J, Chen J, Hentschel S, Vecil G, Dembinski J, et
al: Human bone marrow-derived mesenchymal stem cells in the
treatment of gliomas. Cancer Res. 65:3307–3318. 2005.PubMed/NCBI
|
|
28
|
Uchibori R, Okada T, Ito T, Urabe M,
Mizukami H, Kume A and Ozawa K: Retroviral vector-producing
mesenchymal stem cells for targeted suicide cancer gene therapy. J
Gene Med. 11:373–381. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Menon LG, Kelly K, Yang HW, Kim SK, Black
PM and Carroll RS: Human bone marrow-derived mesenchymal stromal
cells expressing S-TRAIL as a cellular delivery vehicle for human
glioma therapy. Stem Cells. 27:2320–2330. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang XJ, Lu JT, Tu HJ, Huang KM, Fu R, Cao
G, Huang M, Cheng LH, Dai LJ and Zhang L: TRAIL-engineered bone
marrow-derived mesenchymal stem cells: TRAIL expression and
cytotoxic effects on C6 glioma cells. Anticancer Res. 34:729–734.
2014.PubMed/NCBI
|
|
31
|
Liu W, Han B, Sun B, Gao Y, Huang Y and Hu
M: Overexpression of interleukin-18 induces growth inhibition,
apoptosis and gene expression changes in a human tongue squamous
cell carcinoma cell line. J Int Med Res. 40:537–544. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu W, Hu M, Wang Y, Sun B, Guo Y, Xu Z,
Li J and Han B: Overexpression of interleukin-18 protein reduces
viability and induces apoptosis of tongue squamous cell carcinoma
cells by activation of glycogen synthase kinase-3β signaling. Oncol
Rep. 33:1049–1056. 2015.PubMed/NCBI
|
|
33
|
Zheng JN, Pei DS, Mao LJ, Liu XY, Sun FH,
Zhang BF, Liu YQ, Liu JJ, Li W and Han D: Oncolytic adenovirus
expressing interleukin-18 induces significant antitumor effects
against melanoma in mice through inhibition of angiogenesis. Cancer
Gene Ther. 17:28–36. 2010. View Article : Google Scholar
|
|
34
|
Takano S, Ishikawa E, Matsuda M, Yamamoto
T and Matsumura A: Interferon-β inhibits glioma angiogenesis
through downregulation of vascular endothelial growth factor and
upregulation of interferon inducible protein 10. Int J Oncol.
45:1837–1846. 2014.PubMed/NCBI
|
|
35
|
Yoshida J, Mizuno M and Yagi K:
Cytotoxicity of human beta-interferon produced in human glioma
cells transfected with its gene by means of liposomes. Biochem Int.
28:1055–1061. 1992.PubMed/NCBI
|
|
36
|
Guo Y, Wang G, Gao WW, Cheng SW, Wang R,
Ju SM, Cao HL and Tian HL: Induction of apoptosis in glioma cells
and upregulation of Fas expression using the human interferon-β
gene. Asian Pac J Cancer Prev. 13:2837–2840. 2012. View Article : Google Scholar
|
|
37
|
Zhang Y, Wang C, Zhang Y and Sun M: C6
glioma cells retrovi-rally engineered to express IL-18 and Fas
exert FasL-dependent cytotoxicity against glioma formation. Biochem
Biophys Res Commun. 325:1240–1245. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Marshall DJ, Rudnick KA, McCarthy SG,
Mateo LR, Harris MC, McCauley C and Snyder LA: Interleukin-18
enhances Th1 immunity and tumor protection of a DNA vaccine.
Vaccine. 24:244–253. 2006. View Article : Google Scholar
|
|
39
|
Robertson MJ, Mier JW, Logan T, Atkins M,
Koon H, Koch KM, Kathman S, Pandite LN, Oei C, Kirby LC, et al:
Clinical and biological effects of recombinant human interleukin-18
administered by intravenous infusion to patients with advanced
cancer. Clin Cancer Res. 12:4265–4273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kikuchi T, Akasaki Y, Joki T, Abe T,
Kurimoto M and Ohno T: Antitumor activity of interleukin-18 on
mouse glioma cells. J Immunother. 23:184–189. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yamanaka R and Xanthopoulos KG: Induction
of antigen-specific immune responses against malignant brain tumors
by intramuscular injection of sindbis DNA encoding gp100 and IL-18.
DNA Cell Biol. 24:317–324. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yamanaka R, Honma J, Tsuchiya N, Yajima N,
Kobayashi T and Tanaka R: Tumor lysate and IL-18 loaded dendritic
cells elicits Th1 response, tumor-specific CD8+
cytotoxic T cells in patients with malignant glioma. J Neurooncol.
72:107–113. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yamanaka R, Tsuchiya N, Yajima N, Honma J,
Hasegawa H, Tanaka R, Ramsey J, Blaese RM and Xanthopoulos KG:
Induction of an antitumor immunological response by an intratumoral
injection of dendritic cells pulsed with genetically engineered
Semliki Forest virus to produce interleukin-18 combined with the
systemic administration of interleukin-12. J Neurosurg. 99:746–753.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Meijer DH, Maguire CA, LeRoy SG and
Sena-Esteves M: Controlling brain tumor growth by intraventricular
administration of an AAV vector encoding IFN-beta. Cancer Gene
Ther. 16:664–671. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Streck CJ, Dickson PV, Ng CY, Zhou J, Hall
MM, Gray JT, Nathwani AC and Davidoff AM: Antitumor efficacy of
AAV-mediated systemic delivery of interferon-beta. Cancer Gene
Ther. 13:99–106. 2006. View Article : Google Scholar
|